1. Introduction
Biostimulants are organic and inorganic compounds or microorganisms that when applied to plants can enhance plants’ growth, yield, and tolerance to stress [
1]. They have been found to increase plants’ nutrient use efficiency, reduce the supply of fertilizers to the soil, and increase agricultural sustainability [
2]. Protein hydrolysates (PHs) are a particular category of plant biostimulants constituted of a complex mixture of amino acids and oligo- and polypeptides, obtained from chemically or enzymatically hydrolyzed proteins, and deriving from animal and vegetable agro-industrial byproducts, such as blood, viscera, seeds, hay, and plant residues [
2]. From this point of view, they can contribute to the recycling of agro-food waste and reduce the impact that these byproducts have on human and environmental health [
3]. There has been less consensus among the scientific community in regard to animal-derived PHs, produced via alkaline or acid hydrolysis, compared to plant-derived PHs. This is mainly due to the potential adverse effects that can be observed after repeated foliar applications of animal-derived PHs, such as increased leaf NaCl accumulation [
4], leaf chlorination, a reduction in the photosynthetic rate, or other phytotoxic effects [
5,
6]. Moe [
7] also attributed the negative effects of animal-derived PHs to their higher content of free amino acids. Rouphael et al. [
8] found that proline levels increased after repeated foliar applications of animal-derived PHs in basil, with a consequent inhibitory effect on root elongation, N uptake, plant growth, and yield.
A high content of free amino acids is avoided during the production of plant-based biostimulants by exploiting enzymes that have selective cleavage sites on proteins. In this case, the biostimulant products are mixtures of amino acids and peptides of different lengths [
9].
It has been documented that the application of foliar or root PHs improved the absorption of water and macro- and microelements in several horticultural crops [
9,
10]. These results were attributed mostly to PHs’ effect on root architecture and their ability to increase soil mineral solubility, as well as the activity of key enzymes related to nitrogen metabolism, such as nitrate reductase and glutamine synthetase [
11]. PHs have also been found to improve crop quality by increasing the concentration of phytonutrients, such as carotenoids, polyphenols, and flavonoids [
12], and the constraint of nitrate content in many species of leafy vegetables, such as rocket, radish, cabbage, pak choi, onion, swiss chard, spinach, lettuce, celery, and parsley [
13]. In addition, PHs have been shown to alleviate the adverse effects on crop productivity of certain abiotic stresses, such as salinity, high temperature, and drought [
14,
15,
16]. The latter has been related to an increase in antioxidant molecules and/or enzyme activities able to detoxify the reactive oxygen species (ROS) generated during stress conditions.
Lactuca sativa L. is a vegetable that is widespread all over the world and is a widely cultivated species in Mediterranean areas [
17]. Lettuce is classified into seven major groups which differ in their textures, leaves, head shapes, and colors [
18]. It has a short development cycle and it is valued for its high content of water, minerals, vitamins, chlorophylls, fibers, and polyphenols [
19]. Polyphenols, including flavonoids and phenolic acids, have a well-known antioxidant activity and play an important role in extending plant shelf life, as well as in human nutrition, because they are capable of preventing or reducing the oxidative states typical of many inflammatory states related to neurodegenerative diseases and cancer [
20,
21].
In this study we aimed to understand the effect of two biostimulants—protein hydrolysates (legume-derived protein hydrolysates Trainer®) and natural plant extracts (namely tropical plant extracts Auxym®)—on the primary and secondary metabolism of two lettuce cultivars, green and red salanova®, grown in a glasshouse. Mineral composition and morpho-physiological and qualitative traits (SPAD index, gas exchanges, concentration of organic acids, phenolic acids, carotenoid flavonoids) were evaluated in relation to yield. This study can improve our knowledge on the ways in which hydrolyzed proteins and plant extract biostimulants affect plants.
4. Discussion
The leaf area and number of lettuce leaves are primarily genetically regulated since some cultivars are characterized by a higher number of leaves, whereas others are characterized by fewer but larger leaves [
29]. In this work, green salanova had a higher number of leaves, dry biomass, and dry matter, compared to the red cultivar, in the absence of biostimulant application. The latter, on the other hand, produced a greater leaf area and higher fresh yield than the green cultivar. Fresh yield and dry matter (%) differentiation, among lettuce cultivars of the same species has also been reported by Rouphael et al. [
18], whereas other studies have shown that green lettuce cultivars had greater plant vegetative growth and productivity than red ones [
30]. The latter has also been observed for green cultivars compared to the red ones of other species, as in the case of
Perilla frutescens [
31].
In our work, the effect of biostimulants was significant in all the biometric parameters examined. Both biostimulants favored the fresh yield of lettuce cultivars, whereas the biostimulant effects on the other biometric parameters depended on the type of biostimulant treatment, as well as the cultivar type. Most lettuce biometric parameters were favored more through TPE application, apart from lettuce dry matter content, which was actually decreased by both biostimulant applications.
Increases in yield and biometric parameters after PH biostimulant treatments were documented in the recent literature for various vegetable crops, such as lettuce, spinach, rocket, and baby leaf [
32,
33,
34,
35]. An increase in the yield of lettuce plants [
36] and in the roots and shoots of tomato plants [
4] was observed following a radical application of Trainer
®. This latter treatment of lettuce plants grown in greenhouses and under conditions of saline stress improved plants’ performance in terms of their marketable yield, dry biomass, and root dry weight, as shown by Lucini et al. [
11]. The authors found a correlation between the better performance of the treated plants with an increase in photosynthetic activity and nitrogen metabolism. The use of Auxym
® on four tomato (
Solanum lycopersicum L.) landraces [
37] showed that it increased the yield of two landraces by 21% and 35%, compared to untreated plants. Ertani et al. [
12] examined the response of
Capsicum chinensis L. plants to two biostimulants, one obtained from enzymatic hydrolysis of
Medicago sativa L. plants (AH), and another one obtained from cool extraction of the red grape skin of
Vitis vinifera L (RG). AH differed from RG in terms of having a higher content of total organic carbon, total phenols, and indoleacetic acid. RG had a slightly higher content of sugars and the hormone isopentenyladenosine than AH. The authors showed a strong increase in the fresh weight of leaves and fruits, and in the number of fruits, compared to the control plants, after the application of the two biostimulants. These results are comparable to ours, where the lettuce leaf number and the weight of green and red Salanova increased when both type of biostimulants were used. However, the effect was variant on pepper, based on the biostimulant used and the dose.
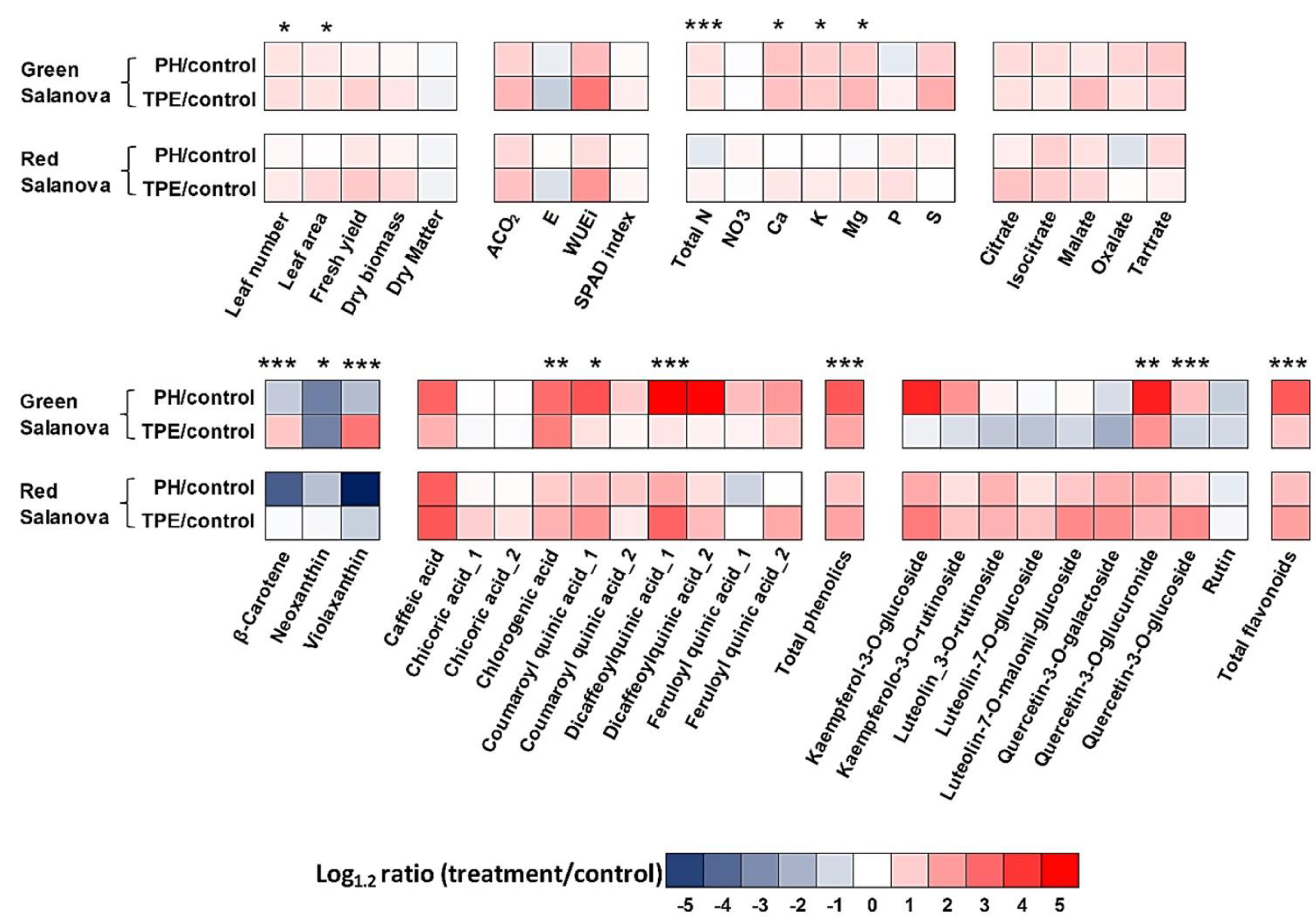
In our work, the two cultivars differed statistically in terms of mineral content, with the red cultivar exhibiting the highest total N content, and all the elements examined (
Table 2). The nitrate content was also higher in the red cultivar (3487 mg kg
−1 FW), compared to the green one, and this value was below the limits established by EU regulations [
38]. A significant variation in the mineral content (K, Ca, Mg, Na) in seven cultivars of the same lettuce species was shown by Rouphael et al. [
18]. Furthermore, greater retention of K in the cytoplasm and vacuole of red lettuce (Salad Bowl lettuce (
Lactuca sativa L. var. Acephal), compared to the green cultivar, was found by Carillo et al. [
39]. In this study, biostimulant application had a significant effect on all minerals but NO
3− and S. The two biostimulants had a similar effect on the NO
3−, S, and P contents of both cultivars. However, the biostimulant effect on certain lettuce mineral components proved to be influenced also by cultivar type, as only green lettuce appeared to respond positively to both biostimulants, at least in terms of the K, Ca, and Mg concentrations. Furthermore, using one or both biostimulants, the concentrations of N, K, Ca, and Mg in the green cultivar were enhanced and reached similar levels to those of the red cultivar’s control levels. The latter is crucial because it could enable us to bio-enhance commercial cultivars in which the low concentrations of certain minemals beneficial for human health can be determined genetically and not by other exogenous factors. That was also one of the main reasons that originally led to the examination of biostimulants as a potential tool for the bio-enhancement of edible plant parts (leaves, fruits) with minerals that are considered essential for sustaining human health [
37,
39].
The enhanced absorption of certain nutrients (N, K, Ca, and Mg) by only the green cultivar could be related with the increase in green salanova’s total N (%) after the application of PH and TPE biostimulants. In contrast, the two biostimulants reduced (PH) or did not have an effect (TPE) on the total N (%) of red lettuce. Several studies have highlighted an increase in the metabolism and assimilation of nitrogen, and the enzymes involved (nitrate reductase, nitrite reductase, glutamine synthetase, glutamate synthase, and aspartate aminotransferase), following the application of protein hydrolysates [
10,
40,
41,
42]. Amino acids, absorbed by plants from the environment, are important sources of organic N, used for the synthesis of proteins or other organic molecules. As a mode of action, amino acids and peptides can also act as elicitors by binding to receptors on cell membranes and triggering morphological, physiological, and biochemical changes in the plant. The microorganisms of the rhizosphere secrete enzymes that hydrolyze the peptides into smaller fractions and that can act as signal molecules in the plant to increase the absorption of minerals [
9]. A root hair growth-promoting peptide was identified in Trainer
® [
9,
43], which can lead to an improvement in the root system, and greater absorption of nutrients by the treated plants compared to those not treated with the biostimulant. In particular, Trainer
® has been found to increase the expression of genes involved in both nitrogen uptake and transport [
42,
44]. Similarly to Trainer
®, Auxym
® application is clearly involved in the expression of genes responsible for the absorption of certain nutrients. For example, an increase in Mg and K content has recently been demonstrated for tomato plants treated with Auxym
® [
37]. The authors emphasized the importance of this result, because the use of Auxym
® would reduce the contribution of K as a fertilizer since it is one of the main elements required for tomato cultivation.
The organic acid differentiation (isocitrate, citrate, and tartrate) observed among the two salanova types examined in this study could be related to nitrate or potassium uptake and assimilation and their influence on the uptake and distribution of other ions. When nitrates are supplied to plants, the anion uptake exceeds the cation uptake, which may lead to an increase in the concentration of certain organic acids in order to maintain the required electroneutrality among the plant and the nutrient medium [
45]. Thus, the nitrate itself, when accumulated, could upregulate the biosynthesis of organic acids [
46]. In our work, the biosynthesis of higher concentrations of organic acids (isocitric, citric, and tartrate acids) in the green cultivar than in the red lettuce cultivar may have been necessary due to the possible regulatory role of organic acids as anti-anions for nitrates. Concerning the red lettuce, the role of the counter-ion was probably exerted more efficiently by K
+, which was present at significantly higher concentrations in the red lettuce than in the green lettuce [
47]. The higher levels of K
+ and Ca
2+ found in red lettuce could be responsible for the higher recorded amount of oxalate, as the latter is usually involved in the regulation of Ca
2+ levels and the Na to K balance [
48]. The two biostimulants used increased the content of malate and citrate, compared to the control. Ertani et al. [
10], applying alfalfa PH to corn plant roots, showed an increase in the activity of genes that code for enzymes of the Krebs cycle and glycolysis. Furthermore, the authors recorded an increase in the activity of the enzymes involved in these pathways of primary metabolisms, such as malate dehydrogenase, isocitrate dehydrogenase, and citrate synthase. The citric acid content increased in four tomato (
Solanum lycopersicum L.) landraces examined by Rouphael et al. [
37] after Auxym
® application. As previously illustrated, both PH and TPE modulated the physiological mechanisms of the treated plants.
Red lettuce showed a higher SPAD index than the green one, as expected, since the SPAD index is significantly correlated to the content of chlorophylls and total flavonoids [
49]. Total flavonoids can play a role as a sunscreen, protecting leaf tissues from UV-B radiation and filtering out yellow-green light, which is partly responsible for chlorophyll excitation [
50]. The lower WUEi and higher transpiration observed in the green lettuce cultivar (
Table 4) could be the result of its reduced ability to accumulate K, Ca, and Mg in the leaves when compared to the red cultivar. The lower K, Ca, and Mg concentrations in the green lettuce leaves could have also increased stomatal resistance and decreased photosynthetic activity [
39]. K is an indispensable element for stomatal movements. If its concentration is reduced, physiological functions can be compromised [
51]. A reduction in Ca levels can also have negative effects on stomatal opening, compromising the assimilation of CO
2 [
39]. Mg is important for the activation of Rubisco, the essential enzyme for photosynthesis (ribulose 1,5-bisphosphate carboxylase-oxygenase). Magnesium is also involved in the subsequent conversion reaction of 3PGA into 1,3-bisphosphoglycerate acid (BPGA) by means of an enzyme that uses MgATP [
45]. A reduction in Mg was also associated with a lower translocation of sucrose from the leaves to the phloem, resulting in an increase in the leaf dry matter [
52], as shown in the green cultivar in our work. The biostimulant effect was significant for all the physiological parameters examined (SPAD, A
CO2, E, and WUEi) in this study. An increase in the SPAD index, photosynthetic activity, and other physiological parameters after the application of plant-derived PH (Trainer
®), was also shown in other studies regarding tomatoes [
18] and lettuce [
36]. The positive effect of the plant-derived protein hydrolysate (Trainer
®) on plant physiological parameters could be attributed to the stimulation of nitrogen uptake and assimilation, which could enhance plant photosynthesis and carbon assimilation [
4].
The content of total phenols and phenolic acids is genotype-dependent, as demonstrated for different leafy vegetables [
53]. For lettuce in particular, the red pigmentation is associated with the presence of antioxidant molecules, such as phenolic acids, and flavonoids [
54]. In our work, the red cultivar had a higher total phenol content than the green one. Chlorogenic acid was the most abundant phenolic acid in lettuce, as already documented in several other papers [
18,
55]. Depending on the cultivar type, the two biostimulants were proved able to increase the concentration of total phenols and some of the phenolic acids identified in this work (chlorogenic acid, caffeic acid, coumaroyl quinic acid isomer 1, and dicaffeoylquinic acid isomer 1), compared to the control. Similarly, total phenolic acids including caffeic acid were strongly enhanced in pepper leaves after the application of two similar biostimulants [
12]. The accumulation of antioxidant molecules, such as phenols, flavonoids, and carotenoids, has been associated with the PH’s biostimulant modification of plant primary and secondary metabolism [
9]. TPEs were also documented to improve plants’ metabolism via the accumulation of starch, fructose, glucose, and lycopene [
37].
In our work, the red cultivar showed a 14-fold higher content of total flavonoids and single flavonoids than the green one. Similar results were observed by Kim et al. [
54]. A higher flavonoid content is considered an advantage, as it allows for more choices among consumers [
19,
56]. The two biostimulants significantly increased total flavonoids, quercetin-3-
O-glucuronide, and quercetin-3-
O-glucoside in the red cultivar (CxB,
Table 6). According to many authors, plant-derived PHs, as a mode of action, activate secondary metabolism through an increase in the expression of genes encoding phenylalanine (tyrosine), an ammonia-lyase enzyme [
57,
58]. Cinnamic acid and subsequently coumaric acid originate from phenylalanine, transformed by the PAL enzyme. Starting from coumaric acid, the synthesis of flavonoids begins. The class of flavonoids includes many plant pigments (anthocyanidins with red and blue colors, yellow flavonols, and others), and condensed tannins. An increase in the synthesis of flavonoids in maize plants, treated with alfalfa PH in saline conditions, was found bobservedy Ertani et al. [
10]. Similarly, Boselli et al. [
59] observed an increase in soluble solids, total phenols, and anthocyanins in grapes following the foliar application of plant-derived PHs.
Carotenoid differentiation among the two lettuce types was expected since the two cultivars had different pigmentation. As expected, all carotenoids exhibited higher values in the red cultivar compared to the green one under control conditions. The effects of biostimulant application on all lettuce carotenoids were subjected to significant interactions with the type of cultivar. More experimentation is required regarding how biostimulants could affect lettuce pigmentation since both biostimulants reduced and/or did not have any effect on the pigmentation of these lettuce plants.
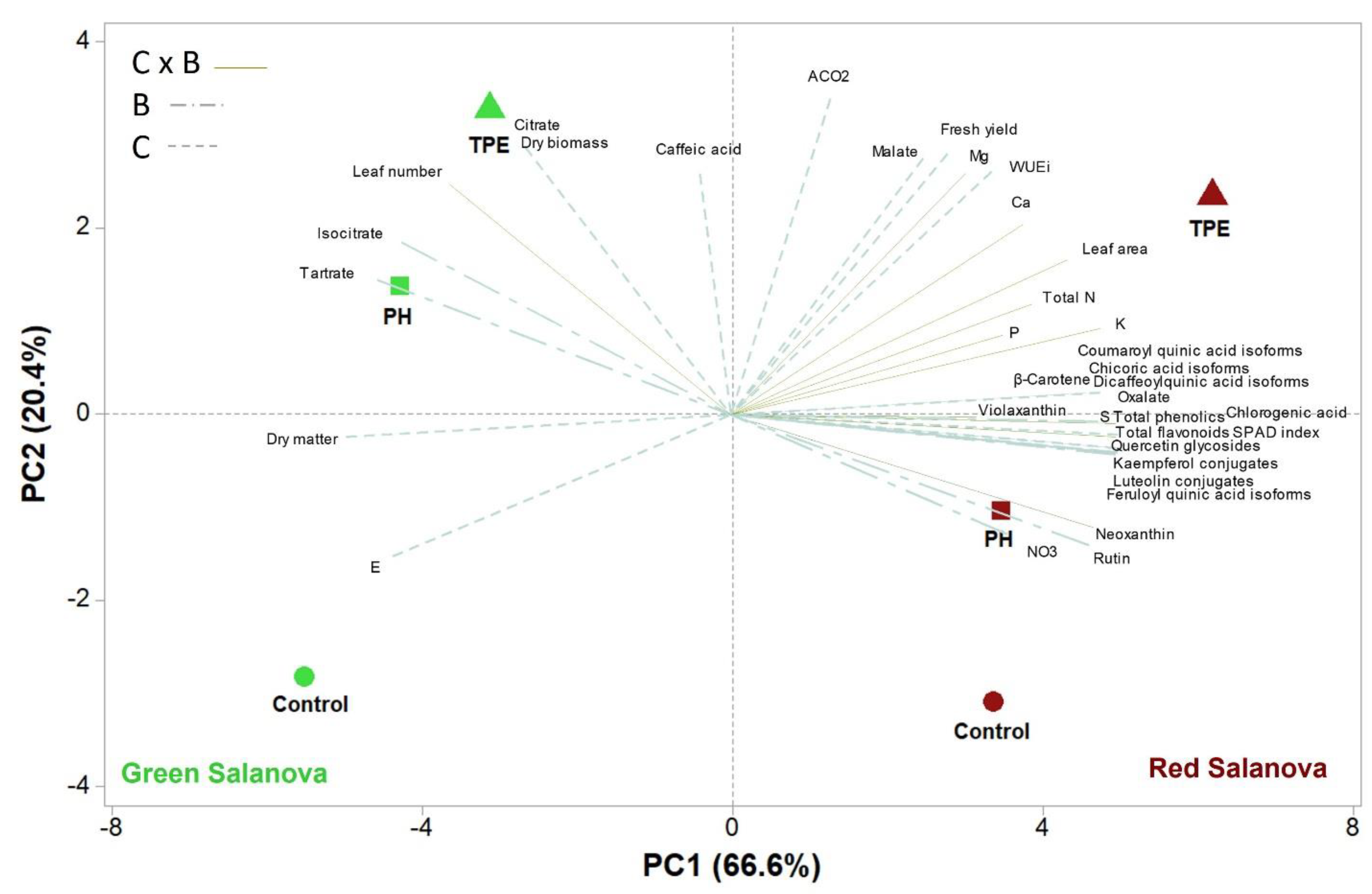
Principal component analysis provided a comprehensive framework for evaluating the two biostimulants’ effects on green and red lettuce cultivar yields and the modulation of primary and secondary metabolism. The two cultivars were clearly separated via the PCA. However, PCA analysis confirmed that several primary and secondary compounds were subjected to cultivar × biostimulant interaction since only green lettuce biostimulant treatments (PH and TPE) were grouped in the same positive quadrant of PC2. The latter verified our previous results indicating that the effects of biostimulant application may differ depending on lettuce cultivar types.


 ,
, 





{kind=link}
{kind=link}