
Cannabis sativa L.: Crop Management and Abiotic Factors That Affect Phytocannabinoid Production

 , , and
, , and 
Abstract
:1. Introduction
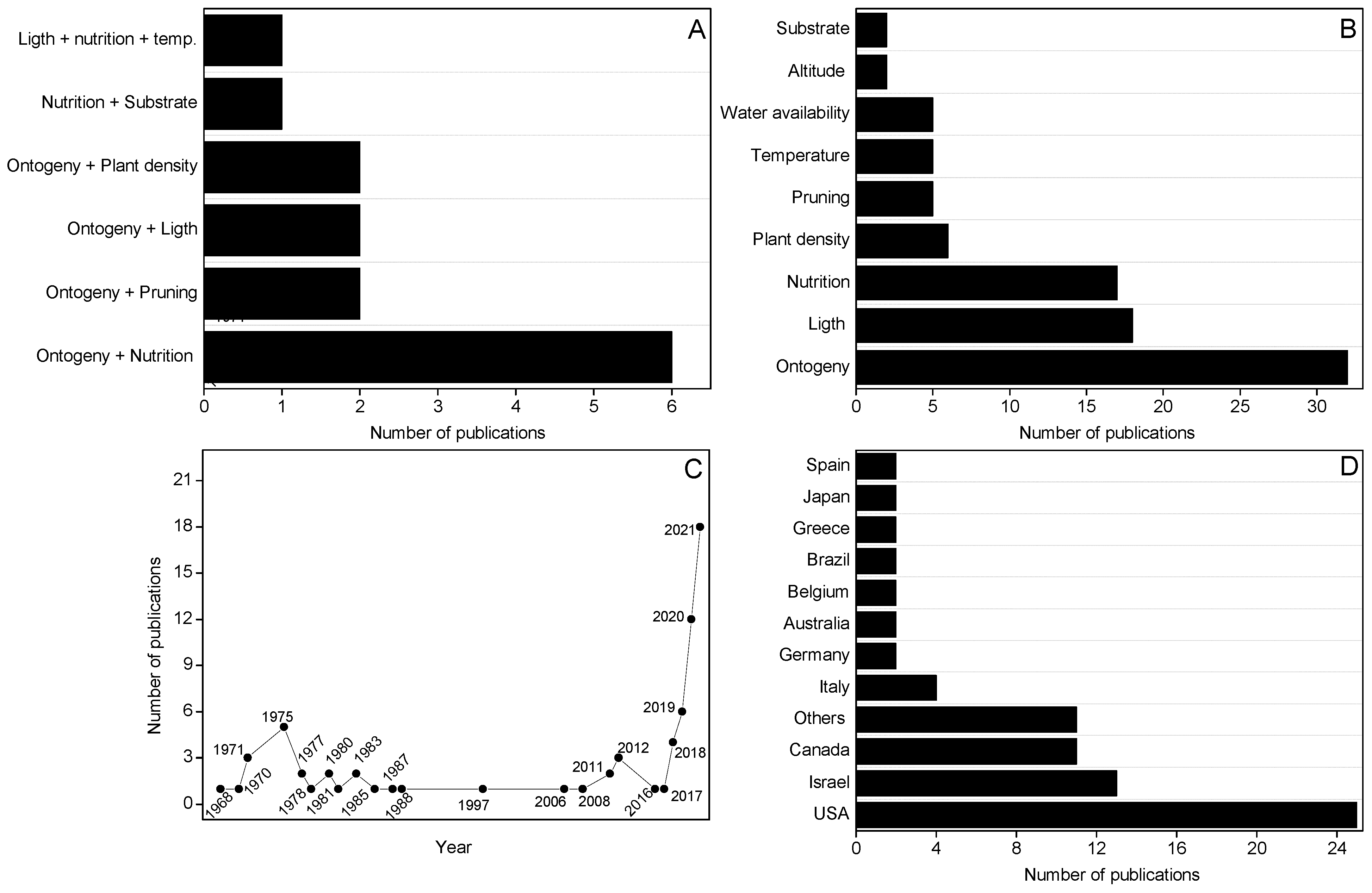
2. State-of-the-Art Research That Evaluates Crop Management and Abiotic Factors That Affect Phytocannabinoid Production
3. Phytocannabinoids Production and Management Techniques in Cannabis Cultivation
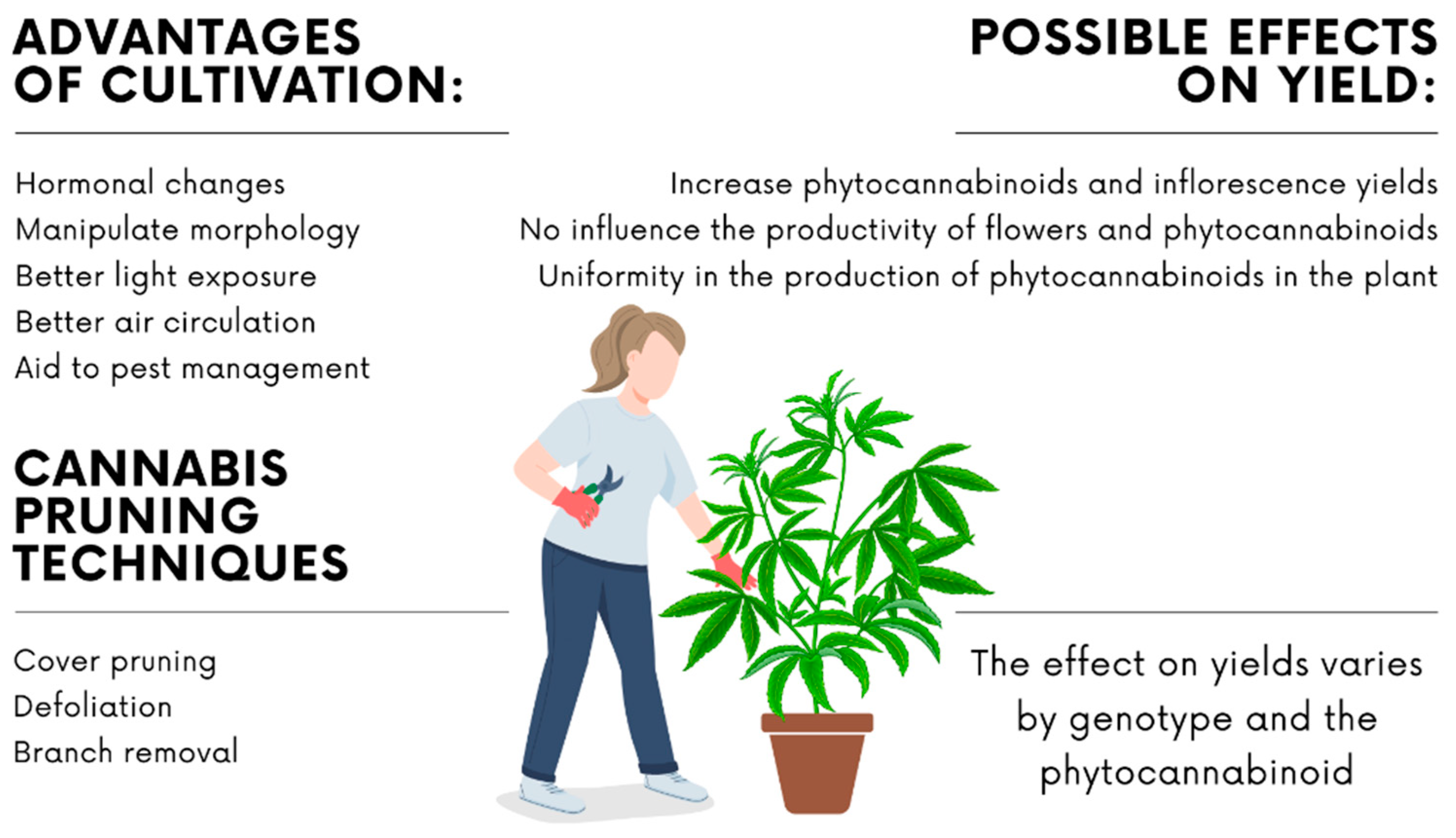
3.1. Pruning
3.2. Effects of Light
3.3. Plant Density
3.4. Ontogeny
3.5. Temperature, Altitude and CO2 Concentration
3.6. Nutrition and Substrate
3.7. Water Availability
4. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
References
- Musto, D.F. The Marihuana Tax Act of 1937. Arch. Gen. Psychiatry 1972, 26, 101. [Google Scholar] [CrossRef] [PubMed]
- Saloner, A.; Sacks, M.M.; Bernstein, N. Response of Medical Cannabis (Cannabis sativa L.) Genotypes to K Supply under Long Photoperiod. Front. Plant Sci. 2019, 10, 1369. [Google Scholar] [CrossRef] [PubMed]
- Li, H.-L. An Archaeological and Historical Account of Cannabis in China. Econ. Bot. 1974, 28, 437–448. [Google Scholar] [CrossRef]
- Singh Chopra, G. Man and Marijuana. Int. J. Addict. 1969, 4, 215–247. [Google Scholar] [CrossRef]
- Mechoulam, R.; Carlini, E.A. Toward Drugs Derived from Cannabis. Naturwissenschaften 1978, 65, 174–179. [Google Scholar] [CrossRef]
- McPartland, J.M. Cannabis Systematics at the Levels of Family, Genus, and Species. Cannabis Cannabinoid Res. 2018, 3, 203–212. [Google Scholar] [CrossRef] [Green Version]
- Mechoulam, R. Cannabis—The Israeli Perspective. J. Basic Clin. Physiol. Pharmacol. 2016, 27, 181–187. [Google Scholar] [CrossRef]
- Treister-Goltzman, Y.; Freud, T.; Press, Y.; Peleg, R. Trends in Publications on Medical Cannabis from the Year 2000. Popul. Health Manag. 2019, 22, 362–368. [Google Scholar] [CrossRef]
- Schluttenhofer, C.; Yuan, L. Challenges towards Revitalizing Hemp: A Multifaceted Crop. Trends Plant Sci. 2017, 22, 917–929. [Google Scholar] [CrossRef] [Green Version]
- UNODC WHO Scheduling Recommendations on Cannabis and Cannabis-Related Substances. Available online: https://www.unodc.org/unodc/en/commissions/CND/Mandate_Functions/current-scheduling-recommendations.html (accessed on 24 May 2021).
- Hussain, T.; Jeena, G.; Pitakbut, T.; Vasilev, N.; Kayser, O. Cannabis sativa Research Trends, Challenges, and New-Age Perspectives. iScience 2021, 24, 103391. [Google Scholar] [CrossRef]
- Andre, C.M.; Hausman, J.F.; Guerriero, G. Cannabis sativa: The Plant of the Thousand and One Molecules. Front. Plant Sci. 2016, 7, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Small, E. Evolution and Classification of Cannabis sativa (Marijuana, Hemp) in Relation to Human Utilization. Bot. Rev. 2015, 81, 189–294. [Google Scholar] [CrossRef]
- de Meijer, E.P.M.; Bagatta, M.; Carboni, A.; Crucitti, P.; Moliterni, V.M.C.; Ranalli, P.; Mandolino, G. The Inheritance of Chemical Phenotype in Cannabis sativa L. Genetics 2003, 163, 335–346. [Google Scholar] [CrossRef]
- Hesami, M.; Pepe, M.; Alizadeh, M.; Rakei, A.; Baiton, A.; Phineas Jones, A.M. Recent Advances in Cannabis Biotechnology. Ind. Crops Prod. 2020, 158, 113026. [Google Scholar] [CrossRef]
- ElSohly, M.A.; Radwan, M.M.; Gul, W.; Chandra, S.; Galal, A. Phytochemistry of Cannabis sativa L. Prog. Chem. Org. Nat. Prod. 2017, 103, 1–36. [Google Scholar]
- Faux, A.M.; Draye, X.; Flamand, M.C.; Occre, A.; Bertin, P. Identification of QTLs for Sex Expression in Dioecious and Monoecious Hemp (Cannabis sativa L.). Euphytica 2016, 209, 357–376. [Google Scholar] [CrossRef]
- Cristiana Moliterni, V.M.; Cattivelli, L.; Ranalli, P.; Mandolino, G. The Sexual Differentiation of Cannabis sativa L.: A Morphological and Molecular Study; Kluwer Academic Publishers: Norwell, MA, USA, 2004; Volume 140. [Google Scholar]
- Dayanandan, P.; Kaufman, P.B. Trichomes of Cannabis sativa L. (Cannabaceae). Am. J. Bot. 1976, 63, 578–591. [Google Scholar] [CrossRef] [Green Version]
- Stout, J.M.; Boubakir, Z.; Ambrose, S.J.; Purves, R.W.; Page, J.E. The Hexanoyl-CoA Precursor for Cannabinoid Biosynthesis Is Formed by an Acyl-Activating Enzyme in Cannabis sativa Trichomes. Plant J. 2012, 71, 353–365. [Google Scholar] [CrossRef] [Green Version]
- Upton, R.; Craker, L.; ElSohly, M.; Romm, A.; Russo, E.; Sexton, M. American Herbal Pharmacopoeia; CRC Press: Boca Raton, FL, USA, 2013; pp. 1–60. [Google Scholar]
- Tanney, C.A.S.; Backer, R.; Geitmann, A.; Smith, D.L. Cannabis Glandular Trichomes: A Cellular Metabolite Factory. Front. Plant Sci. 2021, 12, 1923. [Google Scholar] [CrossRef]
- Welling, M.T.; Deseo, M.A.; Bacic, A.; Doblin, M.S. Untargeted Metabolomic Analyses Reveal Chemical Complexity of Dioecious Cannabis Flowers. Aust. J. Chem. 2021, 74, 463–479. [Google Scholar] [CrossRef]
- Valle, J.R.; Lapa, A.J.; Barros, G.G. Pharmacological Activity of Cannabis According to the Sex of the Plant. J. Pharm. Pharmacol. 1968, 20, 798–799. [Google Scholar] [CrossRef] [PubMed]
- Ohlsson, A.; Abou-Chaar, C.I.; Agurell, S.; Nilsson, I.M.; OIofsson, K.; Sandberg, F. Cannabinoid Constituents of Male and Female Cannabis sativa. Bull. Narc. 1971, 23, 29–32. [Google Scholar]
- Fetterman, P.S.; Keith, E.S.; Waller, C.W.; Guerrero, O.; Doorenbos, N.J.; Quimby, M.W. Mississippi-grown Cannabis sativa L.: Preliminary Observation on Chemical Definition of Phenotype and Variations in Tetrahydrocannabinol Content versus Age, Sex, and Plant Part. J. Pharm. Sci. 1971, 60, 1246–1249. [Google Scholar] [CrossRef]
- Doorenbos, N.J.; Fetterman, P.S.; Quimby, M.W.; Turner, C.E. Cultivation, Extraction, and Analysis of Cannabis sativa L. Ann. N. Y. Acad. Sci. 1971, 191, 3–14. [Google Scholar] [CrossRef]
- Kushima, H.; Shoyama, Y.; Nishioka, I. Cannabis. XII. Variations of Cannabinoid Contents in Several Strains of Cannabis sativa L. with Leaf-Age, Season and Sex. Chem. Pharm. Bull. 1980, 28, 594–598. [Google Scholar] [CrossRef] [Green Version]
- Kimura, M.; Okamoto, K. Distribution of Tetrahydrocannabinolic Acid in Fresh Wild Cannabis. Experientia 1970, 26, 819–820. [Google Scholar] [CrossRef] [PubMed]
- Bruci, Z.; Papoutsis, I.; Athanaselis, S.; Nikolaou, P.; Pazari, E.; Spiliopoulou, C.; Vyshka, G. First Systematic Evaluation of the Potency of Cannabis sativa Plants Grown in Albania. Forensic Sci. Int. 2012, 222, 40–46. [Google Scholar] [CrossRef]
- Lipson Feder, C.; Cohen, O.; Shapira, A.; Katzir, I.; Peer, R.; Guberman, O.; Procaccia, S.; Berman, P.; Flaishman, M.; Meiri, D. Fertilization Following Pollination Predominantly Decreases Phytocannabinoids Accumulation and Alters the Accumulation of Terpenoids in Cannabis Inflorescences. Front. Plant Sci. 2021, 12, 753847. [Google Scholar] [CrossRef]
- Olejar, K.J.; Park, S.-H. Industry-Based Misconceptions Regarding Cross-Pollination of Cannabis spp. Front. Plant Sci. 2022, 13, 793264. [Google Scholar] [CrossRef]
- Adesina, I.; Bhowmik, A.; Sharma, H.; Shahbazi, A. A Review on the Current State of Knowledge of Growing Conditions, Agronomic Soil Health Practices and Utilities of Hemp in the United States. Agriculture 2020, 10, 129. [Google Scholar] [CrossRef] [Green Version]
- Giroud, C. Analysis of Cannabinoids in Hemp Plants. Chimia 2002, 56, 80–83. [Google Scholar] [CrossRef]
- Spitzer-Rimon, B.; Duchin, S.; Bernstein, N.; Kamenetsky, R. Architecture and Florogenesis in Female Cannabis sativa Plants. Front. Plant Sci. 2019, 10, 350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandra, S.; Lata, H.; ElSohly, M.A.; Walker, L.A.; Potter, D. Cannabis Cultivation: Methodological Issues for Obtaining Medical-Grade Product. Epilepsy Behav. 2017, 70, 302–312. [Google Scholar] [CrossRef] [PubMed]
- Riboulet-Zemouli, K. ‘Cannabis’ Ontologies I: Conceptual Issues with Cannabis and Cannabinoids Terminology. Drug Sci. Policy Law 2020, 6, 205032452094579. [Google Scholar] [CrossRef]
- Gorguet, B.; van Heusden, A.W.; Lindhout, P. Parthenocarpic Fruit Development in Tomato. Plant Biol. 2005, 7, 131–139. [Google Scholar] [CrossRef]
- European Monitoring Centre for Drugs and Drug Addiction (EMCDDA). Cannabis Production and Markets in Europe; Publications Office of the European Union: Luxembourg, 2012; p. 268.
- Clarke, R.C. Marijuana Botany: The Propagation and Breeding of Distintive Cannabis, 2nd ed.; Ronin Publishing: Berkeley, CA, USA, 1981. [Google Scholar]
- Potter, D.J. A Review of the Cultivation and Processing of Cannabis (Cannabis Sativa L.) for Production of Prescription Medicines in the UK. Drug Test. Anal. 2014, 6, 31–38. [Google Scholar] [CrossRef]
- van der Werf, H.M.G.; Haasken, H.J.; Wijlhuizen, M. The Effect of Daylength on Yield and Quality of Fibre Hemp (Cannabis sativa L.). Eur. J. Agron. 1994, 3, 117–123. [Google Scholar] [CrossRef]
- Isah, T. Stress and Defense Responses in Plant Secondary Metabolites Production. Biol. Res. 2019, 52, 39. [Google Scholar] [CrossRef] [Green Version]
- Theis, N.; Lerdau, M. The Evolution of Function in Plant Secondary Metabolites. Int. J. Plant Sci. 2003, 164, S93–S102. [Google Scholar] [CrossRef]
- Haney, A.; Kutscheid, B.B. Quantitative Variation in the Chemical Constituents of Marihuana from Stands of Naturalized Cannabis sativa L. in East-Central Illinois. Econ. Bot. 1973, 27, 193–203. [Google Scholar] [CrossRef]
- Danziger, N.; Bernstein, N. Plant Architecture Manipulation Increases Cannabinoid Standardization in ‘Drug-Type’ Medical Cannabis. Ind. Crops Prod. 2021, 167, 113528. [Google Scholar] [CrossRef]
- Lermusieau, G.; Collin, S. Hop Aroma Extraction and Analysis. In Analysis of Taste and Aroma; Springer: Berlin/Heidelberg, Germany, 2002; pp. 69–88. [Google Scholar] [CrossRef]
- Bevan, L.; Jones, M.; Zheng, Y. Optimisation of Nitrogen, Phosphorus, and Potassium for Soilless Production of Cannabis sativa in the Flowering Stage Using Response Surface Analysis. Front. Plant Sci. 2021, 12, 2587. [Google Scholar] [CrossRef] [PubMed]
- Coffman, C.B.; Gentner, W.A. Cannabinoid Profile and Elemental Uptake of Cannabis sativa L. as Influenced by Soil Characteristics 1. Agron. J. 1975, 67, 491–497. [Google Scholar] [CrossRef]
- Naim-Feil, E.; Breen, E.J.; Pembleton, L.W.; Spooner, L.E.; Spangenberg, G.C.; Cogan, N.O.I. Empirical Evaluation of Inflorescences’ Morphological Attributes for Yield Optimization of Medicinal Cannabis Cultivars. Front. Plant Sci. 2022, 13, 858519. [Google Scholar] [CrossRef]
- Fischedick, J.T.; Hazekamp, A.; Erkelens, T.; Choi, Y.H.; Verpoorte, R. Metabolic Fingerprinting of Cannabis sativa L., Cannabinoids and Terpenoids for Chemotaxonomic and Drug Standardization Purposes. Phytochemistry 2010, 71, 2058–2073. [Google Scholar] [CrossRef]
- Glivar, T.; Eržen, J.; Kreft, S.; Zagožen, M.; Čerenak, A.; Čeh, B.; Tavčar Benković, E. Cannabinoid Content in Industrial Hemp (Cannabis sativa L.) Varieties Grown in Slovenia. Ind. Crops Prod. 2020, 145, 112082. [Google Scholar] [CrossRef]
- Aizpurua-Olaizola, O.; Soydaner, U.; Öztürk, E.; Schibano, D.; Simsir, Y.; Navarro, P.; Etxebarria, N.; Usobiaga, A. Evolution of the Cannabinoid and Terpene Content during the Growth of Cannabis sativa Plants from Different Chemotypes. J. Nat. Prod. 2016, 79, 324–331. [Google Scholar] [CrossRef]
- Gorelick, J.; Bernstein, N. Chemical and Physical Elicitation for Enhanced Cannabinoid Production in Cannabis. In Cannabis sativa L.—Botany and Biotechnology; Springer International Publishing: Cham, Switzerland, 2017; pp. 439–456. ISBN 9783319545646. [Google Scholar]
- Amaducci, S.; Scordia, D.; Liu, F.H.; Zhang, Q.; Guo, H.; Testa, G.; Cosentino, S.L. Key Cultivation Techniques for Hemp in Europe and China. Ind. Crops Prod. 2015, 68, 2–16. [Google Scholar] [CrossRef]
- Van Der Werf, H.; Mathussen, E.W.J.M.; Haverkort, A. The Potential of Hemp (Cannabis sativa L.) for Sustainable Fibre Production: A Crop Physiological Appraisal. Ann. Appl. Biol. 1996, 129, 109–123. [Google Scholar] [CrossRef]
- Calzolari, D.; Magagnini, G.; Lucini, L.; Grassi, G.; Appendino, G.B.; Amaducci, S. High Added-Value Compounds from Cannabis Threshing Residues. Ind. Crops Prod. 2017, 108, 558–563. [Google Scholar] [CrossRef]
- Clarke, R.C.; Merlin, M.D. Letter to the Editor: Small, Ernest. 2015. Evolution and Classification of Cannabis sativa (Marijuana, Hemp) in Relation to Human Utilization. Botanical Review 81(3): 189–294. Bot. Rev. 2015, 81, 295–305. [Google Scholar] [CrossRef]
- Hammami, N.; Privé, J.-P.; Joly, D.L.; Moreau, G. Associations between Cannabinoids and Growth Stages of Twelve Industrial Hemp Cultivars Grown Outdoors in Atlantic Canada. Ind. Crops Prod. 2021, 172, 113997. [Google Scholar] [CrossRef]
- de Backer, B.; Maebe, K.; Verstraete, A.G.; Charlier, C. Evolution of the Content of THC and Other Major Cannabinoids in Drug-Type Cannabis Cuttings and Seedlings During Growth of Plants. J. Forensic Sci. 2012, 57, 918–922. [Google Scholar] [CrossRef]
- García-Tejero, I.F.; Hernández, A.; Ferreiro-Vera, C.; Zuazo, V.H.D.; García, J.H.; Sánchez-Carnerero, C.; Casano, S. Yield of New Hemp Varieties for Medical Purposes under Semi-Arid Mediterranean Environment Conditions. Comun. Sci. 2020, 11, e3264. [Google Scholar] [CrossRef]
- Toonen, M.; Ribot, S.; Thissen, J. Yield of Illicit Indoor Cannabis Cultivation in The Netherlands. J. Forensic Sci. 2006, 51, 1050–1054. [Google Scholar] [CrossRef]
- Vanhove, W.; van Damme, P.; Meert, N. Factors Determining Yield and Quality of Illicit Indoor Cannabis (Cannabis spp.) Production. Forensic Sci. Int. 2011, 212, 158–163. [Google Scholar] [CrossRef]
- Hawley, D.; Graham, T.; Stasiak, M.; Dixon, M. Improving Cannabis Bud Quality and Yield with Subcanopy Lighting. HortScience 2018, 53, 1593–1599. [Google Scholar] [CrossRef]
- Rocha, S.B.F. Potencial Brasileiro Para o Cultivo de Cannabis sativa L.; Para Uso Medicinal e Industrial: Viçosa, Brazil, 2019. [Google Scholar]
- Nemati, R.; Fortin, J.-P.; Craig, J.; Donald, S.; Fonteno, C.; Michel, J.-C.; Jackson, B.E. Growing Mediums for Medical Cannabis Production in North America. Agronomy 2021, 11, 1366. [Google Scholar] [CrossRef]
- de Prato, L.; Ansari, O.; Hardy, G.E.S.J.; Howieson, J.; O’Hara, G.; Ruthrof, K.X. The Cannabinoid Profile and Growth of Hemp (Cannabis sativa L.) Is Influenced by Tropical Daylengths and Temperatures, Genotype and Nitrogen Nutrition. Ind. Crops Prod. 2022, 178, 114605. [Google Scholar] [CrossRef]
- Caplan, D.; Dixon, M.; Zheng, Y. Optimal Rate of Organic Fertilizer during the Flowering Stage for Cannabis Grown in Two Coir-Based Substrates. HortScience 2017, 52, 1796–1803. [Google Scholar] [CrossRef]
- Green, G. The Cannabis Grow Bible, 2nd ed.; Green Candy Press: São Francisco, CA, USA, 2010. [Google Scholar]
- Rosenthal, E. Marijuana Grower’s Handbook; Quick American Publishing: Oakland, CA, USA, 2010; ISBN 0-932551-46-7. [Google Scholar]
- Bernstein, N.; Gorelick, J.; Koch, S. Interplay between Chemistry and Morphology in Medical Cannabis (Cannabis sativa L.). Ind. Crops Prod. 2019, 129, 185–194. [Google Scholar] [CrossRef]
- Crispim Massuela, D.; Hartung, J.; Munz, S.; Erpenbach, F.; Graeff-Hönninger, S. Impact of Harvest Time and Pruning Technique on Total CBD Concentration and Yield of Medicinal Cannabis. Plants 2022, 11, 140. [Google Scholar] [CrossRef] [PubMed]
- Danziger, N.; Bernstein, N. Shape Matters: Plant Architecture Affects Chemical Uniformity in Large-Size Medical Cannabis Plants. Plants 2021, 10, 1834. [Google Scholar] [CrossRef] [PubMed]
- Verma, N.; Shukla, S. Impact of Various Factors Responsible for Fluctuation in Plant Secondary Metabolites. J. Appl. Res. Med. Aromat. Plants 2015, 2, 105–113. [Google Scholar] [CrossRef]
- Folina, A.; Kakabouki, I.; Tourkochoriti, E.; Roussis, I.; Pateroulakis, H.; Bilalis, D. Evaluation of the Effect of Topping on Cannabidiol (CBD) Content in Two Industrial Hemp (Cannabis sativa L.) Cultivars. Bull. Univ. Agric. Sci. Vet. Med. Cluj-Napoca. Hortic. 2020, 77, 46. [Google Scholar] [CrossRef]
- Cervantes, J. Marijuana: Horticulture: The Indoor/Outdoor Medical Grower’s Bible, 1st ed.; Van Patten Publishing: Vancouver, WA, USA, 2007. [Google Scholar]
- Campbell, B.J.; Berrada, A.F.; Hudalla, C.; Amaducci, S.; McKay, J.K. Genotype × Environment Interactions of Industrial Hemp Cultivars Highlight Diverse Responses to Environmental Factors. Agrosyst. Geosci. Environ. 2019, 2, 180057. [Google Scholar] [CrossRef] [Green Version]
- Eichhorn Bilodeau, S.; Wu, B.S.; Rufyikiri, A.S.; MacPherson, S.; Lefsrud, M. An Update on Plant Photobiology and Implications for Cannabis Production. Front. Plant Sci. 2019, 10, 296. [Google Scholar] [CrossRef]
- Magagnini, G.; Grassi, G.; Kotiranta, S. The Effect of Light Spectrum on the Morphology and Cannabinoid Content of Cannabis sativa L. Med. Cannabis Cannabinoids 2018, 1, 19–27. [Google Scholar] [CrossRef]
- Danziger, N.; Bernstein, N. Light Matters: Effect of Light Spectra on Cannabinoid Profile and Plant Development of Medical Cannabis (Cannabis sativa L.). Ind. Crops Prod. 2021, 164, 113351. [Google Scholar] [CrossRef]
- Islam, M.J.; Ryu, B.R.; Azad, M.O.K.; Rahman, M.H.; Cheong, E.J.; Lim, J.-D.; Lim, Y.-S. Cannabinoids Accumulation in Hemp (Cannabis sativa L.) Plants under LED Light Spectra and Their Discrete Role as a Stress Marker. Biology 2021, 10, 710. [Google Scholar] [CrossRef]
- Lydon, J.; Teramura, A.H.; Coffman, C.B. UV-B Radiation Effects on Photosynthesis, Growth and Cannabinoid Production of Two Cannabis sativa Chemotypes. Photochem. Photobiol. 1987, 46, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Morrison, V.; Llewellyn, D.; Zheng, Y. Cannabis Inflorescence Yield and Cannabinoid Concentration Are Not Increased with Exposure to Short-Wavelength Ultraviolet-B Radiation. Front. Plant Sci. 2021, 12, 725078. [Google Scholar] [CrossRef] [PubMed]
- Llewellyn, D.; Golem, S.; Foley, E.; Dinka, S.; Maxwell, A.; Jones, P.; Zheng, Y. Cannabis Yield Increased Proportionally with Light Intensity, but Additional Ultraviolet Radiation Did Not Affect Yield or Cannabinoid Content. Anat. Morphol. 2021, 1–19. [Google Scholar] [CrossRef]
- Jenkins, M.W. Cannabis sativa L. Response to Narrow Bandwidth UV and the Combination of Blue and Red Light during the Final Stages of Flowering on Leaf Level Gas-Exchange Parameters, Secondary Metabolite Production, and Yield. Agric. Sci. 2021, 12, 1414–1432. [Google Scholar] [CrossRef]
- Rodriguez-Morrison, V.; Llewellyn, D.; Zheng, Y. Cannabis Yield, Potency, and Leaf Photosynthesis Respond Differently to Increasing Light Levels in an Indoor Environment. Front. Plant Sci. 2021, 12, 456. [Google Scholar] [CrossRef]
- Valle, J.R.; Vieira, J.E.; Aucélio, J.G.; Valio, I.F. Influence of Photoperiodism on Cannabinoid Content of Cannabis sativa L. Bull. Narc. 1978, 30, 67–68. [Google Scholar]
- Khajuria, M.; Rahul, V.P.; Vyas, D. Photochemical Efficiency Is Negatively Correlated with the Δ9- Tetrahydrocannabinol Content in Cannabis sativa L.: Effect of THC in Cannabis. Plant Physiol. Biochem. 2020, 151, 589–600. [Google Scholar] [CrossRef]
- Mahlberg, P.G.; Hemphill, J.K. Effect of Light Quality on Cannabinoid Content of Cannabis sativa L. (Cannabaceae). Bot. Gaz. 1983, 144, 43–48. [Google Scholar] [CrossRef]
- Eaves, J.; Eaves, S.; Morphy, C.; Murray, C. The Relationship between Light Intensity, Cannabis Yields, and Profitability. Agron. J. 2020, 112, 1466–1470. [Google Scholar] [CrossRef]
- Westmoreland, F.M.; Kusuma, P.; Bugbee, B. Cannabis Lighting: Decreasing Blue Photon Fraction Increases Yield but Efficacy Is More Important for Cost Effective Production of Cannabinoids. PLoS ONE 2021, 16, e0248988. [Google Scholar] [CrossRef]
- Namdar, D.; Charuvi, D.; Ajjampura, V.; Mazuz, M.; Ion, A.; Kamara, I.; Koltai, H. LED Lighting Affects the Composition and Biological Activity of Cannabis sativa Secondary Metabolites. Ind. Crops Prod. 2019, 132, 177–185. [Google Scholar] [CrossRef]
- Pattison, P.M.; Tsao, J.Y.; Brainard, G.C.; Bugbee, B. LEDs for Photons, Physiology and Food. Nature 2018, 563, 493–500. [Google Scholar] [CrossRef] [PubMed]
- Lefsrud, M.G.; Kopsell, D.A.; Sams, C.E. Irradiance from Distinct Wavelength Light-Emitting Diodes Affect Secondary Metabolites in Kale. HortScience 2008, 43, 2243–2244. [Google Scholar] [CrossRef] [Green Version]
- Wei, X.; Zhao, X.; Long, S.; Xiao, Q.; Guo, Y.; Qiu, C.; Qiu, H.; Wang, Y. Wavelengths of LED Light Affect the Growth and Cannabidiol Content in Cannabis sativa L. Ind. Crops Prod. 2021, 165, 113433. [Google Scholar] [CrossRef]
- Jenkins, M.W.; Livesay, C.B. Photosynthetic Performance and Potency of Cannabis sativa L. Grown under LED and HPS Illumination. Agric. Sci. 2021, 12, 293–304. [Google Scholar] [CrossRef]
- Pate, D.W. Possible Role of Ultraviolet Radiation in Evolution of Cannabis Chemotypes. Econ. Bot. 1983, 37, 396–405. [Google Scholar] [CrossRef]
- Chandra, S.; Lata, H.; Khan, I.A.; Elsohly, M.A. Photosynthetic Response of Cannabis sativa Photosynthetic Response of Cannabis sativa L. to Variations in Photosynthetic Photon Flux Densities, Temperature and CO2 Conditions. Physiol. Mol. Biol. Plants 2008, 14, 299–306. [Google Scholar] [CrossRef] [Green Version]
- Potter, D.J.; Duncombe, P. The Effect of Electrical Lighting Power and Irradiance on Indoor-Grown Cannabis Potency and Yield. J. Forensic Sci. 2012, 57, 618–622. [Google Scholar] [CrossRef]
- Chandra, S.; Lata, H.; Mehmedic, Z.; Khan, I.A.; ElSohly, M.A. Light Dependence of Photosynthesis and Water Vapor Exchange Characteristics in Different High Δ9-THC Yielding Varieties of Cannabis sativa L. J. Appl. Res. Med. Aromat. Plants 2015, 2, 39–47. [Google Scholar] [CrossRef]
- Bouchard, M. Towards a Realistic Method to Estimate Cannabis Production in Industrialized Countries. Contemp. Drug Probl. 2008, 35, 291–320. [Google Scholar] [CrossRef]
- Dang, M.; Arachchige, N.M.; Campbell, L.G. Optimizing Photoperiod Switch to Maximize Floral Biomass and Cannabinoid Yield in Cannabis sativa L.: A Meta-Analytic Quantile Regression Approach. Front. Plant Sci. 2022, 12, 797425. [Google Scholar] [CrossRef] [PubMed]
- Lenton, S.; Frank, V.A.; Barratt, M.J.; Potter, G.R.; Decorte, T. Growing Practices and the Use of Potentially Harmful Chemical Additives among a Sample of Small-Scale Cannabis Growers in Three Countries. Drug Alcohol Depend. 2018, 192, 250–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Backer, R.; Schwinghamer, T.; Rosenbaum, P.; McCarty, V.; Eichhorn Bilodeau, S.; Lyu, D.; Ahmed, M.B.; Robinson, G.; Lefsrud, M.; Wilkins, O.; et al. Closing the Yield Gap for Cannabis: A Meta-Analysis of Factors Determining Cannabis Yield. Front. Plant Sci. 2019, 10, 495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, R. Hemp as an Agricultural Commodity; Congressional Research Service: Washington, DC, USA, 2014.
- Vanhove, W.; Surmont, T.; van Damme, P.; de Ruyver, B. Yield and Turnover of Illicit Indoor Cannabis (Cannabis spp.) Plantations in Belgium. Forensic Sci. Int. 2012, 220, 265–270. [Google Scholar] [CrossRef]
- Benevenute, S.S.; Freeman, J.H.; Yang, R. How Do Pinching and Plant Density Affect Industrial Hemp Produced for Cannabinoids in Open Field Conditions? Agron. J. 2022, 114, 618–626. [Google Scholar] [CrossRef]
- García-Tejero, I.F.; Durán Zuazo, V.H.; Sánchez-Carnenero, C.; Hernández, A.; Ferreiro-Vera, C.; Casano, S. Seeking Suitable Agronomical Practices for Industrial Hemp (Cannabis sativa L.) Cultivation for Biomedical Applications. Ind. Crops Prod. 2019, 139, 111524. [Google Scholar] [CrossRef]
- Ingallina, C.; Sobolev, A.P.; Circi, S.; Spano, M.; Fraschetti, C.; Filippi, A.; di Sotto, A.; di Giacomo, S.; Mazzoccanti, G.; Gasparrini, F.; et al. Cannabis sativa L. Inflorescences from Monoecious Cultivars Grown in Central Italy: An Untargeted Chemical Characterization from Early Flowering to Ripening. Molecules 2020, 25, 1908. [Google Scholar] [CrossRef]
- Stack, G.M.; Toth, J.A.; Carlson, C.H.; Cala, A.R.; Marrero-González, M.I.; Wilk, R.L.; Gentner, D.R.; Crawford, J.L.; Philippe, G.; Rose, J.K.C.; et al. Season-long Characterization of High-cannabinoid Hemp (Cannabis sativa L.) Reveals Variation in Cannabinoid Accumulation, Flowering Time, and Disease Resistance. GCB Bioenergy 2021, 13, 546–561. [Google Scholar] [CrossRef]
- Richins, R.D.; Rodriguez-Uribe, L.; Lowe, K.; Ferral, R.; O’Connell, M.A. Accumulation of Bioactive Metabolites in Cultivated Medical Cannabis. PLoS ONE 2018, 13, e0201119. [Google Scholar] [CrossRef] [Green Version]
- Turner, J.C.; Mahlberg, P.G.; Lanyon, V.S.; Pleszczynska, J. A Temporal Study of Cannabinoid Composition in Continual Clones of Cannabis sativa L. (Cannabaceae). Bot. Gaz. 1985, 146, 32–38. [Google Scholar] [CrossRef] [Green Version]
- Latta, R.P.; Eaton, B.J. Seasonal Fluctuations in Cannabinoid Content of Kansas Marijuana. Econ. Bot. 1975, 29, 153–163. [Google Scholar] [CrossRef]
- Bernstein, N.; Gorelick, J.; Zerahia, R.; Koch, S. Impact of N, P, K, and Humic Acid Supplementation on the Chemical Profile of Medical Cannabis (Cannabis sativa L.). Front. Plant Sci. 2019, 10, 736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiponi, S.; Bernstein, N. The Highs and Lows of P Supply in Medical Cannabis: Effects on Cannabinoids, the Ionome, and Morpho-Physiology. Front. Plant Sci. 2021, 12, 657323. [Google Scholar] [CrossRef] [PubMed]
- Hemphill, J.K.; Turner, J.C.; Mahlberg, P.G. Cannabinoid Content of Individual Plant Organs from Different Geographical Strains of Cannabis sativa L. J. Nat. Prod. 1980, 43, 112–122. [Google Scholar] [CrossRef]
- Jin, D.; Dai, K.; Xie, Z.; Chen, J. Secondary Metabolites Profiled in Cannabis Inflorescences, Leaves, Stem Barks, and Roots for Medicinal Purposes. Sci. Rep. 2020, 10, 3309. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.C.; Hemphill, J.K.; Mahlberg, P.G. Interrelationships of Glandular Trichomes and Cannabinoid Content. II. Developing Vegetative Leaves of Cannabis sativa L. (Cannabaceae). Bull. Narc. 1981, 33, 63–71. [Google Scholar]
- Turner, J.C.; Hemphill, J.K.; Mahlberg, P.G. Cannabinoid Composition and Gland Distribution in Clones of Cannabis sativa L. (Cannabaceae). Bull. Narc. 1978, 30, 55–65. [Google Scholar]
- Yang, R.; Berthold, E.C.; McCurdy, C.R.; da Silva Benevenute, S.; Brym, Z.T.; Freeman, J.H. Development of Cannabinoids in Flowers of Industrial Hemp (Cannabis sativa L.): A Pilot Study. J. Agric. Food Chem. 2020, 68, 6058–6064. [Google Scholar] [CrossRef]
- Vogelmann, A.F.; Turner, J.C.; Mahlberg, P.G. Cannabinoid Composition in Seedlings Compared to Adult Plants of Cannabis sativa. J. Nat. Prod. 1988, 51, 1075–1079. [Google Scholar] [CrossRef]
- Pacifico, D.; Miselli, F.; Carboni, A.; Moschella, A.; Mandolino, G. Time Course of Cannabinoid Accumulation and Chemotype Development during the Growth of Cannabis sativa L. Euphytica 2008, 160, 231–240. [Google Scholar] [CrossRef]
- Fairbairn, J.W.; Liebmann, J.A. The Cannabinoid Content of Cannabis sativa L. Grown in England. J. Pharm. Pharmacol. 1974, 26, 413–419. [Google Scholar] [CrossRef] [PubMed]
- Namdar, D.; Mazuz, M.; Ion, A.; Koltai, H. Variation in the Compositions of Cannabinoid and Terpenoids in Cannabis sativa Derived from Inflorescence Position along the Stem and Extraction Methods. Ind. Crops Prod. 2018, 113, 376–382. [Google Scholar] [CrossRef]
- Saloner, A.; Bernstein, N. Nitrogen Supply Affects Cannabinoid and Terpenoid Profile in Medical Cannabis (Cannabis sativa L.). Ind. Crops Prod. 2021, 167, 113516. [Google Scholar] [CrossRef]
- Bauerle, W.L.; McCullough, C.; Iversen, M.; Hazlett, M. Leaf Age and Position Effects on Quantum Yield and Photosynthetic Capacity in Hemp Crowns. Plants 2020, 9, 271. [Google Scholar] [CrossRef] [Green Version]
- Sharma, G.K. Altitudinal Variation in Leaf Epidermal Patterns of Cannabis sativa. Bull. Torrey Bot. Club 1975, 102, 199–200. [Google Scholar] [CrossRef]
- Giupponi, L.; Leoni, V.; Pavlovic, R.; Giorgi, A. Influence of Altitude on Phytochemical Composition of Hemp Inflorescence: A Metabolomic Approach. Molecules 2020, 25, 1381. [Google Scholar] [CrossRef] [Green Version]
- Paris, M.; Boucher, F.; Cosson, L. The Constituents of Cannabis sativa Pollen. Econ. Bot. 1975, 29, 245–253. [Google Scholar] [CrossRef]
- Pate, D.W. Chemical Ecology of Cannabis. Int. Hemp Assoc. 1994, 2, 32–37. [Google Scholar]
- Bazzaz, F.A.; Dusek, D.; Seigler, D.S.; Haney, A.W. Photosynthesis and Cannabinoid Content of Temperate and Tropical Populations of Cannabis sativa; Pergamon Press: Oxford, UK, 1975; Volume 3. [Google Scholar]
- Chandra, S.; Lata, H.; Khan, I.A.; ElSohly, M.A. Temperature Response of Photosynthesis in Different Drug and Fiber Varieties of Cannabis sativa L. Physiol. Mol. Biol. Plants 2011, 17, 297–303. [Google Scholar] [CrossRef] [Green Version]
- Kirschbaum, M.U.F. Does Enhanced Photosynthesis Enhance Growth? Lessons Learned from CO2 Enrichment Studies. Plant Physiol. 2011, 155, 117–124. [Google Scholar] [CrossRef] [Green Version]
- Murari, G.; Puccini, A.M.; de Sanctis, R.; Lombardi, S. Influence of Environmental Conditions on Tetrahydrocannabinol (Delta (′9) TCH) in Different Cultivars on Cannabis sativa L. Fitoterapia 1983, 54, 195–201. [Google Scholar]
- Li, X.; Kang, S.; Li, F.; Zhang, X.; Huo, Z.; Ding, R.; Tong, L.; Du, T.; Li, S. Light Supplement and Carbon Dioxide Enrichment Affect Yield and Quality of Off-Season Pepper. Agron. J. 2017, 109, 2107–2118. [Google Scholar] [CrossRef]
- Saloner, A.; Bernstein, N. Response of Medical Cannabis (Cannabis sativa L.) to Nitrogen Supply under Long Photoperiod. Front. Plant Sci. 2020, 11, 1517. [Google Scholar] [CrossRef] [PubMed]
- Coffman, C.B.; Gentner, W.A. Responses of Greenhouse-grown Cannabis sativa L. to Nitrogen, Phosphorus, and Potassium 1. Agron. J. 1977, 69, 832–836. [Google Scholar] [CrossRef]
- Bócsa, I.; Máthé, P.; Hangyel, L. Effect of Nitrogen on Tetrahydrocannabinol (THC) Content in Hemp (Cannabis sativa L.) Leaves at Different Positions. J. Int. Hemp Assoc. 1997, 4, 80–81. [Google Scholar]
- de Prato, L.; Ansari, O.; Hardy, G.E.S.J.; Howieson, J.; O’Hara, G.; Ruthrof, K.X. Morpho-Physiology and Cannabinoid Concentrations of Hemp (Cannabis sativa L.) Are Affected by Potassium Fertilisers and Microbes under Tropical Conditions. Ind. Crops Prod. 2022, 182, 114907. [Google Scholar] [CrossRef]
- Caplan, D.; Dixon, M.; Zheng, Y. Optimal Rate of Organic Fertilizer during the Vegetative-Stage for Cannabis Grown in Two Coir-Based Substrates. HortScience 2017, 52, 1307–1312. [Google Scholar] [CrossRef] [Green Version]
- Saloner, A.; Bernstein, N. Nitrogen Source Matters: High NH4/NO3 Ratio Reduces Cannabinoids, Terpenoids, and Yield in Medical Cannabis. Front. Plant Sci. 2022, 13, 830224. [Google Scholar] [CrossRef]
- Shiponi, S.; Bernstein, N. Response of Medical Cannabis (Cannabis sativa L.) Genotypes to P Supply under Long Photoperiod: Functional Phenotyping and the Ionome. Ind. Crops Prod. 2021, 161, 113154. [Google Scholar] [CrossRef]
- Saloner, A.; Bernstein, N. Effect of Potassium (K) Supply on Cannabinoids, Terpenoids and Plant Function in Medical Cannabis. Agronomy 2022, 12, 1242. [Google Scholar] [CrossRef]
- Yep, B.; Gale, N.V.; Zheng, Y. Comparing Hydroponic and Aquaponic Rootzones on the Growth of Two Drug-Type Cannabis sativa L. Cultivars during the Flowering Stage. Ind. Crops Prod. 2020, 157, 112881. [Google Scholar] [CrossRef]
- Yep, B.; Gale, N.V.; Zheng, Y. Aquaponic and Hydroponic Solutions Modulate NaCl-Induced Stress in Drug-Type Cannabis sativa L. Front. Plant Sci. 2020, 11, 1169. [Google Scholar] [CrossRef] [PubMed]
- Kakabouki, I.; Kousta, A.; Folina, A.; Karydogianni, S.; Zisi, C.; Kouneli, V.; Papastylianou, P. Effect of Fertilization with Urea and Inhibitors on Growth, Yield and Cbd Concentration of Hemp (Cannabis sativa L.). Sustainability 2021, 13, 2157. [Google Scholar] [CrossRef]
- Malík, M.; Velechovský, J.; Praus, L.; Janatová, A.; Kahánková, Z.; Klouček, P.; Tlustoš, P. Amino Acid Supplementation as a Biostimulant in Medical Cannabis (Cannabis sativa L.) Plant Nutrition. Front. Plant Sci. 2022, 13, 868350. [Google Scholar] [CrossRef]
- Burgel, L.; Hartung, J.; Graeff-Hönninger, S. Impact of Different Growing Substrates on Growth, Yield and Cannabinoid Content of Two Cannabis sativa L. Genotypes in a Pot Culture. Horticulturae 2020, 6, 62. [Google Scholar] [CrossRef]
- Carah, J.K.; Howard, J.K.; Thompson, S.E.; Short Gianotti, A.G.; Bauer, S.D.; Carlson, S.M.; Dralle, D.N.; Gabriel, M.W.; Hulette, L.L.; Johnson, B.J.; et al. High Time for Conservation: Adding the Environment to the Debate on Marijuana Liberalization. BioScience 2015, 65, 822–829. [Google Scholar] [CrossRef] [Green Version]
- Bauer, S.; Olson, J.; Cockrill, A.; van Hattem, M.; Miller, L.; Tauzer, M.; Leppig, G. Impacts of Surface Water Diversions for Marijuana Cultivation on Aquatic Habitat in Four Northwestern California Watersheds. PLoS ONE 2015, 10, e0120016. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Z.; Fiddes, K.; Yang, L. A Narrative Review on Environmental Impacts of Cannabis Cultivation. J. Cannabis Res. 2021, 3, 35. [Google Scholar] [CrossRef]
- Yadav, B.; Jogawat, A.; Rahman, M.S.; Narayan, O.P. Secondary Metabolites in the Drought Stress Tolerance of Crop Plants: A Review. Gene Rep. 2021, 23, 101040. [Google Scholar] [CrossRef]
- Caplan, D.; Dixon, M.; Zheng, Y. Increasing Inflorescence Dry Weight and Cannabinoid Content in Medical Cannabis Using Controlled Drought Stress. HortScience 2019, 54, 964–969. [Google Scholar] [CrossRef] [Green Version]
- Sheldon, K.; Shekoofa, A.; Walker, E.; Kelly, H. Physiological Screening for Drought-Tolerance Traits among Hemp (Cannabis sativa L.) Cultivars in Controlled Environments and in Field. J. Crop Improv. 2021, 35, 816–831. [Google Scholar] [CrossRef]
- García-Tejero, I.F.; Zuazo, V.H.D.; Pérez-Álvarez, R.; Hernández, A.; Cassano, S.; Moron, M.; Muriel, J.L. Impact of Plant Density and Irrigation on Yield of Hemp (Cannabis sativa L.) in a Mediterranean Semi-Arid Environment. J. Agric. Sci. Technol. 2014, 16, 887–895. [Google Scholar]
- Sikora, V.; Berenji, J.; Latkovic, D. Influence of Agroclimatic Conditions on Content of Main Cannabinoids in Industrial Hemp (Cannabis sativa L.). Genetika 2011, 43, 449–456. [Google Scholar] [CrossRef]
- Toth, J.A.; Smart, L.B.; Smart, C.D.; Stack, G.M.; Carlson, C.H.; Philippe, G.; Rose, J.K.C. Limited Effect of Environmental Stress on Cannabinoid Profiles in High-Cannabidiol Hemp (Cannabis sativa L.). GCB Bioenergy 2021, 13, 1666–1674. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Reference Number | Objective | Advantage | Disadvantages |
|---|---|---|---|
| [82] | Evaluate daily doses of 0, 6.7 and 13.4 kJ m−2 UV-B radiation | THC content increased linearly with UV-B dose radiation | Not reported |
| [83] | Evaluate supplemental UV radiation ranging from 0.01 to 0.8 μmol m−2 s−1. | Not reported | Decreased inflorescence yield with increasing UV exposure level; no induced enhancements to THC, CBD, and CBG |
| [84] | UVA+UVB radiation and LI ranging from 350 to 1400 μmol m−2 s−1 | Inflorescence yield increased linearly from 19.4 to 57.4 g plant−1 as LI increased from 350 to 1400 μmol m−2 s−1. | No effect in phytocannabinoid concentration and inflorescence yield with supplemental UV. UV radiation decreased harvest index. LI had no effect on cannabinoid content |
| [85] | Evaluate supplemental UV radiation | Increased THC concentration | Decreased inflorescence yield |
| [80] | Evaluate LEDs and HPS lamps | LED lamps (with highest proportion of blue light) provided high flower productivity and increased CBGA accumulation (up to 400%) | Decreased concentrations of other phytocannabinoids (such as THCA, CDBA or CBCA) by up to 40% under LED lamps. |
| [79] | Evaluate LEDs and HPS lamps | HPS lamps induced higher flower yield. LEDs provided highest CBG concentration (increasing by about 200% with a higher incidence of blue light). | HPS lamps induced lower THC and CBD production (about 30%) than when employing LED lamps. |
| [86] | Evaluate LI ranging from 120 to 1800 μmol·m−2·s−1. | Inflorescence yields increased linearly from 116 to 519 g m−2 (i.e., 4.5 times higher the apical inflorescence density increased linearly as the LI increased | No effects on the production of any of the phytocannabinoids measured |
| [87] | Evaluate the influence of photoperiod duration (10 and 12 h) | An increase of 2 h a day of the photoperiod doubled the average amount of THC. | An increase of 2 h a day of the photoperiod decreased cannabichromene content. |
| Reference Number | Objective | Results |
|---|---|---|
| [53] | Evaluate the evolution of major phytocannabinoids during cannabis growth | Maximum concentrations of THCA and CBDA in flowers were attained at the end of flowering, during senescence, but varied according to chemotype. |
| [72] | Investigate the impact of harvest time on phytocannabinoid yield. | Dry weight of inflorescence plant−1 increased from 7.7 g at fifth week of flowering to a maximum of 25.1 g at eleventh week of flowering; the CBDA yield increased from 415 mg plant−1 at the fifth week of flowering, reaching a maximum (an average of 1300·mg plant−1) at the ninth and eleventh weeks of flowering |
| [71] | Evaluate the differences in the concentration of phytocannabinoids between the apical and basal parts of the plant | THC, CBD, CBG, CBC, and THCV showed a significant increase in the parts of the plant close to the apex; only CBT had an inverse correlation with plant height (highest concentration was in the lower organs); the levels of CBC and CBG were similar or higher in the inflorescence leaves than in flowers. |
| [114] | Evaluate the potential of nutritional supplementations (humic acids and inorganic N, P, and K) to affect the cannabinoid profile throughout the plant. | Higher concentrations of THC, CBD, CBG, CBC, and THCV in the flowers closest to the apex, while higher CBT and CBN concentrations were found in the flowers located lower; nutritional supplements influenced cannabinoid profile and may reduce cannabinoid variability throughout the plant, |
| [115] | Evaluate the hypothesis that P uptake, distribution, and availability in the plant affect the biosynthesis of phytocannabinoids. | The apical flowers showed higher concentrations of CBDA and THCA under low P availability (5 mg L−1); under high P availability (90 mg L−1), the concentrations of these compounds did not differ in these two locations. |
| [61] | Evaluate the differences in the concentration of phytocannabinoids in different plant organ positions in the plant. | Variations in phytocannabinoid biosynthesis according to the position of the organs of the plants so that the upper and median parts showed higher phytocannabinoid and flower yields when compared to the basal plant parts, probably due to higher flower yield. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trancoso, I.; de Souza, G.A.R.; dos Santos, P.R.; dos Santos, K.D.; de Miranda, R.M.d.S.N.; da Silva, A.L.P.M.; Santos, D.Z.; García-Tejero, I.F.; Campostrini, E. Cannabis sativa L.: Crop Management and Abiotic Factors That Affect Phytocannabinoid Production. Agronomy 2022, 12, 1492. https://doi.org/10.3390/agronomy12071492
Trancoso I, de Souza GAR, dos Santos PR, dos Santos KD, de Miranda RMdSN, da Silva ALPM, Santos DZ, García-Tejero IF, Campostrini E. Cannabis sativa L.: Crop Management and Abiotic Factors That Affect Phytocannabinoid Production. Agronomy. 2022; 12(7):1492. https://doi.org/10.3390/agronomy12071492
Chicago/Turabian StyleTrancoso, Ingrid, Guilherme A. R. de Souza, Paulo Ricardo dos Santos, Késia Dias dos Santos, Rosana Maria dos Santos Nani de Miranda, Amanda Lúcia Pereira Machado da Silva, Dennys Zsolt Santos, Ivan F. García-Tejero, and Eliemar Campostrini. 2022. "Cannabis sativa L.: Crop Management and Abiotic Factors That Affect Phytocannabinoid Production" Agronomy 12, no. 7: 1492. https://doi.org/10.3390/agronomy12071492