
Maize Rotation Combined with Streptomyces rochei D74 to Eliminate Orobanche cumana Seed Bank in the Farmland
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of S. rochei D74 Fermentation Filtrates
2.2. Petri Dish Bioassay Experiments
2.3. Co-Culture Experiments
2.4. Design of Pot Experiments
2.5. Measurement of Plant Physiological Parameters
2.6. Sunflower Rhizosphere Soil Collection, Isolation and Identification of Bacteria
2.7. Extraction and Sequencing of Total DNA from Rhizosphere Soil
2.8. Statistical Analyses
3. Results
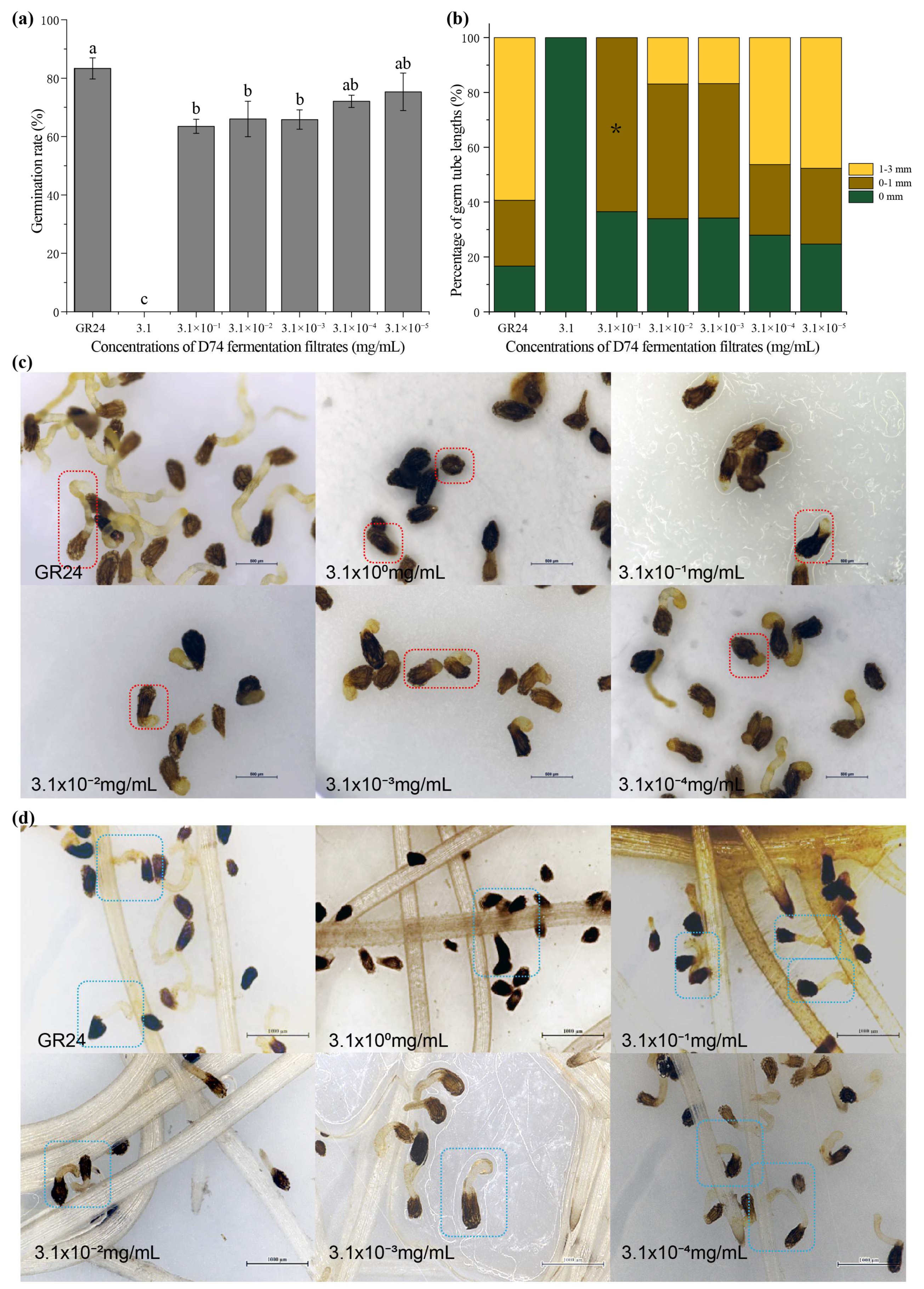
3.1. Effects of D74 Fermentation Filtrates on the Germination of O. cumana Seeds
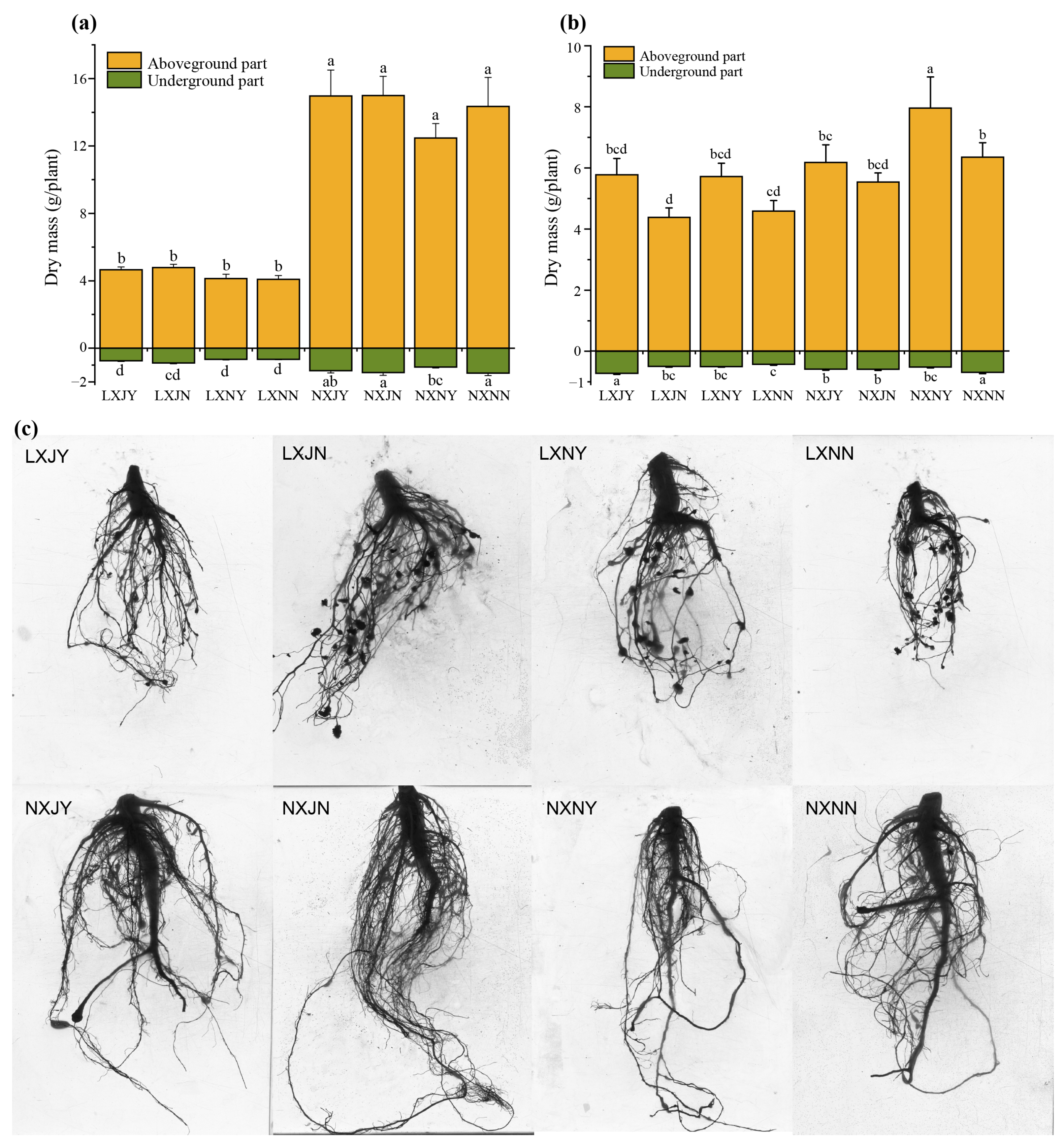
3.2. Effects of Maize Rotation and S. rochei D74 Agents on Sunflower Growth
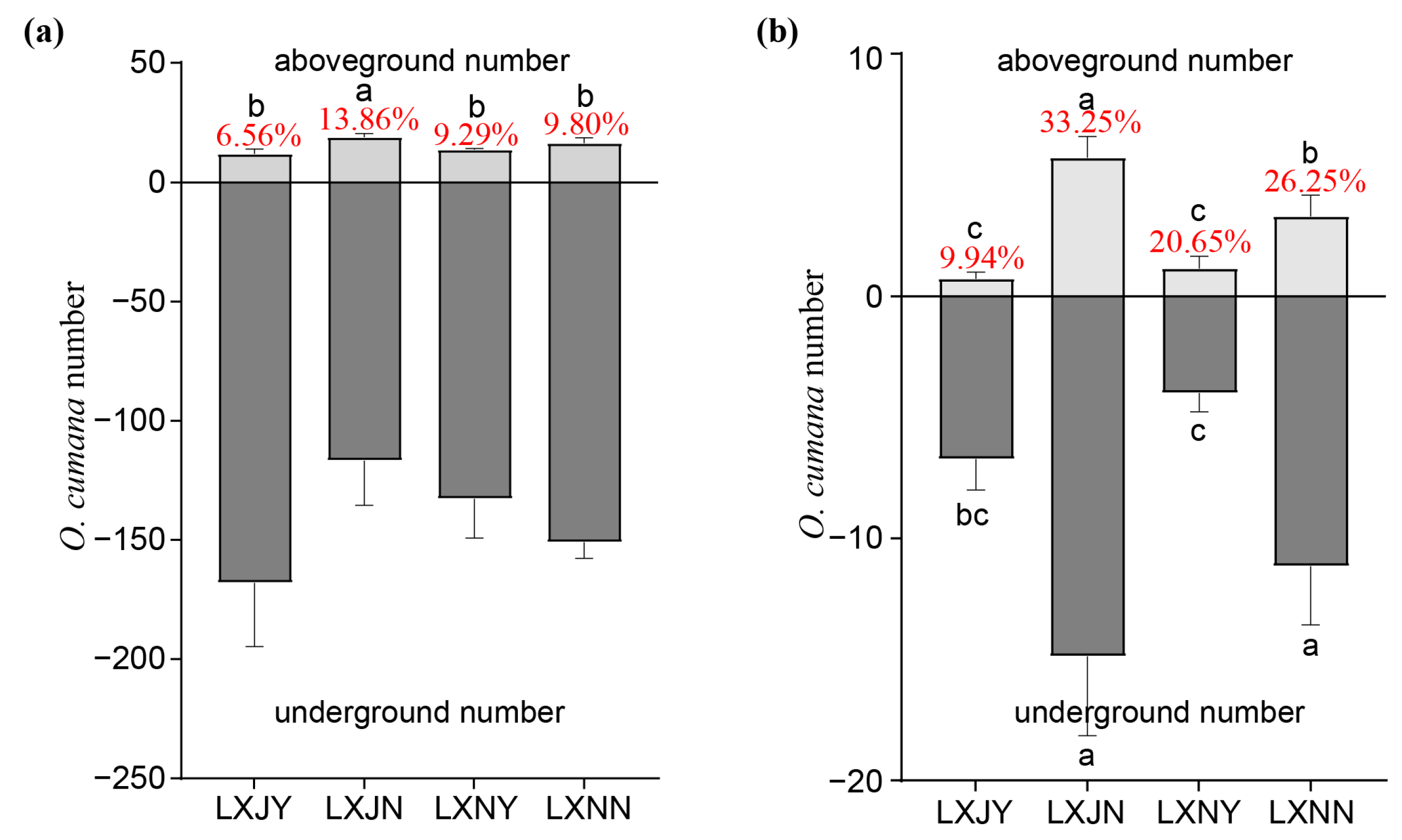
3.3. Effect of Maize Rotation and S. rochei D74 Agents on O. cumana Parasitism
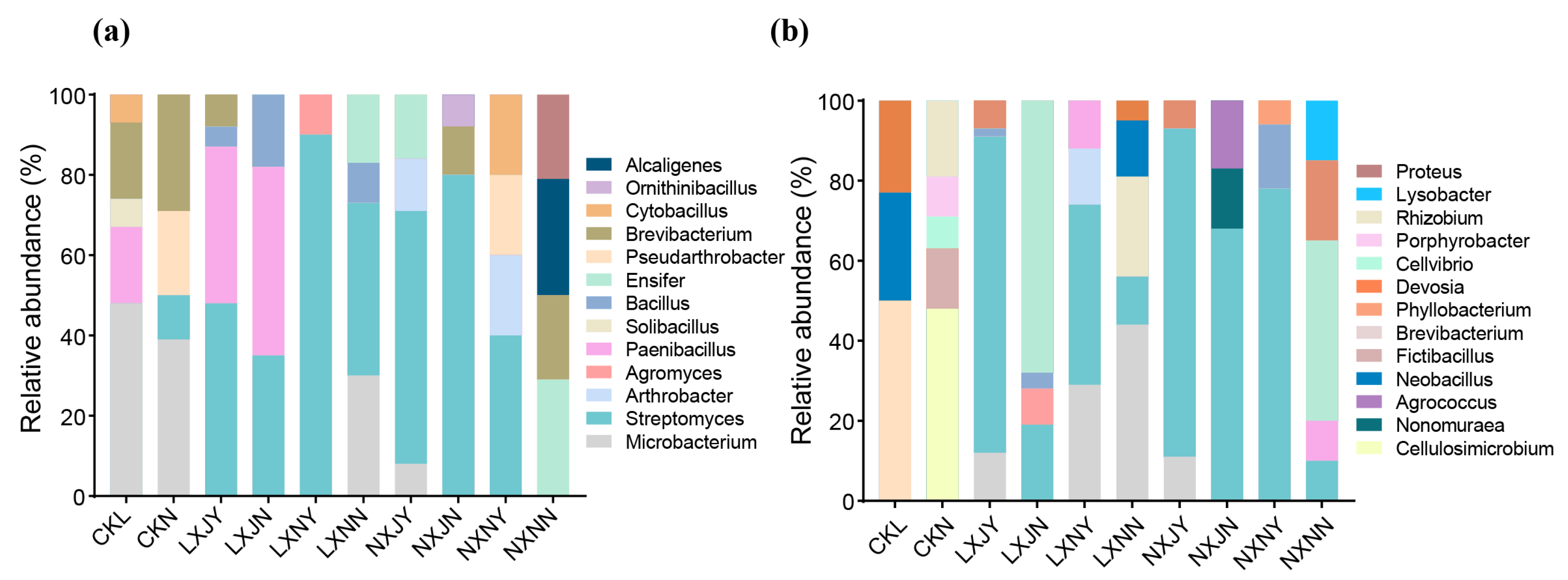
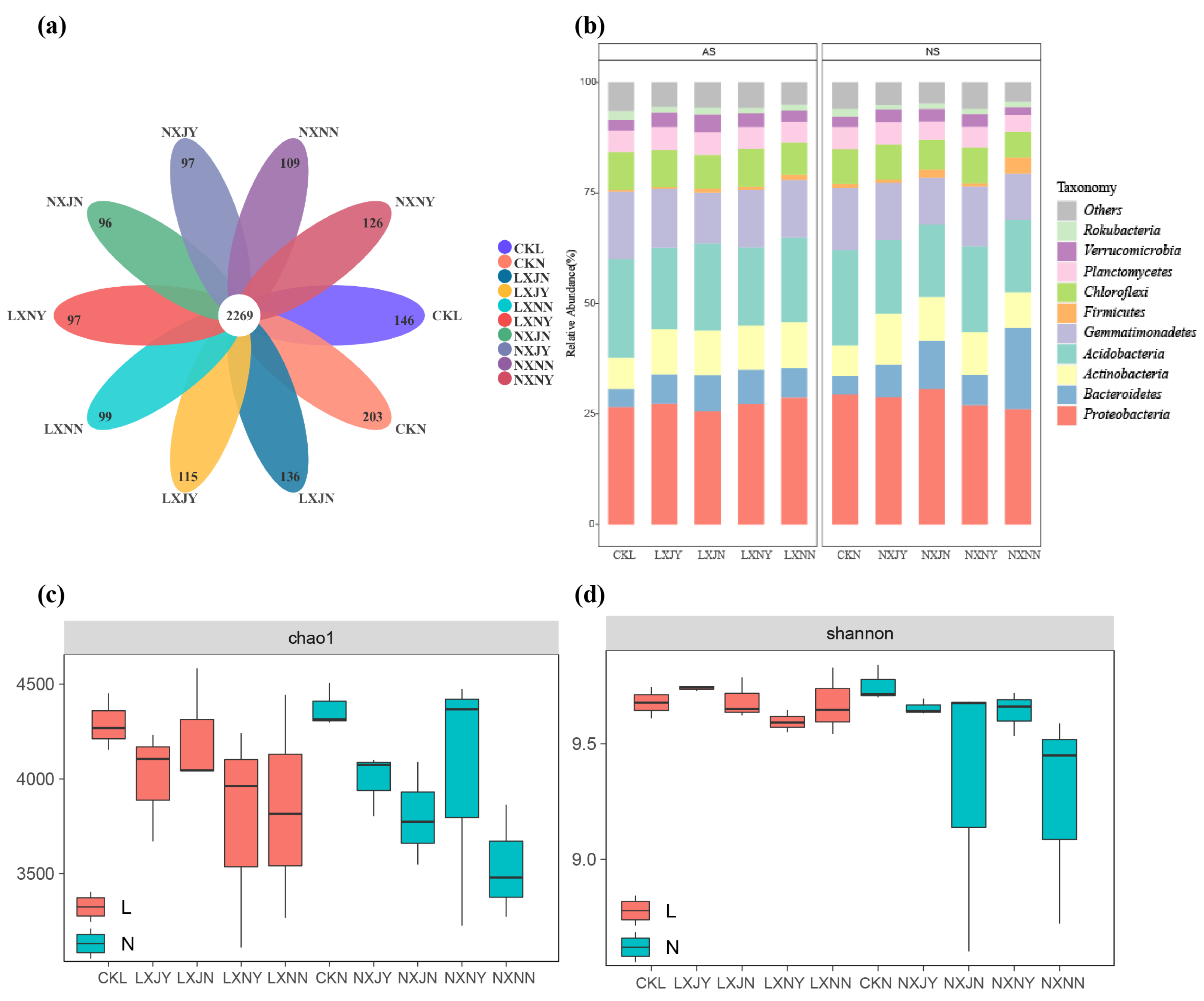
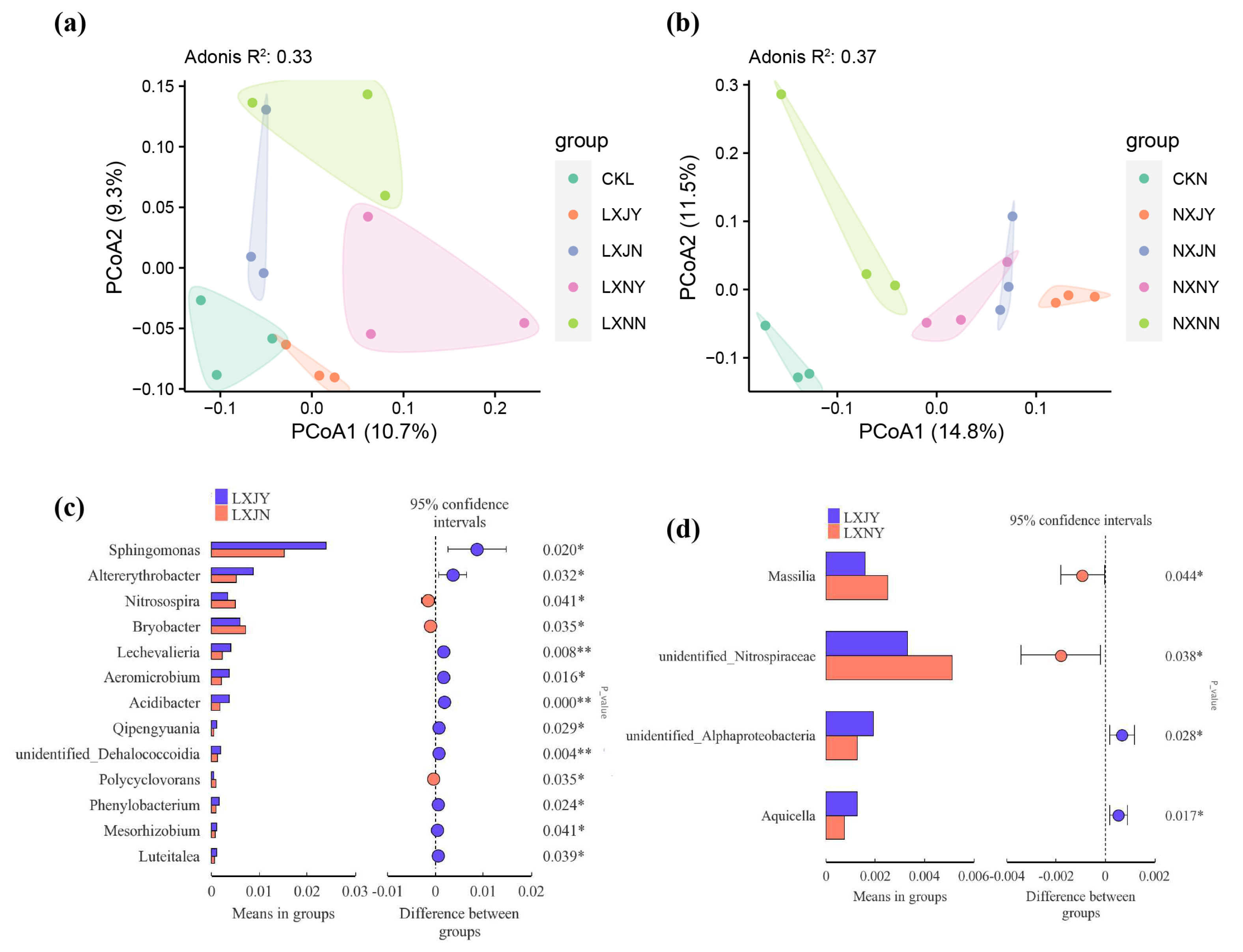
3.4. Effect of Maize Rotation and S. rochei D74 Agents on Rhizosphere Microbiota
3.4.1. Comparison between Culturable Microbial Dominant Strains in Sunflower Rhizosphere Soil
3.4.2. Effects of S. rochei D74 Agents and Maize Rotation on Sunflower Rhizosphere Bacterial Community
4. Discussion
4.1. Effects of D74 Fermentation Filtrates on the Germination of O. cumana Seeds
4.2. Effects of Maize Rotation and S. rochei D74 Agents on Sunflower Growth and O. cumana Parasitism
4.3. Effects of Maize Rotation and S. rochei D74 Agents on Rhizosphere Microbial Community
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yoshida, S.; Cui, S.; Ichihashi, Y.; Shirasu, K. The haustorium, a specialized invasive organ in parasitic plants. Annu. Rev. Plant Biol. 2016, 67, 643–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westwood, J.H.; Yoder, J.I.; Timko, M.P.; Depamphilis, C.W. The evolution of parasitism in plants. Trends Plant Sci. 2010, 15, 227–235. [Google Scholar] [CrossRef]
- Uraguchi, D.; Kuwata, K.; Hijikata, Y.; Yamaguchi, R.; Imaizumi, H.; Am, S.; Rakers, C.; Mori, N.; Akiyama, K.; Irle, S.; et al. A femtomolar–range suicide germination stimulant for the parasitic plant Striga hermonthica. Science 2018, 362, 1301–1305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plakhine, D.; Joel, D.M. Ecophysiological consideration of Orobanche cumana germination. Helia 2010, 33, 13–18. [Google Scholar] [CrossRef]
- Mutuku, J.M.; Cui, S.; Yoshida, S.; Shirasu, K. Orobanchaceae parasite–host interactions. New Phytol. 2021, 230, 46–59. [Google Scholar] [CrossRef]
- Goldwasser, Y.; Rodenburg, J. Integrated agronomic management of parasitic weed seed banks. In Parasitic Orobanchaceae; Joel, D.M., Ed.; Springer: Berlin, Germany, 2013; pp. 393–413. [Google Scholar]
- Krupp, A.; Rücker, E.; Heller, A.; Spring, O. Seed structure characteristics of Orobanche cumana populations. Helia 2015, 38, 1–14. [Google Scholar] [CrossRef]
- Parker, C. Observations on the current status of Orobanche and Striga problems worldwide. Pest Manag. Sci. 2009, 65, 453–459. [Google Scholar] [CrossRef]
- Cimmino, A.; Masi, M.; Rubiales, D.; Evidente, A.; Fernández–Aparicio, M. Allelopathy for parasitic plant management. Nat. Prod. Commun. 2018, 13, 289–294. [Google Scholar] [CrossRef] [Green Version]
- Goldwasser, Y.; Kleifeld, Y. Recent approaches to Orobanche management. Weed Biol. Manag. 2004, 1, 439–466. [Google Scholar]
- Duriez, P.; Vautrin, S.; Auriac, M.-C.; Bazerque, J.; Boniface, M.-C.; Callot, C.; Carrère, S.; Cauet, S.; Chabaud, M.; Gentou, F. A receptor–like kinase enhances sunflower resistance to Orobanche cumana. Nat. Plants 2019, 5, 1211–1215. [Google Scholar] [CrossRef]
- Van Bruggen, A.H.; He, M.; Shin, K.; Mai, V.; Jeong, K.; Finckh, M.; Morris Jr, J. Environmental and health effects of the herbicide glyphosate. Sci. Total Environ. 2018, 616, 255–268. [Google Scholar] [CrossRef] [PubMed]
- Parker, C. A personal history in parasitic weeds and their control. Plants 2021, 10, 2249. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Zhang, M.; Li, Y.; Shui, J.; Zhou, Y. Allelopathy of rice (Oryza sativa L.) root exudates and its relations with Orobanche cumana Wallr. and Orobanche minor Sm. germination. J. Plant Interact. 2014, 9, 722–730. [Google Scholar] [CrossRef]
- Fernández-Aparicio, M.; GarcÍa-Garrido, J.M.; Ocampo, J.A.; Rubiales, D. Colonisation of field pea roots by arbuscular mycorrhizal fungi reduces Orobanche and Phelipanche species seed germination. Weed Res. 2010, 50, 262–268. [Google Scholar] [CrossRef]
- Xi, J.; Lei, B.; Liu, Y.X.; Ding, Z.; Liu, J.; Xu, T.; Hou, L.; Han, S.; Qian, X.; Ma, Y. Microbial community roles and chemical mechanisms in the parasitic development of Orobanche cumana. iMeta 2022, 1, e31. [Google Scholar] [CrossRef]
- Zermane, N.; Souissi, T.; Kroschel, J.; Sikora, R. Biocontrol of broomrape (Orobanche crenata Forsk. and Orobanche foetida Poir.) by Pseudomonas fluorescens isolate Bf7–9 from the faba bean rhizosphere. Biocontrol Sci. Technol. 2007, 17, 483–497. [Google Scholar] [CrossRef]
- Gonsior, G.; Buschmann, H.; Szinicz, G.; Spring, O.; Sauerborn, J. Induced resistance–an innovative approach to manage branched broomrape (Orobanche ramosa) in hemp and tobacco. Weed Sci. 2004, 52, 1050–1053. [Google Scholar] [CrossRef]
- Chen, J.; Xue, Q.; McErlean, C.; Zhi, J.; Ma, Y.; Jia, X.; Zhang, M.; Ye, X. Biocontrol potential of the antagonistic microorganism Streptomyces enissocaesilis against Orobanche cumana. BioControl 2016, 61, 781–791. [Google Scholar] [CrossRef]
- Li, Y.; Li, H.; Han, X.; Han, G.; Xi, J.; Liu, Y.; Zhang, Y.; Xue, Q.; Guo, Q.; Lai, H. Actinobacterial biofertilizer improves the yields of different plants and alters the assembly processes of rhizosphere microbial communities. Appl. Soil Ecol. 2022, 171, 104345. [Google Scholar] [CrossRef]
- Li, Y.; Guo, Q.; He, F.; Li, Y.; Xue, Q.; Lai, H. Biocontrol of root diseases and growth promotion of the tuberous plant Aconitum carmichaelii induced by Actinomycetes are related to shifts in the rhizosphere microbiota. Microb. Ecol. 2020, 79, 134–147. [Google Scholar] [CrossRef]
- Fei, H.; Zhong–liang, Z.; Ming, C.; Quan–hong, X.; Dong–sheng, W. Disease prevention and growth promotion effects of actinomycete strain D74 on Amorphophallus konjac. Acta Hortic. Sin. 2015, 42, 367. [Google Scholar]
- Shi, L.; Nwet, T.T.; Ge, B.; Zhao, W.; Liu, B.; Cui, H.; Zhang, K. Antifungal and plant growth–promoting activities of Streptomyces roseoflavus strain NKZ–259. Biol. Control 2018, 125, 57–64. [Google Scholar] [CrossRef]
- Hothem, S.D.; Marley, K.A.; Larson, R.A. Photochemistry in Hoagland’s nutrient solution. J. Plant Nutr. 2003, 26, 845–854. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, Y.-X.; Guo, X.; Qin, Y.; Garrido-Oter, R.; Schulze-Lefert, P.; Bai, Y. High–throughput cultivation and identification of bacteria from the plant root microbiota. Nat. Protoc. 2021, 16, 988–1012. [Google Scholar] [CrossRef] [PubMed]
- Perry, Z.H.; Barak, A.-T.; Neumann, L.; Levy, A. Computer–based learning: The use of SPSS statistical program for improving biostatistical competence of medical students. J. Biomed. Educ. 2014, 2014, 298140. [Google Scholar] [CrossRef]
- Moberly, J.G.; Bernards, M.T.; Waynant, K.V. Key features and updates for origin 2018. J. Cheminf. 2018, 10, 5. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.; Liu, Y.X.; Huang, L. ImageGP: An easy-to-use data visualization web server for scientific researchers. iMeta 2022, 1, e5. [Google Scholar] [CrossRef]
- Chen, T.; Zhang, H.; Liu, Y.; Liu, Y.-X.; Huang, L. EVenn: Easy to create repeatable and editable Venn diagrams and Venn networks online. J. Genet. Genom. 2021, 48, 863–866. [Google Scholar] [CrossRef]
- Bouwmeester, H.J.; Roux, C.; Lopez–Raez, J.A.; Becard, G. Rhizosphere communication of plants, parasitic plants and AM fungi. Trends Plant Sci. 2007, 12, 224–230. [Google Scholar] [CrossRef]
- Cardoso, C.; Ruyter–Spira, C.; Bouwmeester, H.J. Strigolactones and root infestation by plant–parasitic Striga, Orobanche and Phelipanche spp. Plant Sci. 2011, 180, 414–420. [Google Scholar] [CrossRef]
- Albert, M.; Axtell, M.J.; Timko, M.P. Mechanisms of resistance and virulence in parasitic plant–host interactions. Plant Physiol. 2021, 185, 1282–1291. [Google Scholar] [CrossRef] [PubMed]
- Müller–Stöver, D.; Batchvarova, R.; Kohlschmid, E.; Sauerborn, J. Mycoherbicidal management of Orobanche cumana: Observations from three years of field experiments. In Proceedings of the 10th International World Congress of Parasitic Plants, Kusadasi, Turkey, 8–12 June 2009; p. 86. [Google Scholar]
- Boari, A.; Vurro, M. Evaluation of Fusarium spp. and other fungi as biological control agents of broomrape (Orobanche ramosa). Biol. Control 2004, 30, 212–219. [Google Scholar] [CrossRef]
- Anteyi, W.O.; Klaiber, I.; Rasche, F. Diacetoxyscirpenol, a Fusarium exometabolite, prevents efficiently the incidence of the parasitic weed Striga hermonthica. BMC Plant Biol. 2022, 22, 84. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Li, Y.; Luo, W.; Zhou, J.; Zhao, S.; Xu, J. Herbicidal secondary metabolites from Bacillus velezensis JTB8–2 against Orobanche aegyptiaca. AMB Express 2022, 12, 52. [Google Scholar] [CrossRef]
- Ma, Y.; Jia, J.; An, Y.; Wang, Z.; Mao, J. Potential of some hybrid maize lines to induce germination of sunflower broomrape. Crop. Sci. 2013, 53, 260–270. [Google Scholar] [CrossRef] [Green Version]
- Guorong, N.; Guoquan, T.; Saijin, W.; Jianfu, W.; Qinghua, S.; Chunhuo, Z.; Xiaohua, P. Effects of combination of straw returning and a microbial agent on microorganisms and enzyme activity in rhizosphere soil and yield of late rice. Meteorol. Environ. Res. 2017, 8, 78–82. [Google Scholar]
- Singh, G.; Mukerji, K.G. Root exudates as determinant of rhizospheric microbial biodiversity. In Microbial Activity in the Rhizoshere; Springer: Berlin, Germany, 2006; pp. 39–53. [Google Scholar]
- Zhalnina, K.; Louie, K.B.; Hao, Z.; Mansoori, N.; da Rocha, U.N.; Shi, S.; Cho, H.; Karaoz, U.; Loqué, D.; Bowen, B.P.; et al. Dynamic root exudate chemistry and microbial substrate preferences drive patterns in rhizosphere microbial community assembly. Nat. Microbiol. 2018, 3, 470–480. [Google Scholar] [CrossRef] [Green Version]
- Jiang, N.; Wu, M.; Li, G.; Li, P.; Liu, M.; Li, Z. Comparative effects of two humic substances on microbial dysbiosis in the rhizosphere soil where cucumber (Cucumis sativus L.) is grown. Land Degrad. Dev. 2022, 33, 1944–1953. [Google Scholar] [CrossRef]
- Mendes, R.; Kruijt, M.; De Bruijn, I.; Dekkers, E.; Van Der Voort, M.; Schneider, J.H.; Piceno, Y.M.; DeSantis, T.Z.; Andersen, G.L.; Bakker, P.A. Deciphering the rhizosphere microbiome for disease-suppressive bacteria. Science 2011, 332, 1097–1100. [Google Scholar] [CrossRef]
- Ezziyyani, M.; Requena, M.; Egea-Gilabert, C.; Candela, M. Biological control of Phytophthora root rot of pepper using Trichoderma harzianum and Streptomyces rochei in combination. J. Phytopathol. 2007, 155, 342–349. [Google Scholar] [CrossRef]
- Suwitchayanon, P.; Chaipon, S.; Chaichom, S.; Kunasakdakul, K. Potentials of Streptomyces rochei ERY1 as an endophytic actinobacterium inhibiting damping–off pathogenic fungi and growth promoting of cabbage seedling. Chiang Mai J. Sci 2018, 45, 692–700. [Google Scholar]
- Meena, R.K.; Singh, R.K.; Singh, N.P.; Meena, S.K.; Meena, V.S. Isolation of low temperature surviving plant growth–promoting rhizobacteria (PGPR) from pea (Pisum sativum L.) and documentation of their plant growth promoting traits. Biocatal. Agric. Biotechnol. 2015, 4, 806–811. [Google Scholar] [CrossRef]
- Hua, L.Q.; Yang, S.Q.; Xia, Z.F.; Zeng, H. Application of Sophora alopecuroides organic fertilizer changes the rhizosphere microbial community structure of melon plants and increases the fruit sugar content. J. Sci. Food Agric. 2022, 103, 164–175. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Time | Group | Plant Height/cm | Stalk Diameter /mm | Root Length/cm |
|---|---|---|---|---|
| First year | LXJY | 62.00 ± 1.59 b | 9.13 ± 0.17 bc | 13.71 ± 0.32 bc |
| LXJN | 60.50 ± 1.4 b | 9.28 ± 0.19 bc | 10.64 ± 0.92 c | |
| LXNY | 60.75 ± 1.28 b | 8.99 ± 0.21 d | 12.29 ± 0.21 c | |
| LXNN | 61.00 ± 2.25 b | 8.75 ± 0.12 d | 11.06 ± 1.05 c | |
| NXJY | 126.76 ± 5.65 a | 9.63 ± 0.36 bc | 17.38 ± 2.38 b | |
| NXJN | 140.81 ± 5.23 a | 10.57 ± 0.64 a | 22.31 ± 2.68 a | |
| NXNY | 135.34 ± 7.65 a | 9.58 ± 0.21 bc | 23.24 ± 1.97 a | |
| NXNN | 137.10 ± 7.91 a | 10.05 ± 0.21 ab | 24.50 ± 1.93 a | |
| Second year | LXJY | 103.06 ± 0.10 c | 8.23 ± 0.22 ab | 14.70 ± 0.51 c |
| LXJN | 79.84 ± 0.60 e | 7.85 ± 0.18 b | 10.27 ± 0.36 e | |
| LXNY | 92.67 ± 2.38 d | 8.28 ± 0.19 ab | 13.14 ± 0.56 cd | |
| LXNN | 87.26 ± 1.53 d | 7.05 ± 0.17 c | 10.87 ± 0.68 de | |
| NXJY | 116.29 ± 2.14 a | 7.81 ± 0.08 b | 28.90 ± 1.09 a | |
| NXJN | 108.94 ± 1.58 b | 7.79 ± 0.24 b | 21.71 ± 1.33 b | |
| NXNY | 114.16 ± 2.55 ab | 8.42 ± 0.18 a | 23.79 ± 1.01 b | |
| NXNN | 117.97 ± 2.86 a | 8.03 ± 0.08 ab | 22.36 ± 1.11 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xi, J.; Ding, Z.; Xu, T.; Qu, W.; Xu, Y.; Ma, Y.; Xue, Q.; Liu, Y.; Lin, Y. Maize Rotation Combined with Streptomyces rochei D74 to Eliminate Orobanche cumana Seed Bank in the Farmland. Agronomy 2022, 12, 3129. https://doi.org/10.3390/agronomy12123129
Xi J, Ding Z, Xu T, Qu W, Xu Y, Ma Y, Xue Q, Liu Y, Lin Y. Maize Rotation Combined with Streptomyces rochei D74 to Eliminate Orobanche cumana Seed Bank in the Farmland. Agronomy. 2022; 12(12):3129. https://doi.org/10.3390/agronomy12123129
Chicago/Turabian StyleXi, Jiao, Zanbo Ding, Tengqi Xu, Wenxing Qu, Yanzhi Xu, Yongqing Ma, Quanhong Xue, Yongxin Liu, and Yanbing Lin. 2022. "Maize Rotation Combined with Streptomyces rochei D74 to Eliminate Orobanche cumana Seed Bank in the Farmland" Agronomy 12, no. 12: 3129. https://doi.org/10.3390/agronomy12123129