
The Role of Salicylic Acid in Mitigating the Adverse Effects of Chilling Stress on “Seddik” Mango Transplants



1
Pomology Department, Faculty of Agriculture, Cairo University, 4, El-Gamaa St., Giza 12613, Egypt
2
Central Laboratory for Agricultural Climate (CLAC), Agriculture Research Center (ARC), 6, Michael Bakhom St., Dokki, Giza 12411, Egypt
*
Author to whom correspondence should be addressed.
Agronomy 2022, 12(6), 1369; https://doi.org/10.3390/agronomy12061369
Submission received: 25 April 2022
/
Revised: 29 May 2022
/
Accepted: 30 May 2022
/
Published: 6 June 2022
(This article belongs to the Special Issue Developing Temperature-Resilient Plants: Responses and Mitigation Strategies)
Abstract
:Salicylic acid (SA) was sprayed on “Seddik” mango transplants at concentrations of 0, 0.5, 1, and 1.5 mM. Then, the mango transplants were subjected to 72 h of chilling stress at 4 ± 1 °C, followed by a six-day recovery under greenhouse conditions. Untreated transplants exposed to chilling stress represented the positive control, while those not exposed were the negative control. SA-pretreated mango transplants were compared to the positive and negative controls, evaluating physiological and biochemical changes. The SA concentration of 1.5 mM L−1 was the most efficient in mitigating chilling injury (CI) in mango transplants by maintaining the integrity of the leaves’ cell membrane and minimizing electrolyte leakage (EL), specifically after six days of recovery. SA increased photosynthetic pigment content, total sugar content, and 2, 2-diphenyl-1-picrylhydrazyl (DPPH) radical scavenging activity and decreased proline and total phenolic content in the “Seddik” mango transplants’ leaves. After exposure to chilling stress, the antioxidant enzymes’ internal activities in SA-pretreated chilled mango transplants improved, especially on the sixth day of recovery, compared to the negative control; the transplants nearly attained normal growth levels. Thus, SA can protect plants against the adverse effects of chilling stress.
Keywords:
antioxidant enzymes; chilling stress; climate; Mangifera indica; salicylic acid; Seddik; transplant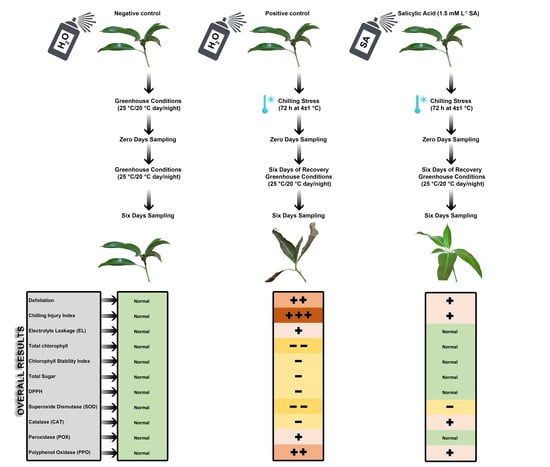
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
Mango (Mangifera indica L.), a tropical tree, is highly sensitive to low temperatures and needs protection against chilling damage. Mango grows very well in a warm, frost-free environment with a specified winter dry season. Mango’s optimum temperature ranges from 24 to 26.7 °C, with a minimum temperature of 10–12 °C, and chilling injury (CI) symptoms occur below these temperatures [1]. Currently, mango is experiencing CI during winter in Egypt, which affects its normal growth and development. According to the Central Laboratory for Agricultural Climate (CLAC) data, the minimum temperatures during the winter season have fluctuated over the last decade. This has caused cold symptoms in plants, specifically in tropical and subtropical fruit trees. Several cytological, biochemical, physiological, and molecular activities are altered due to chilling stress. They include photosynthesis, plasma membrane permeability, water status, osmotic balance, antioxidant activities, and other processes [2]. Many plant species have accrued SA when exposed to chilling stress. Salicylic acid (SA), an endogenous plant hormone, plays a vital role in growth, photosynthetic activity, and pigment content and has crucial physiological and biological roles in normal and stressed plants’ metabolism [3,4,5,6]. SA application improves CI tolerance in many fruit tree species, including apple [7], apricot [8], banana [9], citrus [10], guava [11], kiwifruit [12], loquat [13], mango [14], peach [15], cactus pear [16], plum [17], and pomegranate [18,19]. The vast majority of SA applications has the goal to alleviate CI to fruits during cold storage, and SA use to ease CI on fruit trees is very limited. Therefore, this study aimed to investigate SA role in mitigating the effects of chilling stress on Egyptian “Seddik” mango cultivar transplants.
2. Materials and Methods
This study was conducted during the 2021/2022 growing season at the Department of Pomology, Faculty of Agriculture, Cairo University, Egypt. All chemicals used in this study were reagent-grade and purchased from many worldwide suppliers (Bio Basic Inc., Markham, ON, Canada; Caisson Laboratories Inc., North Logan, UT, USA; Chem-Lab, Zedelgem, Belgium; Loba Chemie Pvt. Ltd., Maharashtra, India; Merck Ltd., Darmstadt, Germany; SDFCL, Chennai, India, and Sigma-Aldrich, Taufkirchen, Germany). Unless otherwise specified, all solutions were prepared in distilled water.
2.1. Greenhouse, Chilling Chamber Preparation, and Plant Materials
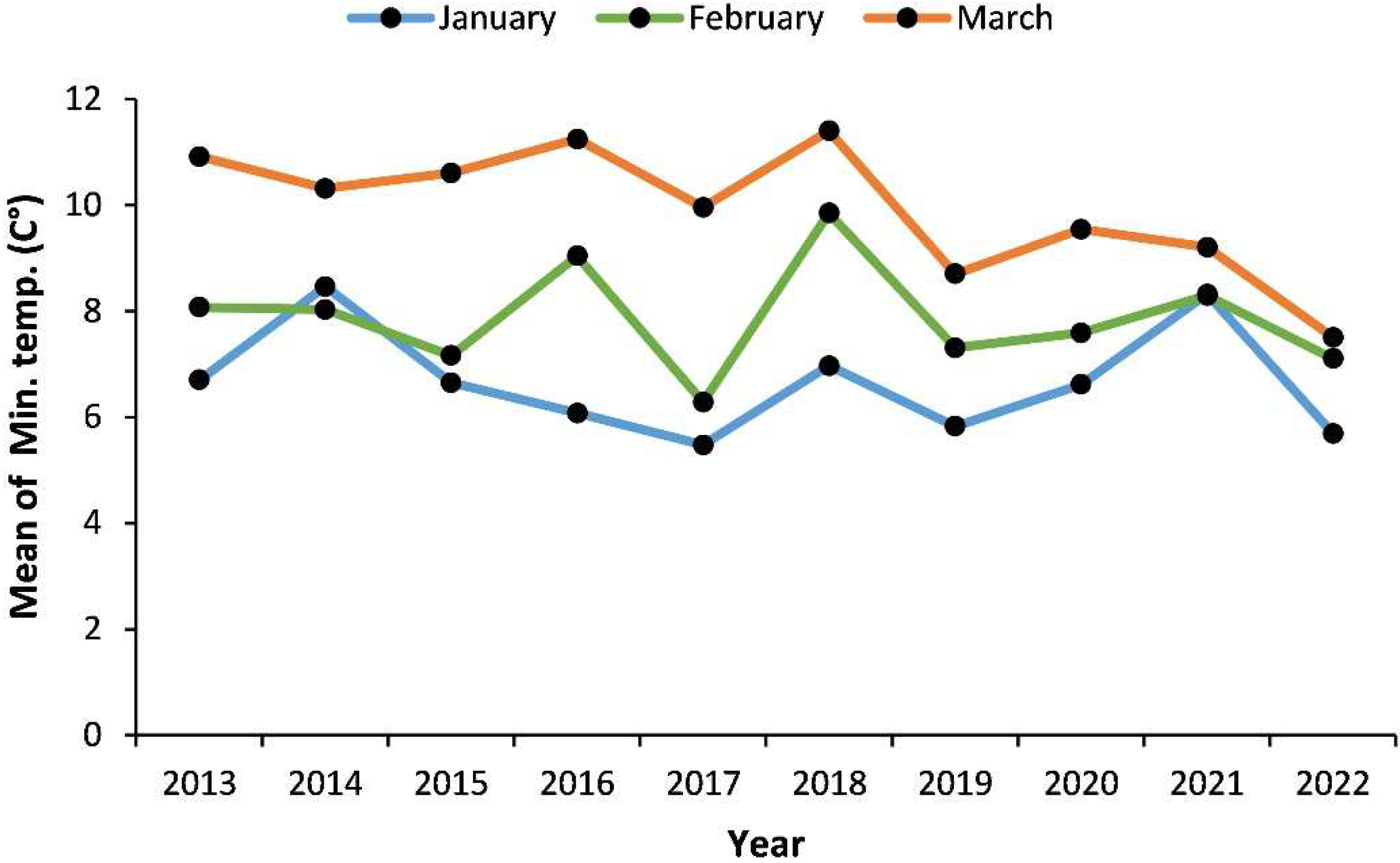
One-year-old mango (Mangifera indica L., cv. “Seddik”) transplants grafted on “Sukkary” rootstock were used. The selected plants had uniform sizes, received the recommended water amounts to prevent the development of water deficit, were supplemented with macro- and microelements, and were treated with suitable pesticides to prevent other biotic stresses. A greenhouse was prepared for mango transplants’ growth and recovery under normal growth conditions (25 °C/20 °C, day/night) before and after exposure to chilling. A chilling room was prepared to expose the mango transplants to chilling stress at 4 ± 1 °C. This temperature was determined based on the last decade’s data (Figure 1) provided by the Central Laboratory for Agricultural Climate (CLAC).
2.2. Exogenous Salicylic Acid Treatments and Chilling Stress Induction
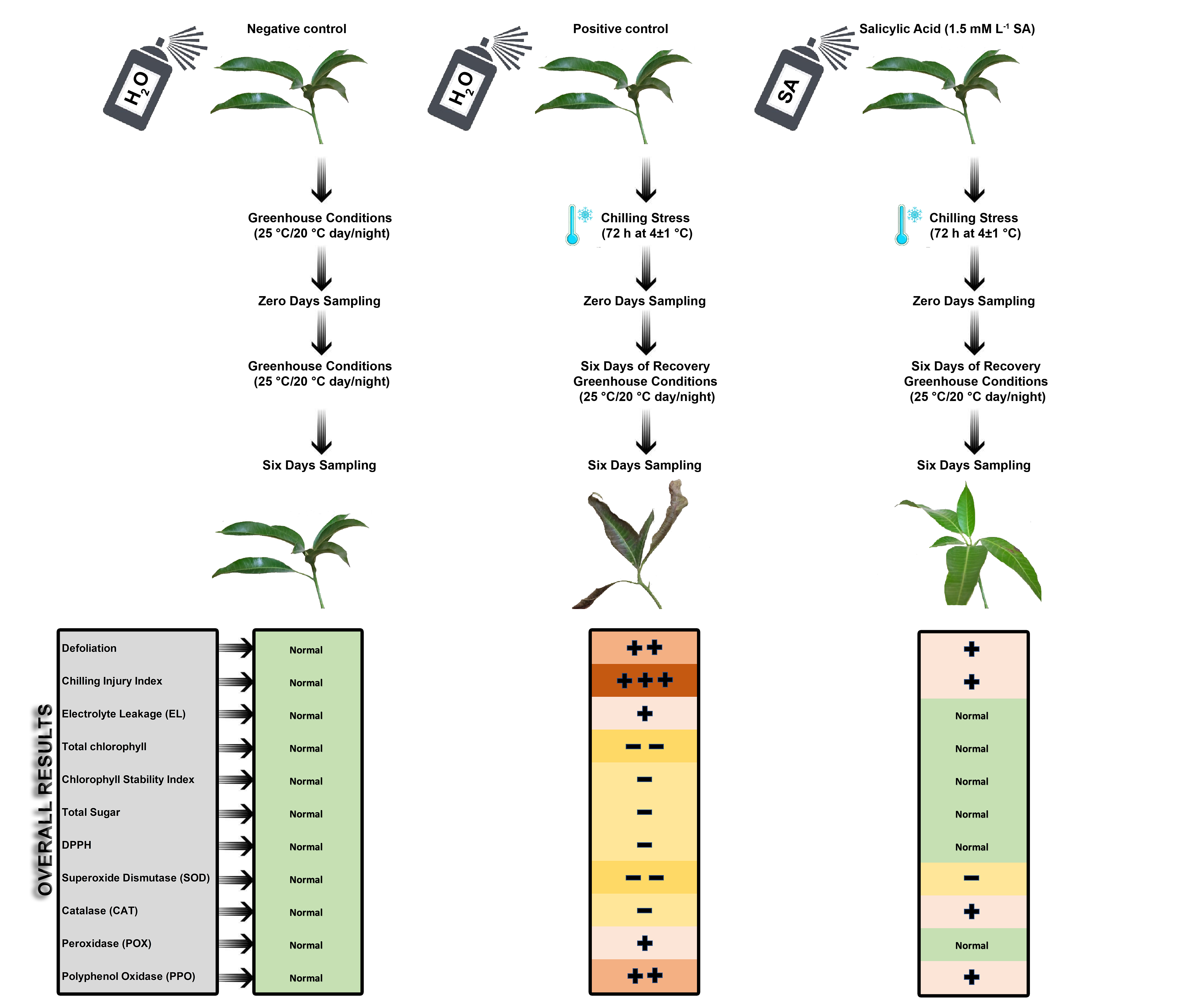
Salicylic acid (SA) was sprayed 48 and 24 h before the chilling treatment for 3 d at different concentrations (0, 0.5, 1, and 1.5 mM) on uniform juvenile mango transplants of “Seddik” cultivar. SA was dissolved in a NaOH solution (0.002 N); the pH was adjusted to 6.8 before the treatment. Then, Tween 80 was added at 0.1% as a surfactant. Untreated (treated with tap water) transplants exposed to chilling represented the positive control, while untreated transplants not exposed to chilling and kept under greenhouse conditions (25 °C/20 °C, day/night) throughout the entire period were the negative control.
For chilling exposure, the transplants were transferred to the Agriculture Development Systems (ADS) project’s growth chamber, Faculty of Agriculture, Cairo University, at 4 ± 1 °C for 72 consecutive hours, followed by a period of six days of recovery under greenhouse conditions (25 °C/20 °C, day/night).
2.3. Measurements
Leaf samples were collected from the third to fifth leaf from the top of the transplant at 0 and 6 days after the culmination of the period of chilling exposure at 4 ± 1 °C for 72 consecutive hours. All treatments were assessed by the following measurements:
2.3.1. Defoliation
Defoliation percentage was estimated visually as the mean number of fallen leaves from each transplant with and without treatment after three weeks of chilling and compared to the initial mean number of leaves in each transplant.
2.3.2. Chilling Injury Index
Chilling injury (CI) was visually assessed after three weeks of exposure to chilling by employing the following scale: normal, no visible symptoms; trace, small necrotic areas on leaves but without growth restrictions (less than 5% of necrotic leaf area); slight, small necrotic areas on leaves (less than 15% of necrotic leaf area); moderate, well-defined necrotic areas on leaves (less than 30% of necrotic leaf area); and severe, extensive necrotic areas with severe growth restrictions (more than 50% of necrotic leaf area, plant still alive). By assigning a score of 1, 2, 3, 4, or 5, respectively, to each group, the average injury for each treatment was calculated [20].
2.3.3. Chlorophyll Content
Fresh leaf samples (0.25 g) were randomly chosen from three plants per replicate and homogenized in 20 mL of acetone (80% v/v). The absorbance was measured using a spectrophotometer (JENWAY 6300, Staffordshire, UK) at 663 and 646 nm, and the chlorophyll content was determined using the following equations [21]:
Chl a (mg g−1 fw) = 12.21 × A663 − 2.81 × A646
Chl b (mg g−1 fw) = 20.13 × A646 − 5.03 × A663
Total chlorophyll content (mg g−1 fw) = Chl a + Chl b
2.3.4. Chlorophyll Stability Index (CSI)
The CSI was determined by heating fresh leaf samples (0.25 g) in 20 mL of distilled water at 56 °C in a water bath for 30 min [22]. Normal and heated leaf samples were homogenized in 80% acetone, then the total chlorophylls content was computed as above; the CSI was determined by using the following formula:
CSI (%) = (Total chlorophyll without heating − Total chlorophyll after heating) × 100
2.3.5. Electrolyte Leakage (EL) and Membrane Stability Index (MSI)
Electrolyte leakage was determined to evaluate membrane permeability, following the procedure of Guo et al. [23] with some modifications. Ten leaf discs of randomly selected plants per replicate were taken from the youngest fully expanded leaf. Then, the discs were placed in a 50 mL falcon tube and washed three times with distilled water to remove surface contamination. Next, the discs were placed in a 50 mL falcon tube containing 20 mL of deionized water (Aquinity2 P10, MembraPure GmbH, Hennigsdorf, Germany) and incubated, at room temperature, for 24 h. The bathing solution’s (EC1) electrical conductivity (EC) was read after incubation using an electrical conductivity meter (BALRAMA, Digital EC Meter, New Delhi, India). Afterward, the same samples were placed into a boiling water bath for 20 min, and a second reading (EC2) was carried out after cooling the solution to room temperature. The EL was expressed as a percent value using the following formula:
EL (%) = (EC1/EC2) × 100
The membrane stability index (MSI) was computed based on electrolyte leakage data and expressed as a percent value using the following formula:
MSI (%) = [1 − (EC1/EC2)] × 100
2.3.6. Total Sugar Content
The total sugar content was determined utilizing the phenol–sulfuric acid method, according to Dubois et al. [24]. To this aim, 0.5 g of fresh leaf was homogenized in 20 mL of 80% ethanol (v/v). One mL ethanolic solution was mixed with 1 mL of 5% phenol dissolved in water (v/v), followed by the addition of 5 mL of concentrated sulfuric acid. The absorbance was read at 490 nm by a spectrophotometer (JENWAY 6300, Staffordshire, UK). A standard curve was generated, employing a pure glucose solution, and the total sugar content was expressed in mg glucose equivalent g−1 of fresh weight.
2.3.7. Total Phenolic Content
A fresh leaf sample (0.5 g) was extracted with 20 mL of 80% (v/v) methanol and then used for determining the total phenolic content. Total phenols were determined with a spectrophotometer, employing the modified Folin–Ciocalteu colorimetric method [25]. The methanolic extract (1 mL) was diluted 1:10, then mixed with 1 mL of Folin–Ciocalteu reagent in a test tube and allowed to stand for 6 min, followed by the addition of 5 mL of 1M Na2CO3 (w/v). Then, 3 mL of distilled water was added. The samples were incubated for 90 min at room temperature in the dark, and the absorbance at 760 nm was measured using the JENWAY 6300 spectrophotometer, Staffordshire, UK. The results were expressed as mg gallic acid (GAE) g−1 fw.
2.3.8. Proline Content
Proline content was determined using the Bates’s method [26]. Leaf tissue (0.5 g) was homogenized in 10 mL of 3% aqueous sulfosalicylic acid for 10 min, followed by filtration. Two milliliters of the filtrate were mixed with 2 mL of acid ninhydrin and 2 mL of glacial acetic acid, then placed in a boiling water bath for 1 h at 90 °C, and the reaction was completed in an ice bath. The developed color was extracted in 4 mL of toluene, and the intensity of the reaction mixture was determined spectrophotometrically (JENWAY 6300, Staffordshire, UK) at the wavelength of 520 nm. Proline concentration was determined from a standard curve and calculated on a fresh weight basis as follows: µmoles proline/g of fresh weight = [(µg proline/mL × mL toluene)/115.5 µg/µmole]/[(g sample)/5].
2.3.9. DPPH Free Radical Scavenging Assay
The antioxidant activity was assessed by the 1,1-diphenyl-2-picrylhydrazyl (DPPH) free radical scavenging method [27,28]. We added 2.4 mL of 0.1 mM DPPH to 1.6 mL of methanolic leaf extract, vortexed the mixture, and incubated it at room temperature in the dark. The absorbance of the samples was measured after 30 min at 517 nm using a spectrophotometer (JENWAY 6300, Staffordshire, UK). The percentage of DPPH scavenging activity was calculated as % inhibition of DPPH = (A517 control –A517 sample/A517 control) × 100.
2.3.10. Extraction and Determination of Antioxidant Enzymes Activity
We used about 0.5 g of fresh young mango leaves to determine the concentration of superoxide dismutase (SOD), catalase (CAT), peroxidase (POX), and polyphenol oxidase (PPO) enzymes. Crude enzyme extracts were prepared using ground fresh leaves samples in liquid nitrogen that were homogenized in 10 mL of phosphate buffer pH 6.8 (0.1 M) and then centrifuged at 2 °C for 20 min at 20,000 rpm in a refrigerated centrifuge. The clear supernatant was the crude enzyme extract [29].
- Superoxide Dismutase (SOD) Activity
SOD (EC 1.15.1.1) activity was determined by measuring the inhibition of the auto-oxidation of pyrogallol by employing a method described by Marklund and Marklund [30]. We used a 10 mL reaction mixture consisting of 3.6 mL of distilled water, 0.1 mL of enzyme extract, 5.5 mL of 50 mM phosphate buffer (pH 7.8), and 0.8 mL of 3 mM pyrogallol (dissolved in 10 mM HCl). The pyrogallol reduction rate was measured at 325 nm with a UV–VIS spectrophotometer (PD 303, Apel Co., Ltd., Saitama, Japan). One unit of enzyme activity is the amount of the enzyme leading to 50% inhibition of the auto-oxidation rate of pyrogallol at 25 °C [31].
- Catalase (CAT) Activity
Catalase (E.C.1.11.1.6) activity was assayed based on Chen et al. [32]. The reaction mixture with a final volume of 10 mL, containing 40 µL of enzyme extract, was added to 9.96 mL of H2O2 phosphate buffer pH 7.0 (0.16 mL of 30% H2O2 to 100 mL of 50 mM phosphate buffer). Catalase activity was measured as the change in H2O2 absorbance in 60 s against a buffer blank at 250 nm using a UV–VIS spectrophotometer (PD 303, Apel Co., Ltd., Saitama, Japan). The blank sample was made by using buffer instead of the enzyme extract. One unit of enzyme activity is the amount of the enzyme reducing 50% of H2O2 in 60 s at 25 °C [31].
- Peroxidase (POX) Activity
Peroxidase (EC 1.11.1.7) activity was assayed in a reaction mixture containing 5.8 mL of 50 mM phosphate buffer pH 7.0, 0.2 mL of the enzyme extract, and 2 mL of 20 mM H2O2. The change in optical density was determined spectrophotometrically (PD 303, Apel Co., Ltd., Saitama, Japan) within 60 s at 470 nm at 25 °C after adding 2 mL of 20 mM pyrogallol [33]. One unit of enzyme activity is the amount of the enzyme catalyzing one micromole of H2O2 per minute at 25 °C [31].
- Polyphenol Oxidase (PPO) Activity
Polyphenol oxidase (EC 1.10.3.1) activity was determined using 125 µmol of phosphate buffer (pH 6.8), 100 µmol of pyrogallols, and 2 mL of enzyme extract. After incubating the mixture for 5 min at 25 °C, the reaction was stopped by adding 1 mL of 5% H2SO4. The developed color was read spectrophotometrically at 430 nm (PD 303, Apel Co., Ltd., Saitama, Japan) [34].
2.4. Statistical Analysis
The experiment used a completely randomized design (CRD) with three replicates; each replicate contained four mango transplants. The statistical analysis was performed using the R software, version 4.0.5, R Core Team, Vienna, Austria [35]. The main treatment effects at each sampling time were analyzed, and the means were compared by Duncan’s multiple range tests [36] at a significance level of 0.05. Pearson’s correlation coefficient was also calculated to specify associations between any measured parameters at each sampling time in response to chilling and SA application.
3. Results and Discussion
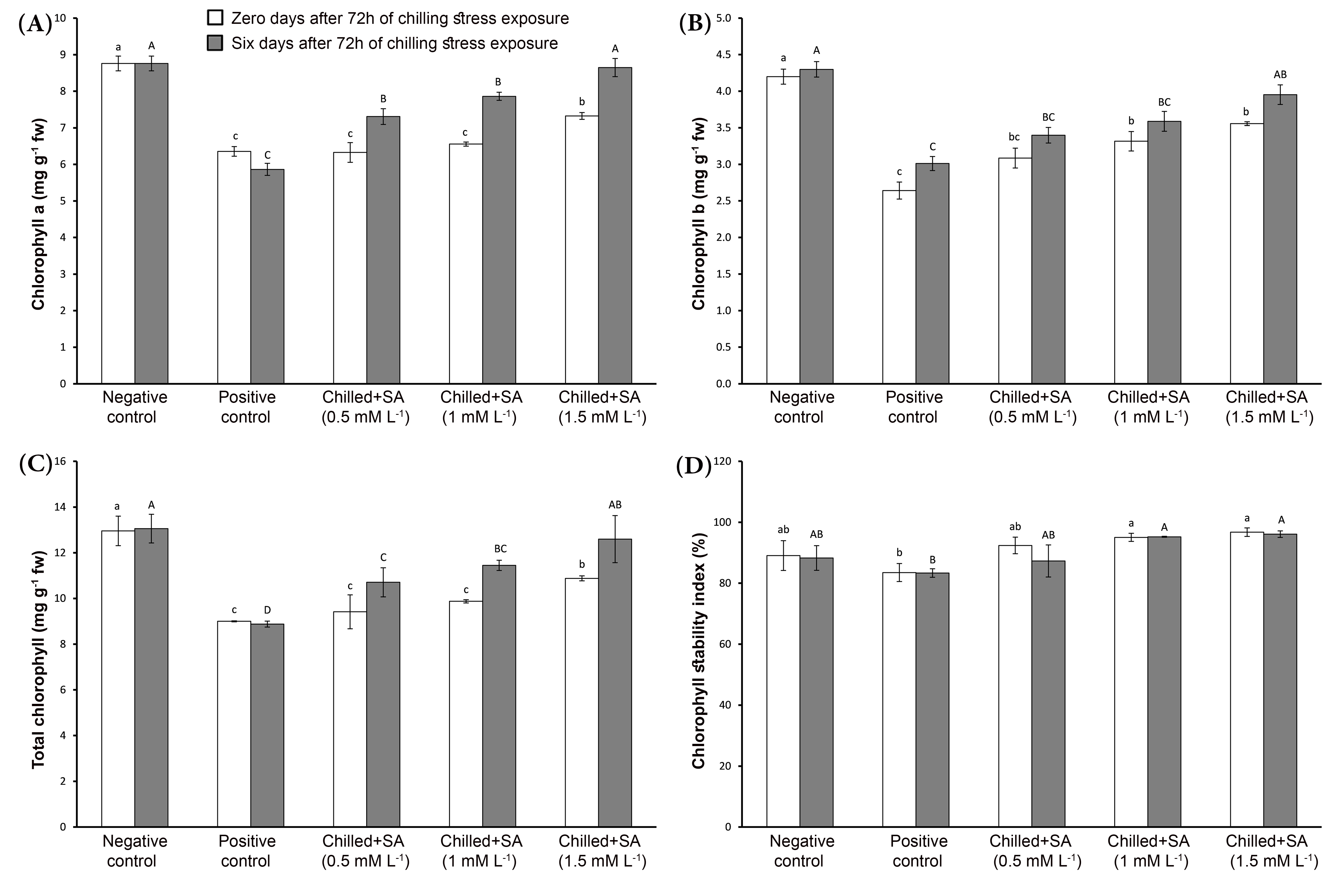
The effects of the exogenous SA application on chilled “Seddik” mango transplants were observed. Defoliation percentages for all treatments, after 21 days of exposure to chilling stress, were significantly high (p ≤ 0.05) compared to those of the negative control (8.33%), but the chilled transplants pretreated with 1.5 mM L−1 SA yielded an acceptable defoliation percentage (23.33%). The positive control revealed the highest defoliation percentage (45.14%) after 21 days of exposure to chilling stress (Figure 2A). Chilling temperatures (lower than 10 °C) cause many physiological changes in chilling-sensitive plants, inducing CI and even mortality in tropical and subtropical species [37]. The plants exhibited a steady increase in leaf fall in the early stages of cold stress exposure and, as days progressed, they became even more defoliated [38]. Similarly, the chilling injury index (CII) of all chilled transplants not treated (positive control) or pretreated with SA was significantly higher (p ≤ 0.05) than that of the negative control (Figure 2B). Thus, SA pretreatment at 1 and 1.5 mM alleviated mango transplants’ CI symptoms (Figure 2E,F, respectively) compared with the positive control (Figure 2D). In fact, the application led to a vital reduction in CI incidence [2,39]. Exposure of the mango transplants to chilling stress for 72 h critically affected plant photosynthetic pigments, chlorophyll a, b, and total pigments, even in transplants pretreated with SA. However, after six days of recovery, the SA-treated mango transplants, specifically, those treated with SA at 1.5 mM, showed pigment values similar to those of the negative control (normal growth conditions). Hence, SA application mitigated the chilled mango transplants’ chilling stress during the recovery period (Figure 3A–C). The lowest total chlorophyll values were 8.99 and 8.87 mg g−1 for positive control treatment after zero and six days of chilling stress, respectively. Similarly, the chlorophyll stability index (CSI) was the lowest for the untreated transplants (positive control) exposed to chilling stress; SA increased mango leaves’ CSI compared with the positive control (Figure 3D).
Under low-temperature conditions, chlorophyll-degrading enzymes’ (chlorophyllases) activity increases, and their biosynthesis is inhibited, leading to a decrease in chlorophyll content in chilled plants [40,41].
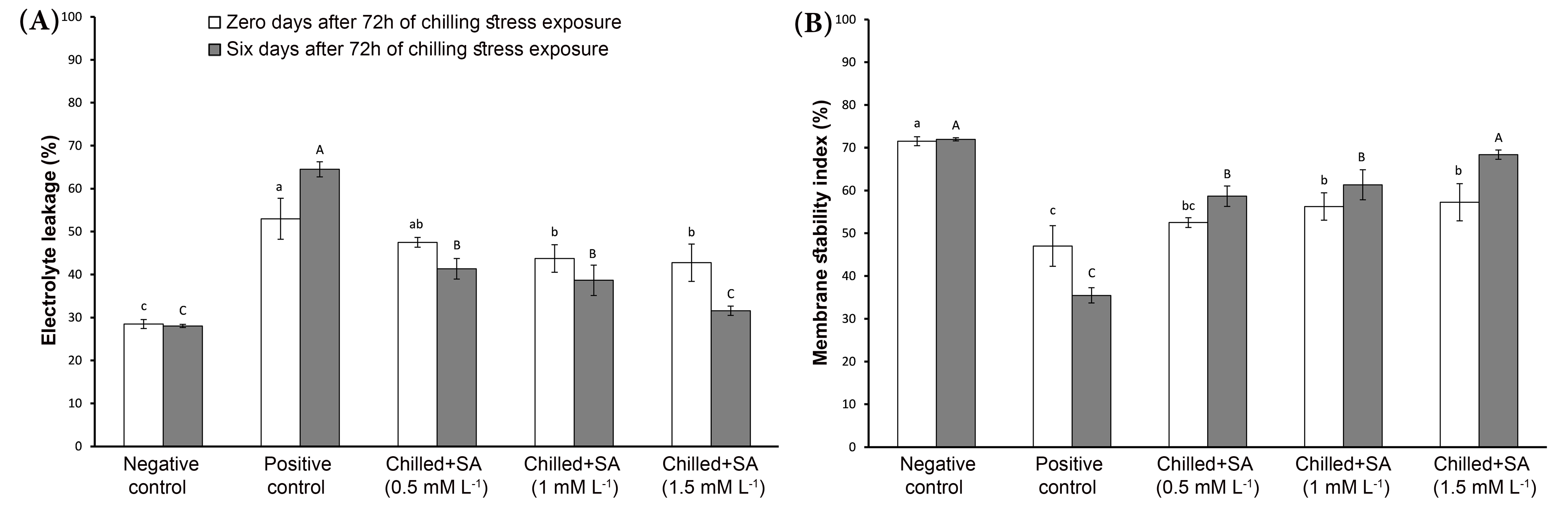
The leaves’ low chlorophyll content at low temperatures can be interpreted as a lack of photosynthetic efficiency [42,43]. Moreover, the reduction in photosynthetic capacity at low temperatures is associated with a decrease in PSII quantum efficiency, the primary target of damage at low temperatures [1,44]. Chilling damage occurs when membranes acquire more saturated fatty acids due to the exposure to low temperatures [44,45,46]. In Figure 4A, it is discernible that the transplants exposed to chilling stress showed an increase in electrolyte leakage percentage after zero (52.99%) or six days (64.51%) compared to those pretreated with SA. The negative control recorded the lowest electrolyte leakage percentage values after zero (28.49%) and six days (28.05%) of exposure to chilling stress.
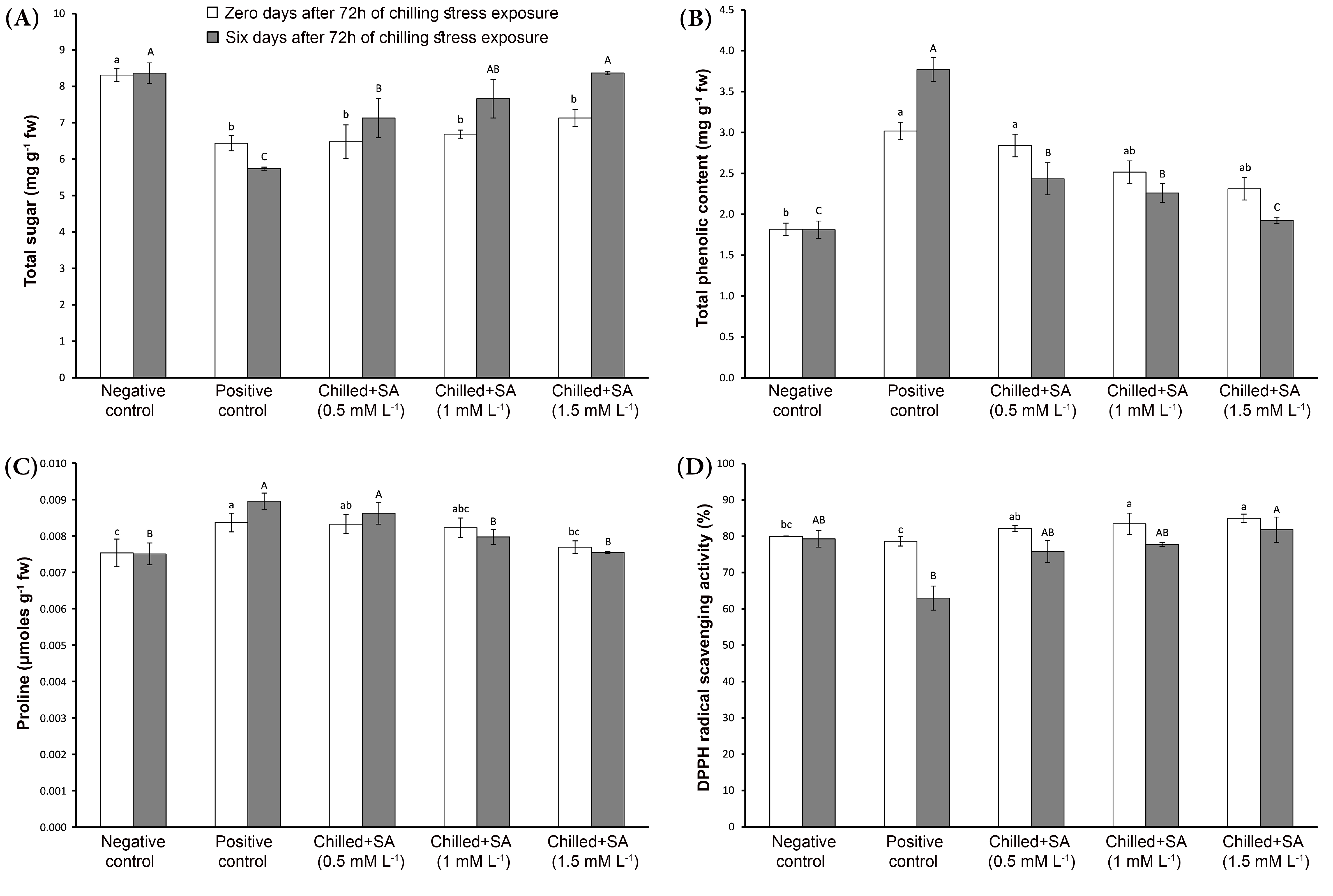
Contrarily, the membrane stability index was the highest for the negative control compared to the positive control, with the lowest values (47.01 and 35.49%) after zero and six days of chilling stress exposure, respectively (Figure 4B). Usually, electrolytes leakage is used to assess the chilling damage [47]. Guinn [48] suggested that the increase in electrolyte leakage is likely due to chilling-induced water stress. Furthermore, increased electrolyte leakage from chilled plants was attributed to membrane deterioration and corresponded to the presence of leaked inorganic and organic ions [46,49,50,51,52]. SA role in maintaining the fatty acids’ content and ratio in the cell membranes could explain its protection of the cell membrane structure [53]. Differences in total sugar content were not statistically significant but accrued gradually with increased SA concentrations, specifically, after six days of recovery, in the chilled mango transplants compared with the positive control (Figure 5A). Generally, plants amass many relevant solutes, such as soluble sugars and amino acids, in response to cold and other osmotic stresses [54,55,56]. During the recovery period, the highest significant (p ≤ 0.05) total phenolic content was recorded in the positive control, with 3.76 mg g−1, and the lowest (p ≤ 0.05) in the negative control, with 1.81 mg g−1. The same trend was also evident after exposure to chilling stress for all treatments (Figure 5B). The transplants pretreated with increased SA concentrations before being subjected to chilling had a lower proline content after zero or six days of stress exposure, as shown in Figure 5C. Rivero et al. [57] observed that mango tissues accrued phenolic compounds under cold stress. A decreased amount of phenolics was observed in SA-pretreated transplants, depicting the effect of SA in alleviating CI in mango under chilling. This finding agrees with Han et al. [58]. However, Wongsheree et al. [45] found that total phenolic compounds in lemon basil leaves, whether young or mature, were not affected by chilling stress exposure at 4 °C. Under cold stress, exogenous SA application caused an increase of soluble carbohydrates in Phaselous vulgaris [59]. SA treatment substantially increased solutes and total soluble sugars, and these osmolytes promoted cryostability in the cell membranes, protecting the plants from cold stress [60,61]. Moreover, the stress conditions increased proline metabolism, attributable to an increase in proline biosynthesis enzymes (pyrolline-5-carboxylate reductase and -glutamyl kinase) [62,63]. Sayyari et al. [64] reported that SA ameliorated CI by inhibiting proline accumulation; the variation in proline content in response to chilling stress primarily depended on the plant genotype [64,65].
Concerning DPPH radical-scavenging activity, SA application positively impacted the chilled mango transplants during recovery by enhancing the function of DPPH. It increased with increases in SA concentration, and the lowest value was measured for the positive control (Figure 5D). The induction of DPPH scavenging activity in chilled mango transplants by SA depended on the concentration applied. This was also observed for banana [66], mango [67,68], and lemons [69] when cold-exposed fruits were treated with SA. Under chilling conditions, SA pre-treatment reduced SOD activity in mango leaves (Figure 6A). After 72 h of exposure to chilling stress, SOD activity significantly (p ≤ 0.05) decreased compared to that in the negative control (0.81 U g−1) but significantly (p ≤ 0.05) increased with the gradual increase of SA concentration (0.65, 0.93, and 1.00 U g−1 with 0.5, 1, and 1.5 mM SA, respectively). The positive control exhibited the lowest significant (p ≤ 0.05) value (0.56 U g−1). This trend was also evident for the recovery period with fewer responses, whereas the negative control showed the highest (0.80 U g−1) and the lowest values (0.38 U g−1). The SOD values in chilled transplants pretreated with SA were 0.42, 0.61, and 0.64 U g−1 with 0.5, 1, and 1.5 mM SA, respectively. Similarly, chilling stress (positive control) significantly (p ≤ 0.05) reduced CAT activity during recovery compared to the negative control, but SA pre-treatment gradually increased CAT activity (p ≤ 0.05) in chilled mango transplants (Figure 6B). Under chilling conditions, POX activity in mango leaves was significantly (p ≤ 0.05) high for the positive control, reaching the highest value (0.73 U g−1) compared to the negative control (0.104 U g−1). As expected, SA pre-treatment gradually decreased POX activity after six days of chilling stress to its normal level (0.104 U g−1) with 1.5 mM L−1 SA treatment (Figure 6C).
Moreover, PPO activity in chilled mango leaves was also significantly (p ≤ 0.05) high after exposure to chilling. However, SA gradually decreased the enzyme activity by about half compared to the negative control. After six days of exposure to chilling stress, PPO activity in chilled mango leaves was significantly (p ≤ 0.05) higher, and the SA treatments significantly (p ≤ 0.05) reduced PPO activity until it approximately returned to the normal level compared to the negative control (Figure 6D).
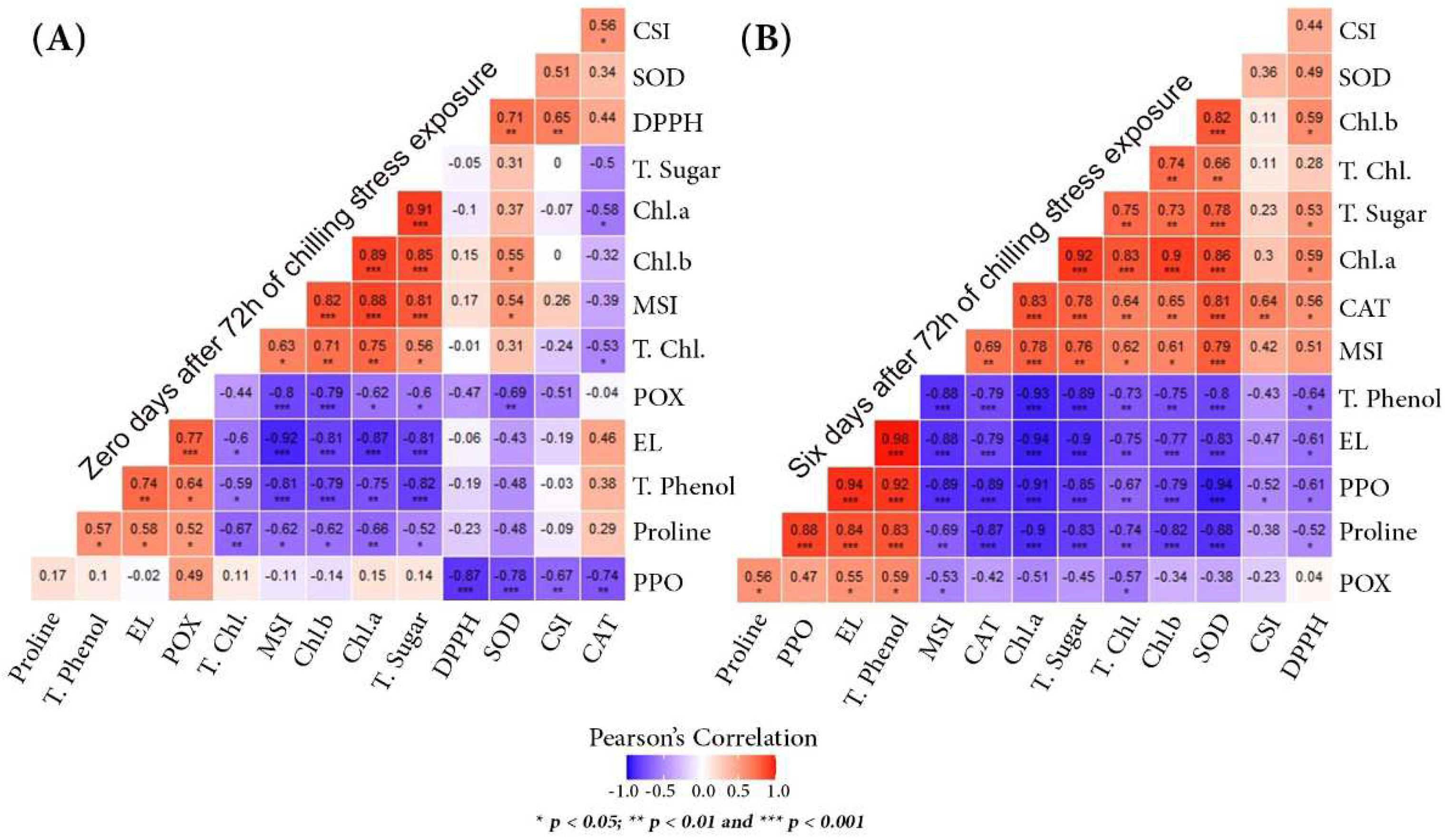
The untreated mango leaves exhibited lower SOD and CAT enzyme activities after six days of chilling stress exposure than the treated ones. It implies less protection against membrane oxidation in untreated leaves [45]. Accordingly, a positive correlation between SOD and CAT enzyme activities was detected after zero (r = 0.34) and six days (r = 0.81) of chilling stress exposure (Figure 7A,B). Generally, SA treatment effectively alleviated the chilled mango transplants’ CI compared to the negative control, reaching the normal levels after six days of stress (recovery). These findings are aligned with those of Chen et al. [9] and Khademi et al. [66], who found that SA treatment effectively reduced banana CI by maintaining membrane integrity and improving antioxidants’ activity. SOD and CAT enzymatic activities exhibited the same trend. SOD, the primary line of defense against ROS-induced oxidative damages, catalyzes the dismutation of two superoxide radicals into H2O2 and O2. CAT converts H2O2 into H2O and O2. Still, POX enables H2O2 oxidation and yields water and another oxidizing molecule. This was evident in the highly positive significant correlation (p < 0.001) between POX enzyme activity and EL (r = 0.77) after 72 h of exposure to chilling stress (Figure 7A). PPO causes tissue browning in most horticultural crops by oxidizing phenolic compounds to quinones. CI alleviation by SA application decreased PPO activity and increased antioxidant systems and specific bioactive chemicals’ concentration, as reported for banana [70], cherry [71], litchi [72], pomegranate [18], and wax apple [73]. After six days of exposure to chilling, PPO had significant (p < 0.001) correlations with EL, total phenol, and proline (Figure 7B). Meanwhile, it showed strong negative correlations with MSI, plant photosynthetic pigments (chlorophyll a, b, and total), SOD, and CAT. SA treatment of cold-stressed plants altered the activities of different enzymatic antioxidants such as CAT, SOD, and POX [74,75]. In the correlation analysis, a significant positive correlation (p < 0.001) between CAT, chlorophyll a, b, MSI, SOD, and total sugar (Figure 7B) was found. SA was shown to enable some crops to recover from cold damage by regulating antioxidative mechanisms [39,59,76,77].
4. Conclusions
The results revealed that exogenous SA application minimized the adverse effects of chilling in local mango transplants of the “Seddik” cultivar, as evidenced by reduced membrane damage and enhanced endogenous production of photosynthetic pigments, total sugar, and DPPH. SA also reduced the content of total phenolic compounds and proline and regulated the activity of antioxidant enzymes, including SOD, CAT, PPO, and POX, especially during the recovery period. These findings suggest that exogenous SA treatment can help mango transplants recover from chilling stress damage.
Author Contributions
Conceptualization, I.H.; methodology, I.H., A.E.M.A., N.K. and A.A; investigation, I.H. and A.A; resources, S.M.S. and A.A.; formal analysis, I.H.; writing—original draft preparation, I.H.; writing—review and editing, I.H. and A.A.; visualization, I.H. and S.M.S.; validation, A.E.M.A. and N.K.; supervision, I.H.; project administration, I.H.; funding acquisition, I.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded and supported by a research grant from the Science, Technology and Innovation Funding Authority (STIFA), under STDF-RG, No. 35967.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors would like to express special gratitude to the Science and Technology Development Fund (STDF) for funding and to the Faculty of Agriculture, Cairo University, for the use of its facilities during this work. Current name: Science, Technology and Innovation Funding Authority (STIFA).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Allen, D.J.; Ort, D.R. Impacts of chilling temperatures on photosynthesis in warm-climate plants. Trends Plant Sci. 2001, 6, 36–42. [Google Scholar] [CrossRef]
- Saleem, M.; Fariduddin, Q.; Janda, T. Multifaceted role of salicylic acid in combating cold stress in plants: A review. J. Plant Growth Regul. 2021, 40, 464–485. [Google Scholar] [CrossRef]
- Cheng, F.; Lu, J.; Gao, M.; Shi, K.; Kong, Q.; Huang, Y.; Bie, Z. Redox signaling and CBF-responsive pathway are involved in salicylic acid-improved photosynthesis and growth under chilling stress in watermelon. Front. Plant. Sci. 2016, 7, 1519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, P.; Ding, Y. Stories of salicylic acid: A plant defense hormone. Trends Plant. Sci. 2020, 25, 549–565. [Google Scholar] [CrossRef] [PubMed]
- Poór, P. Effects of salicylic acid on the metabolism of mitochondrial reactive oxygen species in plants. Biomolecules 2020, 10, 341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, A.K.; Baek, K.H. Salicylic acid biosynthesis and metabolism: A divergent pathway for plants and bacteria. Biomolecules 2021, 11, 705. [Google Scholar] [CrossRef] [PubMed]
- Hadian-Deljou, M.; Esna-Ashari, M.; Sarikhani, H. Effect of pre-and post-harvest salicylic acid treatments on quality and antioxidant properties of ‘Red Delicious’ apples during cold storage. Adv. Hortic. Sci. 2017, 31, 31–38. [Google Scholar] [CrossRef]
- Ezzat, A.; Ammar, A.; Szabó, Z.; Holb, I. Salicylic acid treatment saves quality and enhances antioxidant properties of apricot fruit. Hortic. Sci. 2017, 44, 73–81. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Zhao, X.; Wu, J.E.; He, Y.; Yang, H. Metabolic analysis of salicylic acid-induced chilling tolerance of banana using NMR. Food Res. Int. 2020, 128, 108796. [Google Scholar] [CrossRef]
- Supapvanich, S.; Polpakdee, R.; Wongsuwan, P. Chilling injury alleviation and quality maintenance of lemon basil by preharvest salicylic acid treatment. Emir. J. Food Agric. 2015, 27, 801–807. [Google Scholar] [CrossRef]
- Killadi, B.; Lenka, J.; Chaurasia, R.; Shukla, D.K. Effect of salicylic acid and acylated salicylic acid on the shelf-life of guava (Psidium guajava L.) fruit during storage. Int. J. Chem. Stud. 2019, 7, 1848–1852. [Google Scholar]
- Zhang, Y.; Chen, K.; Zhang, S.; Ferguson, I. The role of salicylic acid in postharvest ripening of kiwifruit. Postharvest Biol. Technol. 2003, 28, 67–74. [Google Scholar] [CrossRef]
- Cai, C.; Li, X.; Chen, K. Acetylsalicylic acid alleviates chilling injury of postharvest loquat (Eriobotrya japonica Lindl.) fruit. Eur. Food Res. Technol. 2006, 223, 533–539. [Google Scholar] [CrossRef]
- Reddy, S.V.R.; Sharma, R.R.; Srivastava, M.; Kaur, C. Effect of pre-harvest application of Salicylic acid on the postharvest behavior of ’Amrapali’ mango fruits during storage. Indian J. Hortic. 2016, 73, 405–409. [Google Scholar] [CrossRef]
- Yang, Z.; Cao, S.; Zheng, Y.; Jiang, Y. Combined salicyclic acid and ultrasound treatments for reducing the chilling injury on peach fruit. J. Agric. Food Chem. 2012, 60, 1209–1212. [Google Scholar] [CrossRef]
- Al-Qurashi, A.D.; Awad, M.A. Postharvest salicylic acid treatment reduces chilling injury of ‘Taify’ cactus pear fruit during cold storage. J. Food Agric. Environ. 2012, 10, 120–124. [Google Scholar]
- Davarynejad, G.H.; Zarei, M.; Nasrabadi, M.E.; Ardakani, E. Effects of salicylic acid and putrescine on storability, quality attributes and antioxidant activity of plum cv. ‘Santa Rosa’. J. Food Sci. Technol. 2015, 52, 2053–2062. [Google Scholar] [CrossRef] [Green Version]
- Sayyari, M.; Babalar, M.; Kalantari, S.; Serrano, M.; Valero, D. Effect of salicylic acid treatment on reducing chilling injury in stored pomegranates. Postharvest Biol. Technol. 2009, 53, 152–154. [Google Scholar] [CrossRef]
- Güneş, N.T.; Yildiz, M.; Variş, B.; Horzum, Ö. Postharvest salicylic acid treatment influences some quality attributes in air-stored pomegranate fruit. J. Agric. Sci. 2020, 26, 499–506. [Google Scholar] [CrossRef]
- Wang, C.Y. Modification of chilling susceptibility in seed¬lings of cucumber and zucchini squash by the bio regulator paclobutrazol (PP333). Sci. Hortic. 1985, 26, 293–298. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef] [Green Version]
- Thakur, A. Use of easy and less expensive methodology to rapidly screen fruit crops for drought tolerance. Acta Hortic. 2004, 662, 231–235. [Google Scholar] [CrossRef]
- Guo, Z.; Huang, M.; Lu, S.; Yaqing, Z.; Zhong, Q. Differential response to paraquat induced oxidative stress in two rice cultivars on antioxidants and chlorophyll a fluorescence. Acta Physiol. Plant 2007, 29, 39–46. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.T.; Smith, F. Colorometric method for determination of sugars and related substances. Analyt. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Blois, M.S. Antioxidant determinations by the use of a stable free radical. Nature 1958, 181, 1199–1200. [Google Scholar] [CrossRef]
- Rahman, M.M.; Islam, M.B.; Biswas, M.; Alam, A.K. In vitro antioxidant and free radical scavenging activity of different parts of Tabebuia pallida growing in Bangladesh. BMC Res. Notes 2015, 8, 621. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, S.P.; Choudhuri, M.A. Implication of water stress-induced changes in the level of endogenous ascorbic acid and hydrogen peroxide in Vigna seedlings. Physiol. Plant 1983, 58, 166–170. [Google Scholar] [CrossRef]
- Marklund, S.; Marklund, G. Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxide dismutase. Eur. J. Biochem. 1974, 47, 469–474. [Google Scholar] [CrossRef]
- Kong, F.X.; Hu, W.; Chao, S.Y.; Sang, W.L.; Wang, L.S. Physiological responses of the lichen Xanthoparmelia mexicana to oxidative stress of SO2. Environ. Exp. Bot. 1999, 42, 201–209. [Google Scholar] [CrossRef]
- Chen, Y.; Cao, X.X.D.; Lu, Y.; Wang, X.R. Effect of rare earth metal ions and their EDTA complex on antioxidant enzymes of fish liver. Bull. Environ. Contam. Toxicol. 2000, 65, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Pütter, J. Peroxidases. In Methods of Enzymatic Analysis, 2nd ed.; Bergmeyer, H.U., Ed.; Verlag Chemie Weinheim, Academic Press: New York, NY, USA, 1974; Volume 2, pp. 685–690. [Google Scholar] [CrossRef]
- Kar, M.; Mishra, D. Catalase, peroxidase and polyphenol oxidase activities during rice leaf senescence. Plant Physiol. 1976, 57, 315–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. Available online: https://www.R-project.org/ (accessed on 29 January 2022).
- Duncan, D.B. Multiple range and multiple F tests. Biometrics 1955, 11, 1–42. [Google Scholar] [CrossRef]
- Lukatkin, A.S.; Brazaityte, A.; Bobinas, C.; Duchovskis, P. Chilling injury in chilling-sensitive plants: A review. Agriculture 2012, 99, 111–124. [Google Scholar]
- Chang, Y.C.; Lin, T.C. Temperature effects on fruit development and quality performance of nagami kumquat (Fortunella margarita [Lour.] Swingle). Hort. J. 2020, 89, 351–358. [Google Scholar] [CrossRef]
- Kang, G.; Wang, C.; Sun, G.; Wang, Z. Salicylic acid changes activities of H2O2-metabolizing enzymes and increases the chilling tolerance of banana seedlings. Environ. Exp. Bot. 2003, 50, 9–15. [Google Scholar] [CrossRef]
- Reddy, M.P. Changes in pigment composition. Hill reaction activity and saccharides metabolism in bajra (Penisetum typhoides) leaves under NaCl salinity. Photosynthetica 1986, 20, 50–55. [Google Scholar]
- Wise, R.R.; Naylor, A.W. Chilling-enhanced photooxidation: Evidence for the role of singlet oxygen and superoxide in the breakdown of pigments and endogenous antioxidants. Plant Physiol. 1987, 83, 278–282. [Google Scholar] [CrossRef] [Green Version]
- Schapendonk, A.H.C.M.; Dolstra, O.; Van Kooten, O. The use of chlorophyll fluorescence as a screening method for cold tolerance in maize. Photosynth. Res. 1989, 20, 235–247. [Google Scholar] [CrossRef]
- Kingston-Smith, A.H.; Foyer, C.H. Bundle sheath proteins are more sensitive to oxidative damage than those of the mesophyll in maize leaves exposed to paraquat or low temperatures. J. Exp. Bot. 2000, 51, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Durner, E.F. Principles of Horticultural Physiology; CABI Publishing: Oxford, UK, 2013; p. 391. [Google Scholar]
- Wongsheree, T.; Ketsa, S.; van Doorn, W.G. The relationship between chilling injury and membrane damage in lemon basil (Ocimum × citriodourum) leaves. Postharvest Biol. Technol. 2009, 51, 91–96. [Google Scholar] [CrossRef]
- Gadallah, F.M.; El-Yazal, S.; Abdel-Samad, G.A. Physiological changes in leaves of some mango cultivars as response to exposure to low temperature degrees. Hortic. Int. J. 2019, 3, 266–273. [Google Scholar] [CrossRef]
- McKay, H.M. Electrolyte leakage from fine roots of conifer seedlings: A rapid index of plant vitality following cold storage. Can. J. For. Res. 1992, 22, 1371–1377. [Google Scholar] [CrossRef]
- Guinn, G. Chilling injury in cotton seedlings: Changes in permeability of cotyledons. Crop. Sci. 1971, 11, 101–102. [Google Scholar] [CrossRef]
- Chinnusamy, V.; Zhu, J.; Zhu, J.K. Cold stress regulation of gene expression in plants. Trends Plant Sci. 2007, 12, 444–451. [Google Scholar] [CrossRef]
- El-Moniem, E.A.A.; Ismail, O.M.; Shaban, A.E.A. Changes in leaves component of some mango cultivars in relation to exposure to low temperature degrees. World J. Agric. Res. 2010, 6, 212–217. [Google Scholar]
- Gadallah, F.M.; El-Yazal, M.A.S.; Abdel-Samad, G.A.; Sayed, A.A. Leakage compositional changes accompanying to exposure of some mango cultivars to low temperature under field conditions. Int. J. Curr. Microbiol. App. Sci. 2020, 9, 1448–1463. [Google Scholar] [CrossRef]
- Sayed, A.A.; Gadallah, F.M.; El-Yazal, M.A.S.; Abdel-Samad, G.A. Impact of exposure to low temperature degrees in field conditions on leaf pigments and chlorophyll fluorescence in leaves of mango trees. JHPR 2020, 10, 30–45. [Google Scholar] [CrossRef]
- Popova, L.P.; Maslenkova, L.T.; Ivanova, A.; Stoinova, Z. Role of salicylic acid in alleviating heavy metal stress. In Environmental Adaptations and Stress Tolerance of Plants in the Era of Climate Change; Ahmad, P., Prasad, M., Eds.; Springer: New York, NY, USA, 2012; pp. 447–466. [Google Scholar] [CrossRef]
- Rhodes, D.; Nadolska-Orczyk, A.; Rich, P.J. Salinity, osmolytes and compatible solutes. In Salinity: Environment-Plants-Molecules; Läuchli, A., Lüttge, U., Eds.; Springer: Dordrecht, The Netherlands, 2002; pp. 181–204. [Google Scholar] [CrossRef]
- Ruelland, E.; Zachowski, A. How plants sense temperature. Environ. Exp. Bot. 2010, 69, 225–232. [Google Scholar] [CrossRef]
- Kamal, A.; Kumar, V.; Muthukumar, M.; Bajpai, A. Morphological indicators of salinity stress and their relation with osmolyte associated redox regulation in mango cultivars. J. Plant Biochem. Biotechnol. 2021, 30, 918–929. [Google Scholar] [CrossRef]
- Rivero, R.M.; Ruiz, J.M.; Garcıa, P.C.; Lopez-Lefebre, L.R.; Sánchez, E.; Romero, L. Resistance to cold and heat stress: Accumulation of phenolic compounds in tomato and watermelon plants. Plant Sci. 2001, 160, 315–321. [Google Scholar] [CrossRef]
- Han, J.; Tian, S.P.; Meng, X.H.; Ding, Z.S. Response of physiologic metabolism and cell structures in mango fruit to exogenous methyl salicylate under low temperature stress. Physiol. Plant 2006, 128, 125–133. [Google Scholar] [CrossRef]
- Soliman, M.H.; Alayafi, A.A.; El Kelish, A.A.; Abu-Elsaoud, A.M. Acetylsalicylic acid enhance tolerance of Phaseolus vulgaris L. to chilling stress, improving photosynthesis, antioxidants and expression of cold stress responsive genes. Bot. Stud. 2018, 59, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Gusta, L.V.; Wisniewski, M. Understanding plant cold hardiness: An opinion. Physiol. Plant 2013, 147, 4–14. [Google Scholar] [CrossRef]
- Luo, Y.L.; Su, Z.L.; Bi, T.J.; Cui, X.L.; Lan, Q.Y. Salicylic acid improves chilling tolerance by affecting antioxidant enzymes and osmoregulators in sacha inchi (Plukenetia volubilis). Rev. Bras. Bot. 2014, 37, 357–363. [Google Scholar] [CrossRef]
- Zhu, B.; Su, J.; Chang, M.; Verma, D.P.S.; Fan, Y.L.; Wu, R. Overexpression of a Δ1-pyrroline-5-carboxylate synthetase gene and analysis of tolerance to water-and salt stress in transgenic rice. Plant Sci. 1998, 139, 41–48. [Google Scholar] [CrossRef]
- Forlani, G.; Trovato, M.; Funck, D.; Signorelli, S. Regulation of Proline Accumulation and Its Molecular and Physiological Functions in Stress Defence. In Osmoprotectant-Mediated Abiotic Stress Tolerance in Plants; Hossain, M., Kumar, V., Burritt, D., Fujita, M., Mäkelä, P., Eds.; Springer: Cham, Switzerland, 2019; pp. 73–97. [Google Scholar] [CrossRef]
- Sayyari, M.; Ghanbari, F.; Fatahi, S.; Bavandpour, F. Chilling tolerance improving of watermelon seedling by salicylic acid seed and foliar application. Not. Sci. Biol. 2013, 5, 67–73. [Google Scholar] [CrossRef] [Green Version]
- Bastam, N.; Baninasab, B.; Ghobadi, C. Improving salt tolerance by exogenous application of salicylic acid in seedlings of pistachio. Plant. Growth Regul. 2013, 69, 275–284. [Google Scholar] [CrossRef]
- Khademi, O.; Ashtari, M.; Razavi, F. Effects of salicylic acid and ultrasound treatments on chilling injury control and quality preservation in banana fruit during cold storage. Sci. Hortic. 2019, 249, 334–339. [Google Scholar] [CrossRef]
- Junmatong, C.; Faiyue, B.; Rotarayanont, S.; Uthaibutra, J.; Boonyakiat, D.; Saengnil, K. Cold storage in salicylic acid increases enzymatic and non-enzymatic antioxidants of Nam Dok Mai No. 4 mango fruit. Sci. Asia 2015, 41, 12–21. [Google Scholar] [CrossRef] [Green Version]
- Awad, M.A.; Al-Qurashi, A.D. Postharvest salicylic acid and melatonin dipping delay ripening and improve quality of ‘Sensation’ Mangoes. Philipp. Agric. Sci. 2021, 104, 34–44. [Google Scholar]
- Siboza, X.I.; Bertling, I. The effects of methyl jasmonate and salicylic acid on suppressing the production of reactive oxygen species and increasing chilling tolerance in ‘Eureka’lemon [Citrus limon (L.) Burm. F.]. J. Hortic. Sci. Biotechnol. 2013, 88, 269–276. [Google Scholar] [CrossRef]
- Chen, J.Y.; He, L.H.; Jiang, Y.M.; Kuang, J.F.; Lu, C.B.; Joyce, D.C.; Macnish, A.; He, Y.X.; Lu, W.J. Expression of PAL and HSPs in fresh-cut banana fruit. Environ. Exp. Bot. 2009, 66, 31–37. [Google Scholar] [CrossRef]
- Dokhanieh, A.Y.; Aghdam, M.S.; Fard, J.R.; Hassanpour, H. Postharvest salicylic acid treatment enhances antioxidant potential of cornelian cherry fruit. Sci. Hortic. 2013, 154, 31–36. [Google Scholar] [CrossRef]
- Kumari, P.; Barman, K.; Patel, V.B.; Siddiqui, M.W.; Kole, B. Reducing postharvest pericarp browning and preserving health promoting compounds of litchi fruit by combination treatment of salicylic acid and chitosan. Sci. Hortic. 2015, 197, 555–563. [Google Scholar] [CrossRef]
- Supapvanich, S.; Mitsang, P.; Youryon, P.; Techavuthiporn, C.; Boonyaritthongchai, P.; Tepsorn, R. Postharvest quality maintenance and bioactive compounds enhancement in ‘Taaptimjaan’ wax apple during short-term storage by salicylic acid immersion. Hortic. Environ. Biotechnol. 2018, 59, 373–381. [Google Scholar] [CrossRef]
- Mutlu, S.; Karadağoğlu, Ö.; Atici, Ö.; Nalbantoğlu, B. Protective role of salicylic acid applied before cold stress on antioxidative system and protein patterns in barley apoplast. Biol. Plant 2013, 57, 507–513. [Google Scholar] [CrossRef]
- Mo, Y.; Gong, D.; Liang, G.; Han, R.; Xie, J.; Li, W. Enhanced preservation effects of sugar apple fruits by salicylic acid treatment during postharvest storage. J. Sci. Food Agric. 2008, 88, 2693–2699. [Google Scholar] [CrossRef]
- Mutlu, S.; Atıcı, Ö.; Nalbantoğlu, B.; Mete, E. Exogenous salicylic acid alleviates cold damage by regulating antioxidative system in two barley (Hordeum vulgare L.) cultivars. Front. Life Sci. 2016, 9, 99–109. [Google Scholar] [CrossRef] [Green Version]
- Ignatenko, A.; Talanova, V.; Repkina, N.; Titov, A. Exogenous salicylic acid treatment induces cold tolerance in wheat through promotion of antioxidant enzyme activity and proline accumulation. Acta Physiol. Plant 2019, 41, 1–10. [Google Scholar] [CrossRef]
Figure 1.
Minimum mean January, February, and March temperatures (open-air temperature) during the last decade in Giza district, Egypt.
Figure 1.
Minimum mean January, February, and March temperatures (open-air temperature) during the last decade in Giza district, Egypt.

Figure 2.
Defoliation percentage for each treatment after 21 days of chilling stress (A), chilling injury index (B), chilling injury (CI) symptoms in mango transplants, negative control (C), CI symptoms in chilled mango transplants, positive control (D), CI symptoms in chilled mango transplants pretreated with 1 mM SA (E) and 1.5 mM SA (F). Bars with different letters represent significantly different data at 95% confidence, as determined by Duncan’s Multiple Range Test. Error bars represent the standard deviation.
Figure 2.
Defoliation percentage for each treatment after 21 days of chilling stress (A), chilling injury index (B), chilling injury (CI) symptoms in mango transplants, negative control (C), CI symptoms in chilled mango transplants, positive control (D), CI symptoms in chilled mango transplants pretreated with 1 mM SA (E) and 1.5 mM SA (F). Bars with different letters represent significantly different data at 95% confidence, as determined by Duncan’s Multiple Range Test. Error bars represent the standard deviation.

Figure 3.
Changes in leaf chlorophyll content; chlorophyll a content (A); chlorophyll b content (B); total chlorophyll (C), and chlorophyll stability index (D) in mango transplants under the studied treatments. Different lower-case letters indicate statistical differences between treatments at zero days after 72 h of chilling stress exposure, while the upper-case letters indicate significant differences between treatments at six days at 95% confidence, as determined by Duncan’s Multiple Range Test. Error bars represent the standard deviation.
Figure 3.
Changes in leaf chlorophyll content; chlorophyll a content (A); chlorophyll b content (B); total chlorophyll (C), and chlorophyll stability index (D) in mango transplants under the studied treatments. Different lower-case letters indicate statistical differences between treatments at zero days after 72 h of chilling stress exposure, while the upper-case letters indicate significant differences between treatments at six days at 95% confidence, as determined by Duncan’s Multiple Range Test. Error bars represent the standard deviation.

Figure 4.
Changes in electrolyte leakage (A) and membrane stability index (B) of mango transplants’ leaves under the studied treatments. Different lower-case letters indicate statistical differences between treatments at zero days after 72 h of chilling stress exposure, while the upper-case letters indicate significant differences between treatments at six days at 95% confidence, as determined by Duncan’s Multiple Range Test. Error bars represent the standard deviation.
Figure 4.
Changes in electrolyte leakage (A) and membrane stability index (B) of mango transplants’ leaves under the studied treatments. Different lower-case letters indicate statistical differences between treatments at zero days after 72 h of chilling stress exposure, while the upper-case letters indicate significant differences between treatments at six days at 95% confidence, as determined by Duncan’s Multiple Range Test. Error bars represent the standard deviation.

Figure 5.
Changes in total sugar (A), total phenolic content (B), proline content (C), and DPPH (D) in mango transplant leaves. Different lower-case letters indicate statistical differences between treatments at zero days after 72 h of chilling stress exposure, while the upper-case letters indicate significant differences between treatments at six days at 95% confidence, as determined by Duncan’s Multiple Range Test. Error bars represent the standard deviation.
Figure 5.
Changes in total sugar (A), total phenolic content (B), proline content (C), and DPPH (D) in mango transplant leaves. Different lower-case letters indicate statistical differences between treatments at zero days after 72 h of chilling stress exposure, while the upper-case letters indicate significant differences between treatments at six days at 95% confidence, as determined by Duncan’s Multiple Range Test. Error bars represent the standard deviation.

Figure 6.
Changes in the activities of superoxide dismutase (A), catalase (B), peroxidase (C), and polyphenol oxidase (D) in mango transplant leave. Different lower-case letters indicate statistical differences between treatments at zero days after 72 h of chilling stress exposure, while the various upper-case letters indicate significant differences between treatments at six days at 95% confidence, as determined by Duncan’s Multiple Range Test. Error bars represent the standard deviation.
Figure 6.
Changes in the activities of superoxide dismutase (A), catalase (B), peroxidase (C), and polyphenol oxidase (D) in mango transplant leave. Different lower-case letters indicate statistical differences between treatments at zero days after 72 h of chilling stress exposure, while the various upper-case letters indicate significant differences between treatments at six days at 95% confidence, as determined by Duncan’s Multiple Range Test. Error bars represent the standard deviation.

Figure 7.
Pearson’s correlation coefficient analysis for each sampling time, (A) for day zero and (B) for day six after 72 h of chilling stress exposure. R is presented in different colors; the legend shows the color range of different R values with * p < 0.05, ** p < 0.01, and *** p < 0.001 as indicators of statistical significance.
Figure 7.
Pearson’s correlation coefficient analysis for each sampling time, (A) for day zero and (B) for day six after 72 h of chilling stress exposure. R is presented in different colors; the legend shows the color range of different R values with * p < 0.05, ** p < 0.01, and *** p < 0.001 as indicators of statistical significance.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hmmam, I.; Ali, A.E.M.; Saleh, S.M.; Khedr, N.; Abdellatif, A. The Role of Salicylic Acid in Mitigating the Adverse Effects of Chilling Stress on “Seddik” Mango Transplants. Agronomy 2022, 12, 1369. https://doi.org/10.3390/agronomy12061369
AMA Style
Hmmam I, Ali AEM, Saleh SM, Khedr N, Abdellatif A. The Role of Salicylic Acid in Mitigating the Adverse Effects of Chilling Stress on “Seddik” Mango Transplants. Agronomy. 2022; 12(6):1369. https://doi.org/10.3390/agronomy12061369
Chicago/Turabian StyleHmmam, Ibrahim, Amr E. M. Ali, Samir M. Saleh, Nagwa Khedr, and Abdou Abdellatif. 2022. "The Role of Salicylic Acid in Mitigating the Adverse Effects of Chilling Stress on “Seddik” Mango Transplants" Agronomy 12, no. 6: 1369. https://doi.org/10.3390/agronomy12061369
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.