
Agronomic and Physiological Indices for Reproductive Stage Heat Stress Tolerance in Green Super Rice

 ,
,  , ,
, ,  , ,
, ,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site and Design
2.2. Cell Membrane Thermostability
2.3. Pollen Fertility Test
2.4. NDVI (Normalized Difference Vegetation Index)
2.5. Agronomic Parameters and Heat Susceptiblity Index
2.6. Statistical Analysis
3. Results
3.1. Assessment of Genetic Diversity Using Principal Component Analysis
3.2. Analysis of Variance Showed Significant Variation in Green Super Rice
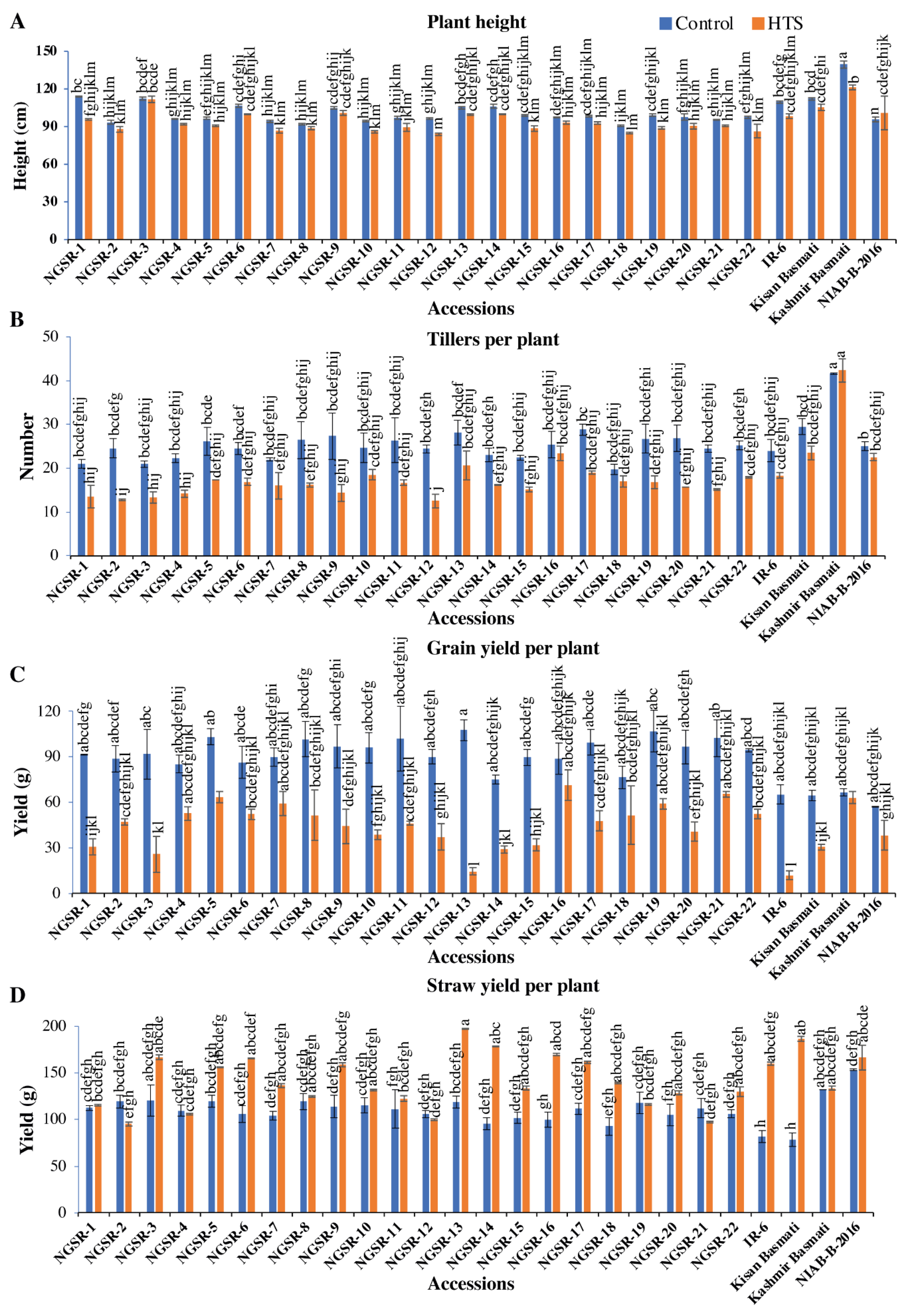
3.3. Mean Performance of Green Super Rice under Heat Stress
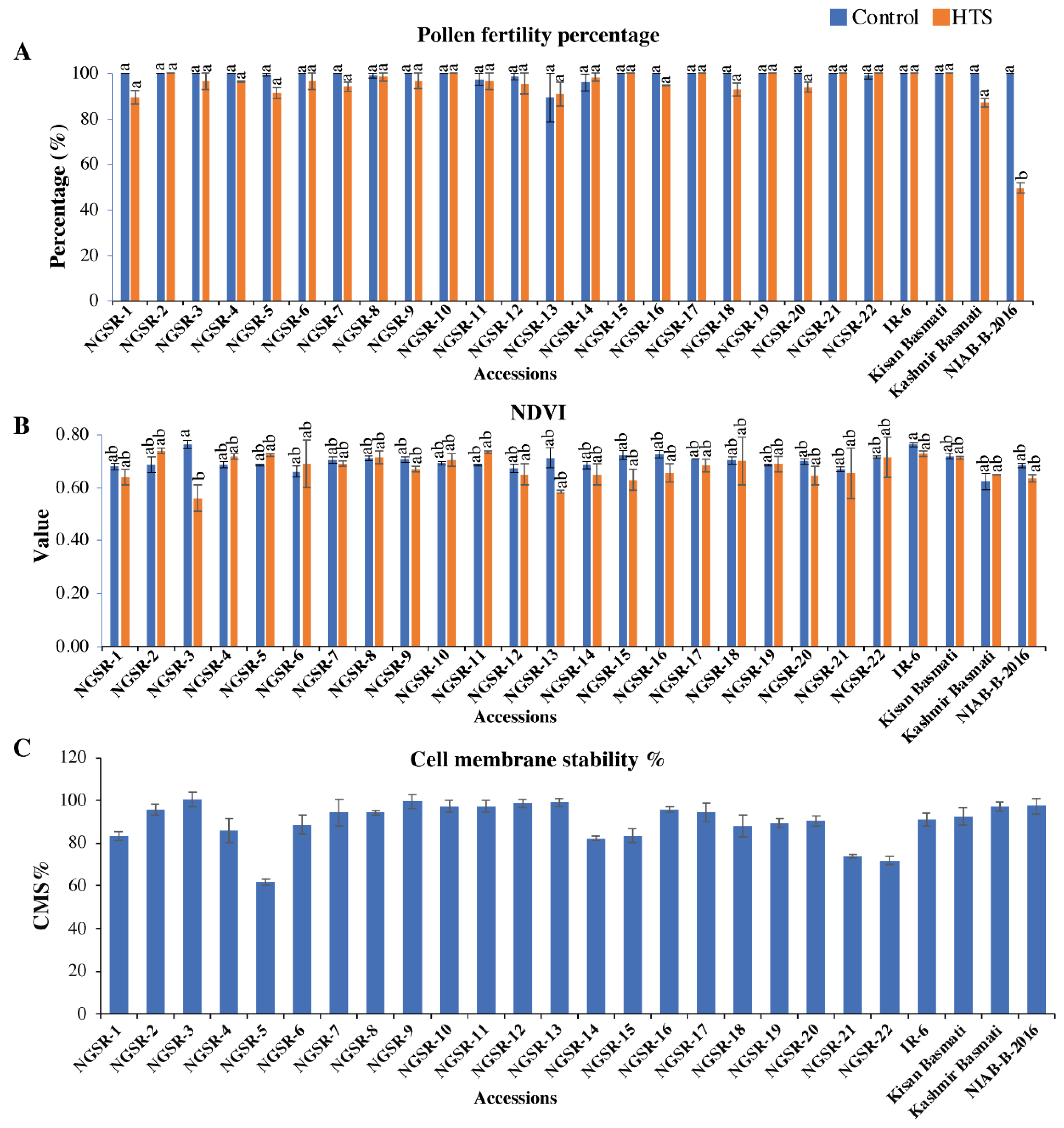
3.4. Normalized Difference Vegetative Index (NDVI)
3.5. Effect of Heat Stress on Cell Membrane Stability (CMS%)
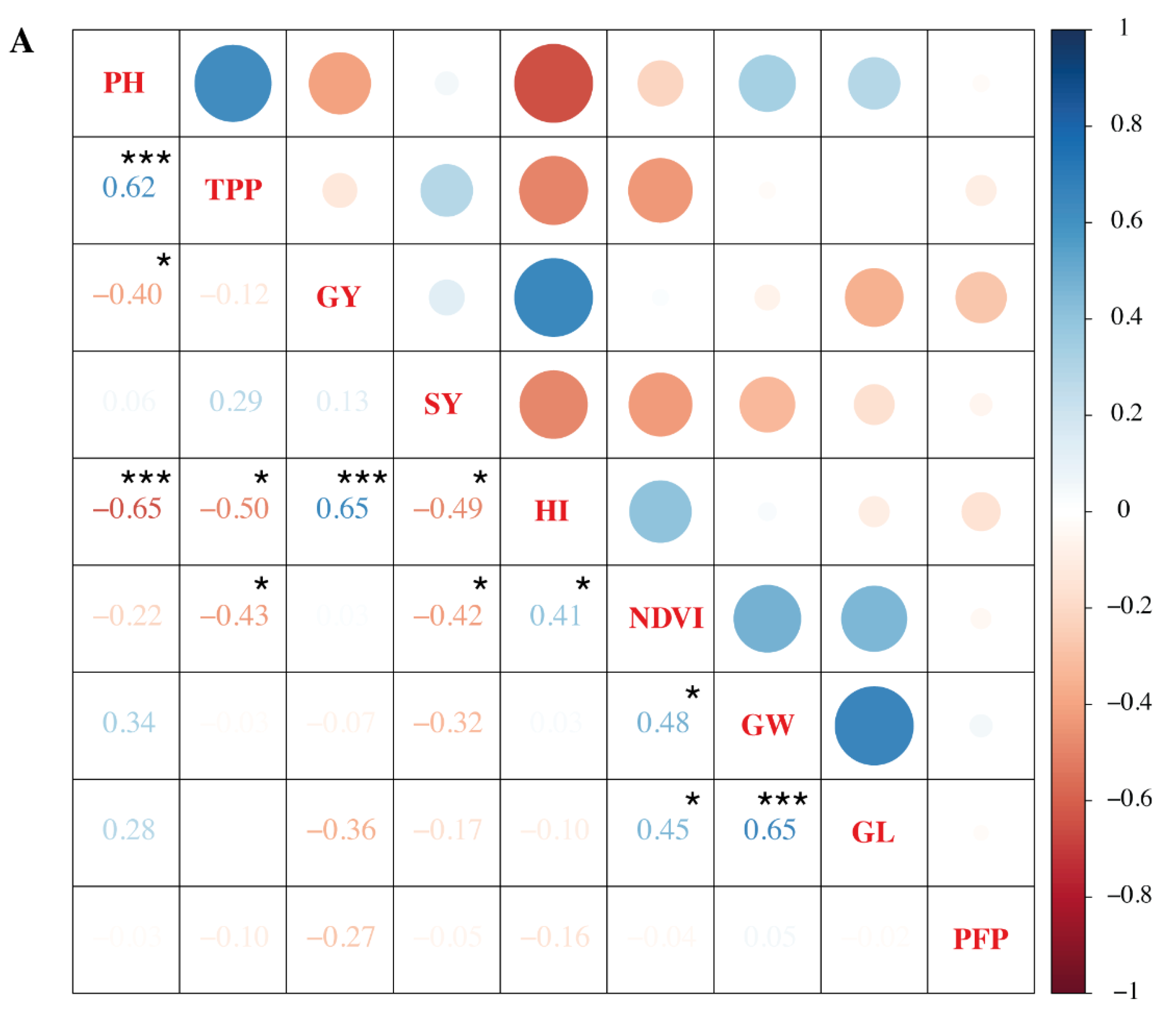
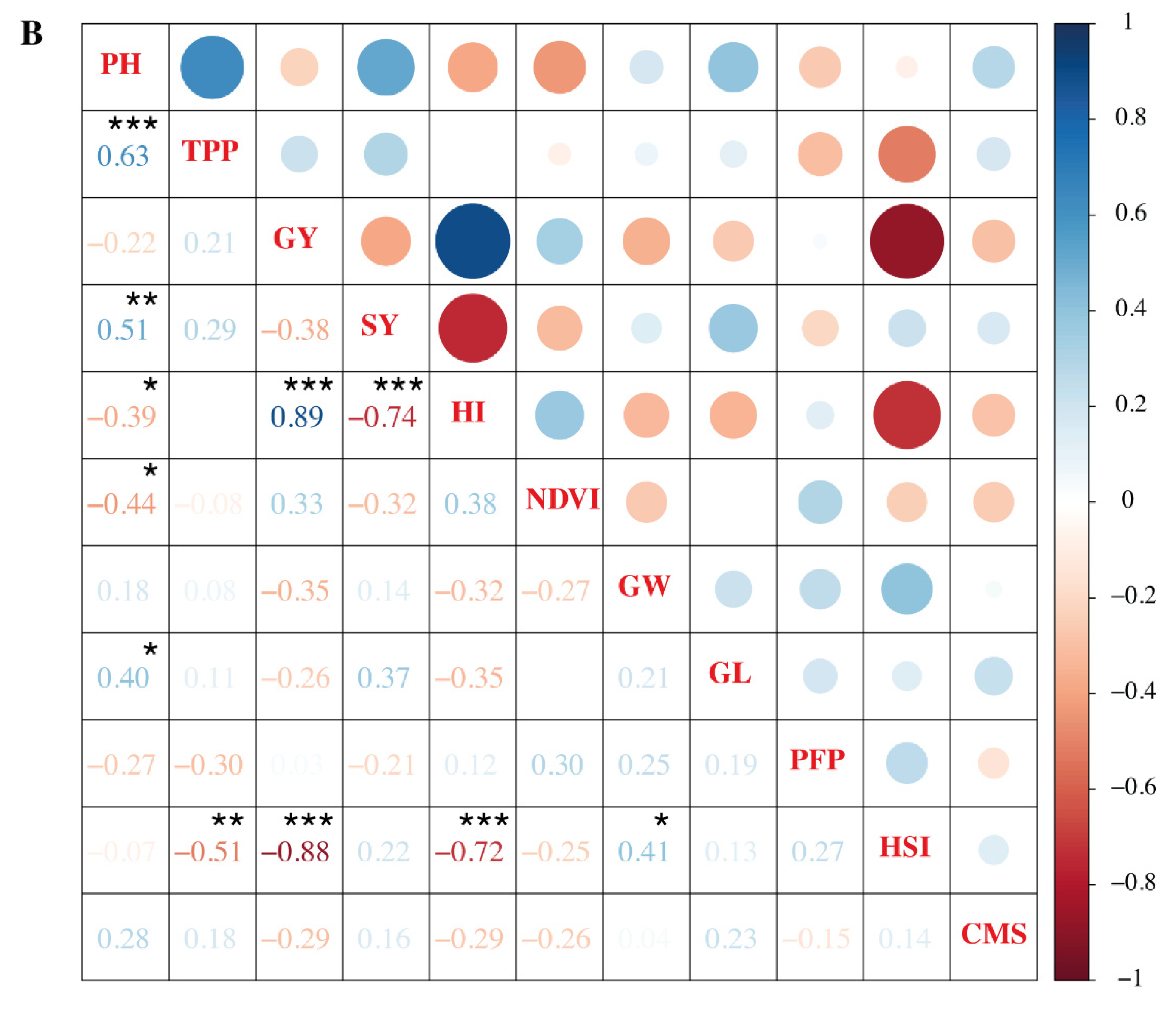
3.6. Association of Grain Yield and Other Parameters under Heat Stress
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Compliance with Ethical Standards
References
- Zafar, S.A.; Hameed, A.; Nawaz, M.A.; Ma, W.; Noor, M.A.; Hussain, M.; Mehboobur, R. Mechanisms and molecular approaches for heat tolerance in rice (Oryza sativa L.) under climate change scenario. J. Integr. Agric. 2018, 17, 726–738. [Google Scholar] [CrossRef]
- Fukagawa, N.K.; Ziska, L.H. Rice: Importance for global nutrition. J. Nutr. Sci. Vitaminol. 2019, 65, S2–S3. [Google Scholar] [CrossRef] [PubMed]
- Mahajan, G.; Sekhon, N.; Singh, N.; Kaur, R.; Sidhu, A. Yield and nitrogen-use efficiency of aromatic rice cultivars in response to nitrogen fertilizer. J. New Seeds 2010, 11, 356–368. [Google Scholar] [CrossRef]
- Ahmad, H.; Zafar, S.; Naeem, M.; Shokat, S.; Inam, S.; Rehman, M.A.u.; Naveed, S.A.; Xu, J.; Li, Z.; Ali, G.; et al. Impact of Pre-Anthesis Drought Stress on Physiology, Yield-Related Traits, and Drought-Responsive Genes in Green Super Rice. Front. Genet. 2022, 13, 832542. [Google Scholar] [CrossRef]
- Uzair, M.; Patil, S.B.; Zhang, H.; Kumar, A.; Mkumbwa, H.; Zafar, S.A.; Chun, Y.; Fang, J.; Zhao, J.; Khan, M.R. Screening Direct Seeding-Related Traits by Using an Improved Mesocotyl Elongation Assay and Association between Seedling and Maturity Traits in Rice. Agronomy 2022, 12, 975. [Google Scholar] [CrossRef]
- Raza, A.; Tabassum, J.; Kudapa, H.; Varshney, R.K. Can omics deliver temperature resilient ready-to-grow crops? Crit. Rev. Biotechnol. 2021, 41, 1209–1232. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Yu, H.; Qu, J.; Cao, Y.; Ding, L.; Feng, W.; Khalid, M.H.B.; Li, W.; Fu, F. Maize ZmBES1/BZR1-5 Decreases ABA Sensitivity and Confers Tolerance to Osmotic Stress in Transgenic Arabidopsis. Int. J. Mol. Sci. 2020, 21, 996. [Google Scholar] [CrossRef]
- Lu, F.; Li, W.; Peng, Y.; Cao, Y.; Qu, J.; Sun, F.; Yang, Q.; Lu, Y.; Zhang, X.; Zheng, L.; et al. ZmPP2C26 Alternative Splicing Variants Negatively Regulate Drought Tolerance in Maize. Front. Plant Sci. 2022, 13, 851531. [Google Scholar] [CrossRef]
- Feng, W.; Liu, Y.; Cao, Y.; Zhao, Y.; Zhang, H.; Sun, F.; Yang, Q.; Li, W.; Lu, Y.; Zhang, X.; et al. Maize ZmBES1/BZR1-3 and -9 Transcription Factors Negatively Regulate Drought Tolerance in Transgenic Arabidopsis. Int. J. Mol. Sci. 2022, 23, 6025. [Google Scholar] [CrossRef]
- Li, B.; Gao, K.; Ren, H.; Tang, W. Molecular mechanisms governing plant responses to high temperatures. J. Integr. Plant Biol. 2018, 60, 757–779. [Google Scholar] [CrossRef] [PubMed]
- Waqas, M.A.; Wang, X.; Zafar, S.A.; Noor, M.A.; Hussain, H.A.; Azher Nawaz, M.; Farooq, M. Thermal Stresses in Maize: Effects and Management Strategies. Plants 2021, 10, 293. [Google Scholar] [CrossRef] [PubMed]
- Raza, A. Metabolomics: A systems biology approach for enhancing heat stress tolerance in plants. Plant Cell Rep. 2020, 41, 741–763. [Google Scholar] [CrossRef] [PubMed]
- Tollefson, J. How hot will Earth get by 2100? Nature 2020, 580, 443–446. [Google Scholar] [CrossRef] [PubMed]
- Stocker, T. Climate Change 2013: The Physical Science Basis: Working Group I Contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Sun, Q.; Miao, C.; Hanel, M.; Borthwick, A.G.; Duan, Q.; Ji, D.; Li, H. Global heat stress on health, wildfires, and agricultural crops under different levels of climate warming. Environ. Int. 2019, 128, 125–136. [Google Scholar] [CrossRef]
- Shuangyi, L.; Zhihong, J.; Jieming, C.; Gang, T.; Shuyu, W. Response of temperature-related rice disaster to different warming levels under RCP8. 5 emission scenario in major rice production region of China. Front. Clim. 2022, 3, 736459. [Google Scholar] [CrossRef]
- Shah, F.; Huang, J.; Cui, K.; Nie, L.; Shah, T.; Chen, C.; Wang, K. Impact of high-temperature stress on rice plant and its traits related to tolerance. J. Agric. Sci. 2011, 149, 545–556. [Google Scholar] [CrossRef]
- Aryan, S.; Gulab, G.; Habibi, N.; Kakar, K.; Sadat, M.I.; Zahid, T.; Rashid, R.A. Phenological and physiological responses of hybrid rice under different high-temperature at seedling stage. Bull. Natl. Res. Cent. 2022, 46, 45. [Google Scholar] [CrossRef]
- Chunlin, S.; Zongqiang, L.; Min Jiang, Y.S.; Yingxue, L.; Shouli, X.; Yang, L.; Shenbin, Y.; Gengkang, Y. An quantitative analysis of high temperature effects during meiosis stage on rice grain number per panicle. Chin. J. Rice Sci. 2017, 31, 658. [Google Scholar]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.; Roychowdhury, R.; Fujita, M. Physiological, biochemical, and molecular mechanisms of heat stress tolerance in plants. Int. J. Mol. Sci. 2013, 14, 9643–9684. [Google Scholar] [CrossRef]
- Liu, W.; Yin, T.; Zhao, Y.; Wang, X.; Wang, K.; Shen, Y.; Ding, Y.; Tang, S. Effects of high temperature on rice grain development and quality formation based on proteomics comparative analysis under field warming. Front. Plant Sci. 2021, 12, 746180. [Google Scholar] [CrossRef]
- Nagai, T.; Makino, A. Differences between rice and wheat in temperature responses of photosynthesis and plant growth. Plant Cell Physiol. 2009, 50, 744–755. [Google Scholar] [CrossRef] [PubMed]
- Zafar, S.A.; Hameed, A.; Khan, A.S.; Ashraf, M. Heat shock induced morpho-physiological response in indica rice (Oryza sativa L.) at early seedling stage. Pak. J. Bot 2017, 49, 453–463. [Google Scholar]
- Zafar, S.A.; Hameed, A.; Ashraf, M.; Khan, A.S.; Li, X.; Siddique, K.H. Agronomic, physiological and molecular characterisation of rice mutants revealed the key role of reactive oxygen species and catalase in high-temperature stress tolerance. Funct. Plant Biol. 2020, 47, 440–453. [Google Scholar] [CrossRef]
- Jagadish, S.K.; Craufurd, P.Q.; Wheeler, T. High temperature stress and spikelet fertility in rice (Oryza sativa L.). J. Exp. Bot. 2007, 58, 1627–1635. [Google Scholar] [CrossRef] [PubMed]
- Satake, T.; Yoshida, S. High temperature-induced sterility in indica rices at flowering. Jpn. J. Crop Sci. 1978, 47, 6–17. [Google Scholar] [CrossRef]
- Mathur, S.; Agrawal, D.; Jajoo, A. Photosynthesis: Response to high temperature stress. J. Photochem. Photobiol. B Biol. 2014, 137, 116–126. [Google Scholar] [CrossRef]
- Wu, C.; Cui, K.; Wang, W.; Li, Q.; Fahad, S.; Hu, Q.; Huang, J.; Nie, L.; Mohapatra, P.K.; Peng, S. Heat-induced cytokinin transportation and degradation are associated with reduced panicle cytokinin expression and fewer spikelets per panicle in rice. Front. Plant Sci. 2017, 8, 371. [Google Scholar] [CrossRef]
- Ya-liang, W.; Yu-ping, Z.; Yan-hua, Z.; Hui, W.; Jing, X.; Hui-zhe, C.; Yi-kai, Z.; De-feng, Z. Effect of high temperature stress on rice spikelet differentiation and degeneration during panicle initiation stage. Chin. J. Agrometeorol. 2015, 36, 724. [Google Scholar]
- Fu, G.; Zhang, C.; Yang, X.; Yang, Y.; Chen, T.; Zhao, X.; Fu, W.; Feng, B.; Zhang, X.; Tao, L. Action mechanism by which SA alleviates high temperature-induced inhibition to spikelet differentiation. Chin. J. Rice Sci. 2015, 29, 637–647. [Google Scholar]
- Zafar, S.A.; Patil, S.B.; Uzair, M.; Fang, J.; Zhao, J.; Guo, T.; Yuan, S.; Uzair, M.; Luo, Q.; Shi, J. DEGENERATED PANICLE AND PARTIAL STERILITY 1 (DPS1) encodes a cystathionine β-synthase domain containing protein required for anther cuticle and panicle development in rice. New Phytol. 2020, 225, 356–375. [Google Scholar] [CrossRef]
- Xu, Y.; Chu, C.; Yao, S. The impact of high-temperature stress on rice: Challenges and solutions. Crop J. 2021, 9, 963–976. [Google Scholar] [CrossRef]
- Deng, Y.; Tian, X.; Wu, C.; Matsui, T.; Xiao, B. Early signs of heat stress-induced abnormal development of anther in rice. Zhongguo Shengtai Nongye Xuebao Chin. J. Eco-Agric. 2010, 18, 377–383. [Google Scholar] [CrossRef]
- Zhao, L.; Zhu, Z.; Tian, X.; Kobayasi, K.; Hasegawa, T.; Zhang, Y.; Chen, Z.; Wang, C.; Matsui, T. Inheritance analysis of anther dehiscence as a trait for the heat tolerance at flowering in japonica hybrid rice (Oryza sativa L.). Euphytica 2016, 211, 311–320. [Google Scholar] [CrossRef]
- Hu, Q.; Wang, W.; Lu, Q.; Huang, J.; Peng, S.; Cui, K. Abnormal anther development leads to lower spikelet fertility in rice (Oryza sativa L.) under high temperature during the panicle initiation stage. BMC Plant Biol. 2021, 21, 428. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, L.; Zhou, J.; Hu, S.; Chen, H.; Xiang, J.; Zhang, Y.; Zeng, Y.; Shi, Q.; Zhu, D. Research progress on heat stress of rice at flowering stage. Rice Sci. 2019, 26, 1–10. [Google Scholar] [CrossRef]
- Zhang, C.; Li, G.; Chen, T.; Feng, B.; Fu, W.; Yan, J.; Islam, M.R.; Jin, Q.; Tao, L.; Fu, G. Heat stress induces spikelet sterility in rice at anthesis through inhibition of pollen tube elongation interfering with auxin homeostasis in pollinated pistils. Rice 2018, 11, 14. [Google Scholar] [CrossRef] [PubMed]
- Santiago, J.P.; Sharkey, T.D. Pollen development at high temperature and role of carbon and nitrogen metabolites. Plant Cell Environ. 2019, 42, 2759–2775. [Google Scholar] [CrossRef] [PubMed]
- Rieu, I.; Twell, D.; Firon, N. Pollen development at high temperature: From acclimation to collapse. Plant Physiol. 2017, 173, 1967–1976. [Google Scholar] [CrossRef] [PubMed]
- Panda, D.; Mishra, S.S.; Behera, P.K. Drought tolerance in rice: Focus on recent mechanisms and approaches. Rice Sci. 2021, 28, 119–132. [Google Scholar] [CrossRef]
- Xiong, H.; Yu, J.; Miao, J.; Li, J.; Zhang, H.; Wang, X.; Liu, P.; Zhao, Y.; Jiang, C.; Yin, Z. Natural variation in OsLG3 increases drought tolerance in rice by inducing ROS scavenging. Plant Physiol. 2018, 178, 451–467. [Google Scholar] [CrossRef]
- Ali, J.; Anumalla, M.; Murugaiyan, V.; Li, Z. Green super rice (GSR) traits: Breeding and genetics for multiple biotic and abiotic stress tolerance in rice. In Rice Improvement; Springer: Berlin/Heidelberg, Germany, 2021; pp. 59–97. [Google Scholar]
- Zaid, I.U.; Zahra, N.; Habib, M.; Naeem, M.K.; Asghar, U.; Uzair, M.; Latif, A.; Rehman, A.; Ali, G.M.; Khan, M.R. Estimation of genetic variances and stability components of yield-related traits of Green Super Rice at multi-environmental conditions of Pakistan. Agronomy 2022, 12, 1157. [Google Scholar] [CrossRef]
- Jewel, Z.A.; Ali, J.; Pang, Y.; Mahender, A.; Acero, B.; Hernandez, J.; Xu, J.; Li, Z. Developing green super rice varieties with high nutrient use efficiency by phenotypic selection under varied nutrient conditions. Crop J. 2019, 7, 368–377. [Google Scholar] [CrossRef]
- Yu, S.; Ali, J.; Zhang, C.; Li, Z.; Zhang, Q. Genomic Breeding of Green Super Rice Varieties and Their Deployment in Asia and Africa. Appl. Genet. 2020, 133, 1427–1442. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q. Strategies for developing green super rice. Proc. Natl. Acad. Sci. USA 2007, 104, 16402–16409. [Google Scholar] [CrossRef]
- Chaturvedi, A.K.; Bahuguna, R.N.; Shah, D.; Pal, M.; Jagadish, S. High temperature stress during flowering and grain filling offsets beneficial impact of elevated CO2 on assimilate partitioning and sink-strength in rice. Sci. Rep. 2017, 7, 8227. [Google Scholar] [CrossRef] [PubMed]
- Tripathy, J.; Zhang, J.; Robin, S.; Nguyen, T.T.; Nguyen, H. QTLs for cell-membrane stability mapped in rice (Oryza sativa L.) under drought stress. Theor. Appl. Genet. 2000, 100, 1197–1202. [Google Scholar] [CrossRef]
- Govaerts, B.; Verhulst, N. The Normalized Difference Vegetation Index (NDVI) Greenseeker (TM) Handheld Sensor: Toward the Integrated Evaluation of Crop Management Part A: Concepts and Case Studies; CIMMYT: Mexico City, Mexico, 2010. [Google Scholar]
- Shanmugavadivel, P.; Sv, A.M.; Prakash, C.; Ramkumar, M.; Tiwari, R.; Mohapatra, T.; Singh, N.K. High resolution mapping of QTLs for heat tolerance in rice using a 5K SNP array. Rice 2017, 10, 28. [Google Scholar]
- Zafar, S.A.; Uzair, M.; Khan, M.R.; Patil, S.B.; Fang, J.; Zhao, J.; Singla-Pareek, S.L.; Pareek, A.; Li, X. DPS1 regulates cuticle development and leaf senescence in rice. Food Energy Secur. 2021, 10, e273. [Google Scholar] [CrossRef]
- Shi, W.; Ishimaru, T.; Gannaban, R.; Oane, W.; Jagadish, S. Popular rice (Oryza sativa L.) cultivars show contrasting responses to heat stress at gametogenesis and anthesis. Crop Sci. 2015, 55, 589–596. [Google Scholar] [CrossRef]
- Cheabu, S.; Moung-Ngam, P.; Arikit, S.; Vanavichit, A.; Malumpong, C. Effects of heat stress at vegetative and reproductive stages on spikelet fertility. Rice Sci. 2018, 25, 218–226. [Google Scholar] [CrossRef]
- Karwa, S.; Bahuguna, R.N.; Chaturvedi, A.K.; Maurya, S.; Arya, S.S.; Chinnusamy, V.; Pal, M. Phenotyping and characterization of heat stress tolerance at reproductive stage in rice (Oryza sativa L.). Acta Physiol. Plant. 2020, 42, 29. [Google Scholar] [CrossRef]
- Haider, S.; Raza, A.; Iqbal, J.; Shaukat, M.; Mahmood, T. Analyzing the regulatory role of heat shock transcription factors in plant heat stress tolerance: A brief appraisal. Mol. Biol. Rep. 2022, 49, 5771–5785. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.S.; Uzair, M.; Raza, A.; Habib, M.; Xu, Y.; Yousuf, M.; Yang, S.H.; Khan, M.R. Uncovering the Research Gaps to Alleviate the Negative Impacts of Climate Change on Food Security: A Review. Front. Plant Sci. 2022, 13, 927535. [Google Scholar] [CrossRef]
- Easterling, W.E.; Aggarwal, P.K.; Batima, P.; Brander, K.M.; Erda, L.; Howden, S.M.; Kirilenko, A.; Morton, J.; Soussana, J.-F.; Schmidhuber, J. Food, fibre and forest products. Clim. Chang. 2007, 2007, 273–313. [Google Scholar]
- Uzair, M.; Ali, M.; Fiaz, S.; Attia, K.; Khan, N.; Al-Doss, A.A.; Khan, M.R.; Ali, Z. The Characterization of Wheat Genotypes for Salinity Tolerance Using Morpho-Physiological Indices under Hydroponic Conditions. Saudi J. Biol. Sci. 2022, 29, 103299. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.; Sakai, H.; Yagi, K.; Hasegawa, T. Interactions of elevated [CO2] and night temperature on rice growth and yield. Agric. For. Meteorol. 2009, 149, 51–58. [Google Scholar] [CrossRef]
- Farooq, J.; Khaliq, I.; Ali, M.A.; Kashif, M.; Rehman, A.U.; Naveed, M.; Ali, Q.; Nazeer, W.; Farooq, A. Inheritance pattern of yield attributes in spring wheat at grain filling stage under different temperature regimes. Aust. J. Crop Sci. 2011, 5, 1745–1753. [Google Scholar]
- Ilík, P.; Špundová, M.; Šicner, M.; Melkovičová, H.; Kučerová, Z.; Krchňák, P.; Fürst, T.; Večeřová, K.; Panzarová, K.; Benediktyová, Z. Estimating heat tolerance of plants by ion leakage: A new method based on gradual heating. New Phytol. 2018, 218, 1278–1287. [Google Scholar] [CrossRef] [PubMed]
- Rehman, S.U.; Bilal, M.; Rana, R.M.; Tahir, M.N.; Shah, M.K.N.; Ayalew, H.; Yan, G. Cell membrane stability and chlorophyll content variation in wheat (Triticum aestivum) genotypes under conditions of heat and drought. Crop Pasture Sci. 2016, 67, 712–718. [Google Scholar] [CrossRef]
- Lu, Z.; Cui, J.; Wang, L.; Teng, N.; Zhang, S.; Lam, H.-M.; Zhu, Y.; Xiao, S.; Ke, W.; Lin, J. Genome-wide DNA mutations in Arabidopsis plants after multigenerational exposure to high temperatures. Genome Biol. 2021, 22, 160. [Google Scholar] [CrossRef]
- Iqbal, N.; Farooq, S.; Arshad, R.; Hameed, A. Differential accumulation of high and low molecular weight heat shock proteins in Basmati rice (Oryza sativa L.) cultivars. Genet. Resour. Crop Evol. 2010, 57, 65–70. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SOV | G | Rep | T | G × T | H2 |
|---|---|---|---|---|---|
| DF | 25 | 1 | 1 | 25 | |
| PH | 358.2 *** | 1.4 ns | 828.9 *** | 76.7 *** | 96.43 |
| TPP | 90.0 *** | 15.9 ns | 1155.6 *** | 10.0 ns | 92.60 |
| GY | 458 *** | 461 ns | 45,179 *** | 318 * | 64.85 |
| SY | 797 *** | 920 ns | 34,194 *** | 1158 *** | 63.36 |
| HI | 0.007 *** | 0.007 ns | 1.1028 *** | 0.0078 *** | 88.31 |
| GW | 18.26 *** | 15.69 * | 0.122 ns | 4.122 ns | 81.80 |
| GL | 3.09 *** | 0.08 ns | 1.1839 ** | 0.223 ns | 95.71 |
| NDVI | 0.003 ns | 0.000047 ns | 0.006 ns | 0.003 ns | 35.83 |
| PFP | 110.0 *** | 80.1 * | 467.0 *** | 107.4 *** | 85.18 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zafar, S.A.; Arif, M.H.; Uzair, M.; Rashid, U.; Naeem, M.K.; Rehman, O.U.; Rehman, N.; Zaid, I.U.; Farooq, M.S.; Zahra, N.; et al. Agronomic and Physiological Indices for Reproductive Stage Heat Stress Tolerance in Green Super Rice. Agronomy 2022, 12, 1907. https://doi.org/10.3390/agronomy12081907
Zafar SA, Arif MH, Uzair M, Rashid U, Naeem MK, Rehman OU, Rehman N, Zaid IU, Farooq MS, Zahra N, et al. Agronomic and Physiological Indices for Reproductive Stage Heat Stress Tolerance in Green Super Rice. Agronomy. 2022; 12(8):1907. https://doi.org/10.3390/agronomy12081907
Chicago/Turabian StyleZafar, Syed Adeel, Muhammad Hamza Arif, Muhammad Uzair, Umer Rashid, Muhammad Kashif Naeem, Obaid Ur Rehman, Nazia Rehman, Imdad Ullah Zaid, Muhammad Shahbaz Farooq, Nageen Zahra, and et al. 2022. "Agronomic and Physiological Indices for Reproductive Stage Heat Stress Tolerance in Green Super Rice" Agronomy 12, no. 8: 1907. https://doi.org/10.3390/agronomy12081907