
Effect of Application of Biostimulants on the Biomass, Nitrate, Pigments, and Antioxidants Content in Radish and Turnip Microgreens


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Conditions
2.2. Biostimulants Application
2.3. Seedling Measurement
2.4. Chlorophyll and Carotenoid Pigments Measurement
2.5. Total Sugars Measurement
2.6. Nitrate Concentrations Measurement
2.7. Ascorbic Acid Measurement
2.8. Total Phenolics Measurement
2.9. Radical Scavenging Activity Measurement
2.10. Statistical Analysis
3. Results
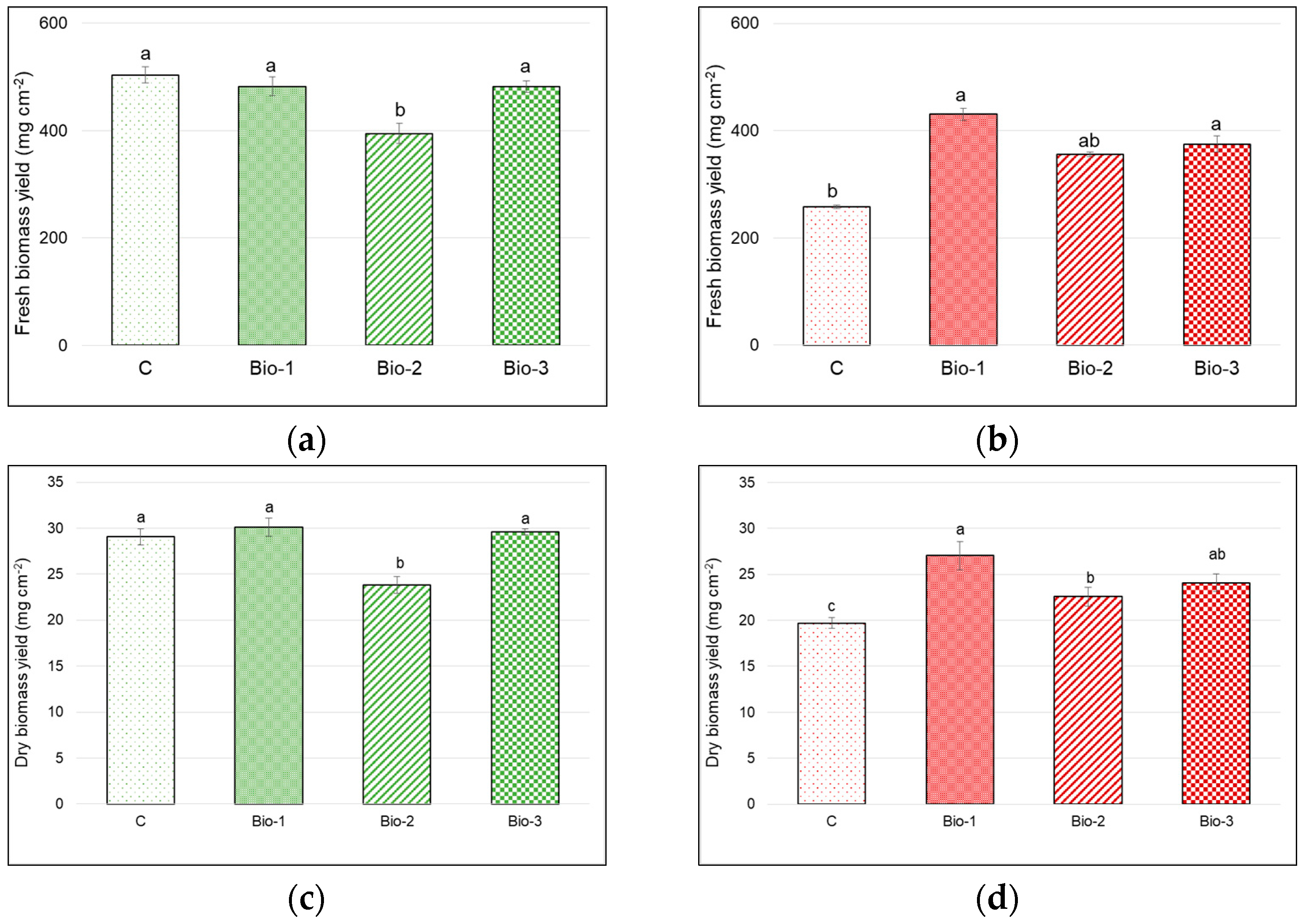
3.1. Fresh and Dry Biomass Yield
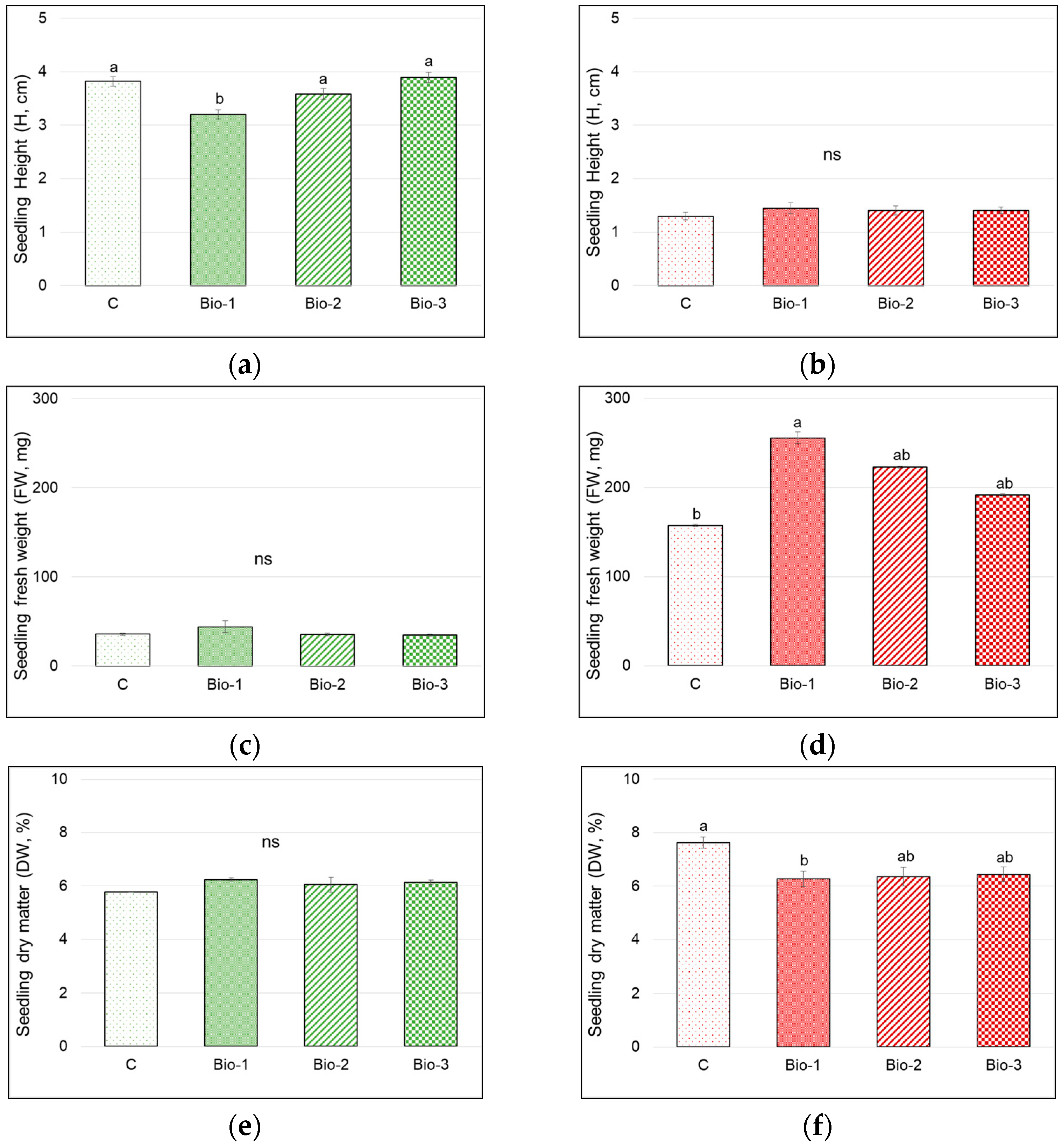
3.2. Seedling Height and Weight
3.3. Chlorophyll (a, b, Total) and Carotenoids
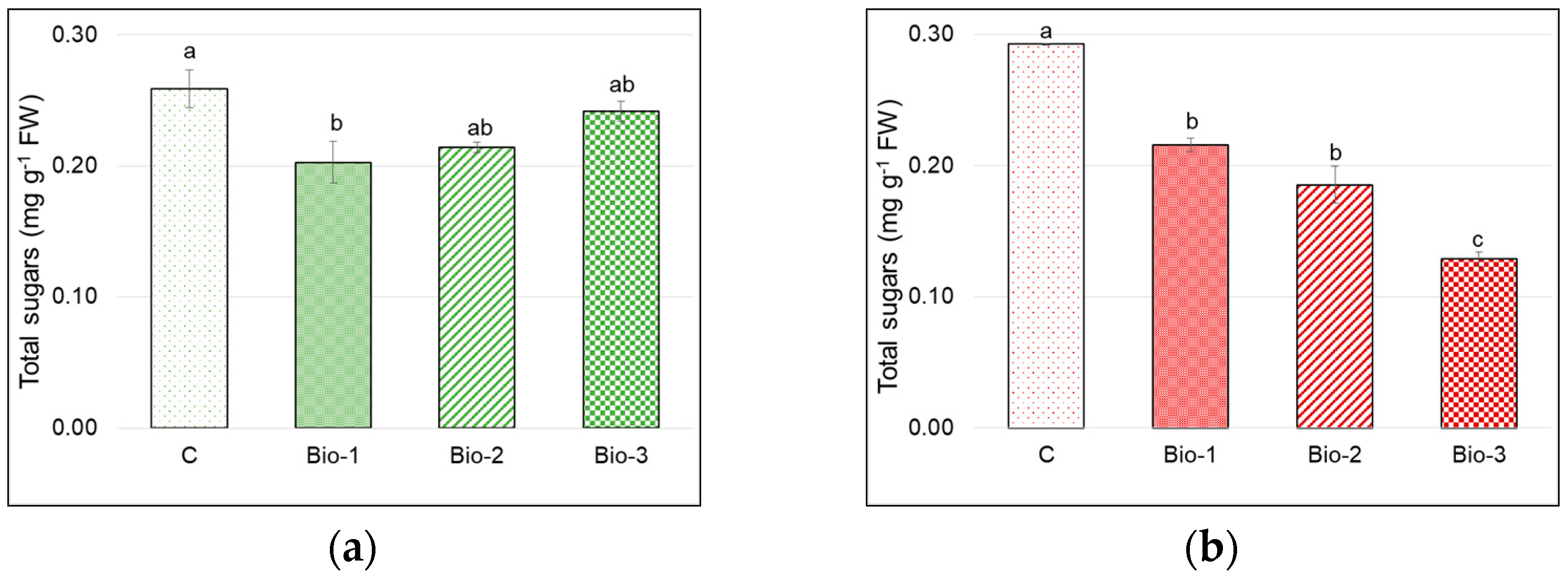
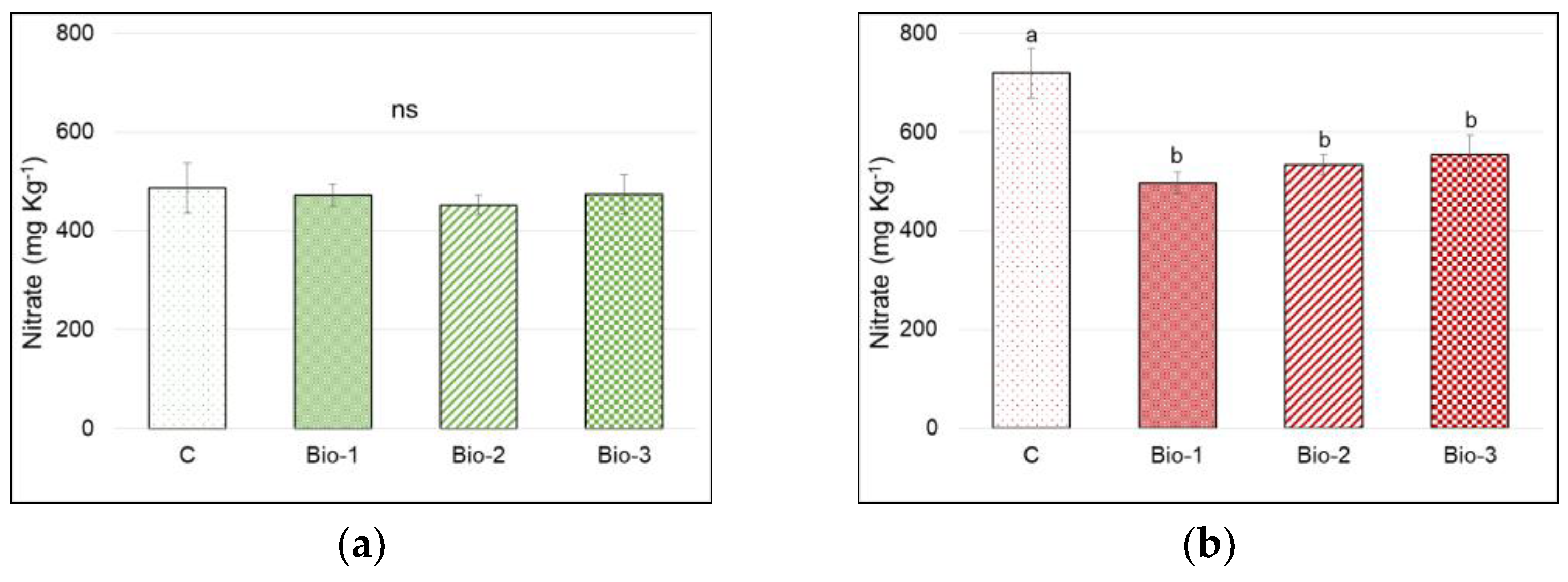
3.4. Total Sugars and Nitrates Content
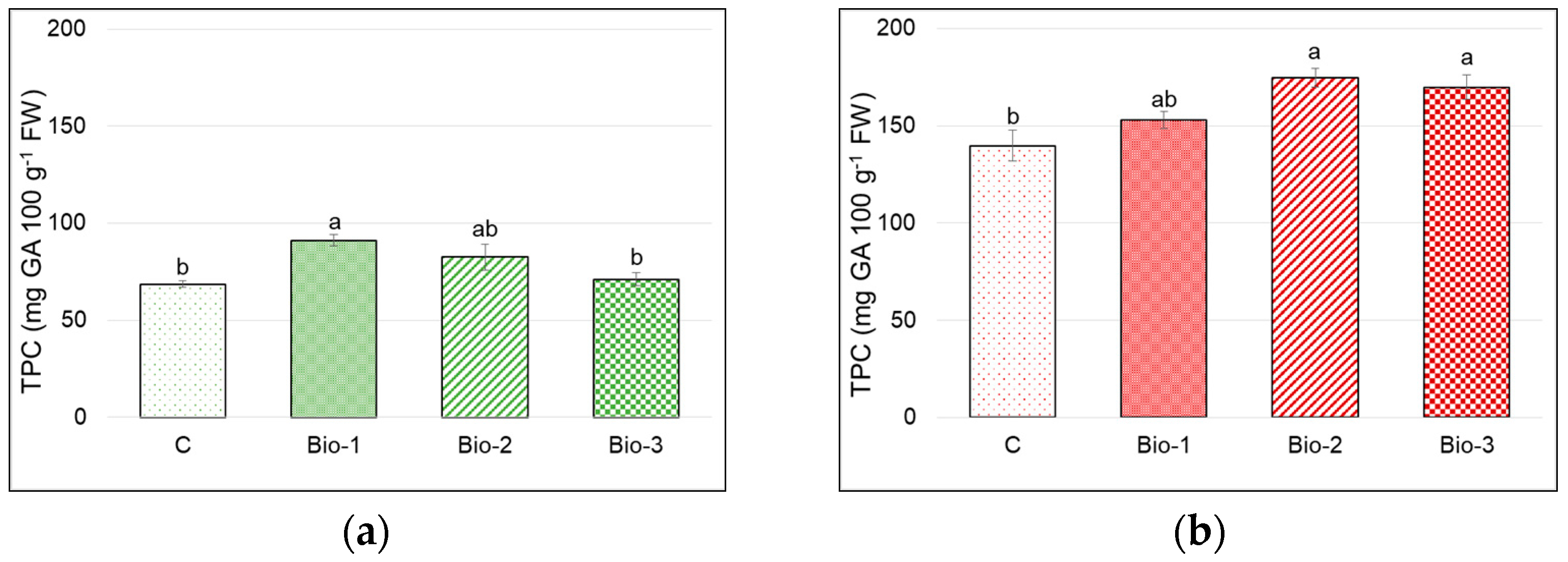
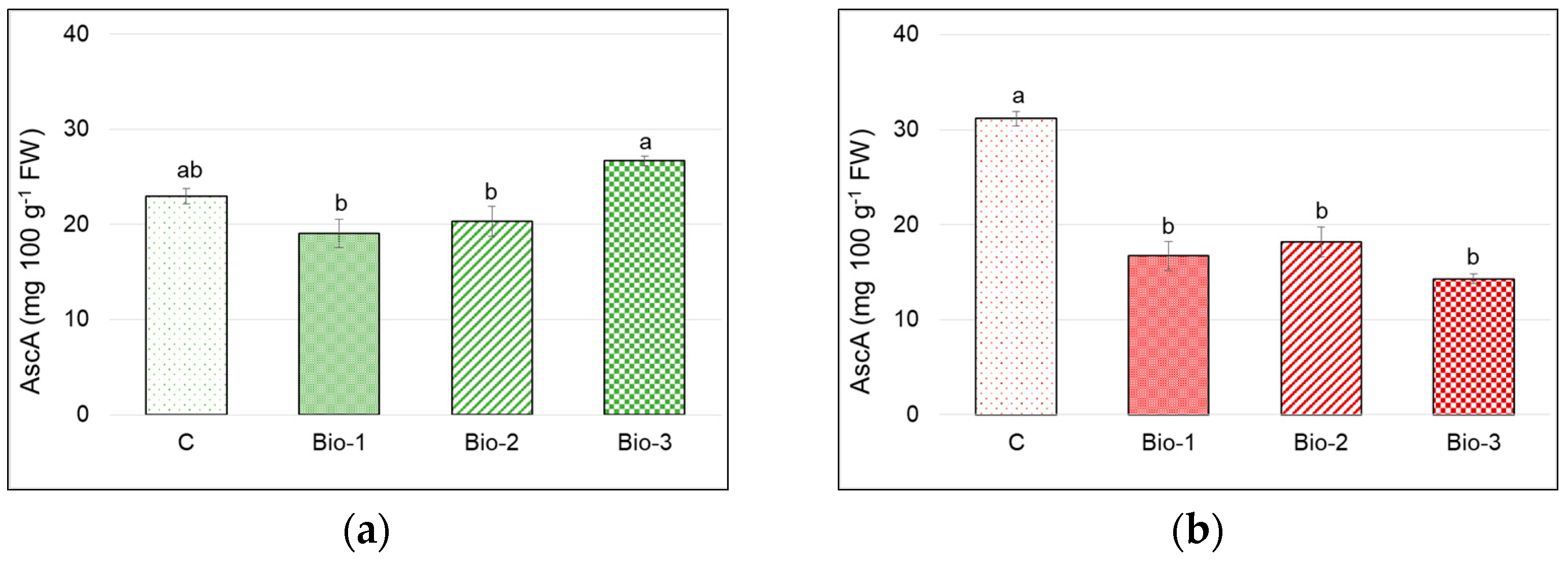
3.5. Antioxidants and Antioxidant Activity
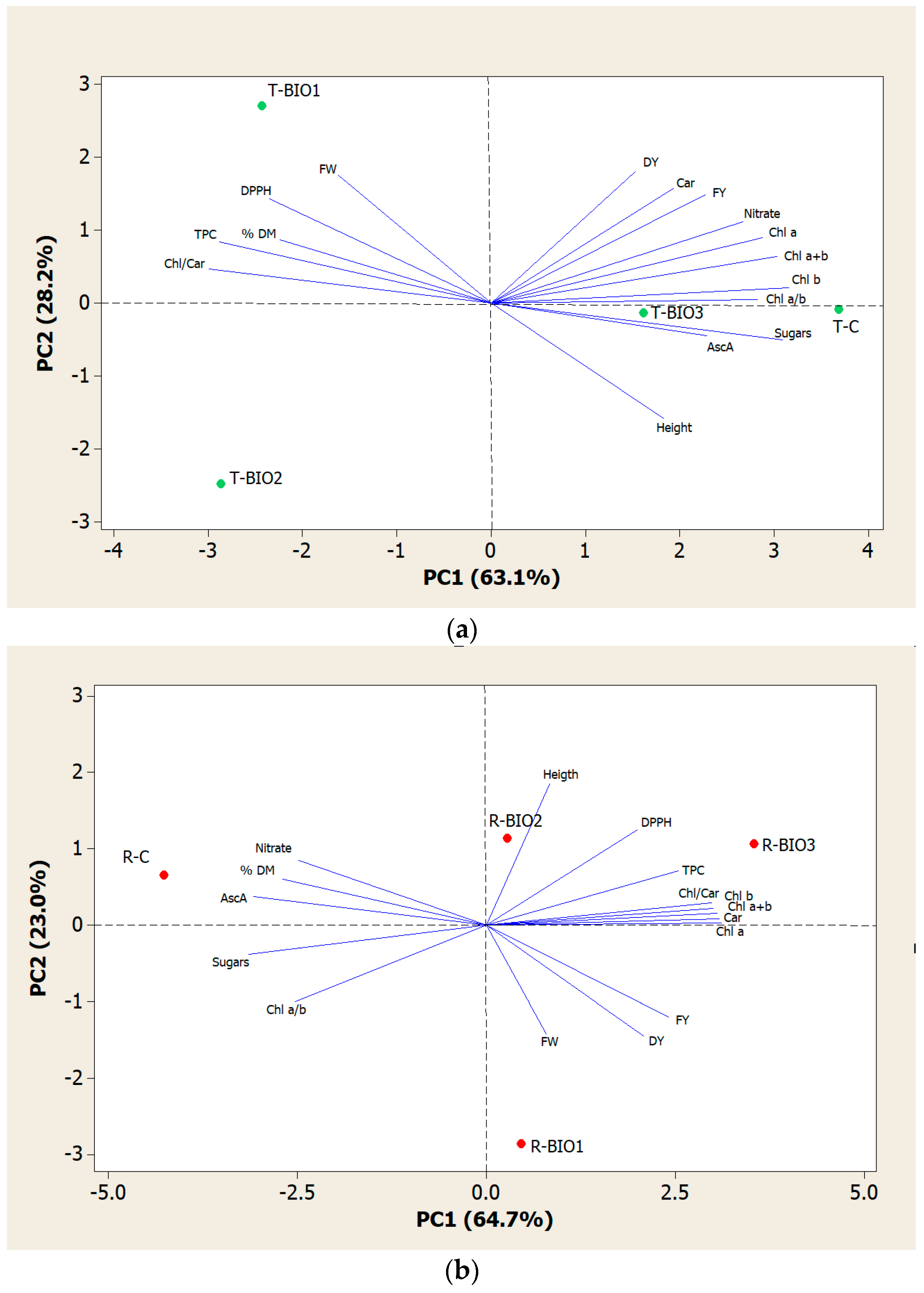
3.6. Analysis of Principal Component
3.7. Pearson’s Correlation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Di Bella, M.C.; Niklas, A.; Toscano, S.; Picchi, V.; Romano, D.; Lo Scalzo, R.; Branca, F. Morphometric characteristics, polyphenols and ascorbic acid variation in Brassica oleracea L. novel foods: Sprouts, microgreens and baby leaves. Agronomy 2020, 10, 782. [Google Scholar] [CrossRef]
- Kyriacou, M.C.; El-Nakhel, C.; Graziani, G.; Pannico, A.; Soteriou, G.A.; Giordano, M. Functional quality in novel food sources: Genotypic variation in the nutritive and phytochemical composition of thirteen microgreens species. Food Chem. 2019, 277, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Yadav, L.; Koley, T.K.; Tripathi, A.; Singh, S.P. Antioxidant potentiality and mineral content of summer season leafy greens: Comparison at mature and microgreen stages using chemometric. Agric. Res. 2019, 8, 165–175. [Google Scholar] [CrossRef]
- Pannico, A.; Kyriacou, M.C.; El-Nakhel, C.; Graziani, G.; Carillo, P.; Corrado, G.; Ritieni, A.; Rouphael, Y.; De Pascale, S. Hemp microgreens as an innovative functional food: Variation in the organic acids, amino acids, polyphenols, and cannabinoids composition of six hemp cultivars. Int. Food Res. J. 2022, 161, 111863. [Google Scholar] [CrossRef]
- Kyriacou, M.C.; El-Nakhel, C.; Pannico, A.; Graziani, G.; Zarrelli, A.; Soteriou, G.A.; Kyratzis, A.; Antoniou, C.; Pizzolongo, F.; Romano, R. Ontogenetic variation in the mineral, phytochemical and yield attributes of Brassicaceous microgreens. Foods 2021, 10, 1032. [Google Scholar] [CrossRef]
- Di Gioia, F.; De Bellis, P.; Mininni, C.; Santamaria, P.; Serio, F. Physicochemical, agronomical and microbiological evaluation of alternative growing media for the production of rapini (Brassica rapa L.) microgreens. J. Sci. Food Agric. 2017, 97, 1212–1219. [Google Scholar] [CrossRef]
- Ertani, A.; Schiavon, M.; Altissimo, A.; Franceschi, C.; Nardi, S. Phenol-containing organic substances stimulate phenylpropanoid metabolism in Zea mays. J. Plant Nutr. Soil. Sci. 2011, 174, 496–503. [Google Scholar] [CrossRef]
- Ertani, A.; Schiavon, M.; Muscolo, A.; Nardi, S. Alfalfa plant-derived biostimulant stimulate short-term growth of salt stressed Zea mays L. plants. Plant Soil 2013, 364, 145–158. [Google Scholar] [CrossRef]
- Toscano, S.; Romano, D.; Massa, D.; Bulgari, R.; Franzoni, G.; Ferrante, A. Biostimulant applications in low input horticultural cultivation systems. Italus Hortus 2019, 25, 27–36. [Google Scholar] [CrossRef]
- EC. REGULATION (EU) 2019/1009 of the European Parliament and of the Council of 5 June 2019 Laying Down Rules on the Making Available on the Market of EU Fertilising Products and Amending Regulations (EC) No 1069/2009 and (EC) No 1107/2009 and Repealing Regulation (EC) No 2003/2003. 2019. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=celex%3A32019R1009 (accessed on 27 October 2022).
- Pannacci, E.; Baratta, S.; Falcinelli, B.; Farneselli, M.; Tei, F. Mugwort (Artemisia vulgaris L.) Aqueous extract: Hormesis and biostimulant activity for seed germination and seedling growth in vegetable crops. Agriculture 2022, 12, 1329. [Google Scholar] [CrossRef]
- Colla, G.; Rouphael, Y.; Canaguier, R.; Svecova, E.; Cardarelli, M. Biostimulant action of a plant-derived protein hydrolysate produced through enzymatic hydrolysis. Front. Plant Sci. 2014, 5, 448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaafsma, G. Safety of protein hydrolysates, fractions thereof and bioactive peptides in human nutrition. Eur. J. Clin. Nutr. 2009, 63, 1161–1168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colla, G.; Nardi, S.; Cardarelli, M.; Ertani, A.; Lucini, L.; Canaguier, R.; Rouphael, Y. Protein hydrolysates as biostimulants in horticulture. Sci. Hortic. 2015, 196, 28–38. [Google Scholar] [CrossRef]
- Carillo, P.; Colla, G.; Fusco, G.M.; Dell’Aversana, E.; El-Nakhel, C.; Giordano, M.; Pannico, A.; Cozzolino, E.; Mori, M.; Reynaud, H.; et al. Morphological and physiological responses induced by protein hydrolysate-based biostimulant and nitrogen rates in greenhouse spinach. Agronomy 2019, 9, 450. [Google Scholar] [CrossRef] [Green Version]
- Sestili, F.; Rouphael, Y.; Cardarelli, M.; Pucci, A.; Bonini, P.; Canaguier, R.; Colla, G. Protein hydrolysate stimulates growth in tomato coupled with N-dependent gene expression involved in N assimilation. Front. Plant Sci. 2018, 9, 1233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bulgari, R.; Cocetta, G.; Trivellini, A.; Vernieri, P.; Ferrante, A. Biostimulants and crop responses: A review. Biol. Agric. Hortic. 2015, 31, 1–17. [Google Scholar] [CrossRef]
- Howladar, S.M. A novel Moringa oleifera leaf extract can mitigate the stress effects of salinity and cadmium in bean (Phaseolus vulgaris L.) plants. Ecotoxicol. Environ. Saf. 2014, 100, 69–75. [Google Scholar] [CrossRef]
- Rady, M.M.; Mohamed, G.F. Modulation of salt stress effects on the growth, physio-chemical attributes and yields of Phaseolus vulgaris L. plants by the combined application of salicylic acid and Moringa oleifera leaf extract. Sci. Hortic. 2015, 193, 105–113. [Google Scholar] [CrossRef]
- Arif, Y.; Bajguz, A.; Hayat, S. Moringa oleifera extract as a natural plant biostimulant. J. Plant Growth Regul. 2022, 1–16. [Google Scholar] [CrossRef]
- Phiri, C. Influence of Moringa oleifera leaf extracts on germination and early seedling development of major cereals. Am. J. Agric. Biol. Sci. 2010, 1, 774–777. [Google Scholar] [CrossRef]
- Mashamaite, C.V.; Ngcobo, B.L.; Manyevere, A.; Bertling, I.; Fawole, O.A. Assessing the usefulness of Moringa oleifera leaf extract as a biostimulant to supplement synthetic fertilizers: A Review. Plants 2022, 11, 2214. [Google Scholar] [CrossRef] [PubMed]
- Hala, H.; El-Noor, A.; Ewais, N.A. Effect of Moringa oleifera leaf extract (MLE) on pepper seed germination, seedlings improvement, growth, fruit yield and its quality. Middle East J. Agric. Res. 2017, 6, 448–463. [Google Scholar]
- Colla, G.; Hoagland, L.; Ruzzi, M.; Cardarelli, M.; Bonini, P.; Canaguier, R.; Rouphael, Y. Biostimulant action of protein hydrolysates: Unraveling their effects on plant physiology and microbiome. Front. Plant Sci. 2017, 8, 2202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, A.; Verma, J. Does plant-microbe interaction confer stress tolerance in plants: A review? Microbiol. Res. 2018, 207, 41–52. [Google Scholar] [CrossRef]
- Valencia, E.; Gross, N.; Quero, J.L.; Carmona, C.P.; Ochoa, V.; Gozalo, B.; Delgado-Baquerizo, M.; Dumack, K.; Hamonts, K.; Singh, B.K.; et al. Cascading effects from plants to soil microorganisms explain how plant species richness and simulated climate change affect soil multifunctionality. Glob. Chang. Biol. 2018, 24, 5642–5654. [Google Scholar] [CrossRef]
- Paradìković, N.; Teklić, T.; Zeljković, S.; Lisjak, M.; Špoljarević, M. Biostimulants research in some horticultural plant species—A review. Food Energy Secur. 2019, 8, e00162. [Google Scholar] [CrossRef]
- Toscano, S.; Ferrante, A.; Branca, F.; Romano, D. Enhancing the quality of two species of baby leaves sprayed with Moringa leaf extract as biostimulant. Agronomy 2021, 11, 1399. [Google Scholar] [CrossRef]
- Zulfiqar, F.; Casadesús, A.; Brockman, H.; Munné-Bosch, S. An overview of plant-based natural biostimulants for sustainable horticulture with a particular focus on moringa leaf extracts. Plant Sci. 2020, 295, 110194. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Method Enzymol. 1987, 148, 350–382. [Google Scholar]
- Cocetta, G.; Rossoni, M.; Gardana, C.; Mignani, I.; Ferrante, A.; Spinardi, A. Methyl jasmonate affects phenolic metabolism and gene expression in blueberry (Vaccinium corymbosum). Physiol. Plant 2015, 153, 269–283. [Google Scholar] [CrossRef]
- Cataldo, D.A.; Haroon, M.; Sehrader, L.E.; Youngs, V.L. Rapid colorimetric determination of nitrate in plant tissue by titration of salicylic acid. Commun. Soil Sci. Plant Anal. 1975, 6, 71–80. [Google Scholar] [CrossRef]
- Janghel, E.K.; Gupta, V.K.; Rai, M.K.; Rai, J.K. Micro determination of ascorbic acid using methyl viologen. Talanta 2007, 72, 1013–1016. [Google Scholar] [CrossRef] [PubMed]
- Toscano, S.; Ferrante, A.; Leonardi, C.; Romano, D. PAL activities in asparagus spears during storage after ammonium sulfate treatments. Postharvest Biol. Technol. 2018, 140, 34–41. [Google Scholar] [CrossRef]
- Kyriacou, M.C.; El-Nakhel, C.; Pannico, A.; Graziani, G.; Soteriou, G.A.; Giordano, M.; Zarrelli, A.; Ritieni, A.; De Pascale, S.; Rouphael, Y. Genotype-specific modulatory effects of select spectral bandwidths on the nutritive and phytochemical composition of microgreens. Front. Plant Sci. 2019, 10, 1501. [Google Scholar] [CrossRef]
- Wojdyło, A.; Nowicka, P.; Tkacz, K.; Turkiewicz, I.P. Sprouts vs. microgreens as novel functional foods: Variation of nutritional and phytochemical profiles and their in vitro bioactive properties. Molecules 2020, 25, 4648. [Google Scholar] [CrossRef]
- Zhang, Y.; Xiao, Z.; Ager, E.; Kong, L.; Tan, L. Nutritional quality and health benefits of microgreens, a crop of modern agriculture. J. Future Food 2021, 1, 58–66. [Google Scholar] [CrossRef]
- Amengual, J. Bioactive properties of carotenoids in human health. Nutrients 2019, 11, 2388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrante, A.; Incrocci, L.; Maggini, R.; Serra, G.; Tognoni, F. Colour changes of fresh-cut leafy vegetables during storage. J. Food Agric. Environ. 2004, 2, 40–44. [Google Scholar]
- Lenzi, A.; Orlandini, A.; Bulgari, R.; Ferrante, A.; Bruschi, P. Antioxidant and mineral composition of three wild leafy species: A comparison between microgreens and baby greens. Foods 2019, 8, 487. [Google Scholar] [CrossRef] [Green Version]
- Renna, M.; Stellacci, A.M.; Corbo, F.; Santamaria, P. The use of a nutrient quality score is effective to assess the overall nutritional value of three Brassica microgreens. Foods 2020, 9, 1226. [Google Scholar] [CrossRef]
- Marchioni, I.; Martinelli, M.; Ascrizzi, R.; Gabbrielli, C.; Flamini, G.; Pistelli, L.; Pistelli, L. Small functional foods: Comparative phytochemical and nutritional analyses of five microgreens of the Brassicaceae family. Foods 2021, 10, 427. [Google Scholar] [CrossRef] [PubMed]
- Bulgari, R.; Negri, M.; Santoro, P.; Ferrante, A. Quality evaluation of indoor-grown microgreens cultivated on three different substrates. Horticulturae 2021, 7, 96. [Google Scholar] [CrossRef]
- Cavaiuolo, M.; Cocetta, G.; Bulgari, R.; Spinardi, A.; Ferrante, A. Identification of innovative potential quality markers in rocket and melon fresh-cut produce. Food Chem. 2015, 188, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Chandra, D.; Kim, J.G.; Kim, Y.P. Changes in microbial population and quality of microgreens treated with different sanitizers and packaging films. Hortic. Environ. Biotechnol. 2012, 53, 32–40. [Google Scholar] [CrossRef]
- Xiao, Z.; Luo, Y.; Lester, G.E.; Kou, L.; Yang, T.; Wang, Q. Postharvest quality and shelf life of radish microgreens as impacted by storage temperature, packaging film, and chlorine wash treatment. LWT Food Sci. Technol. 2014, 55, 551–558. [Google Scholar] [CrossRef]
- Tan, L.; Nuffer, H.; Feng, J.; Kwan, S.H.; Chen, H.; Tong, X.; Kong, L. Antioxidant properties and sensory evaluation of microgreens from commercial and local farms. Food Sci. Hum. Wellness 2020, 9, 45–51. [Google Scholar] [CrossRef]
- Niroula, A.; Khatri, S.; Timilsina, R.; Khadka, D.; Khadka, A.; Ojha, P. Profile of chlorophylls and carotenoids of wheat (Triticum aestivum L.) and barley (Hordeum vulgare L.) microgreens. J. Food Sci. Technol. 2019, 56, 2758–2763. [Google Scholar] [CrossRef]
- Santamaria, P. Nitrate in vegetables: Toxicity, content, intake and EC regulation. J. Sci. Food Agric. 2006, 86, 10–17. [Google Scholar] [CrossRef]
- European Commission Commission. Regulation (EU) No 1258/2011 of 2 December 2011 amending regulation (EC) No. 1881/2006 as regards maximum levels for nitrates in foodstuffs. Off. J. Eur. Union 2011, 320, 15–17. [Google Scholar]
- Xiao, Z.; Rausch, S.R.; Luo, Y.; Sun, J.; Yu, L.; Wang, Q.; Chen, P.; Yu, L.; Stommel, J.R. Microgreens of Brassicaceae: Genetic diversity of phytochemical concentrations and antioxidant capacity. LWT 2019, 101, 731–737. [Google Scholar] [CrossRef]
- Li, Z.; Lee, H.W.; Liang, X.; Liang, D.; Wang, Q.; Huang, D.; Ong, C.N. Profiling of phenolic compounds and antioxidant activity of 12 cruciferous vegetables. Molecules 2018, 23, 1139. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Choi, Y.; Jeong, H.S.; Lee, J.; Sung, J. Effect of different cooking methods on the content of vitamins and true retention in selected vegetables. Food Sci. Biotechnol. 2018, 27, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Ulla, H.; Antti, K.; Oskar, L.; Mari, S. Genetic basis of flavor sensitivity and food preferences. In Flavor; Etiévant, P., Guichard, E., Salles, C., Voilley, A., Eds.; Woodhead Publishing: Sawston, UK, 2016; pp. 203–227. [Google Scholar] [CrossRef]
- Balestrini, R.; Brunetti, C.; Cammareri, M.; Caretto, S.; Cavallaro, V.; Cominelli, E.; De Palma, M.; Docimo, T.; Giovinazzo, G.; Grandillo, S.; et al. Strategies to Modulate Specialized Metabolism in Mediterranean Crops: From Molecular Aspects to Field. Int. J. Mol. Sci. 2021, 22, 2887. [Google Scholar] [CrossRef] [PubMed]
- Cartea, M.E.; Francisco, M.; Soengas, P.; Velasco, P. Phenolic compounds in Brassica vegetables. Molecules 2011, 16, 251. [Google Scholar] [CrossRef] [PubMed]
- Kyriacou, M.C.; Rouphael, Y. Towards a new definition of quality for fresh fruits and vegetables. Sci. Hortic. 2018, 234, 463–469. [Google Scholar] [CrossRef]
- Hura, K.; Hura, T.; Dziurka, K.; Dziurka, M. Carbohydrate, phenolic and antioxidant level in relation to chlorophyll a content in oilseed winter rape (Brassica napus L.) inoculated with Leptosphaeria maculans. Eur. J. Plant Pathol. 2015, 143, 291–303. [Google Scholar] [CrossRef] [Green Version]
- Neugart, S.; Baldermann, S.; Hanschen, F.S.; Klopsch, R.; Wiesner-Reinhold, M.; Schreiner, M. The intrinsic quality of brassicaceous vegetables: How secondary plant metabolites are affected by genetic, environmental, and agronomic factors. Sci. Hortic. 2018, 233, 460–478. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| df | MS | F | p | |
|---|---|---|---|---|
| Turnip greens | ||||
| FY | 3 | 25.4626 | 12.97 | 0.0019 |
| DY | 3 | 7026.77 | 9.57 | 0.0050 |
| H | 3 | 4.5819 | 11.4107 | ≤0.001 |
| FW | 3 | 53.829 | 1.3702 | 0.3198 |
| DM | 3 | 0.12438 | 1.8585 | 0.2149 |
| Radish | ||||
| FY | 3 | 15,464.18 | 7.983 | 0.0086 |
| DY | 3 | 27.847 | 4.604 | 0.0374 |
| H | 3 | 0.1851 | 0.6289 | 0.5973 |
| FW | 3 | 5267.36 | 4.2204 | 0.0459 |
| DM | 3 | 1.2215 | 4.885 | 0.0324 |
| df | MS | F | p | |
|---|---|---|---|---|
| Turnip greens | ||||
| Chl a | 3 | 0.00183 | 1.823 | 0.221 |
| Chl b | 3 | 0.00074 | 1.326 | 0.332 |
| Total Chl | 3 | 0.00474 | 1.616 | 0.261 |
| Carotenoids | 3 | 0.000053 | 1.900 | 0.208 |
| Chl a/Chl b | 3 | 0.2063 | 1.287 | 0.3432 |
| Chl/Car | 3 | 3.0432 | 17.909 | ≤0.001 |
| Radish | ||||
| Chl a | 3 | 0.01134 | 23.019 | ≤0.001 |
| Chl b | 3 | 0.003206 | 12.823 | 0.002 |
| Total Chl | 3 | 0.02651 | 19.600 | ≤0.001 |
| Carotenoids | 3 | 0.000149 | 7.1013 | 0.012 |
| Chl a/Chl b | 3 | 0.1357 | 2.048 | 0.1857 |
| Chl/Car | 3 | 2.6302 | 16.900 | ≤0.001 |
| Chl a | Chl b | Total Chl | Carotenoids | Chl a/Chl b | Chl/Car | |
|---|---|---|---|---|---|---|
| (μg mg−1 FW) | ||||||
| Turnip greens | ||||||
| Control | 0.199 ± 0.022 | 0.084 ± 0.019 | 0.283 ± 0.041 | 0.045 ± 0.002 | 2.484 ± 0.266 | 3.192 ± 0.177a |
| Bio-1 | 0.171 ± 0.002 | 0.055 ± 0.004 | 0.226 ± 0.007 | 0.045 ± 0.001 | 3.116 ± 0.196 | 1.151 ± 0.093b |
| Bio-2 | 0.142 ± 0.015 | 0.050 ± 0.006 | 0.192 ± 0.021 | 0.037 ± 0.004 | 2.885 ± 0.049 | 1.005 ± 0.123b |
| Bio-3 | 0.188 ± 0.025 | 0.071 ± 0.018 | 0.260 ± 0.042 | 0.046 ± 0.004 | 2.783 ± 0.319 | 1.515 ± 0.414a |
| Significance | ns | ns | ns | ns | ns | *** |
| Radish | ||||||
| Control | 0.198 ± 0.008c | 0.072 ± 0.005b | 0.270 ± 0.013b | 0.049 ± 0.002b | 2.761 ± 0.089 | 1.479 ± 0.087b |
| Bio-1 | 0.267 ± 0.017b | 0.099 ± 0.016b | 0.366 ± 0.033b | 0.058 ± 0.002ab | 2.768 ± 0.261 | 2.340 ± 0.375b |
| Bio-2 | 0.246 ± 0.011c | 0.093 ± 0.003b | 0.340 ± 0.013b | 0.055 ± 0.005ab | 2.639 ± 0.110 | 2.097 ± 0.034b |
| Bio-3 | 0.346 ± 0.013a | 0.149 ± 0.006a | 0.495 ± 0.020a | 0.066 ± 0.001a | 2.314 ± 0.005 | 3.699 ± 0.241a |
| Significance | *** | ** | *** | ** | ns | *** |
| df | MS | F | p | |
|---|---|---|---|---|
| Turnip greens | ||||
| TS | 3 | 0.00196 | 4.803 | 0.034 |
| Nitrates | 3 | 624.46 | 0.168 | 0.915 |
| TPC | 3 | 330.006 | 6.275 | 0.017 |
| AscA | 3 | 34.015 | 8.1963 | 0.008 |
| DPPH | 3 | 14,671.44 | 19.53 | ≤0.001 |
| Radish | ||||
| TS | 3 | 0.01399 | 72.662 | ≤0.001 |
| Nitrates | 3 | 29,139.92 | 13.53 | 0.0017 |
| TPC | 3 | 771.027 | 6.999 | 0.0126 |
| AscA | 3 | 170.903 | 41.574 | ≤0.001 |
| DPPH | 3 | 37,524.8 | 32.504 | ≤0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toscano, S.; Romano, D.; Patanè, C. Effect of Application of Biostimulants on the Biomass, Nitrate, Pigments, and Antioxidants Content in Radish and Turnip Microgreens. Agronomy 2023, 13, 145. https://doi.org/10.3390/agronomy13010145
Toscano S, Romano D, Patanè C. Effect of Application of Biostimulants on the Biomass, Nitrate, Pigments, and Antioxidants Content in Radish and Turnip Microgreens. Agronomy. 2023; 13(1):145. https://doi.org/10.3390/agronomy13010145
Chicago/Turabian StyleToscano, Stefania, Daniela Romano, and Cristina Patanè. 2023. "Effect of Application of Biostimulants on the Biomass, Nitrate, Pigments, and Antioxidants Content in Radish and Turnip Microgreens" Agronomy 13, no. 1: 145. https://doi.org/10.3390/agronomy13010145