1. Introduction
Light interception and its direct impact on crop growth have been important concepts with respect to field crops [
1]. Like many other crops, the amounts of incoming intercepted photosynthetically active radiation (I PAR) and radiation use efficiency (RUE) of the canopy for biomass production have been highlighted as the most important determinants of the productive potentiality of the leguminous crop stands like mungbean [
2], pigeon pea [
3], lentil [
4], etc. Basu et al. [
5] recorded up to 97% variation in intercepted PAR, which could be explained by the biomass accumulation in case of transplanted rice. On the other hand, Oluwasemire and Odugbenro [
6] noted the maximum increment in plant biomass for groundnut with a PAR interception to the tune of 55–60%. Further studies indicated that the incoming PAR intercepted by crop canopy is largely governed by the leaf area index (LAI) and canopy architecture [
7]. Basically, leaf area is one of the major determinants of PAR interception and its utilization for biomass accumulation and net photosynthesis [
8]. Expanding leaf area is a commendable attribute to the overall growth rate of any crop leading to extensive interception of solar radiation and eventually contributing to better economic harvests [
9]. On the other hand, the radiation conversion efficiency of a crop into plant biomass equally depends upon the physiological characteristics of the crop [
10] as well as on environmental conditions [
11]. In this context, the leaf chlorophyll content of a plant is one of the fundamental attributing physiological characteristics related to photosynthetic capacity. Accelerated chlorophyll biosynthesis invariably leads to capturing more incoming solar radiation and a greater rate of net photosynthesis [
12]. Notably, RUE is also enhanced with the increase in PAR interception [
13]. However, improvement in RUE clearly indicates a higher rate of photosynthesis, which in turn contributes to better yield and nutrient use efficiency. In this context, Worku and Demisie [
3] observed around 88% correlation between dry matter production and RUE regarding pigeon pea. In addition, Jena et al. [
7] registered up to 4.12 g MJ
−1 RUE in mustard with increasing biomass production.
Grass pea (
Lathyrus sativus L.) is generally relay-cropped using the residual soil moisture in rice-fallow during
rabi season in India [
14]. Basically, it is a protein-rich pulse crop (28%) containing considerable proportions of several minerals like calcium, phosphorus and iron [
15]. It is considered as an ‘insurance crop’ as it produces reliable yields when all other crops fail due to a harsh environment. Compared to the other pulse crops, grass pea is a remarkable drought-tolerant crop that thrives with minimal external inputs and consequently is an ideal legume for resource-poor farmers [
16].
Seed priming is a recent technology to magnify the rate and synchrony of crop seeds germination, vigour and establishment of seedlings and subsequent attainments of biomass, yield attributing characters and yield of pulse crops [
17]. Nutrient seed priming can serve as a simple but effective agronomic practice to meet the nutrient demand of the crop in the early growth stages and eventually increase the final yield in case of relay sowing of pulse crops. In rice fallows, seed priming with KH
2PO
4 [
18], sodium molybdate [
19] has been earlier reported to increase grass pea production owing to accelerated crop growth and better uptake of nutrients from soil. Basically, molybdenum (Mo) is a vital micronutrient regulating different physiological and biochemical mechanisms in grain legumes [
20]. In particular, its direct involvement in the synthesis and activity of nitrogenase and nitrate reductase enzymes, regulating symbiotic N fixation and N assimilation by triggering rhizobial activity has been cited by earlier literature [
21]. Application of ammonium molybdate at a dose of 0.5 g kg
−1 seed has been observed to increase root nodulation of grass pea up to 80–90% along with up-gradation of economic yield to the tune of 30% [
14].
The foliar fertilization technique provides the crops plants with a quick supply of nutrients reaching directly to the site of photosynthesis without any wastage [
22]. Especially in indeterminate legumes, foliar application of nutrients is very much proficient as it provides sufficient time for conversion of late formed flowers into pods in addition to stimulation of balanced partitioning of photoassimilates from source to sink [
23]. Foliar feeding of urea and NPK (19:19:19) was found to be beneficial in the case of green gram, black gram, lentil, grass pea and chickpea [
24,
25] by delaying senescence and thereby facilitating photosynthesis. The positive influences of NPK foliar nutrition and their interactions are inevitably attributed to the indispensable role of nitrogen (N), phosphorus (P) and potassium (K) in the physiological development of plants [
15]. Application of N helps to expand leaf area as N is considered as the primary constituent of leaf chlorophyll maximizing the photosynthetic capacity and overall growth of crop plants [
26]. Generally, fertilization with N increases the vegetative growth, total carbohydrate, soluble sugars and NPK content of plants [
27]. Modulation of dry matter and protein contents in grain legume crops in terms of both qualitative and quantitative points of view through N application is a very well-known fact. Legume crops go through gradual leaf senescence well before their maturity, which obstructs the yield by breaking the normal source–sink relationship [
28]. This specific setback can be overcome through the foliar spray of nitrogen [
29], whereas P stimulates root, seed and fruit development along with aiding in vital metabolic functions of plants [
30]. In addition, P also departs energy in the form of ATP for nitrogen metabolism and hence enhances BNF, increasing rhizobial colonization, leaf area, photosynthesis, carbon partitioning and biomass accumulation [
31]. Phosphorus has a stimulating effect on the growth parameters, total carbohydrate, soluble sugars and minerals contents and influences the productivity by affecting the processes of energy storage and transfer [
32]. Potassium addition significantly stimulates root and shoot growth, and enhances the BNF and protein content of pulse grains [
33], besides regulating the water economy in the plant body through osmoregulation and maintenance of leaf water potential [
34]. Notably, Randhawa et al. [
35] reported an interception of PAR of around 460 MJ m
−2 along with maximum total dry matter and RUE using a nutrient management schedule consisting of NPK in terms of maize.
Indeed, there is a paucity of information regarding the impact of PAR interception and PAR use efficiency on grass pea production in the lower Gangetic plains of Eastern India. This study had been undertaken with the specific objectives of quantifying the amount of intercepted PAR and PAR use efficiency of winter grown grass pea as well as evaluating their interaction with the growth, physiology and seed yield of relay grass pea as influenced by seed priming with Mo and foliar nutrition with urea and NPK.
2. Materials and Methods
2.1. Location of the Study
The field experiment was pursued at the ‘A–B’ block, District Seed Farm (22°93′ N, 88°53′ E, 9.75 m above the mean sea level) of Bidhan Chandra Krishi Viswavidyalaya, Nadia, West Bengal, India during two subsequent rabi seasons (October–March) of 2017–2018 and 2018–2019.
2.2. Soil and Weather Conditions
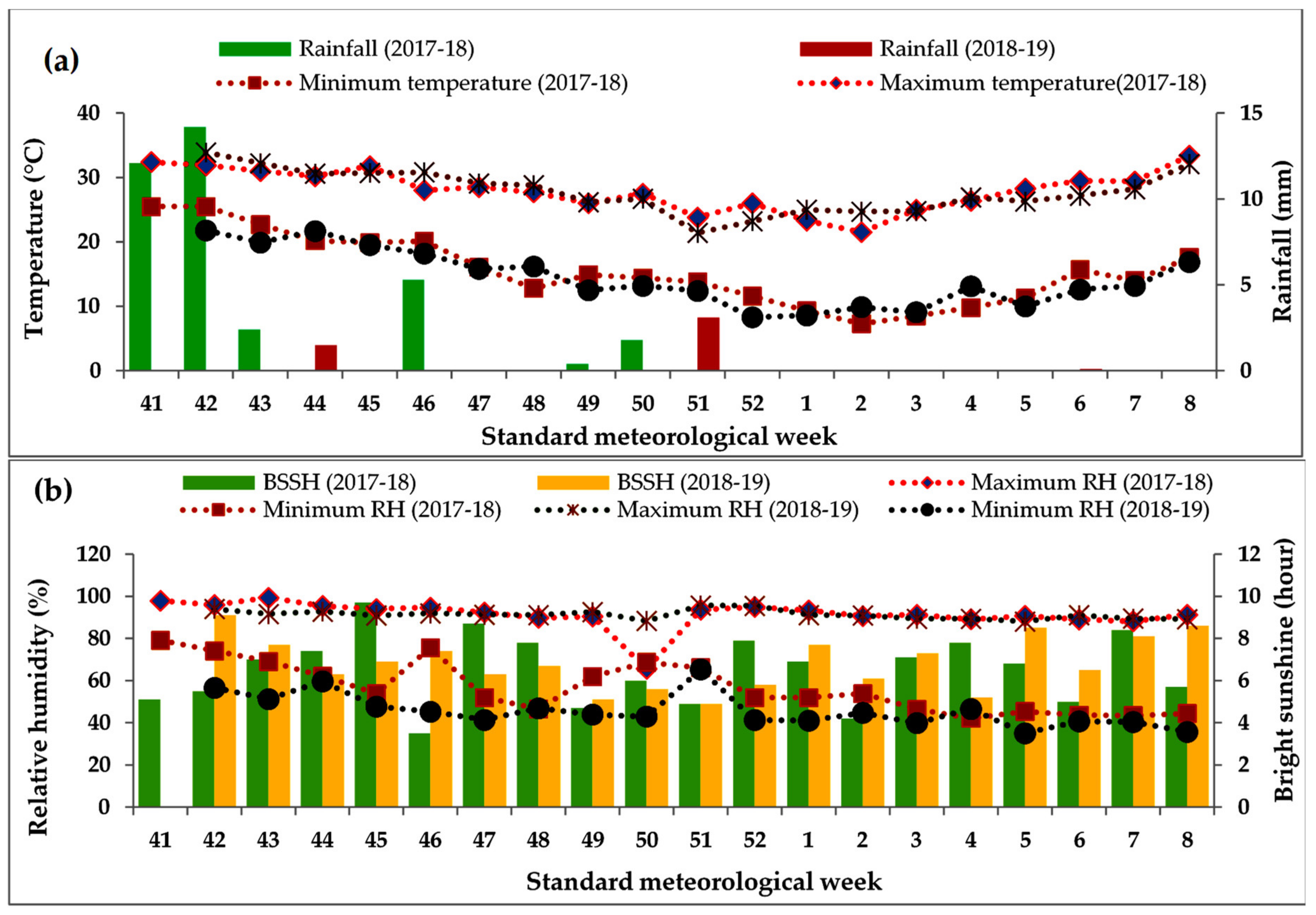
The soil of the study site was well-drained Gangetic alluvium (order: Inceptisol, suborder: Aqepts, great group: Haplaquepts) with moderate fertility and nearly neutral in reaction, categorised under the textural class of sandy loam with a neutral soil reaction. The detailed physicochemical properties of the soil of the research plots have been depicted in
Table 1. Meteorological features of the experimental site in both years have been presented graphically in
Figure 1.
2.3. Treatments and Design
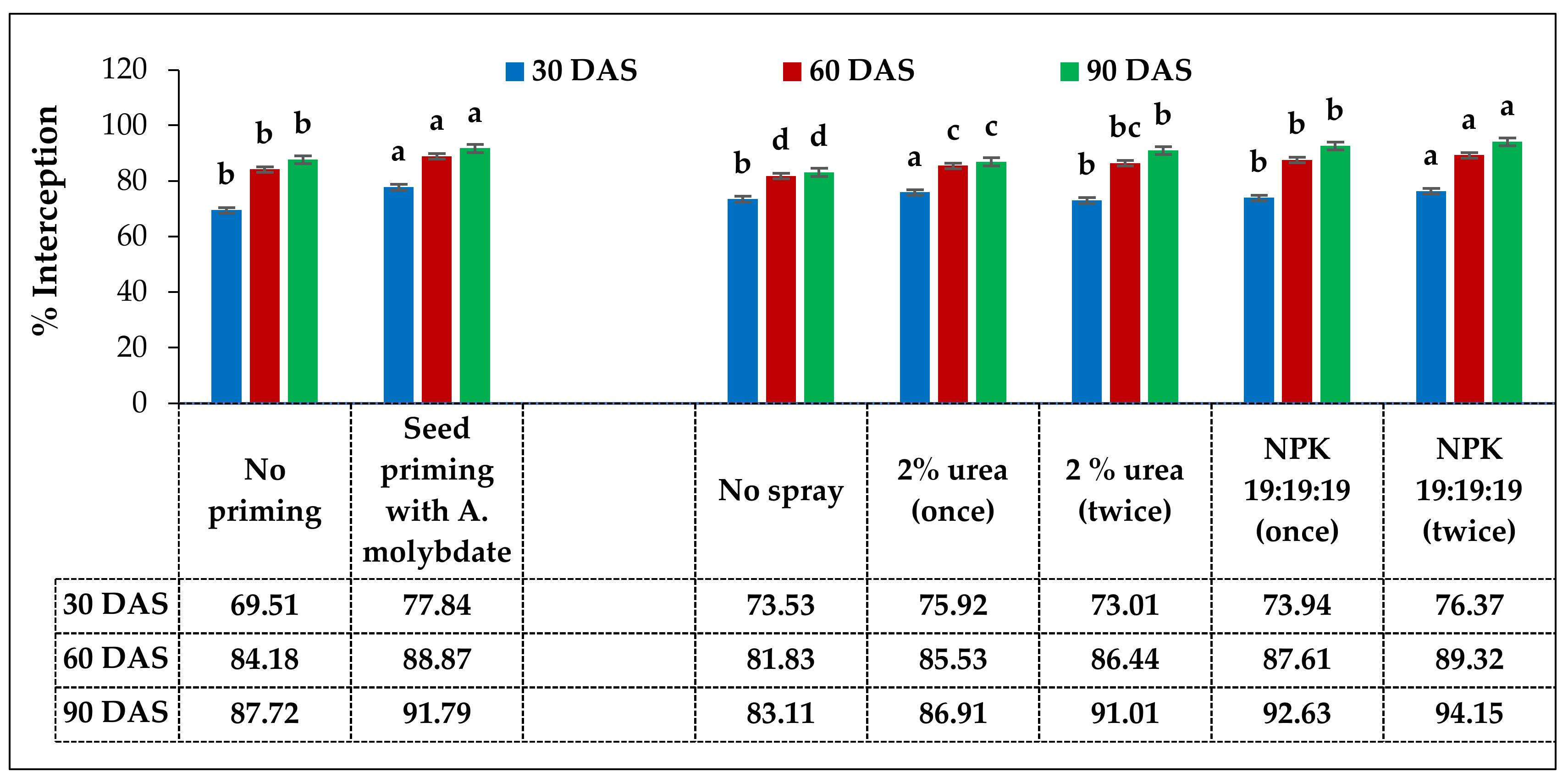
The field experiment was arranged in a factorial randomized block design consisting of 2 levels of the 1st factor (seed priming) and 5 levels of the 2nd factor (foliar sprays) in various combinations with a total of 10 treatments replicated thrice. Grass pea seeds of the variety Ratan (Bio L-212) were used for the whole experiment. Detailed treatments are presented in
Table 2.
2.4. Experimental Procedures
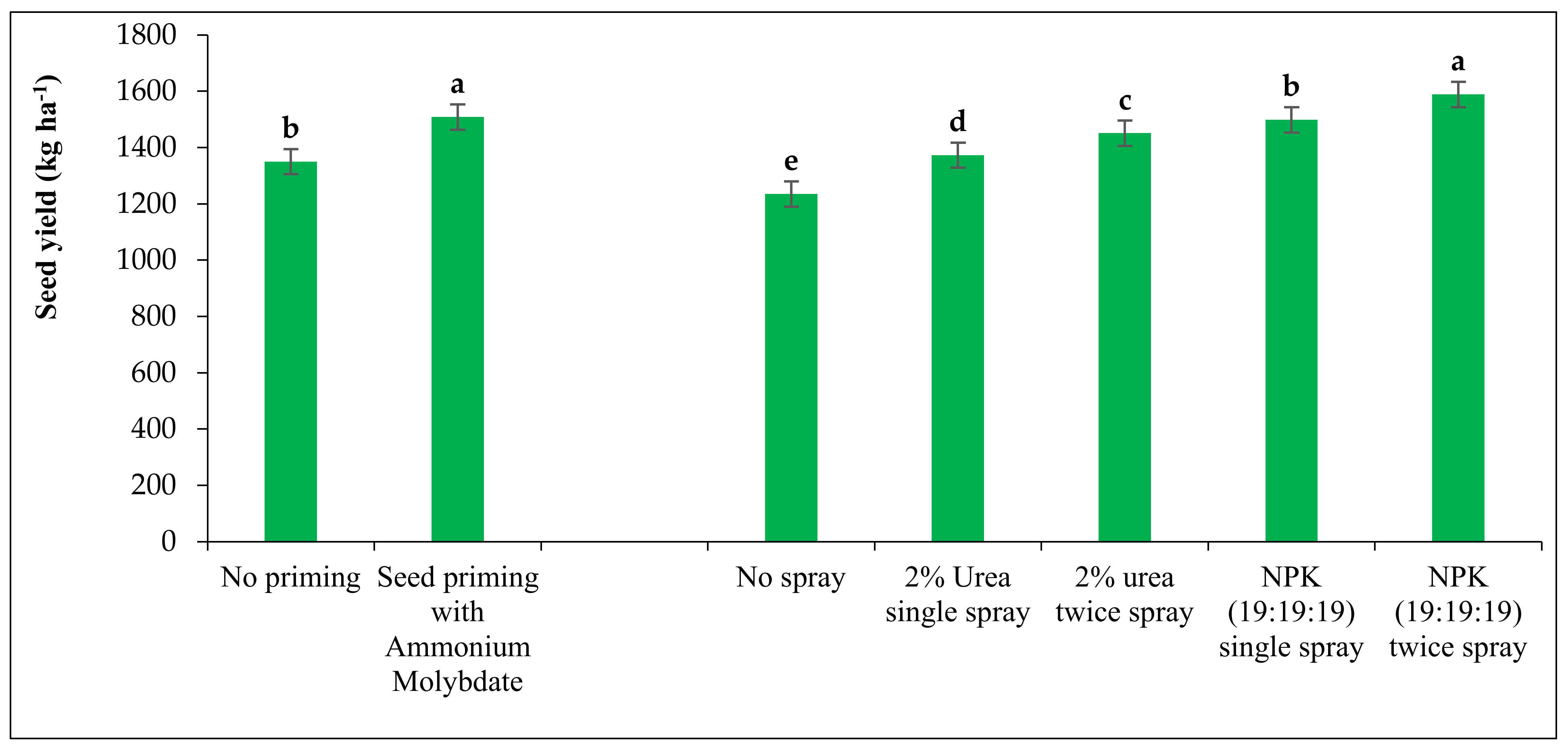
The event of land preparation was completely excluded for relay grass pea crop in this experiment. Generally, grass pea crop requires a seed rate of 40 kg ha−1 for line sowing. However, the seeds were sown at the rate of 80 kg ha−1 in individual experimental plots of 5 m × 3 m through broadcasting on a standing rice crop as per the recommended practices of relay cropping. Half of the seeds were primed with ammonium molybdate at the rate of 0.5 g kg−1 of seed for 8 h followed by shade dry and the rest were kept dry on the day before sowing. Before an hour of sowing, all the seeds were treated with Rhizobium biofertilizer at the rate of 20 g kg−1 of seed for better nodulation. Basal dose of fertilizers application as well as irrigation were completely excluded in case of cultivation of relay grass pea.
One manual weeding was done at 25–30 days after sowing for proper stand establishment of the crop. Foliar sprays with 2% Urea and 0.5% NPK (19:19:19) were done as per the treatment wise allotments in the morning hours spraying with the help of a knapsack sprayer by one labourer simply walking along with the individual plots. The exact amounts per plot requirements of fertilizers were calculated as per the treatment schedule and the same was mixed with the tap water (at the rate of 500 lit ha−1) inside the spray tank for better accuracy of the dose. Spraying of fungicide including SAAF (Mancozeb + Carbendazim) @ 2.5 g lit−1 of water was done at 60 DAS as a plant protection measure.
2.5. Data and Their Estimation Procedures
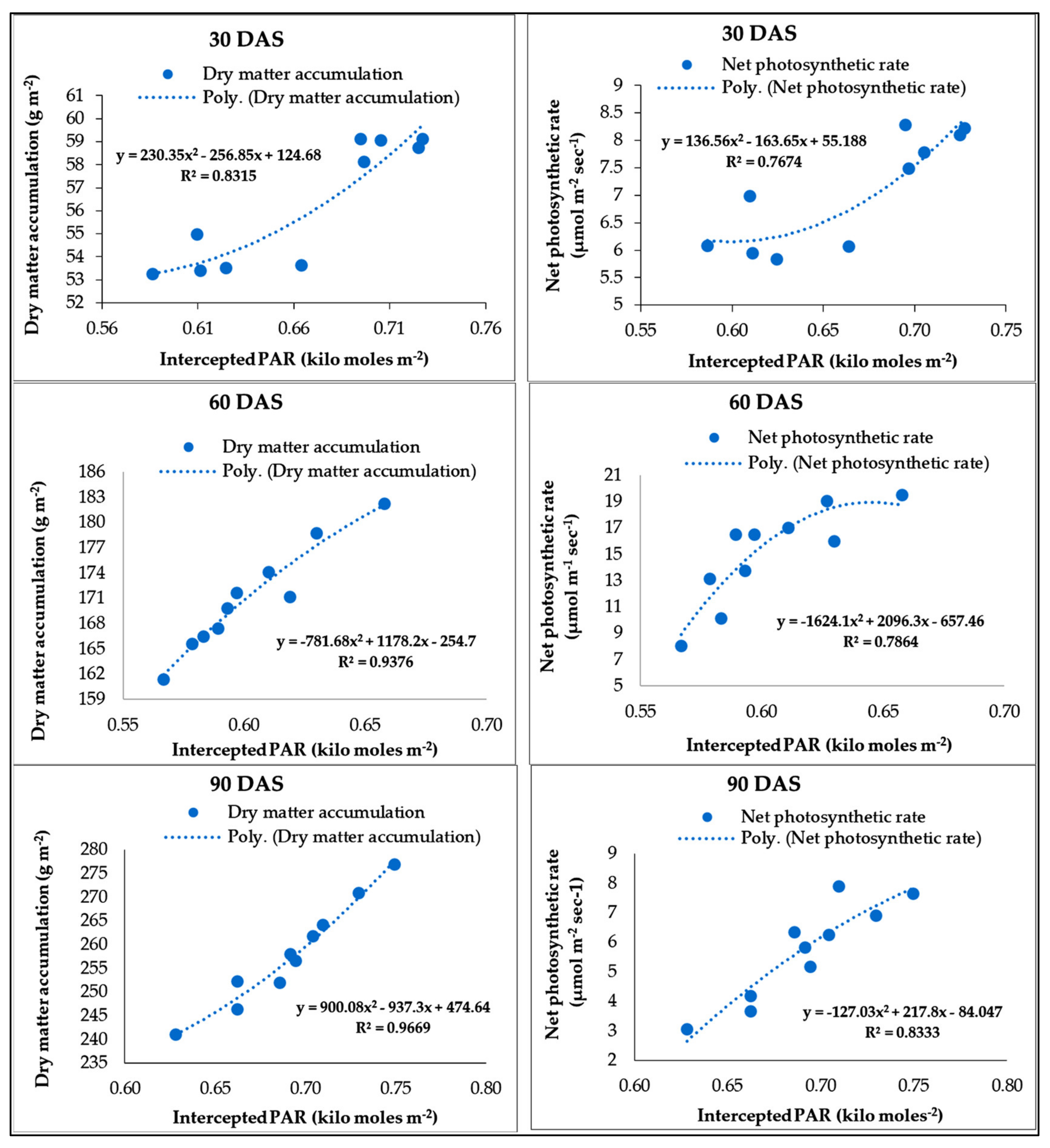
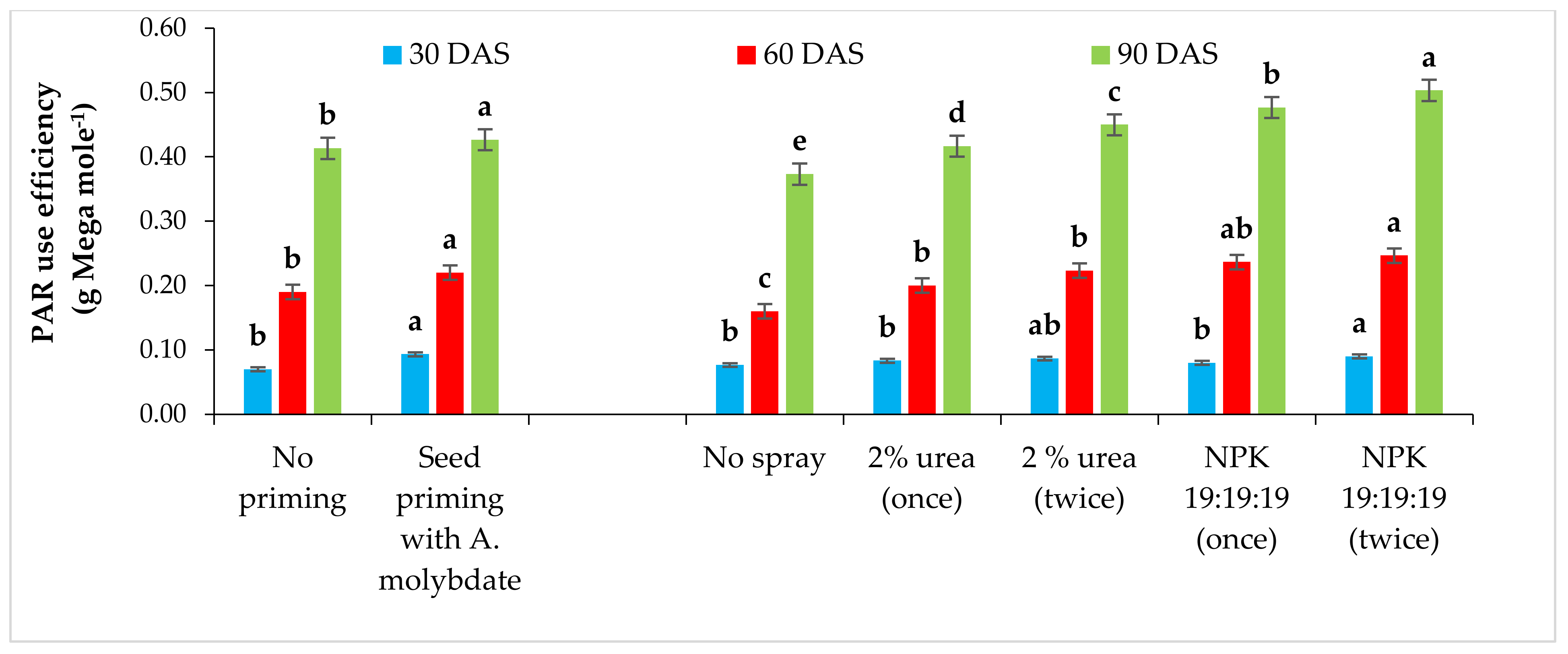
The observations of PAR were measured starting from vegetative (15–45 DAS) up to the pod filling stage (75–105 DAS) at 11.30 h at 30 days intervals using Line quantum sensor (APOGEE Logan UT). The instrument was placed 25 cm above the crop across the rows to estimate incident radiation. Then, it was kept horizontally under the canopy and placed likewise 25 cm higher the soil surface to measure the transmitted radiation from the bottom of the canopy. The reflected PAR was measured from the same position by simply inverting the sensor. Intercepted PAR (I PAR) and PAR use efficiency (PARUE) were calculated following Equations (1) and (2) [
44]:
where
= incident PAR above the canopy, T PAR = transmitted PAR through the canopy towards the soil surface, and R
= reflected PAR from the canopy
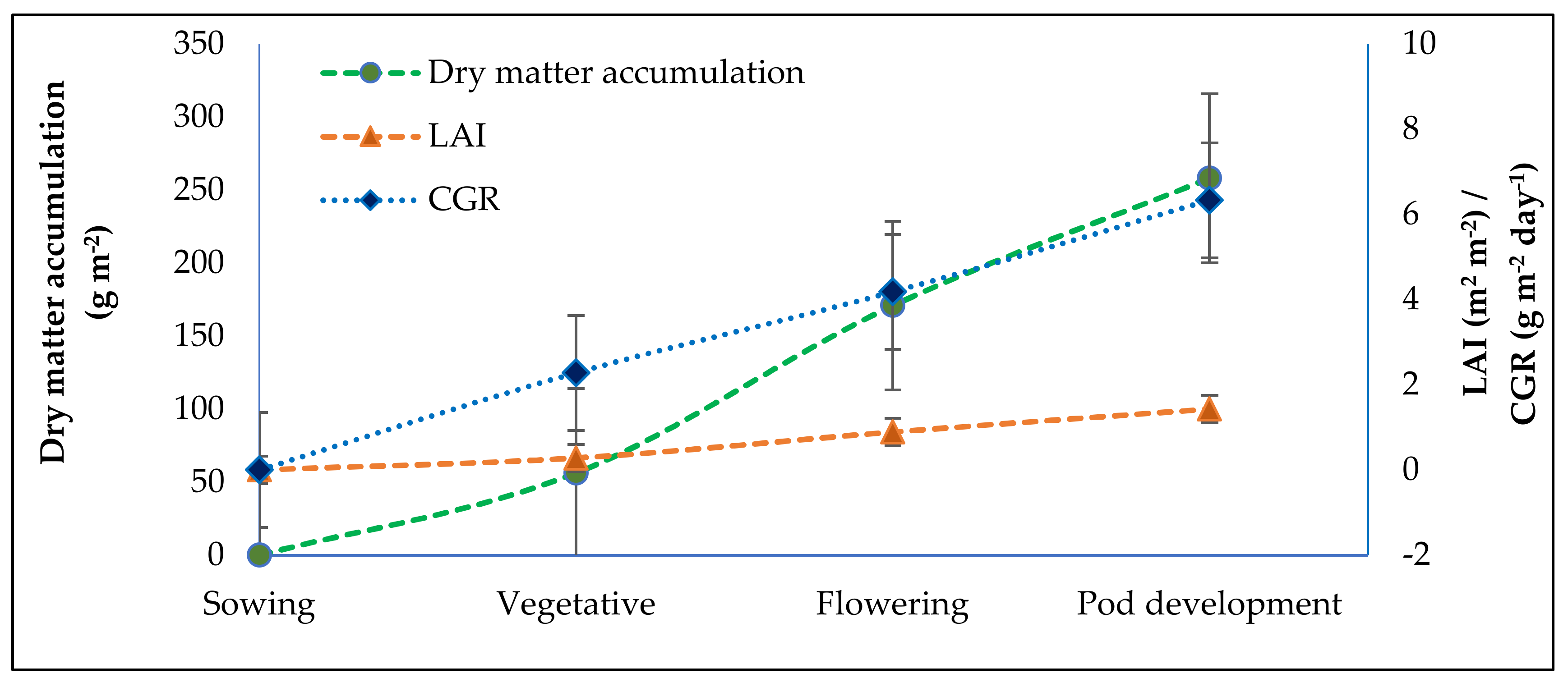
For taking observations of growth attributes of grass pea, 20 plants were tagged through random selection excluding the border rows from each plot. For growth analysis, dry matter accumulation, crop growth rate (CGR), and leaf area index (LAI) of grass pea crop was worked out at vegetative (30 DAS), flowering (60 DAS) and pod filling stage (90 DAS) from 10 randomly selected plants.
LAI was computed following the expression [
45]:
CGR was estimated using the following formula of Watson [
45] and expressed in g m
−2 day
−1:
where
= total dry weight of plant at time
,
= total dry weight of plant at time
and G = ground area.
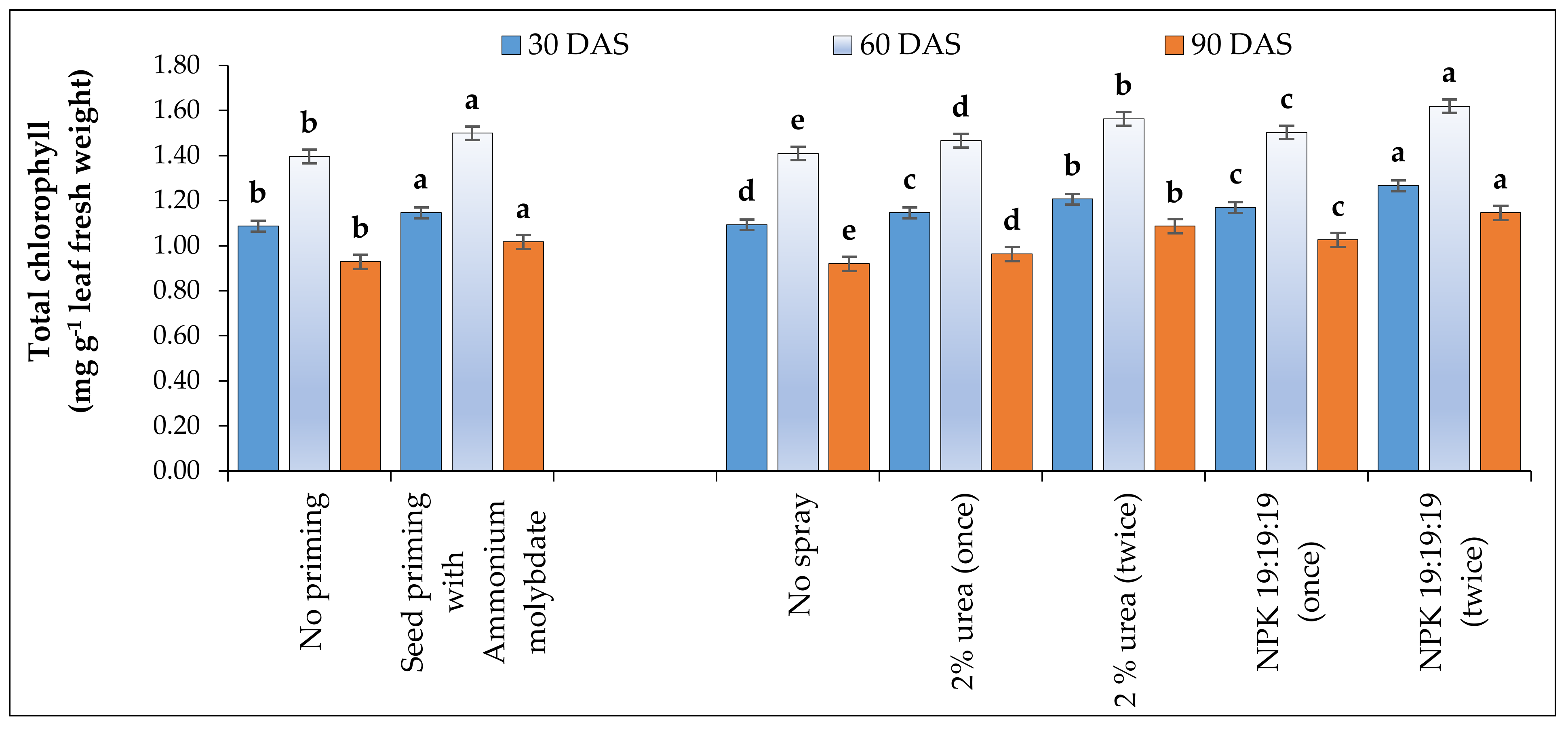
The leaf chlorophyll contents were estimated at 30, 60 and 90 DAS. It was measured by taking absorbance readings at 480, 510, 645 and 663 nm wavelengths against a blank one with only 80% acetone in a Systronics-105 spectrophotometer. The chlorophyll a and b, total chlorophyll and carotenoid were estimated with the following formula given by Arnon [
46], all expressed in mg g
−1 of fresh leaf weight:
where V = Extract volume (mL), W = Fresh weight of leaf tissue (g), and A = Absorbance.
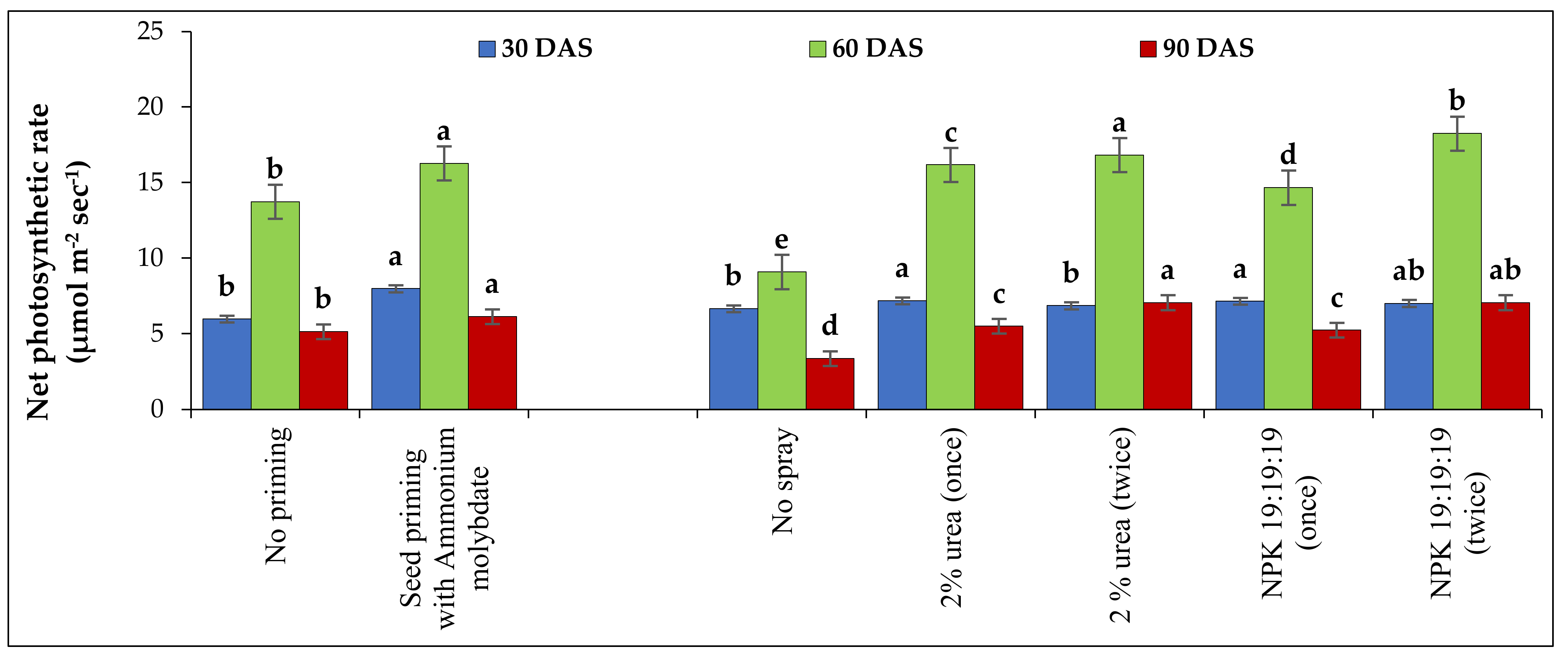
The net photosynthetic rate of grass pea leaves was measured with a portable handheld photosynthesis system (CI-340 Handheld Photosynthesis system, CID Bio-Science, Inc. Camas, WA, USA) and expressed in µmol m−2 s−1. The measurements were obtained on clear sunny days from the fully developed upper leaves of five selected plants from 11:30 a.m. to 12.30 p.m. at 30, 60 and 90 DAS.
The available nitrogen, phosphorous and potassium content in grass pea leaves were determined respectively by the modified Kjeldahl method [
39], Olsen’s method [
40] and flame photometer method [
39].
2.6. Statistical Analysis
Data were statistically analysed by implementing the analysis of variance (ANOVA) techniques proposed by Gomez and Gomez [
47] for factorial randomized block design. Pooled analysis was exercised in case of similar data from both years. Treatment means were compared by employing the F-test. The significant differences between the treatments were compared by a critical difference at a 5% level of significance. The regression analysis was carried out by SPSS 7.5 software, (SPSS 7.5 copyright, 1997 by SPSS Inc., USA Base 7.5 Application guide). Tukey’s posthoc test was performed to compare the differences between mean values.


 ,
, 










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}