

Early Identification of Plant Drought Stress Responses: Changes in Leaf Reflectance and Plant Growth Promoting Rhizobacteria Selection-The Case Study of Tomato Plants
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil Properties and Determination of Field Capacity
2.2. Tomato Plant Growth
2.3. Measurement of Plant Parameters
2.4. Isolation and Screening of Putative PGPR
2.5. M13 DNA Fingerprinting
2.6. Statistical Analysis
3. Results
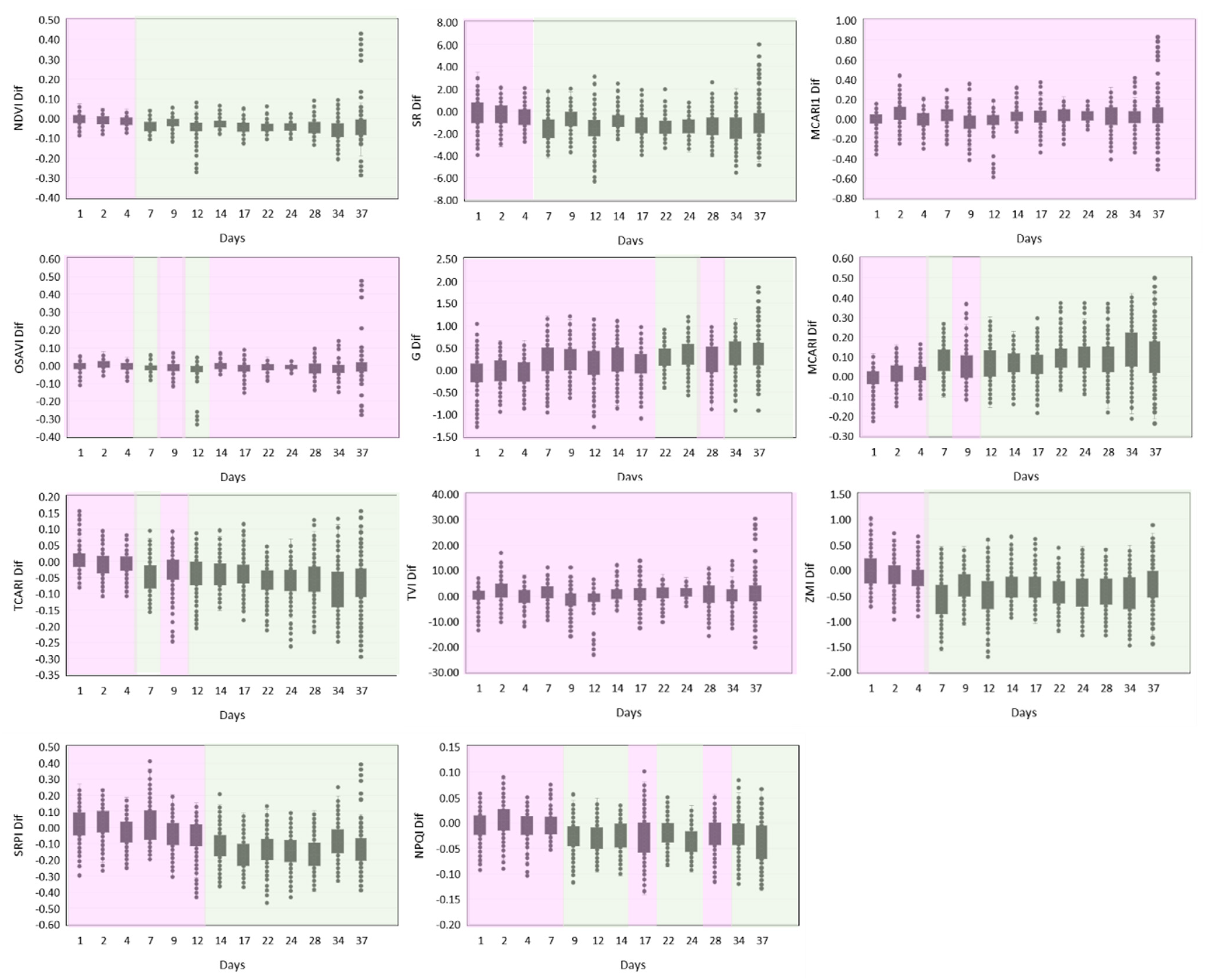
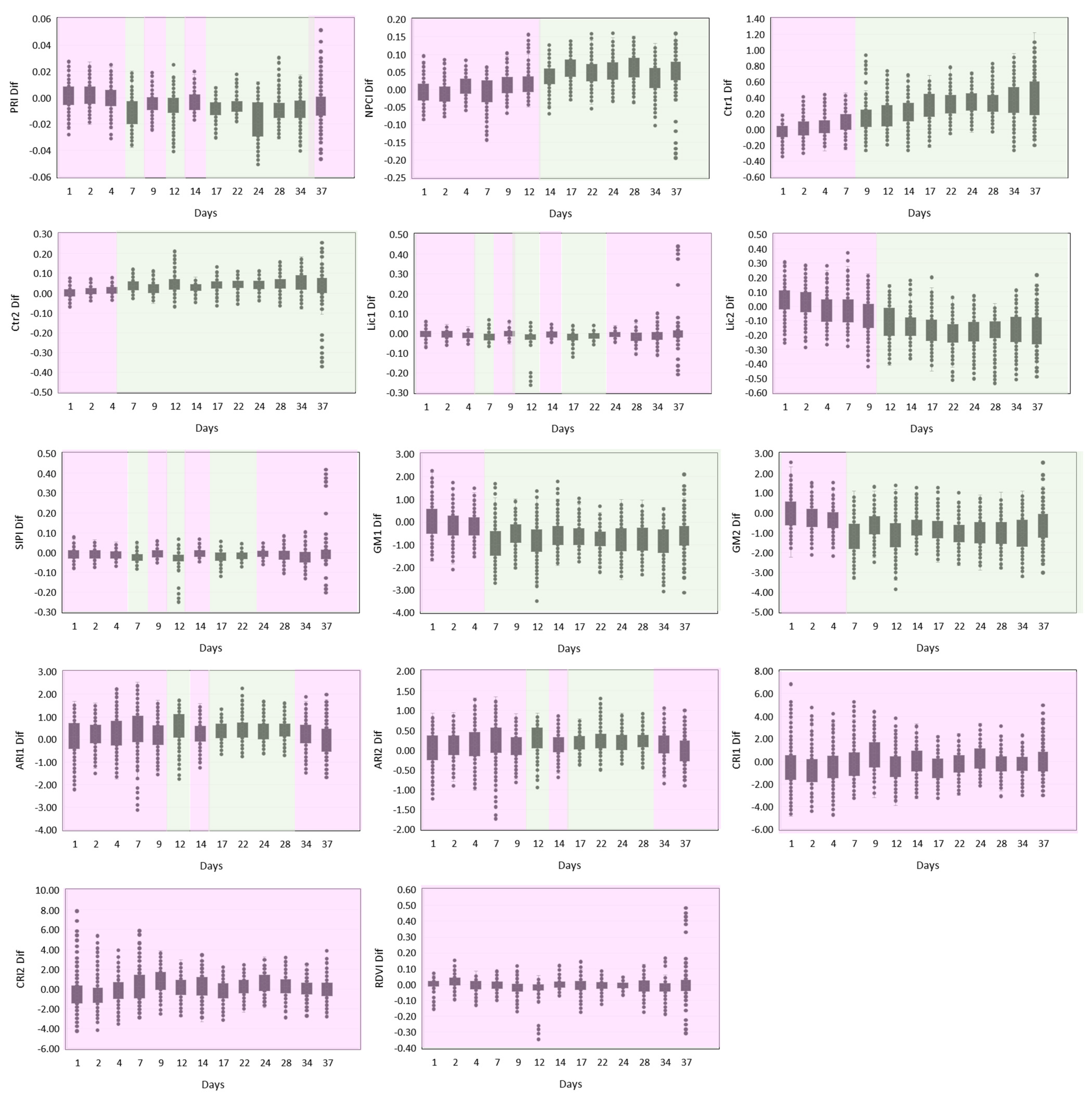
3.1. SRI Differences between the Watering Conditions
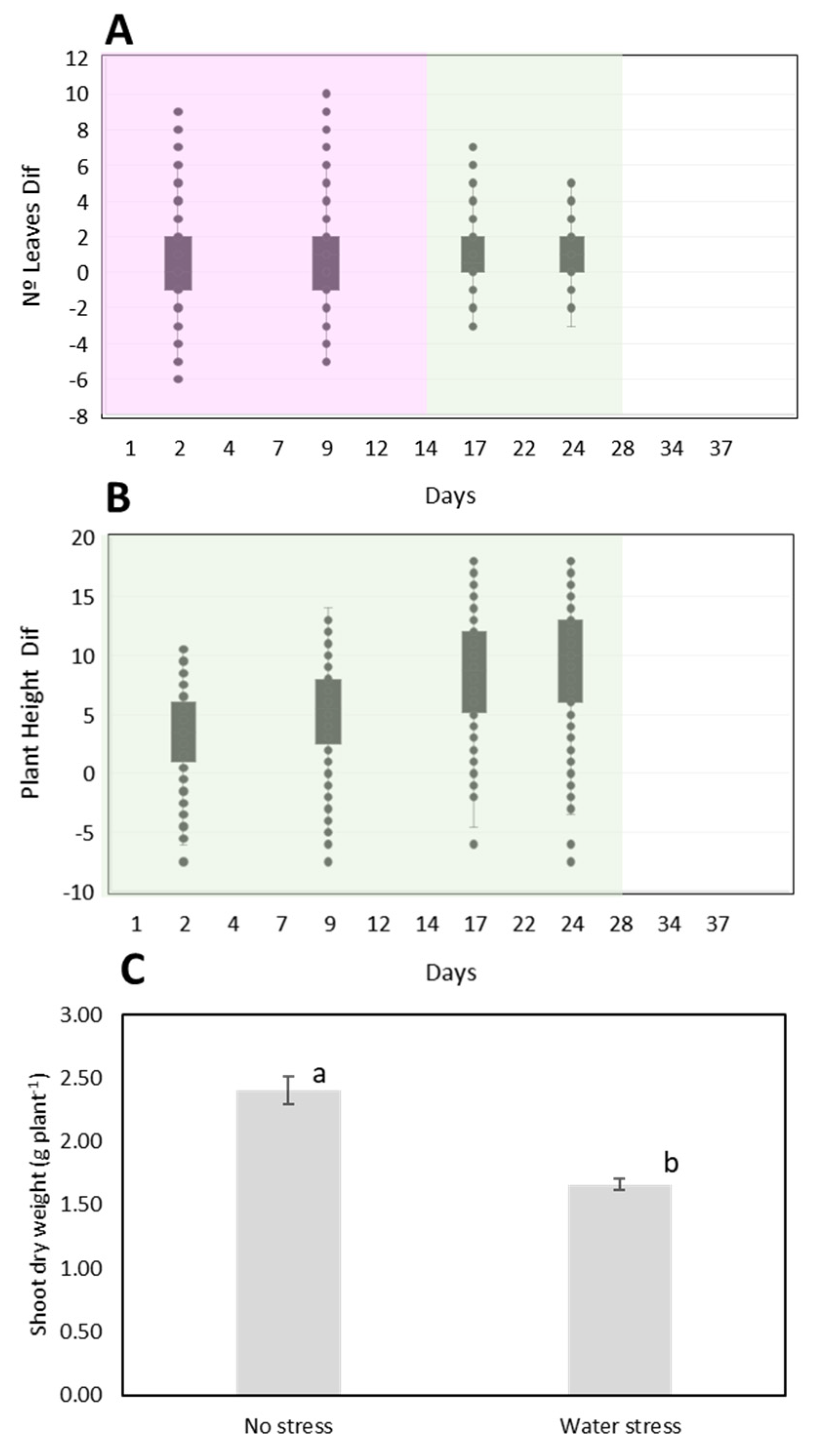
3.2. Plant Morphological Changes under Water Depletion
3.3. Link between SRIs and Plant Morphological Parameters
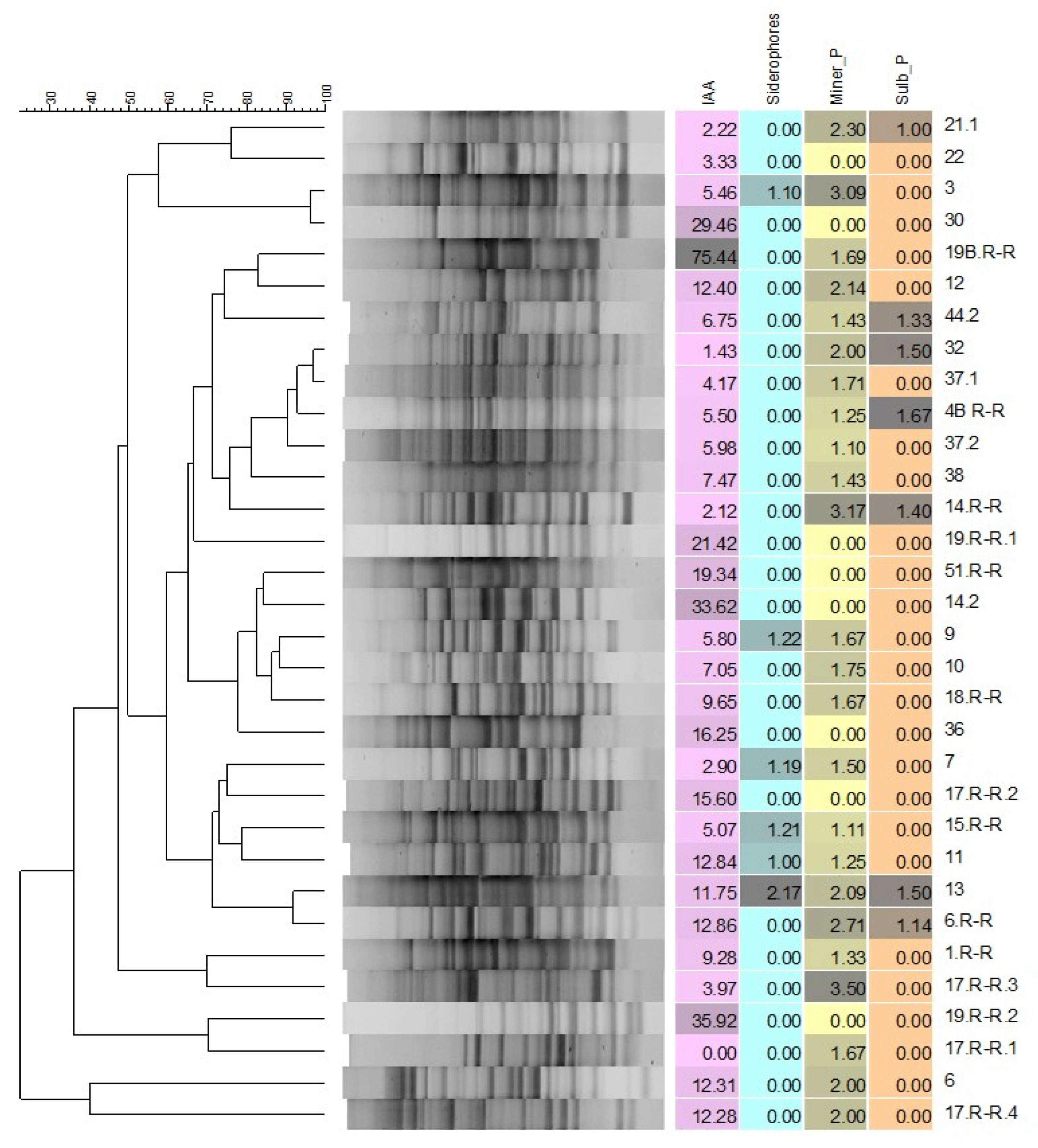
3.4. Characterization of Rhizobacterial Isolates
4. Discussion
4.1. Increased Chlorophyll Content of Tomato under Water Depletion
4.2. Physiological Adaptive Response Mechanisms under Water Depletion
4.3. The Link between SRIs and the Plant Morphology
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kasim, W.A.; Osman, M.E.; Omar, M.N.; Abd El-Daim, I.A.; Bejai, S.; Meijer, J. Control of Drought Stress in Wheat Using Plant Growth Promoting Bacteria. J. Plant Growth Regul. 2013, 32, 122–130. [Google Scholar] [CrossRef]
- Meena, K.K.; Sorty, A.M.; Bitla, U.M.; Choudhary, K.; Gupta, P.; Pareek, A.; Singh, D.P.; Prabha, R.; Sahu, P.K.; Gupta, V.K.; et al. Abiotic Stress Responses and Microbe-Mediated Mitigation in Plants: The Omics Strategies. Front. Plant Sci. 2017, 9, 172. [Google Scholar] [CrossRef] [Green Version]
- Vurukonda, S.S.; Vardharajula, S.; Shrivastava, M.; SkZ, A. Enhancement of Drought Stress Tolerance in Crops by Plant Growth Promoting Rhizobacteria. Microbiol. Res. 2016, 184, 13–24. [Google Scholar] [CrossRef]
- Shrivastava, P.; Kumar, R. Soil Salinity: A Serious Environmental Issue and Plant Growth Promoting Bacteria as One of the Tools for its Alleviation. Saudi J. Biol. Sci. 2015, 22, 123–131. [Google Scholar] [CrossRef] [Green Version]
- Jaleel, C.A.; Manivannan, P.; Wahid, A.; Farooq, M.; Al-Juburi, H.J.; Somasundaram, R.; Panneerselvam, R. Drought Stress in Plants: A Review on Morphological Characteristics and Pigments Composition. Int. J. Agric. Biol. 2009, 11, 100–105. [Google Scholar]
- Khan, N.A.; Khan, M.I.R.; Ferrante, A.; Poor, P. Editorial: Ethylene: A Key Regulatory Molecule in Plants. Front. Plant Sci. 2017, 8, 1782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under Drought and Salt Stress: Regulation Mechanisms from Whole Plant to Cell. Ann. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruz de Carvalho, M.H. Drought Stress and Reactive Oxygen Species: Production, Scavenging and Signaling. Plant Signal. Behav. 2008, 3, 156–165. [Google Scholar] [CrossRef] [Green Version]
- Alharby, H.F.; Al-Zahrani, H.S.; Alzahrani, Y.M.; Alsamadany, H.; Hakeem, K.R.; Rady, M.M. Maize Grain Extract Enriched with Polyamines Alleviates Drought Stress in Triticum aestivum through Up-Regulation of the Ascorbate–Glutathione Cycle, Glyoxalase System, and Polyamine Gene Expression. Agronomy 2021, 11, 949. [Google Scholar] [CrossRef]
- Rady, M.M.; Boriek, S.H.K.; Abd El-Mageed, T.A.; Seif El-Yazal, M.A.; Ali, E.F.; Hassan, F.A.S.; Abdelkhalik, A. Exogenous Gibberellic Acid or Dilute Bee Honey Boosts Drought Stress Tolerance in Vicia faba by Rebalancing Osmoprotectants, Antioxidants, Nutrients, and Phytohormones. Plants 2021, 10, 748. [Google Scholar] [CrossRef]
- Tiwari, S.; Lata, C.; Chauhan, P.S.; Nautiyal, C.S. Pseudomonas putida Attunes Morphophysiological, Biochemical and Molecular Responses in Cicer arietinum L. During Drought Stress and Recovery. Plant Physiol. Biochem. 2016, 99, 108–117. [Google Scholar] [CrossRef]
- Abedini, D.; Jaupitre, S.; Bouwmeester, H.; Dong, L. Metabolic Interactions in Beneficial Microbe Recruitment by Plants. Curr. Opin. Biotechnol. 2021, 70, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Schalk, I.J.; Hannauer, M.; Braud, A. New Roles for Bacterial Siderophores in Metal Transport and Tolerance. Environ. Microbiol. 2011, 13, 2844–2854. [Google Scholar] [CrossRef]
- Backer, R.; Rokem, J.S.; Ilangumaran, G.; Lamont, J.; Praslickova, D.; Ricci, E.; Subramanian, S.; Smith, D.L. Plant Growth-Promoting Rhizobacteria: Context, Mechanisms of Action, and Roadmap to Commercialization of Biostimulants for Sustainable Agriculture. Front. Plant Sci. 2018, 9, 1473. [Google Scholar] [CrossRef] [Green Version]
- Adesemoye, A.O.; Torbert, H.A.; Kloepper, J.W. Plant Growth-Promoting Rhizobacteria Allow Reduced Application Rates of Chemical Fertilizers. Microb. Ecol. 2009, 58, 921–929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le, T.A.; Pék, Z.; Takács, S.; Neményi, A.; Daood, H.G.; Helyes, L. The Effect of Plant Growth Promoting Rhizobacteria on the Water-Yield Relationship and Carotenoid Production of Processing Tomatoes. HortScience 2018, 53, 816–822. [Google Scholar] [CrossRef] [Green Version]
- Akhtar, S.S.; Amby, D.B.; Hegelund, J.N.; Fimognari, L.; Großkinsky, D.K.; Westergaard, J.C.; Müller, R.; Moelbak, L.; Liu, F.; Roitsch, T. Bacillus licheniformis FMCH001 Increases Water Use Efficiency Via Growth Stimulation in Both Normal and Drought Conditions. Front. Plant Sci. 2020, 11, 297. [Google Scholar] [CrossRef]
- Xun, W.; Shao, J.; Shen, Q.; Zhang, R. Rhizosphere Microbiome: Functional Compensatory Assembly for Plant Fitness. Comput. Struct. Biotechnol. J. 2021, 19, 5487–5493. [Google Scholar] [CrossRef]
- Naylor, D.; Coleman-Derr, D. Drought Stress and Root-Associated Bacterial Communities. Front. Plant Sci. 2018, 8, 2223. [Google Scholar] [CrossRef] [PubMed]
- Timmusk, S.; Abd El-Daim, I.A.; Copolovici, L.; Tanilas, T.; Kännaste, A.; Behers, L.; Nevo, E.; Seisenbaeva, G.; Stemström, E.; Niinemets, Ü. Drought-Tolerance of Wheat Improved by Rhizosphere Bacteria from Harsh Environments: Enhanced Biomass Production and Reduced Emissions of Stress Volatiles. PLoS ONE 2014, 9, e96086. [Google Scholar] [CrossRef] [Green Version]
- Jochum, M.D.; McWilliams, K.L.; Borrego, E.J.; Kolomiets, M.V.; Niu, G.; Pierson, E.A.; Jo, Y.-K. Bioprospecting Plant Growth-Promoting Rhizobacteria that Mitigate Drought Stress in Grasses. Front. Microbiol. 2019, 10, 2106. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Gao, X.; Shen, Z.; Zhu, C.; Jiao, Z.; Li, R.; Shen, Q. Pre-colonization of PGPR Triggers Rhizosphere Microbiota Succession Associated with Crop Yield Enhancement. Plant Soil 2019, 439, 553–567. [Google Scholar] [CrossRef]
- Berg, G.; Kusstatscher, P.; Abdelfattah, A.; Cernava, T.; Smalla, K. Microbiome Modulation-Toward a Better Understanding of Plant Microbiome Response to Microbial Inoculants. Front. Microbiol. 2021, 12, 650610. [Google Scholar] [CrossRef]
- Erdle, K.; Mistele, B.; Schmidhalter, U. Spectral High-Throughput Assessments of Phenotypic Differences in Biomass and Nitrogen Partitioning During Grain Filling of Wheat Under High Yielding Western European Conditions. Field Crops Res. 2013, 141, 16–26. [Google Scholar] [CrossRef]
- Becker, E.; Schmidhalter, U. Evaluation of Yield and Drought Using Active and Passive Spectral Sensing Systems at the Reproductive Stage in Wheat. Front. Plant Sci. 2017, 8, 379. [Google Scholar] [CrossRef] [Green Version]
- El-Hendawy, S.E.; Alotaibi, M.; Al-Suhaibani, N.; Al-Gaadi, K.; Hassan, W.; Dewir, Y.H.; Emam, M.A.E.-G.; Elsayed, S.; Schmidhalter, U. Comparative Performance of Spectral Reflectance Indices and Multivariate Modeling for Assessing Agronomic Parameters in Advanced Spring Wheat Lines Under Two Contrasting Irrigation Regimes. Front. Plant Sci. 2019, 10, 1537. [Google Scholar] [CrossRef]
- El-Hendawy, S.; Al-Suhaibani, N.; Elsayed, S.; Hassan, W.M.; Dewir, Y.H.; Refay, Y.; Abdella, K.A. Potential of the Existing and Novel Spectral Reflectance Indices for Estimating the Leaf Water Status and Grain Yield of Spring Wheat Exposed to Different Irrigation Rates. Agric. Water Manag. 2019, 217, 356–373. [Google Scholar] [CrossRef]
- Wahb-Allah, M.A.; Alsadon, A.A.; Ibrahim, A.A. Drought Tolerance of Several Tomato Genotypes under Greenhouse Conditions. World Appl. Sci. J. 2011, 15, 933–940. [Google Scholar]
- Santana, M.M.; Carvalho, L.; Melo, J.; Araújo, M.E.; Cruz, C. Unveiling the Hidden Interaction between Thermophiles and Plant Crops: Wheat and Soil Thermophilic Bacteria. J. Plant Interact. 2020, 15, 127–138. [Google Scholar] [CrossRef]
- Zhou, R.; Yu, X.; Ottosen, C.O.; Rosenqvist, E.; Zhao, L.; Wang, Y.; Yu, W.; Zhao, T.; Wu, Z. Drought Stress had a Predominant Effect over Heat Stress on Three Tomato Cultivars Subjected to Combined Stress. BMC Plant Biol. 2017, 17, 24. [Google Scholar] [CrossRef] [Green Version]
- Bakken, L.R. Separation and Purification of Bacteria from Soil. Appl. Environ. Microbiol. 1985, 49, 1482–1487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yim, W.J.; Poonguzhali, S.; Madhaiyan, M.; Palaniappan, P.; Siddikee, M.A.; Sa, T. Characterization of Plant-Growth Promoting Diazotrophic Bacteria Isolated from Field Grown Chinese Cabbage under Different Fertilization Conditions. J. Microbiol. 2009, 47, 147–155. [Google Scholar] [CrossRef]
- Kelel, M.; Abera, G.; Yisma, A.; Molla, B.; Gebre, N.; Adugna, T.; Gary, M.; Wessel, G.M. Isolation of Phosphate Solubilizing Bacteria from Acacia Tree Rhizophere Soil. J. Microbiol. Biotechnol. Res. 2017, 4, 9–13. [Google Scholar]
- Alexander, D.B.; Zuberer, D.A. Use of Chrome Azurol S Reagents to Evaluate Siderophore Production by Rhizosphere Bacteria. Biol. Fertil. Soils 1991, 12, 39–45. [Google Scholar] [CrossRef]
- Ryskov, A.P.; Jincharadze, A.G.; Prosnyak, M.I.; Ivanov, P.L.; Limborska, S.A. M13 Phage DNA as a Universal Marker for DNA Fingerprinting of Animals, Plants and Microorganisms. FEBS Lett. 1988, 233, 388–392. [Google Scholar] [CrossRef] [Green Version]
- Huey, B.; Hall, J. Hypervariable DNA Fingerprinting in Escherichia coli: Minisatellite Probe from Bacteriophage M13. J. Bacteriol. 1989, 171, 2528–2532. [Google Scholar] [CrossRef] [Green Version]
- Pitcher, D.G.; Saunders, N.A.; Owen, R.J. Rapid Extraction of Bacterial Genomic DNA with Guanidium Thiocyanate. Lett. Appl. Microbiol. 1989, 8, 151–156. [Google Scholar] [CrossRef]
- Rouse, J.W.; Haas, R.H.; Schell, J.A.; Deering, D.W. Monitoring Vegetation Systems in the Great Plains with ERTS. In Proceedings of the 3rd ERTS Symposium, NASA SP-351; Scientific and Technical Information Office, National Aeronautics and Space Administration: Washington, DC, USA, 1973; Volume 1, pp. 309–317. [Google Scholar]
- Rouse, J.W.; Haas, R.H.; Deering, D.W.; Schell, J.A.; Harlan, J.C. Monitoring the Vernal Advancement and Retrogradation (Greenwave Effect) of Natural Vegetation, Type III Final Report; NASA Goddard Space Flight Center: Greenbelt, MD, USA, 1974; p. 371. [Google Scholar]
- Sellers, P.J. Canopy Reflectance, Photosynthesis, and Transpiration. Int. J. Remote Sens. 1985, 6, 1335–1372. [Google Scholar] [CrossRef] [Green Version]
- Birth, G.S.; McVey, G. Measuring the Colour of Growing Turf with a Reflectance Spectrophotometer. Agron. J. 1968, 60, 640–643. [Google Scholar] [CrossRef]
- Jordan, C.F. Derivation of Leaf Area Index from Quality of Light on the Forest Floor. Ecology 1969, 50, 663–666. [Google Scholar] [CrossRef]
- Haboudane, D.; Miller, J.R.; Pattey, E.; Zarco-Tejada, P.J.; Strachan, I.B. Hyperspectral Vegetation Indices and Novel Algorithms for Predicting Green LAI of Crop Canopies: Modeling and Validation in the Context of Precision Agriculture. Remote Sens. Environ. 2004, 90, 337–352. [Google Scholar] [CrossRef]
- Rondeaux, G.; Steven, M.; Barret, F. Optimization of Soil Adjusted Vegetation Indices. Remote Sens. Environ. 1996, 55, 95–107. [Google Scholar] [CrossRef]
- Zarco-Tejada, P.J.; Berjón, A.; López-Lozano, R.; Miller, J.R.; Martín, P.; Cachorro, V.; González, M.R.; de Frutos, A. Assessing Vineyard Condition with Hyperspectral Indices: Leaf and Canopy Reflectance Simulation in a Row-Structured Discontinuous Canopy. Remote Sens. Environ. 2005, 99, 271–287. [Google Scholar] [CrossRef]
- Daughtry, C.S.T.; Walthall, C.L.; Kim, M.S.; De Colstoun, E.B.; McMurtrey, J.E., III. Estimating Corn Leaf Chlorophyll Concentration from Leaf and Canopy Reflectance. Remote Sens. Environ. 2000, 74, 229–239. [Google Scholar] [CrossRef]
- Haboudane, D.; Miller, J.R.; Trembley, N.; Zarco-Tejada, P.J.; Dextraze, L. Integrated Narrow Band Vegetation Indices for Prediction of Crop Chlorophyll Content for Application to Precision Agriculture. Remote Sens. Environ. 2002, 81, 416–426. [Google Scholar] [CrossRef]
- Broge, N.H.; Leblanc, E. Comparing Predictive Power and Stability of Broad-Band and Hyperspectral Vegetation Indices for Estimation of Green Leaf Area Index and Canopy Chlorophyll Density. Remote Sens. Environ. 2000, 76, 156–172. [Google Scholar] [CrossRef]
- Zarco-Tejada, P.J.; Miller, J.R.; Noland, T.L.; Mohammed, G.H.; Sampson, P.H. Scaling-Up and Model Inversion Methods with Narrow-Band Optical Indices for Chlorophyll Content Estimation in Closed Forest Canopies with Hyperspectral Data. IEEE Trans. Geosci. Remote Sens. 2001, 39, 1491–1507. [Google Scholar] [CrossRef] [Green Version]
- Peñuelas, J.; Baret, F.; Filella, I. Semi-Empirical Indices to Assess Carotenoids/Chlorophyll a Ratio from Leaf Spectral Reflectance. Photosynthetica 1995, 31, 221–230. [Google Scholar]
- Barnes, J.D.; Balaguer, L.; Manrique, E.; Elvira, S.; Davison, A.W. A Reappraisal of the Use of DMSO for the Extraction and Determination of Chlorophylls a and b in Lichens and Higher Plants. Environ. Exp. Bot. 1992, 32, 85–100. [Google Scholar] [CrossRef]
- Peñuelas, J.; Filella, I.; Gamon, J.A. Assessment of Photosynthetic Radiation-Use Efficiency with Spectral Reflectance. New Phytol. 1995, 131, 291–296. [Google Scholar] [CrossRef]
- Gamon, J.A.; Serrano, L.; Surfus, J.S. The Photochemical Reflectance Index: An Optical Indicator of Photosynthetic Radiation Use Efficiency across Species, Functional Types, and Nutrient Levels. Oecologia 1997, 112, 492–501. [Google Scholar] [CrossRef] [PubMed]
- Peñuelas, J.; Gamon, J.A.; Fredeen, A.L.; Merino, J.; Field, C.B. Reflectance Indices Associated with Physiological Changes in Nitrogen- and Water-Limited Sunflower Leaves. Remote Sens. Environ. 1994, 48, 135–146. [Google Scholar] [CrossRef]
- Carter, G.A. Ratios of Leaf Reflectance in Narrow Wavebands as Indicators of Plant Stress. Int. J. Remote Sens. 1994, 15, 697–703. [Google Scholar] [CrossRef]
- Carter, G.A.; Cibula, W.G.; Miller, R.L. Narrow-Band Reflectance Imagery Compared with Thermal Imagery for Early Detection of Plant Stress. J. Plant Physiol. 1996, 148, 515–522. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Lang, M.; Sowinska, M.; Heisel, F.; Miehé, J.A. Detection of Vegetation Stress Via a New High Resolution Fluorescence Imaging System. J. Plant Physiol. 1996, 148, 599–612. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Merzlyak, M.N. Remote Estimation of Chlorophyll Content in Higher Plant Leaves. Int. J. Remote Sens. 1997, 18, 2691–2697. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Merzlyak, M.N. Remote Sensing of Chlorophyll Concentration in Higher Plant Leaves. Adv. Space Res. 1998, 22, 689–692. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Merzlyak, M.N.; Chivkunova, O.B. Optical Properties and Nondestructive Estimation of Anthocyanin Content in Plant Leaves. Photochem. Photobiol. 2001, 74, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Gitelson, A.A.; Zur, Y.; Chivkunova, O.B.; Merzlyak, M.N. Assessing Carotenoid Content in Plant Leaves with Reflectance Spectroscopy. Photochem. Photobiol. 2002, 75, 272–281. [Google Scholar] [CrossRef]
- Roujean, J.-L.; Breon, F.-M. Estimating PAR Absorbed by Vegetation from Bidirectional Reflectance Measurements. Remote Sens. Environ. 1995, 51, 375–384. [Google Scholar] [CrossRef]
- Estill, K.; Delaney, R.H.; Smith, W.K.; Ditterline, R.L. Water Relations and Productivity of Alfalfa Leaf Chlorophyll Variants. Crop Sci. 1991, 31, 1229–1233. [Google Scholar] [CrossRef]
- Ashraf, M.; Karim, F. Screening of Some Cultivars/Lines of Black Gram (Vigna mungo L. Hepper) for Resistance to Water Stress. Trop. Agric. 1991, 68, 57–62. [Google Scholar]
- Gubiš, J.; Vaňková, R.; Červená, V.; Dragúňová, M.; Hudcovicová, M.; Lichtnerová, H.; Dokupil, T.; Jureková, Z. Transformed Tobacco Plants with Increased Tolerance to Drought. S. Afr. J. Bot. 2007, 73, 505–511. [Google Scholar] [CrossRef] [Green Version]
- Makela, P.; Karkkainen, J.; Somersalo, S. Effect of Glycinebetaine on Chloroplast Ultrastructure, Chlorophyll and Protein Content, and RuBPCO activities in Tomato Grown Under Drought or Salinity. Biol. Plant. 2000, 43, 471–475. [Google Scholar] [CrossRef]
- Yudina, L.; Sukhova, E.; Mudrilov, M.; Nerush, V.; Pecherina, A.; Smirnov, A.A.; Dorokhov, A.S.; Chilingaryan, N.O.; Vodeneev, V.; Sukhov, V. Ratio of Intensities of Blue and Red Light at Cultivation Influences Photosynthetic Light Reactions, Respiration, Growth, and Reflectance Indices in Lettuce. Biology 2022, 11, 60. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Miehé, J.A. Fluorescence Imaging as a Diagnostic Tool for Plant Stress. Trends Plant Sci. 1997, 2, 316–320. [Google Scholar] [CrossRef]
- Chen, X.L.; Li, Y.L.; Wang, L.C.; Guo, W.Z. Red and Blue Wavelengths Affect the Morphology, Energy Use Efficiency and Nutritional Content of Lettuce (Lactuca sativa L.). Sci. Rep. 2021, 11, 8374. [Google Scholar] [CrossRef]
- Izzo, L.G.; Mickens, M.A.; Aronne, G.; Gómez, C. Spectral Effects of Blue and Red Light on Growth, Anatomy, and Physiology of Lettuce. Physiol. Plant. 2021, 172, 2191–2202. [Google Scholar] [CrossRef]
- Joliot, P.; Joliot, A. Cyclic Electron Flow in C3 Plants. Biochim. Biophys. Acta 2006, 1757, 362–368. [Google Scholar] [CrossRef] [Green Version]
- Sukhov, V.; Sukhova, E.; Vodeneev, V. Long-Distance Electrical Signals as a Link between the Local Action of Stressors and the Systemic Physiological Responses in Higher Plants. Prog. Biophys. Mol. Biol. 2019, 146, 63–84. [Google Scholar] [CrossRef]
- Sukhov, V. Electrical Signals as Mechanism of Photosynthesis Regulation in Plants. Photosynth. Res. 2016, 130, 373–387. [Google Scholar] [CrossRef]
- Murata, N.; Nishiyama, Y. ATP is a Driving Force in the Repair of Photosystem II during Photoinhibition. Plant Cell Environ. 2018, 41, 285–299. [Google Scholar] [CrossRef]
- Huang, W.; Yang, S.J.; Zhang, S.B.; Zhang, J.L.; Cao, K.F. Cyclic Electron Flow Plays an Important Role in Photoprotection for the Resurrection Plant Paraboea rufescens Under Drought Stress. Planta 2012, 235, 819–828. [Google Scholar] [CrossRef] [PubMed]
- Lehtimäki, N.; Lintala, M.; Allahverdiyeva, Y.; Aro, E.M.; Mulo, P. Drought Stress-Induced Upregulation of Components Involved in Ferredoxin-Dependent Cyclic Electron Transfer. J. Plant Physiol. 2010, 167, 1018–1022. [Google Scholar] [CrossRef]
- Suorsa, M. Cyclic Electron Flow Provides Acclimatory Plasticity for the Photosynthetic Machinery Under Various Environmental Conditions and Developmental Stages. Front. Plant Sci. 2015, 6, 800. [Google Scholar] [CrossRef] [PubMed]
- Barton, C.V.M.; North, P.R.J. Remote Sensing of Canopy Light Use Efficiency Using the Photochemical Reflectance Index: Model and Sensitivity Analysis. Remote Sens. Environ. 2001, 78, 264–273. [Google Scholar] [CrossRef]
- Thenot, F.; Méthy, M.; Winkel, T. The Photochemical Reflectance Index (PRI) as a Water-Stress Index. Int. J. Remote Sens. 2002, 23, 5135–5139. [Google Scholar] [CrossRef]
- Wieloch, T.; Augusti, A.; Schleucher, J. Anaplerotic Flux into the Calvin-Benson Cycle: Hydrogen Isotope Evidence for in vivo Occurrence in C3 Metabolism. New Phytol. 2022, 234, 405–411. [Google Scholar] [CrossRef]
- Sunil, B.; Saini, D.; Bapatla, R.B.; Aswani, V.; Raghavendra, A.S. Photorespiration is Complemented by Cyclic Electron Flow and the Alternative Oxidase Pathway to Optimize Photosynthesis and Protect against Abiotic Stress. Photosynth. Res. 2019, 139, 67–79. [Google Scholar] [CrossRef]
- Ahmad, H.M.; Fiaz, S.; Hafeez, S.; Zahra, S.; Shah, A.N.; Gul, B.; Aziz, O.; Mahmood-Ur-Rahman; Fakhar, A.; Rafique, M.; et al. Plant Growth-Promoting Rhizobacteria Eliminate the Effect of Drought Stress in Plants: A Review. Front. Plant Sci. 2022, 13, 875774. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pH (H2O) (20 °C) | 7.4 |
| σ (mS cm−1) | 0.583 |
| Texture | Clay |
| % Organic matter | 4.13 |
| % N | 0.18 |
| Phosphorus (P2O5) mg kg−1 | 46.9 |
| Potassium (K2O) mg kg−1 | 220 |
| Sulfur (S) mg kg−1 | 15.5 |
| Sodium (Na) mg kg−1 | 15.8 |
| Zinc (Zn) mg kg−1 | <2.20 (LQ) 1 |
| Molybdenum (Mo) mg kg−1 | <0.400 (LQ) 1 |
| Acronym | Name | Description | Formula | Reference |
|---|---|---|---|---|
| NDVI | Normalized Difference Vegetation Index | Quantifies green vegetation by calculating the normalized difference between near-infrared (scattered by green leaves) and red light (absorbed by vegetation); directly related to the photosynthetic capacity of the plant. | NDVI = (RNIR − RRED)/(RNIR + RRED) | [38,39,40] |
| SR | Simple Ratio Index | Quantifies green vegetation by calculating the ratio between the reflectance in near-infrared and red bands, taking advantage of the increased red-light absorption by chlorophyll, but increased reflectance of near-infrared energy for healthy plants. | SR = RNIR/RRED | [39,41,42] |
| MCARI1 | Modified Chlorophyll Absorption in Reflectance Index 1 | Less sensitive than MCARI (see below) to the effect of leaf chlorophyll content on the prediction of green leaf area index (LAI). | MCARI1 = 1.2 × [2.5 × (R790 − R670) − 1.3 × (R790 − R550)] | [43] |
| OSAVI | Optimized Soil-Adjusted Vegetation Index | The index is optimized for agricultural monitoring, being a good estimator of green vegetation in homogeneous canopies such as those from agricultural crops, especially at mid-latitudes. | OSAVI = (1 + 0.16) × (R790 − R670)/(R790 − R670 + 0.16) | [44] |
| G | Greenness Index | A chlorophyll index for chlorophyll content estimation from leaf optical properties, i.e., from leaf reflectance in the visible and the red edge spectral region. | G = R554/R677 | [45] |
| MCARI | Modified Chlorophyll Absorption in Reflectance Index | The index responds to leaf chlorophyll concentrations, but shows high sensitivity to variations in LAI, being difficult to interpret for low LAI values. It minimizes the confounding background reflectance of soil and non-photosynthetic elements. | MCARI = [(R700 − R670) − 0.2 × (R700 − R550)] × (R700/R670) | [46] |
| TCARI | Transformed Chlorophyll Absorption Reflectance Index | As MCARI, this index indicates the relative abundance of chlorophyll, but exhibits a better sensitivity at low chlorophyll concentrations. Although relatively less responsive than MCARI to LAI variations, it is still sensitive to the underlying soil reflectance properties, particularly for low LAIs. | TCARI = 3 × [(R700- R670) − 0.2 × (R700-R550) × (R700/R670)] | [47] |
| TVI | Triangular Vegetation Index | Calculated as the area of a hypothetical triangle in spectral space that links green peak reflectance, the chlorophyll absorption minimum, and the NIR shoulder. Both chlorophyll absorption causing a decrease in red reflectance, and leaf tissue abundance causing an increase in NIR reflectance, will increase the triangle area. This index is a good estimator of green LAI. | TVI = 0.5 × [120 × (R750 − R550) − 200 × (R670 − R550)] | [48] |
| ZMI | Zarco-Tejada and Miller Index | The index is used to estimate chlorophyll a+b contents at canopy level, minimizing the effect of LAI variation. | ZMI = R750/R710 | [49] |
| SRPI | Simple Ratio Pigment Index | The index is used to assess the ratio between carotenoids and chlorophyll a concentrations. | SRPI = R430/R680 | [50] |
| NPQI | Normalized Phaeophytinization Index | The index estimates relative contents of chlorophyll a and phaeophytin, the primary electron acceptor in PSII. | NPQI = (R415 − R435)/(R415 + R435) | [49,51] |
| PRI | Photochemical Reflectance Index | The index is sensitive to changes in carotenoid pigments (particularly xanthophylls) in live foliage; xanthophyll cycle pigments are closely linked to PSII light use efficiency and PRI is an optical indicator of photosynthetic radiation use efficiency. | PRI = (R531 − R570)/(R531 + R570) | [52,53] |
| NPCI | Normalized Pigment Chlorophyll Index | Defined to evaluate carotenoids/chlorophyll ratio, and particularly applicable to N limitation, when plants develop greater concentration of carotenoids relative to chlorophyll, this index estimates the proportion of total photosynthetic pigments to chlorophyll. | NPCI = (R680 − R430)/(R680 + R430) | [54] |
| Ctr1 | Carter Index 1 | As a result of decreased absorption by pigments, visible reflectance increases in stressed leaves and is most sensitive to stress in the 535–640 nm and 685–700 nm range; ratios of leaf reflectances Ctr1 and Ctr 2 were those that most strongly indicated plant stress caused by several agents among several species. | Ctr1 = R695/R420 | [55,56] |
| Ctr2 | Carter Index 2 | Ctr2 = R695/R760 | ||
| Lic1 | Lichtenthaler Index 1 | The ratios, namely, blue (440 nm)/red (690 nm) reflectances, are related to chlorophyll fluorescence emission; F440/F690 is very sensitive to environmental changes, permitting an early stress detection in plants’ photosynthetic apparatus. | Lic1 = (R790 − R680)/(R790 + R680) | [45,57] |
| Lic2 | Lichtenthaler Index 2 | Lic2 = R440/R690 | ||
| SIPI | Structure Intensive Pigment Index | The index is used to assess the ratio between carotenoids and chlorophyll a concentrations. | SIPI = (R790 − R450)/(R790 − R650) | [50] |
| GM1 | Gitelson and Merzlyak Index 1 | GM ratios were developed as indices with maximum sensitivity to chlorophyll as they use reflectances corresponding to wavelengths with high sensitivity (550 nm and 700 nm), and insensitivity (750 nm) to variations in chlorophyll content; directly proportional to the leaves’ chlorophyll concentration in several plant species and within a large range of its variation. | GM1 = R750/R550 | [58,59] |
| GM2 | Gitelson and Merzlyak Index 2 | GM2 = R750/R700 | ||
| ARI1 | Anthocyanin Reflectance Index 1 | ARI1 index was developed for the estimation of the accumulation of anthocyanins, which are stress-related pigments. ARI2 corrects for leaf density and thickness, as the near-infrared spectral band (760–800 nm), related to leaf scattering, is added to ARI1. | ARI1 = 1/R550 − 1/R700 | [60] |
| ARI2 | Anthocyanin Reflectance Index 2 | ARI2 = R800 × (1/R550 − 1/R700) | ||
| CRI1 | Carotenoid Reflectance Index 1 | CRI indices estimate the total carotenoid content in plant leaves; the sensitivity of reciprocal reflectance to carotenoid content was maximal around 510 nm, but since chlorophylls affect reflectance in this spectral range, a reciprocal reflectance at either 550 or 700 nm, linearly proportional to the chlorophyll content, was used to remove their effect. | CRI1 = 1/R510 − 1/R550 | [61] |
| CRI2 | Carotenoid Reflectance Index 2 | CRI2 = 1/R510 − 1/R700 | ||
| RDVI | Renormalized Difference Vegetation Index | The index uses the difference between near-infrared and red wavelengths, along with NDVI, to spot healthy vegetation, but it is insensitive to the effects of soil reflectance and sun-viewing geometry. | RDVI = (R780 − R670)/((R780 + R670)^0.5) | [62] |
| Index | No. Leaves | Plant Height |
|---|---|---|
| NDVI | 0.124 | −0.721 ** |
| SR | 0.124 | −0.721 ** |
| MCARI1 | 0.151 | 0.047 |
| OSAVI | 0.194 * | −0.474 ** |
| G | −0.195 * | 0.388 ** |
| MCARI | −0.097 | 0.512 ** |
| TCARI | 0.115 | −0.504 ** |
| TVI | 0.214 ** | −0.011 |
| ZMI | 0.157 * | −0.592 ** |
| SRPI | 0.195 * | −0.575 ** |
| NPQI | 0.015 | −0.489 ** |
| PRI | 0.000 | −0.525 ** |
| NPCI | −0.195 * | 0.575 ** |
| CTr1 | −0.196 * | 0.657 ** |
| CTr2 | −0.179 * | 0.708 ** |
| Lic1 | 0.068 | −0.508 ** |
| Lic2 | 0.213 ** | −0.572 ** |
| SIPI | 0.014 | −0.432 ** |
| GM1 | 0.231 ** | −0.647 ** |
| GM2 | 0.174 * | −0.658 ** |
| ARI1 | 0.107 | 0.254 ** |
| ARI2 | 0.118 | 0.252 ** |
| CRI1 | −0.200 * | −0.063 |
| CRI2 | −0.178 * | 0.057 |
| RDVI | 0.237 ** | −0.306 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosa, A.P.; Barão, L.; Chambel, L.; Cruz, C.; Santana, M.M. Early Identification of Plant Drought Stress Responses: Changes in Leaf Reflectance and Plant Growth Promoting Rhizobacteria Selection-The Case Study of Tomato Plants. Agronomy 2023, 13, 183. https://doi.org/10.3390/agronomy13010183
Rosa AP, Barão L, Chambel L, Cruz C, Santana MM. Early Identification of Plant Drought Stress Responses: Changes in Leaf Reflectance and Plant Growth Promoting Rhizobacteria Selection-The Case Study of Tomato Plants. Agronomy. 2023; 13(1):183. https://doi.org/10.3390/agronomy13010183
Chicago/Turabian StyleRosa, Ana Paula, Lúcia Barão, Lélia Chambel, Cristina Cruz, and Margarida Maria Santana. 2023. "Early Identification of Plant Drought Stress Responses: Changes in Leaf Reflectance and Plant Growth Promoting Rhizobacteria Selection-The Case Study of Tomato Plants" Agronomy 13, no. 1: 183. https://doi.org/10.3390/agronomy13010183