1. Introduction
Weeds are a major production constraint in farming systems worldwide. Weed losses are especially high in small and meso-scale vegetable and small fruit production that rely heavily on labor for weed management. In small-holder agriculture the burden of arduous weed management often constrains the health, income, and advancement of women and children. Weed control options are also limited in organic production systems and farmers consider weeds to be the most significant barrier to organic farming [
1,
2]. Therefore, alternative environmentally sustainable weed management options are needed for organic farmers and others seeking sustainable approaches for crop production
Anaerobic soil disinfestation (ASD), also known as reductive soil disinfestation or biological soil disinfestation, was initially developed for soilborne disease management; however, recent reports suggest that it might be effective in controlling weeds [
3]. Soil treatment using ASD requires the incorporation of easily decomposable carbon sources in soil, irrigation to saturation, and covering with plastic sheeting for longer than three weeks to create anaerobic conditions [
4]. During the first stage of ASD, the growth of aerobic microorganisms is stimulated by the metabolization of labile carbon sources incorporated into the soil, resulting in the rapid decline of oxygen levels and subsequent reduction in soil redox potential [
4,
5,
6]. Under these anaerobic conditions, poorly oxidized compounds such as hydrogen sulfide, methane and ethylene gases, alcohol and fatty acids (acetic, butyric, and propionic acids) are produced by soil microbes [
7]. These compounds can be toxic to weeds, pathogens, and nematode propagules present in the soil [
7,
8].
Several studies have shown that ASD can inactivate weed seeds and tubers. For example, yellow nutsedge (
Cyperus esculentus) tuber germination and reproduction have been reduced in ASD-treated soil [
9]. Germination of seeds of other species has been suppressed in ASD-treated soil, but results have been variable [
10]. The effectiveness of ASD against weed seeds depends on soil temperature, duration of incubation, rate of field application, and type of carbon source. The mechanism of action of ASD for weed germination suppression is not well understood. However, potential mechanisms are associated with the abrupt creation of hypoxic conditions [
11] and reduction of soil pH due to accumulation of organic acids during anaerobic decomposition of carbon sources [
5,
6]. Increased soil temperature due to exothermic reactions during carbon source breakdown [
12] could also affect the viability of weed seeds before or during germination.
Seed survival in soil under extreme environmental conditions differs among weed species [
13]. Some weed seeds and vegetative propagules remain viable for a long time while others survive just for a short period in soil. Various weed species are likely to respond to ASD in different ways. Previous studies on weed management using ASD have focused on one or two species [
3,
9]. However, farmers must deal with several weed species at the same time. Therefore, understanding the influence of ASD on a wide range of weed species is needed to provide a practical perspective for application to farm situations.
The effectiveness of ASD depends on characteristics of the carbon sources and their availability to microbial populations [
6,
14]. The nature and rate of carbon sources used in ASD likely play significant roles in the ability of ASD to kill weed propagules in soil. Therefore, understanding the effect of different carbon sources and their rates of field application on different types of weed species is needed to make effective use of ASD for weed management. This study was designed to evaluate commonly available carbon sources and rates of field application to suppress weed species varying in propagule characteristics and survivability in soil.
2. Materials and Methods
2.1. Weed Species
We studied seven weed species that are commonly problematic for smallholder farmers in many countries and can be used as model species to study ASD effectiveness: common lambsquarters (Chenopodium album), barnyardgrass (Echinochloa crus-galli), black nightshade (Solanum nigrum), yellow nutsedge (Cyperus esculentus), common pokeweed (Phytolacca decandra), dandelion [Taraxacum officinale], and redroot pigweed (Amaranthus retroflexus).
The seeds of S. nigrum were obtained from the Department of Horticulture and Crop Science, The Ohio State University (OSU), C. esculentus tubers were purchased from Azlin Seed Service (Leland, Mississippi) and the remaining weed seeds except E. crus-galli were collected from gardens on the OSU College of Food, Agricultural and Environmental (CFAES) Wooster Campus. The soil used during the experiments was highly infested with E. crus-galli seeds and required no further infestation.
2.2. Soils
The soil used in all greenhouse and growth chamber experiments was collected from a certified organic field on Badger Farm on the OSU CFAES Wooster Campus near Wooster, OH in November 2016. The soil was dried and pulverized, then homogenized in a cement mixer, and screened through 1.0 cm mesh before storage at 10 °C in plastic boxes until use. Soil chemical properties presented in
Table 1 were determined using standard procedures at Spectrum Analytic Inc., Washington Courthouse, OH, USA.
2.3. Baseline Germination Test
Weed seed germination percentage was estimated by using a blotter test. Seeds were surface sterilized with 1% sodium hypochlorite solution for 5 min followed by washing five times with sterile deionized distilled water. Fifty-one seeds per species were plated in 85 mm plastic Petri dishes containing premoistened sterilized blue blotter papers (Seedburo Equipment Co., Des Plaines, IL, USA). The Petri dishes were then incubated at 25 °C under an 8 h light, 16 h dark cycle. Seeds of each species were replicated three times in three plates. Germinated seeds were periodically counted until 28 days after plating. Then, 5 mL of 0.2% KNO3 solution was added to each Petri plate to induce the germination of the remaining seeds. The seeds were then maintained in an incubator at the same light and temperature regime for one additional week. The germination percentage of C. esculentus tubers was determined by planting 16 tubers in a plastic tray containing a soil mix [6 parts sandy soil, 1 part peat, 1.8 g L−1 lime, 0.45 g L−1 fertilizer (20:20:20, N-P-K fertilizer, Peter’s Professional, ICL Fertilizers, Ltd., Beer Sheva, Israel)].
Seed germination was below 85% for P. decandra, A. retroflexus, and S. nigrum during a baseline germination test, so these seeds were treated to break dormancy according to standard procedures. Seeds of P. decandra were treated with concentrated H2SO4 for 5 min followed by washing in running deionized water over a 50 mesh screen for 5 min before plating in Petri dishes. Seeds of S. nigrum and A. retroflexus were refrigerated at 4 °C for three weeks before plating.
2.4. Carbon Sources
The four carbon sources tested were wheat bran (The Mennel Milling Company, Fostoria, OH, USA), molasses (Golden Barrel blackstrap molasses, Good Food, Inc., Honey Brook, PA, USA), raw chicken manure (collected from the Poultry Research Farm, OSU CFAES Wooster Campus, Wooster, OH, USA), and mustard greens (Brassica juncea L.) biomass.
Mustard greens ‘Southern Giant Curled’ seeds (Thiram® treated, Seedway, Hall, NY, USA) were sown in 50-cell plug trays containing Baccto Professional Grower Mix (Houston, TX, USA) and grown for 30–40 days under greenhouse conditions programmed at 25 °C and 14 h light. Plants were fertilized once a week with liquid fertilizer (20-20-20 N-P-K fertilizer, Peter’s Professional, ICL Fertilizers, Ltd., Beer Sheva, Israel). Plants were uprooted and washed then entire plants were chopped and macerated in a blender (Waring Commercial Blender, Waring Commercial, Torrington, CT, USA) before mixing with soil. Fresh chicken manure was collected from the OSU Poultry Research Center, Wooster and stored at 10 °C in buckets until use. Wheat bran, chicken manure, and mustard greens biomass were mixed with soil before placement in the pots. Molasses was mixed with an equal volume of water and poured onto soil in pots. These carbon sources were selected considering the ratio of carbon to nitrogen (C:N), i.e., C:N ratio of wheat bran = 17, chicken manure = 9.6, molasses = 81, and mustard greens biomass = 12, as well as their availability in a diversity of settings and previous results in ASD trials.
2.5. ASD Treatment
Three experiments were conducted to evaluate the effect of ASD with different carbon sources on weed seed germination in growth chambers and a greenhouse. A fourth experiment was conducted under field conditions. For all the experiments, sixteen
C. esculentus tubers weighing 0.1 to 0.2 g each or 150 seeds of other species were thoroughly mixed with 50 mL of dry soil and placed in a nylon mesh packet, one species per packet. Growth chamber/greenhouse experiments were conducted in pots filled with the field soil described above. ASD treatment was initiated by saturating soil with tap water. Pots were allowed to drain for one hour and then enclosed either with plastic sheeting or in 3.78-L Ziploc
® plastic bags (SC Johnson Family Company, Racine, WI, USA). The pots were then incubated in a growth chamber for 21 days with a 12 h day/night cycle; and temperature was maintained at 30 °C/day and 26 °C/night. Soil reducing conditions in each pot were measured based on the depletion of iron oxide paint from IRIS (Indicator of Reduction in Soils) tubes [
16], which were placed in soil in each pot through a guide hole before irrigation. After removal from soil after ASD treatment the percentage of paint loss from the IRIS tubes was estimated visually using a grid [
17]. Germination percentage was determined for weed seeds or tubers recovered from packets buried in the soil. The weed seed packets were washed in tap water to remove the soil followed by soaking in a 1% solution of sodium hypochlorite for 5 min and finally rinsing two times in deionized distilled water. Seeds/tubers were plated on blotter paper (seeds) or planted (tubers) as described above to monitor germination. After retrieval of weed seed packets, soil in each pot was stirred with spatula and pots were kept in the greenhouse for 45 days to induce the germination of resident weed seeds. Irrigation with tap water was done once a day. Liquid fertilizer (20-20-20 N-P-K fertilizer) was applied to pots once weekly. The number of seedlings was counted at 10-day intervals after 20 days of incubation, for 45 days. The percentage of seed mortality was calculated by [(number of seeds plated/number of seeds germinated) × (100/number of seeds plated)].
Soil samples (~100 g) were collected immediately after plastic removal and soil pH was measured in 1:1 slurry with water using a Thermo Scientific Benchtop pH Meter (Thermo Scientific, Agawam, MA, USA). Gravimetric soil moisture (GSM) percentage was measured by drying 100 g of soil from each pot at 60 °C for 96 h and measuring final weight. The gravimetric moisture percentage was calculated as the difference between the final and initial soil weight. All the experiments were conducted two times independently. This ASD treatment procedure, seed viability monitoring, soil pH and gravimetric soil moisture monitoring process were used for all growth chamber and greenhouse experiments as described in
Section 2.6,
Section 2.7 and
Section 2.8.
2.6. Effect of Carbon Source and Burial Depth
This experiment was conducted to evaluate the impact of carbon sources and depth of seed/tuber placement in germination of different weed seeds/tubers under ASD treatments. The experiment was conducted using a completely randomized design (CDR) with a split-plot arrangement in 17 × 17 × 17 cm plastic pots in growth chambers with three replications. Seed or tuber burial depth (5 cm and 15 cm) was the main plot factor, and ASD carbon source (wheat bran, molasses, chicken manure, and mustard greens) was the sub-plot factor. All carbon sources were added at a rate of 37 g kg−1 soil (equivalent to 72 Mg ha−1). Mustard greens biomass was used on a fresh weight basis. Controls included were non-amended covered (anaerobic control) and non-amended non-covered (aerobic control). Mesh packets containing C. esculentus tubers or other weed seeds were buried in the soil at depths of 5 cm or 15 cm from the surface of the soil. The top and bottom of each pot were sealed individually using a doubled black plastic sheet (1.5 mm embossed, black plastic mulch, PolyExpert, Inc., Laval, QC, Canada) and made airtight using string and electrical tape. Soil pH, reducing conditions and gravimetric moisture content, and germination of artificially inoculated and resident weed seeds were recorded as mentioned previously.
2.7. Effect of Rate of Mustard Greens Biomass
This experiment was conducted to evaluate the impact of different rates of mustard greens biomass in suppression of weed seed/tuber germination under ASD treatment. This experiment was conducted using a randomized complete block design (RCBD) with three replications in 11 × 11 × 11 cm plastic pots for 21 days. Rates of mustard greens biomass evaluated were 5, 10, 15, and 20 g dry matter per kg soil (equivalent to 10.1, 20.2, 30.3, and 40.4 Mg ha−1, respectively). The dry weight of the mustard greens was determined by measuring the fresh weight of three 100 g samples of 30–40 day-old plants, then drying them at 60 °C for 72 h. The average fresh and dry weights of the three samples was used to calculate the dry weight percentage of mustard greens. Fresh plant biomass was used in the experiment, and the amount added was uniform based on dry matter percentage. Aerobic and anaerobic controls were included. Pots (except the aerobic control) were double-sealed with two 3.78 L Ziploc bags. Soil pH, reducing conditions, and gravimetric moisture content, and germination of artificially inoculated and resident weed seeds were recorded as mentioned previously.
2.8. Effect of Rates of Wheat Bran, Molasses, and Chicken Manure
This experiment was conducted to evaluate the impact of four rates of wheat bran, molasses, and chicken manure as ASD amendments on weed seed or tuber mortality. The experiment was conducted in RCBD using 11 × 11 × 11 cm plastic pots with three replications of 11 treatments, including aerobic and anaerobic controls without added carbon sources. Soils were amended with wheat bran at rates of 5, 10, and 15 g kg−1 soil (equivalent to 10.1, 20.2, and 30.3 Mg ha−1, respectively); and chicken manure and molasses at rates of 5, 10, and 20 g kg−1 soil (equivalent to 10.1, 20.2, and 40.4 Mg ha−1, respectively). Aerobic and anaerobic controls were maintained as described previously and pots were double sealed with two 3.78-L Ziploc bags. Soil pH, reducing conditions, and gravimetric moisture content and germination of artificially inoculated seeds and resident weed seeds were determined as described above.
2.9. Field Evaluation
Field experiments were conducted at the OSU CFAES Muck Crops Agricultural Research Station in Huron County, OH on Linwood muck soil, pH 5.8, to examine the field efficacy of ASD with selected carbon sources for weed seed mortality. The experiments were conducted using a Latin square design with five treatments: wheat bran 20.2 Mg ha−1, molasses 20.2 Mg ha−1, a combination of wheat bran (20.2 Mg ha−1) plus molasses (10.1 Mg ha−1), and aerobic and anaerobic controls. The plot size in the first iteration of the experiment was 0.91 m by 3 m and in the second iteration it was 1 m by 1 m. Buffer zones were 3 m within the rows and 0.91 m between the rows in the first experiment. In the second experiment, the buffer zones were 1 m within and between the rows. Wheat bran was spread over the plots by hand, followed by incorporation at 15–20 cm depth with a tractor-mounted rototiller in the first experiment and by hand raking in the second experiment. Molasses was diluted 1:4 with water and applied using watering cans. Mesh packets, each containing 16 tubers of C. esculentus, or 100 seeds of C. album, A. retroflexus, or S. nigrum plus 2–3 g muck soil were buried at 15 cm depth before irrigation. Thirty cm long × 1.27 cm diameter IRIS tubes were placed in each plot after making a guide hole with the same size of PVC pipe before covering the plots with plastic. Soils were saturated with water applied by overhead sprinkler irrigation and each plot was covered individually with 1.5 mm embossed black plastic mulch for 4 weeks. Margins of the plastic were tightly sealed to control the exchange of gases by depositing extra soil along the edges of the plastic. Aerobic and anaerobic controls were not amended with a carbon source and the aerobic control was also not covered. After removal of plastic mulch, weed seed germination tests and soil reducing were determined as mentioned above.
2.10. Effect of Organic Acids on Weed Seed Germination
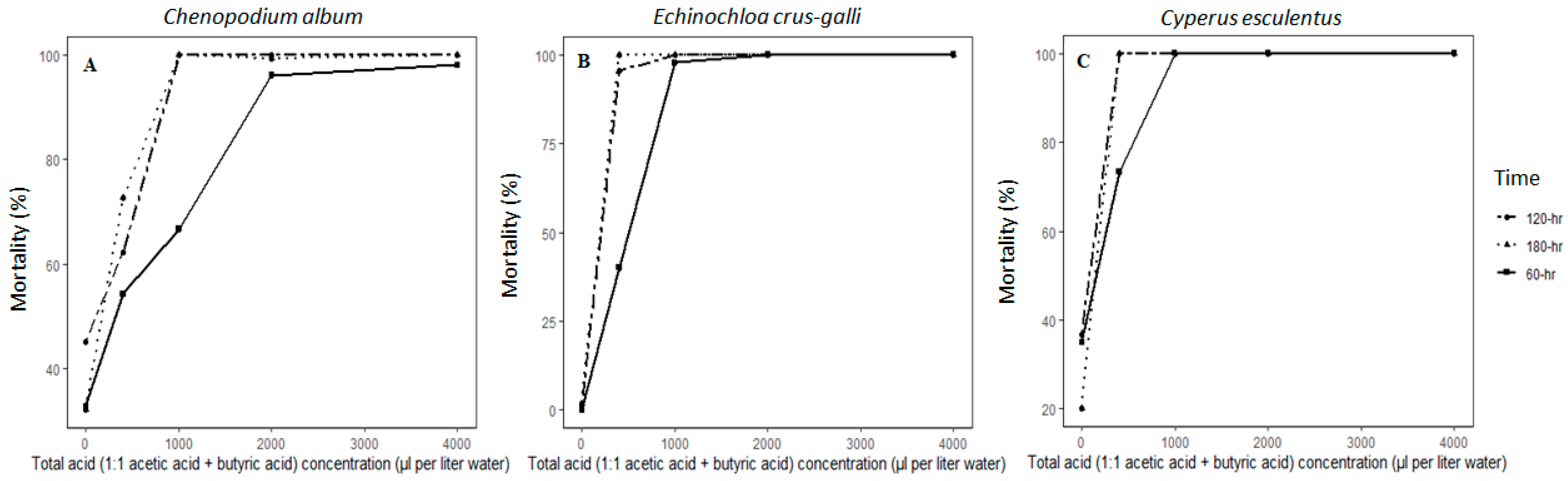
To test organic acids produced during ASD as possible causes of seed mortality, we evaluated seeds of C. album and E. crus-galli and tubers of C. esculentus, exposed to five concentrations of acetic plus butyric acid for three-time durations in a nested factorial design with three replications. The first factor was combined acid concentrations of 0, 400, 1000, 2000 and 4000 μL L−1 and second factor was duration of exposure: 60, 120 and 180 h. Organic acid solutions were prepared by mixing equal amounts of butyric and acetic acids for each concentration. Nylon mesh packets containing 100 seeds or 30 tubers were submerged in 50 mL acid solution in 100 mL plastic bottles.
Nylon mesh packets were retrieved from the acid solutions and distilled water. Seeds were plated in plastic Petri dishes containing premoistened blotter paper, and C. esculentus tubers were planted in soil mix (six parts sandy soil: one part peat; 1.8 g L−1 lime; 0.45 g L−1 fertilizer; 20:20:20) in 50 plug plastic trays (cm3 cell−1) and propagule germination was monitored as described above.
2.11. Data Analysis
Data were subjected to the Shapiro–Wilk test followed by the Bartlett test for normality of residuals and homogeneity for variance, respectively, before proceeding to analysis of variance (ANOVA). Data that deviated from the normal distribution were square root transformed prior to ANOVA. Data analyses were conducted using the linear model function “lm” in RStudio (R version 3.2.5) [
18]. In the case of a significant difference between the treatment means, Fisher’s test of least significant difference (LSD) was derived from the
agricolae package [
19]. The regression analysis of carbon source rates in ASD and weed seed mortality were conducted using the linear model function (lm) in
agricolae in R-studio.
4. Discussion
Anaerobic soil disinfestation (ASD) has potential to contribute to weed management in settings where carbon sources and means to impose anaerobic conditions are available. Like all weed management methods, ASD was more effective for some species than others, and clear trends based on seed characteristics are challenging to determine. The weed species tested in this study represent a broad range of species types in terms of plant physio-morphology and seed characteristics. Results indicate that ASD tends to be more lethal to larger seeds than smaller seeds. We observed high seed mortality in
P. decandra, and
T. officinale, the two perennial species, and germination inhibition of resident
E. crus-galli seeds was higher in ASD-treated soil than for small-seeded weeds such as
A. retroflexus,
S. nigrum and
C. album. Seeds of
A. retroflexus were the smallest tested and were the most resilient to ASD. The reason is unknown, but previous reports, e.g., [
21] indicated that larger seeds absorb moisture faster than small seeds, which means that ASD metabolites including organic/fatty acids enter into the larger seeds faster than smaller seeds, resulting in greater sensitivity of larger seeds to ASD than smaller seeds.
Seed mortality after exposure to ASD varied within the grasses and broadleaf species, depending on the carbon source used. Cyperus esculentus was more sensitive to ASD than E. crus-galli, while C. album and S. nigrum were more sensitive than A. retroflexus. Interestingly, P. decandra and T. officinale were also highly sensitive to flooded, moderately hypoxic conditions in the non-amended anaerobic control. The soil used in the experiments was collected from an organic farm with organic matter content of 1.7%, which could have acted as a natural source of carbon in the anaerobic control, resulting in hypoxic conditions and high seed mortality for P. decandra and T. officinale.
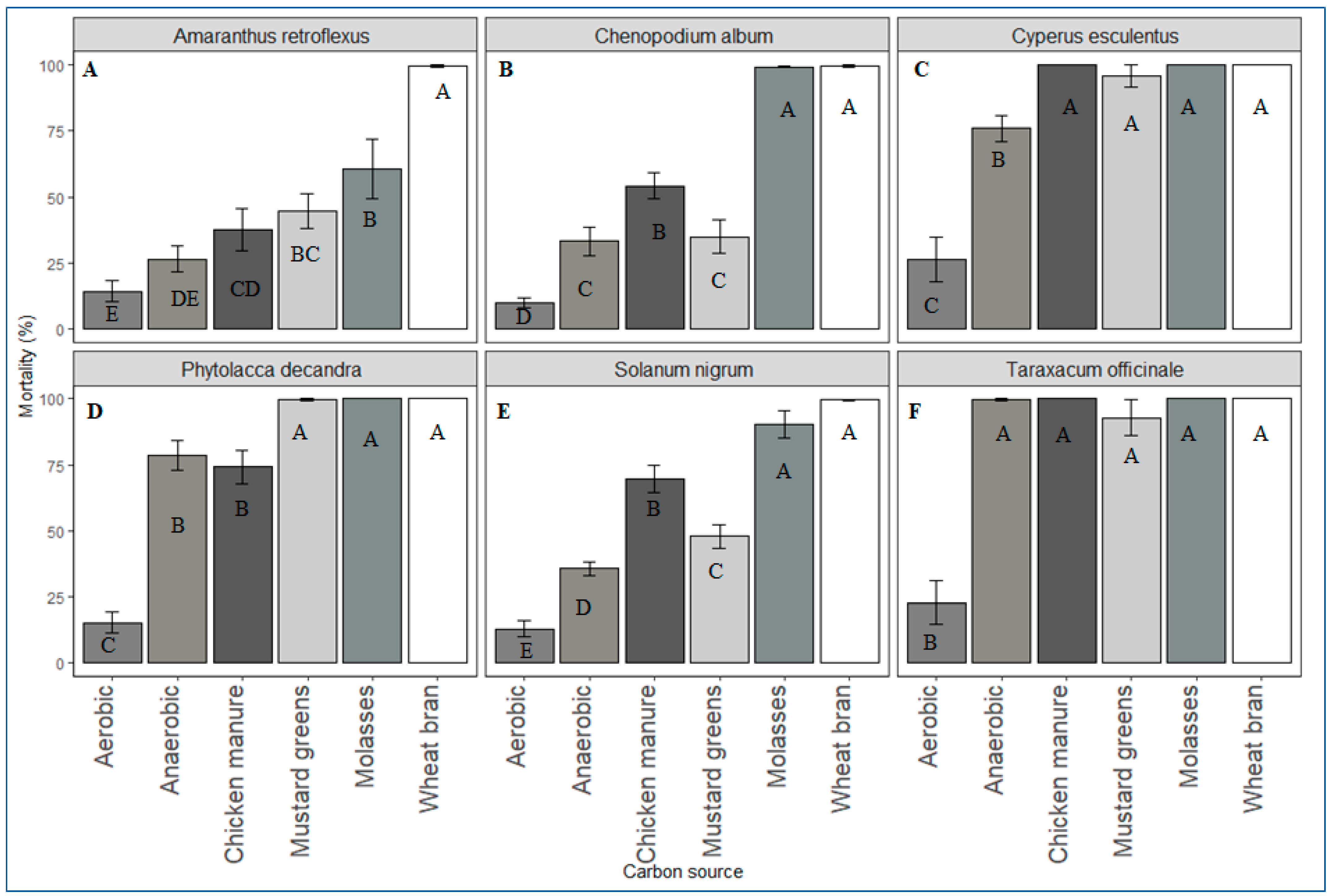
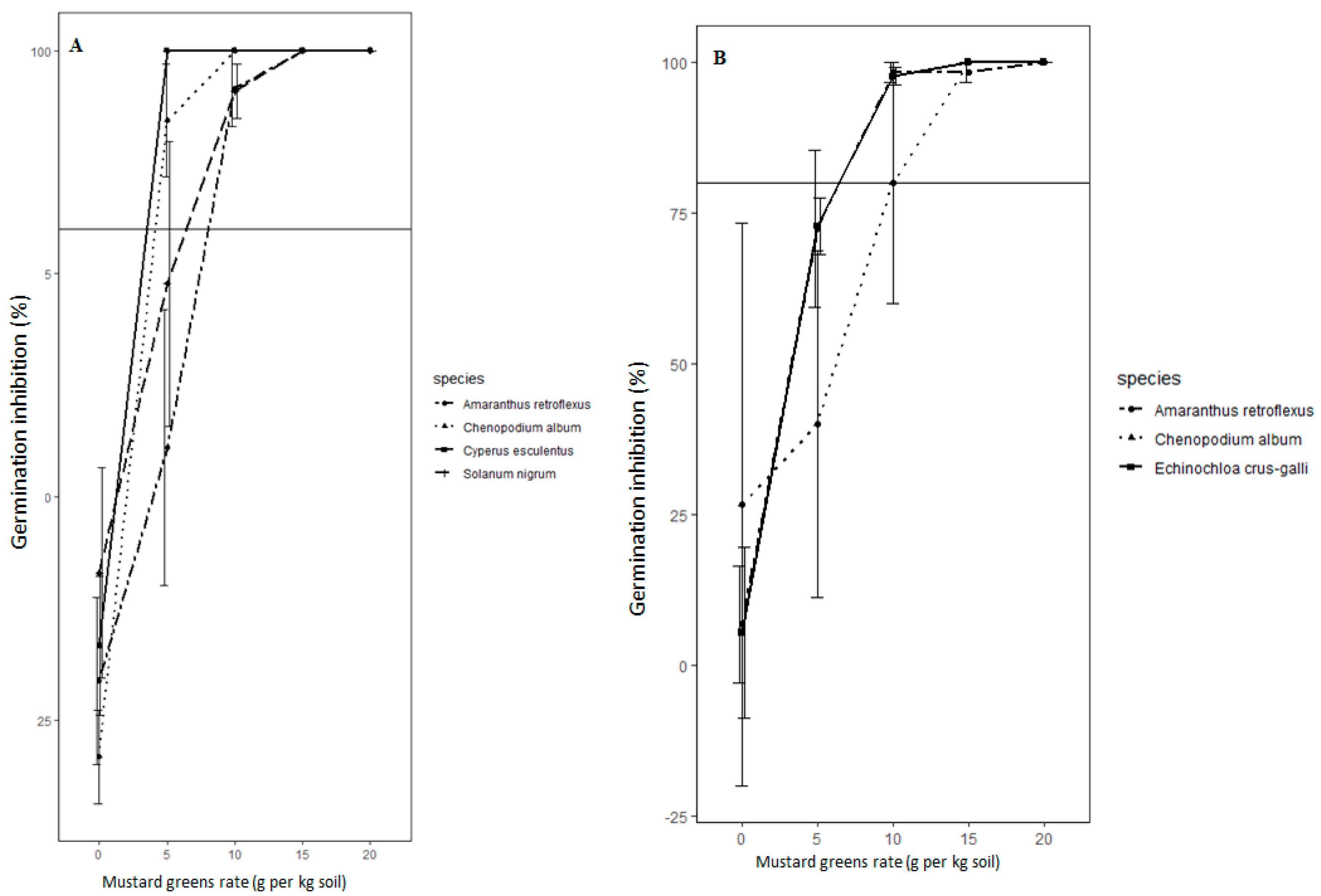
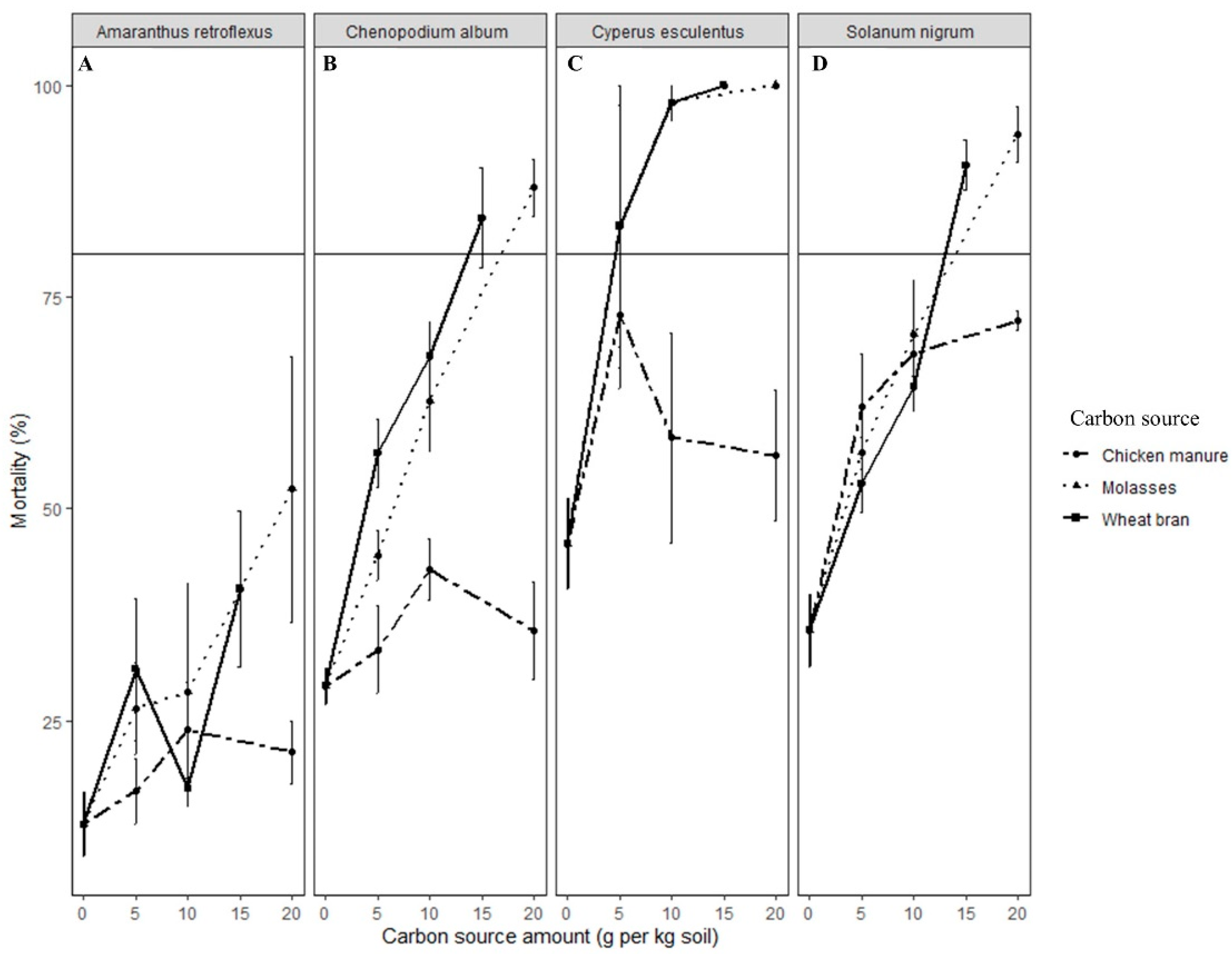
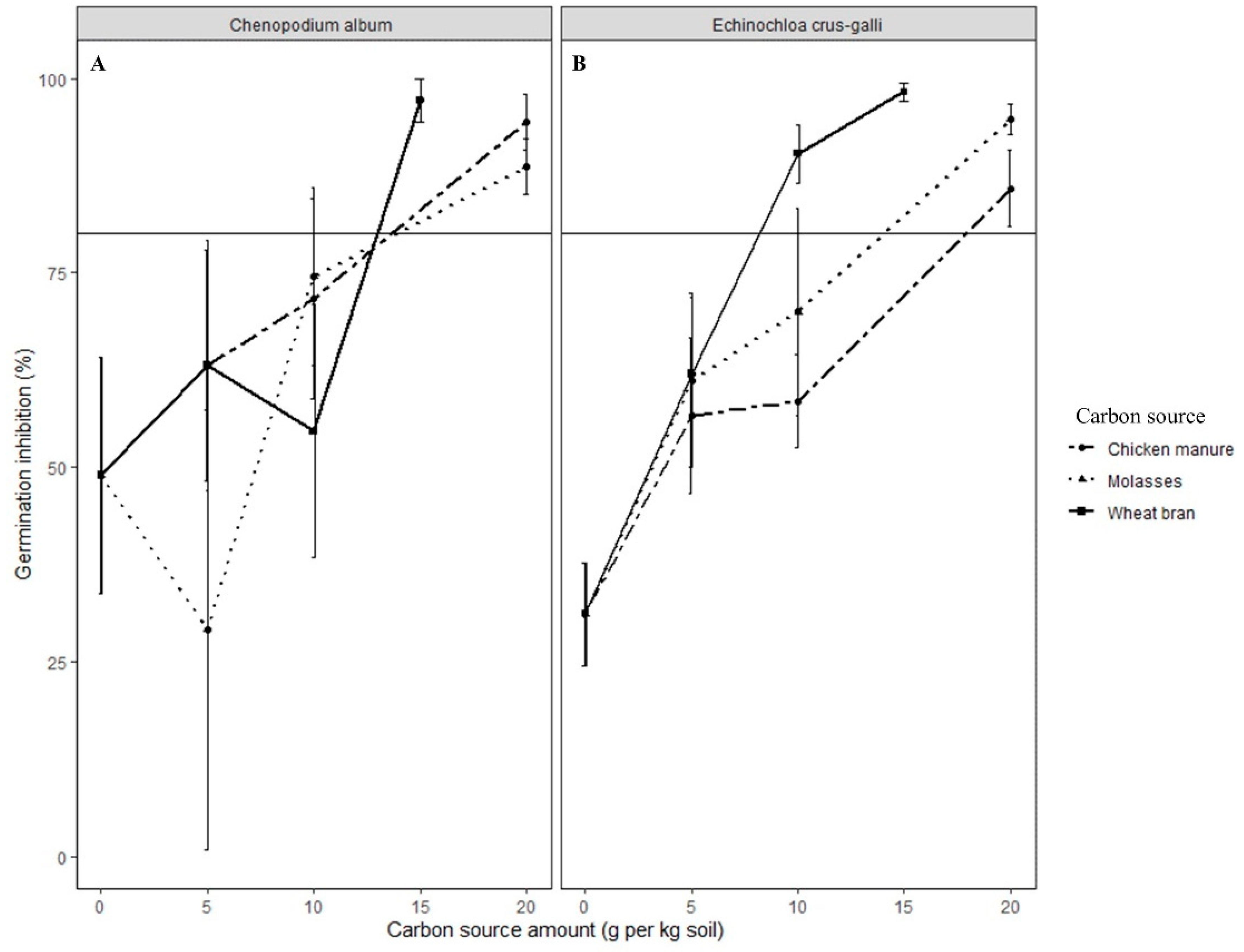
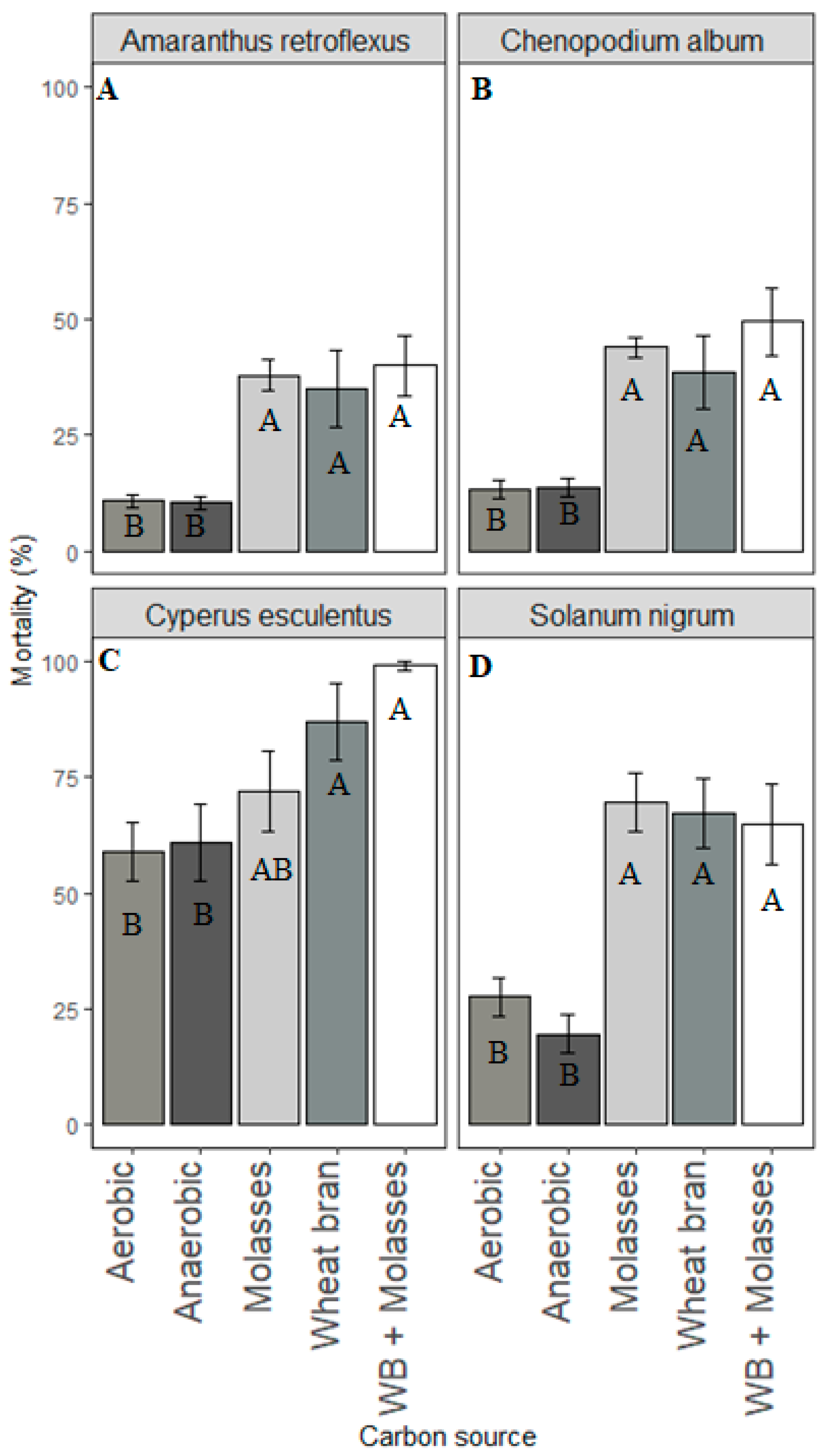
In this study, propagules of all weed species in ASD-treated soil were killed. However, the lethality of ASD varied according to the types and rates of carbon sources used and the weed species evaluated. Wheat bran and molasses were highly effective carbon sources for ASD, causing seed mortality of >90% for most of the weed species tested across all the trials, the exception being A. retroflexus seeds in soil treated with ASD-molasses. However, ASD with chicken manure or mustard greens amendment was not as effective as ASD-wheat bran or -molasses in killing seeds of S. nigrum and C. album. A high rate (≥20 Mg ha−1 dry weight) of mustard greens was necessary to kill >90% of the seeds of the weed species tested during ASD.
The seeds of Taraxacum officinale and P. decandra were excluded from further experiments when initial studies indicated their high sensitivity to flooded, covered soil conditions. Propagules of C. esculentus were the most sensitive, and ASD with 10 Mg ha−1 mustard greens, wheat bran, or molasses would likely be optimal for suppression of tuber germination. Seeds of C. album, S. nigrum and E. crus-galli might require rates of 40 Mg ha−1, 30 Mg ha−1, or 20 Mg ha−1 molasses, wheat bran or mustard greens biomass, respectively. Amaranthus retroflexus seed suppression was not consistent across growth chamber and field experiments. However, it appears that high rates of mustard greens biomass (20 Mg ha−1) and wheat bran (50 Mg ha−1) are necessary to kill A. retroflexus seeds during ASD.
Our results agree with the observations of Shrestha et al. [
3]. In their metanalysis study ASD strongly suppressed
C. esculentus and
C. album, however it did not effectively suppress small-seeded weeds such as red pigweed and crabgrass (
Digitaria anguinalis). In a metanalysis of 88 published studies conducted by Shrestha et al. [
3], weed suppression ranged from 32% to 88%. The effectiveness of ASD in suppressing weeds is mostly influenced by weed species, and to a lesser extent by carbon source [
3,
4]. Germination of common chickweed (
Stellaria media L.),
A. retroflexus, and white clover (
Trifolium repens L.) seeds and
C. esculentus tubers was suppressed similarly by ASD with rice bran, paper mulch, brewer`s spent grain, waste coffee grounds, or peanut shells or sorghum-sudangrass, cowpea, and buckwheat-plant residues used at the rate 4 mg carbon g
−1 soil [
22]. They also observed different degrees of sensitivity of weed species to ASD. For instance,
C. esculentus was more sensitive to ASD than
S. media or
A. retroflexus. The number of sprouted tubers of
C. esculentus recovered after ASD treatment was 42% in wheat bran- compared to 65% in dry molasses-amended ASD [
9].
Thermal, biological, and chemical mechanisms are considered responsible for weed seed inactivation in soil [
23]. All these mechanisms are used in ASD to kill weed seeds in the soil. Disturbance of soil during carbon source incorporation and watering to facilitate the carbon source decomposition might create a false (stale) seedbed situation at the beginning of the ASD process [
24]. Furthermore, increases in soil temperature due to solarization might provide favorable conditions for seed germination. Mirsky et al. [
25] suggested that soil disturbance enhanced the germination of many weed seeds by bringing the seeds to a favorable germination depth in soil [
26], causing a fluctuation in soil temperature [
27] and exposing seeds to light [
28]. After a period, soil oxygen gradually is depleted due to high microbial activity and at the same time, anaerobic fermentation processes yield different types of metabolic products and elevate the soil temperature due to exothermic reactions during decomposition of carbon sources. Accumulation of metabolic products, mostly organic fatty acids, and soil reduction due to depletion of oxygen, also changes the soil pH during ASD [
5,
29,
30,
31]. Overall, there are drastic changes in soil chemical and biological environments that are not favorable for seed juveniles to survive. Furthermore, these conditions enhance the degradation of the weed seed coat or make the seed coat permeable to the metabolites, eventually resulting in a decline in seedbank viability.
The depletion of oxygen levels during seed germination could be a potential mechanism of weed control during ASD [
10]. Benvenuti and MacChia [
23] suggested that hypoxic conditions in the soil only might not be sufficient to kill the weed seeds but the presence of the final product from the fermentation process might play a significant role in influencing seed germination. Later on, several researchers described short-chain fatty acids such as acetic acid, propionic acid, isobutyric acid, butyric acid, and valeric acid as potentially active metabolites produced during anaerobic decomposition of organic amendments in soil to kill weed seed, soilborne pests and pathogens [
5,
6,
30,
32,
33]. Hewavitharana et al. [
6] reported additional biocidal volatile compounds such as dimethyl trisulfide, 2-ethyl-1-hexanol, decanal, and nonanal produced during ASD process. Furthermore, covering with plastic prevented the diffusion of fermentation products and made them readily available to kill the weeds propagules in soil during ASD.
A positive correlation between accumulation of short-chain fatty acids in soil and seed mortality has been reported [
34]. Most of the volatile fatty acid concentrations were sufficient to kill weed propagules under elevated temperatures during ASD [
33] and composting processes [
34]. However, Shiralipour et al. [
35] indicated that acetic acid concentrations over 30 mg kg
−1 are phytotoxic to plants. Reports from different workers indicate that ASD yields far more organic acids than this threshold during the anaerobic decomposition of carbon sources. Hestmark et al. [
36] reported the accumulation of as much as 250 to 500 mg kg
−1 of acetic acid and 300 to 350 mg kg
−1 of propionic acid when green waste and green waste plus compost (GWC) were used as ASD carbon sources. Furthermore, Sanabria-Velazquez et al. [
20] reported 1283 to 4718 mg kg
−1 total organic acids including 1046 to 1442 mg kg
−1 of acetic acid, 122 to 333 mg propionic acid kg
−1, 59 to 308 mg isobutyric acid kg
−1, 419 to 2729 mg butyric acid kg
−1 when using mustard greens, ethanol, wheat bran, and molasses as ASD carbon sources. These reports from diverse working groups working with different carbon sources indicate that organic fatty acids are the major inhibitory compounds for weed seed mortality in ASD. This is also supported by our assays in which seeds of
E. crus-galli and
C. album and tubers of
C. esculentus treated with a combination of 50% acetic acid and 50% butyric acid at concentrations > 400 µL L
−1 water were killed when exposed for at least 60 h.
The ASD treatments with different carbon sources did not change the soil pH drastically. Only high rates of molasses and wheat bran (50 Mg ha
−1) applied to soil resulted in reduced soil pH. Mustard greens biomass did not change the soil pH significantly compared to the anaerobic control; in contrast, chicken manure slightly increased the soil pH. Previous workers reported that ASD treatment resulted in decreased soil pH due to the addition of organic acids in soil during carbon source breakdown [
5,
29,
30,
31]. The soil used in this study was from an organically managed farm; the lack of decrease in soil pH might be due to higher buffering capacity of soil. Generally, the addition of organic matter to soil enhances the humic acid percentage, which increases the elasticity (buffering capacity) of soil to tolerate pH fluctuations due to the addition of acidic and alkaline agents. The slight increase in soil pH in the case of chicken manure-amended ASD treatments might be due to the addition of calcium; generally, chicken manure is a rich source of calcium, which can help to elevate alkalinity in soil. It might also be due to ammonium accumulated under anaerobic decomposition. Elevated soil pH due to the application of chicken manure was also reported by Duruigbo et al. [
37].
An interesting observation of this study was the higher gravimetric soil moisture content in wheat bran-amended ASD-treated soil compared to others. Water absorption during saturation was visibly slower in wheat bran-amended soils than in soils subjected to other treatments. The higher retention of water in soil amended with wheat bran compared to other carbon sources could be due to increased soil porosity, open surface area and potential influence of cation exchange capacity that might change the soil sorptive properties [
38]. However, more studies are warranted on this perspective. Furthermore, soil-reducing conditions were high in all ASD-treated soils in all the experiments with the exception of ASD-treated soil amended with the high rate of wheat bran (50 Mg ha
−1). Interestingly, in the rate optimization experiment, increasing the rate of carbon source amendment increased the intensity of soil-reducing conditions for all the carbon sources, although the results were inconsistent for wheat bran. Wheat bran contains a high percentage of phytic acid [
39], which makes insoluble complexes with many minerals; however, lactic acid bacteria can break down this complex to utilize as an energy source. Under high wheat bran rates, lactic acid bacteria populations present in soil might not be high enough for complete decomposition and that might be the reason for lower soil-reducing conditions in ASD-treated soil amended with a high rate of wheat bran.
Weed management practices that result in inhibition of more than 50% of the germinable seedbank are considered effective for small hard-seeded weed species [
40]. In the controlled environment and field verification experiments conducted in this study, ASD met or exceeded this criterion for all the weed species tested, depending on type and rate of the carbon source.












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}