
Recent Advances in Eco-Friendly and Scaling-Up Bioproduction of Prodigiosin and Its Potential Applications in Agriculture



1
Doctoral Program in Applied Sciences, Tamkang University, New Taipei City 25137, Taiwan
2
Department of Chemistry, Tamkang University, New Taipei City 25137, Taiwan
3
Institute of Biotechnology and Environment, Tay Nguyen University, Buon Ma Thuot 630000, Vietnam
*
Authors to whom correspondence should be addressed.
Agronomy 2022, 12(12), 3099; https://doi.org/10.3390/agronomy12123099
Submission received: 6 November 2022
/
Revised: 29 November 2022
/
Accepted: 5 December 2022
/
Published: 7 December 2022
(This article belongs to the Topic Production of Microbial Secondary Metabolites from Organic Wastes and Investigation of Their Potential Applications)
Abstract
:Prodigiosin is a red pigment produced by various microbial strains, of these, Serratia marcescens has been recorded as the major PG-producing strain. This microbial pigment has attracted much attention because it possesses potential applications in various fields. Thus, this active metabolite has been increasingly studied for bioproduction and investigated for its novel applications. Recently, several prodigiosin-related reviews were reported. These studies covered some aspects of the general physicochemical properties, pathway synthesis, production, and applications of prodigiosin in medicine. However, only a few works discussed the eco-friendly production of prodigiosin from organic wastes. Remarkably, the scaling-up of prodigiosin production and its potential applications in agriculture have rarely been reviewed or discussed. This review extensively presents and discusses the green biosynthesis, enhancement, and scaling-up of prodigiosin production from a wide range of organic byproducts/wastes using different methods of fermentation. Notably, this paper highlights the novel and promising applications of prodigiosin in agriculture via in vitro tests, greenhouse tests, and field studies. The action mechanisms related to some potential bioactivities and the toxicology studies of prodigiosin are also discussed. This review thus supplies scientific data for further research and the application of prodigiosin in the future.
1. Introduction
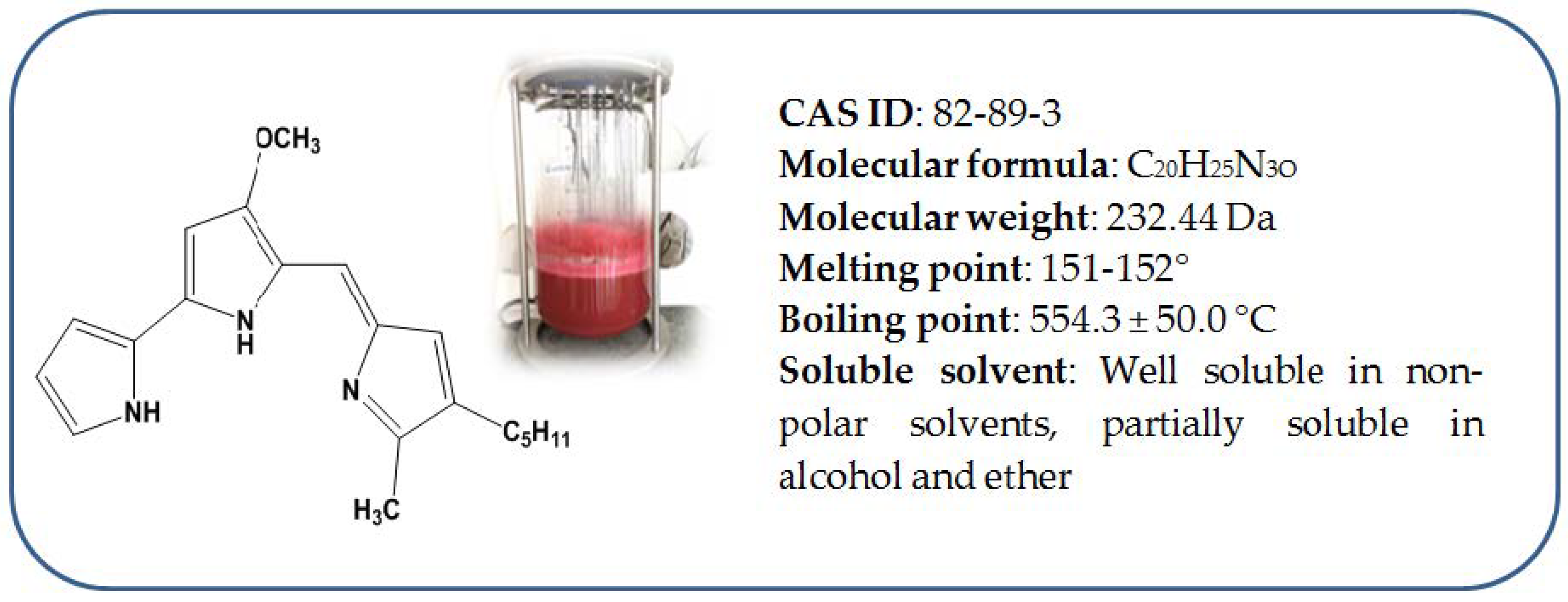
Prodigiosin (PG), a red pigment, is a ring compound belonging to the group of prodiginine with a pyrrolyl pyrromethane skeleton [1]. The structure and physicochemical properties of PG are presented in Figure 1 [2,3]. PG is biosynthesized from various microbial strains, among which, Serratia marcescens is recognized as the major producer of PG [4]. This microbial pigment has been extensively investigated and has evidenced potential for applications in many fields, such as medicine, agriculture, industry, food, and the environment [4,5,6,7,8,9,10,11,12,13,14,15,16,17,18,19,20,21,22]. Furthermore, its safety was also proved in some works [23,24,25,26,27,28].
Due to the various benefits of this pigment, the number of original works concerning PG production and the evaluation of its novel uses has increased dramatically in recent years [2,29,30,31,32,33,34]. To date, various review works on PG have also been published. These studies focused on the topics of general physical–chemical characteristics and biosynthesis pathways of PG [3,35,36], high-level PG production, and the applications of PG in medicine [3,4,35,37,38]. However, the scaling-up of production, action mechanisms, and potential use of PG in agriculture have been rarely mentioned in previous studies. Therefore, this study aims to address these aspects of PG.
2. Production of Prodigiosin via Microbial Fermentation
Natural bioactive pigments are derived from various sources, such as microbes, insects, and plants [39,40]. Of these, the microbial source possesses the advantage of obtaining bioactive pigment compounds [39]. Obtaining pigments and other secondary metabolites from microbial fermentation is of great interest due to the valuable characteristics of microbes (e.g., fast growth and bioproduction of various secondary metabolites by regulating fermentation conditions) and its green, cost-effective production process (e.g., using an abundant and low-cost substrate, fermentation in mild conditions, and no environmental pollution) [41,42,43,44,45,46,47]. Pigments can be produced at high levels by microbial fermentation using low-cost substrates [42,43,44,45], resulting in a high capacity for commercial applications [42]. The objective of reducing the cost of pigment production by using inexpensive media is to raise microbial productivity, causing a decrease in the cost of production [43,44,45]. Subsequently, byproducts/wastes, including agro-industrial byproducts [45], food and kitchen wastes [48,49], and biomass wastes [50], became promising substrate sources for fermentation to produce natural pigments.
PG is also produced via microbial conversion using various kinds of substrates as the carbon/nitrogen source, with the application of scaled-up fermentation to achieve higher productivity in a reduced fermentation time. In this section, different types of substrate sources are discussed to clarify the role of low-cost organic sources and byproducts/wastes compared to the traditional commercial media, with a special focus on the effect of byproduct/waste sources on PG productivity.
2.1. Overview of the Substrate Sources for Prodigiosin Production in Liquid-State Fermentation
Liquid-state fermentation (LSF), also known as submerged fermentation, is characterized by the growth of a microorganism in a liquid with high water content and nutrient ingredients that are supplied into a liquid medium during the culture process. The culture parameters and control growth conditions can be easily managed in this fermentation method. The nutrient ingredients and microorganisms are evenly distributed in the medium [51,52]. In fact, LSF is used very commonly in PG production, and many studies used LSF to produce pigment compounds. The production of PG from Serratia marcescens by the LSF method using various kinds of substrates as the C/N source is summarized in Table 1.
2.1.1. The Commercial and Low-Cost Organic Materials for Prodigiosin Production
As shown in Table 1, many researchers used commercial media, the most popular being Luria–Bertani broth, yeast extract, beef extract, and nutrient broth, for fermentation to biosynthesize PG with a PG yield of up to 5830 mg/L−1 [53,54,55,56,57,58,59,60]. For lower-cost PG production, some studies used inexpensive organic materials for fermentation [4,24,53,56,58,61,63,64]. The medium supplemented with organic materials, such as 4% soybean oil, 4% olive oil, and 6% sunflower oil, recorded an improvement in PG yield of around 525 mg/L−1, 579 mg/L−1, and 790 mg/L−1, respectively, compared to the control commercial medium (50 mg/L−1) [53]. Another report explored effective fermentation in media containing various low-cost substrates, and the highest PG yield (13,622 mg/L−1) was collected from a medium with a final formulation containing peanut powder, olive oil, and beef extract. Organic materials such as fish meal, soybean powder, or corn steep liquor were not suitable for PG production [56]. Some seeds, seed oils, and coconut oil were also used for PG biosynthesis [58]. The PG yield was as high as 16,680 mg/L−1 when using sesame seed broth as substrate and 38,750 mg/L−1 when using the peanut seed broth medium. For the other media, PG productivity ranged from 767 to 2890 mg/L−1 [58]. The potential PG yield (from 16,500–42,030 mg/L−1) was also obtained from media using a substrate, such as seed broth or seed oil [58,63,64]. Thus, seed and seed oil are suitable materials for cost-effective PG production. Overall, low-cost organic sources provide potentially effective PG biosynthesis and reduce the cost of production with improved PG content compared to commercial substrate sources.
2.1.2. The Organic Byproduct/Waste Sources for Prodigiosin Production
To reduce the production cost, some studies used various organic byproducts/wastes (OBWs), including fishery processing byproducts, agro-industrial byproducts, and other organic byproducts/wastes, as C/N sources for microbial production of PG (Table 1).
- Fishery Processing Byproducts
Management of fishery discards, including fishery by-catch and processing byproducts, has been recognized as an emerging research topic [4]. According to data from the Food and Agriculture Organization, around 60% of fishery catch yield is processed, whereas the remaining amount has become a global environmental issue [76]. To resolve this problem, some chitin-rich fishery processing byproducts, such as squid pens, shrimp heads, shrimp, and crab shells, were used to produce high-value microbial secondary metabolites, such as PG (Table 1). Wang et al. [65] used 1.5% squid pens as the sole C/N source for microbial production of PG and obtained a yield of 978 mg/L−1 within 2 days of fermentation. Other studies also used fishery discards as the main substrate and showed effective PG production with a productivity of over 3000 mg/L−1 [2,29,30,31]. Squid pen powder (SPP) was the most suitable substrate for PG production by Serratia marcescens TNU01 with a yield of 3790 mg/L−1 in an optimal medium that contained 1.75% SPP [29]. In fact, squid pens contain chitin/protein in the ratio of 40/60, which might be important for PG biosynthesis by Serratia marcescens [24]. In other studies, the culture media were established for Serratia marcescens TNU02 fermentation using demineralized crab shell powder (de-CSP) [2]. The suitable formulation for high PG productivity (4514 mg/L−1) contained a protein/de-CSP ratio in the range of 3/7 to 4/6 [2]. Two other works also used the byproducts of shrimp processing for fermentation [30,31]. Demineralized shrimp shell powder [30] and shrimp head powder [31] were potential marine byproduct sources for PG production, with high PG yields of 5910 mg/L−1 and 5355 mg/L−1, respectively. Overall, fishery processing byproducts hold a promising potential when used as the main C/N source for PG production.
- Agro-Industrial Byproducts
Production of natural microbial pigments via fermentation technology is an attractive research target that is featured by high productivity and low cost [77]. Agro-industrial byproducts (AIBs) serve as an ideal substrate for microbial pigment production [77]. AIBs are various residues from the agricultural and food industries [39,78]. Reusing these residues is considered necessary because these sources, if untreated, are estimated to create waste of up to 3.40 billion metric tons by 2050 [50,79]. Thus, various materials from AIBs are also used as C/N sources for fermentation to produce the red pigment PG (Table 1).
Cassava wastewater and peanut oil cake (POC) were considered as two potential AIBs for PG production, and their effectiveness was proven in several reports. Cassava is one of six vital agricultural products worldwide, and the cassava processing industry is also very developed [32,80]. The processing of 1 ton of cassava releases around 300 L of cassava waste [80], which also contains cassava wastewater (CWW). This wastewater source is rich in organic components and potentially causes waste and pollution issues if it is directly released into the environment [81]. Thus, reusing CWW has attracted much interest. Considering that this source contains nutrients that are suitable for microbial growth, some studies utilized CWW as the substrate for fermentation to produce PG [32,66]. Moreover, de Araújo et al. [66] reported that S. marcescens UCP1459 fermentation obtained a high PG yield of up to 49,500 mg/L−1 in a medium using 6% CWW and 2% mannitol. Recently, Tran et al. [32] also recorded a potential PG productivity of 5202 mg/L−1 by utilizing CWW as the main substrate for the culture. POC is also a byproduct collected after oil pressing, and it accounts for over 50% of the substance compared to the original material [33]. This POC byproduct contains very rich nutrients that are suitable for fermentation. POC was also reused as a C/N source for PG production in some works [33,67,68]. According to Vijayalakshmi et al. [67], POC is an inexpensive source, better than other commercial materials for PG production, that could obtain a maximum PG yield of 40.9 mg/L−1 in a 5% POC medium. In another report by Naik et al. [68], PG was produced using different agro-wastes, including POC, safflower seed oil cake, cotton seed oil cake, coconut oil cake, and sesame oil cake. When used in agar medium, POC was confirmed to be the most suitable substrate for PG biosynthesis. Furthermore, the PG yield in liquid medium with 8% POC was found to be very promising, achieving up to 39,800 mg/L−1 productivity of PG [68]. The latest work by Nguyen et al. [33] also demonstrated the potential of POC in PG production. Notably, after the optimization of culture ingredients, the final medium was designed to be very simple, containing 1% POC with no other commercial supplements, while still capable of producing a high PG content of 5380 mg/L−1 [33]. Hence, POC may be a promising, low-cost C/N source with a simple culture formulation for PG production.
Some other sources also showed effectiveness for PG production. The soybean meal medium enhanced the PG yield by 5.19-fold compared to the commercial control medium, achieving an optimal yield of 9632 mg/L−1 [69]. PG was produced by Serratia marcescens UCP 1549 using corn bran as an inexpensive substrate for fermentation and achieved a PG yield of 1680 mg/L−1 [70]. Earlier, ram horns (a byproduct from the meat industry) were also utilized for cost-effective PG production, and it was observed that this substrate promoted both bacteria growth and PG yield (277.74 mg/L−1) [71]. Brown sugar was also found to be an inexpensive substrate for PG biosynthesis by Serratia marcescens UTM1, and the optimal PG yield was up to 8109 mg/L−1 [72]. Some studies used raw materials and wastewater for PG production by Streptomyces fusant NRCF69 and found that dairy processing wastewater broth used singly could yield 36,700 mg/L−1 of PG. In addition, the pigment content reached up to 47,000 mg/L−1 when supplemented with 0.5% mannitol [64].
- Other Organic Byproducts/Wastes for Prodigiosin Production
The low-cost and eco-friendly targets for PG production were also examined using other organic byproducts/wastes. Recently, Atim et al. [34] utilized feather and waste frying oil for PG production and achieved a high PG yield of 9660 mg/L−1. Some food waste and fibers are also utilized for PG production [73]. Various fibers, including rice husk, coconut fiber, sawdust, palm oil fiber, sugar cane bagasse, and tender plantain waste, were added to the solid medium used for fermentation. For example, rice husk combined with food waste creates an ideal solid medium for PG biosynthesis that can achieve a PG yield of 7890 mg/L−1 [73]. Tannery solid waste fleshing was also used as the main nutrient for PG biosynthesis, which showed a potential productivity of up to 33,000 mg/L−1 using a medium containing 3% substrate [74]. Xia et al. [75] used kitchen waste as a nutrient for PG production and achieved a yield of 223 mg/L−1 in a medium containing kitchen waste supplemented with 1% peptone and 0.2% proline.
LSF utilizes diverse substrates for fermentation. Considering that many commercial materials are not cost-effective and have low PG productivity, several organic sources have been considered as potential substrates for PG production with high yields in the range of 525–42,030 mg/L−1. Notably, although byproducts/wastes are disposal sources, they are surprisingly effective with outstanding pigment productivity, some of which achieve yields as high as 49,500 mg/L−1 of pigments. In a review, Han et al. [37] reported that PG production using a low-cost substrate could significantly enhance PG yield by around 1.35–33.7-fold. These results also led to novel research that developed the value of inexpensive materials in the microbial production of potentially bioactive compounds. In addition to advancing economic value, the utilization of byproducts/wastes also helps to avoid the surplus of these nutrient-rich sources and alleviates environmental burdens.
2.2. Overview of the Substrate Sources for Prodigiosin Production in Solid-State Fermentation
PG production using the LSF method is popular in research. However, solid-state fermentation (SSF) has also been conducted and assessed for PG production in several investigations (Table 2) [51,82,83,84,85,86]. SSF was conducted on a solid and insoluble substrate with very low moisture (lower limit of 12%), and microorganisms adhered to the solid substrate [87]. Normally, the substrates used in SSF are organic and inexpensive materials which have several advantages including cost-effective fermentation, excellent productivity, and simple technology [87,88,89]. Some reports on PG production using SSF show its potential for PG productivity [51,82,83,84,85,86]. Recently, Nguyen et al. [90] used peanut powder for SSF in PG production and reported that the mutant strain S. marcescens EMS5 supported up to a 1.52-fold improvement in PG yield compared to the wild strain. Khalid et al. [51] screened some substrates for cultivation and found that wheat bran medium obtained the highest PG yield of 47.5 mg/g−1 during early fermentation. Furthermore, a maximal yield of PG (240 mg/g−1) was achieved in an optimal medium containing wheat bran supplemented with 0.5 mL/g−1 of sunflower oil and 0.4 mL/g−1 of live Bacillus subtilis cells [51]. Tannery fleshing waste was also used as a solid medium for PG production with a high yield of 70.4 mg/g−1 [84]. Another study used a mannitol solid medium and found it to be a promising culture medium for microbial fermentation and PG, yielding a productivity of 3.2 mg/g−1 [85]. Some agro-industrial wastes were also utilized as solid mediums for PG biosynthesis [86]. Therein, wheat bran exhibited a maximum PG productivity of 1.307 mg/L−1, whereas the other assessed media yielded PG in a lower range from 0.008–0.066 mg/L−1 [86]. Other reports attempted to use SSF for PG production using agro-industry wastes such as wheat bran, sugarcane bagasse, instant noodle waste, tangerine peels, pineapple peels, and pineapple crown for fermentation [82]. Among those agro-industry wastes, wheat bran medium showed the highest PG yield. The PG productivity in the optimal medium (5 g wheat bran and 5% waste soybean oil) was recorded at 119.8 g/kg−1 dry substrate [82]. PG was also produced by SSF using byproducts, including bagasse, wheat straw powder, and wood chips, as a medium [83]. They recorded bagasse to be the best solid medium for PG production with a yield of 20.13 g/kg−1 dry solid, with the PG productivity in optimal medium reaching up to 40.86 g/kg−1 dry solid (1.17 g/g−1 bagasse, glycerol, and 0.33 g/g−1 bagasse soy peptone) [83].
Both the LSF and SSF methods are very commonly used to produce secondary metabolites from microorganisms. These fermentation methods have some advantages and disadvantages [91]. SSF showed high potential for productivity and low production cost; however, the parameters and process of fermentation were rather difficult to control [92]. These issues have limited the use of SSF on a large scale. Nowadays, productivity can be improved using the LSF method by supplementing some yield-enhancing factors during cultivation and reducing the cost, which can be achieved by using inexpensive substrates or byproducts/wastes for fermentation. Notably, fermentation by the LSF method may be performed and scaled up via bioreactor systems to obtain a high PG yield within a significantly reduced cultivation time [2,29,30,31,32,33]. In the future, more research is needed to identify other novel materials that can be used as substrates for fermentation. To date, only fishery processing byproducts have been used in LSF. Thus, there is a need to expand these findings by using other nutrient-rich byproducts that are available in abundant amounts, such as those obtained from food or agro-processing.
2.3. Enhancement of Prodigiosin Production by Supplementing with Nutrients
- Amino Acids
The synthesis of precursor 4-methoxy-2,2-bipyrrole-5-carbaldehyde (MBC) of PG requires the presence of some amino acids, such as proline, serine, and methionine, in the transformation pathway [93,94]. Thus, supplementing these amino acids may promote PG production. Qadri et al. [95] studied the role of methionine in PG biosynthesis via the methylation of pigment. They also found that the addition of a single methionine did not cause pigment production. This process was stimulated for the supplementation of methionine with other amino acids. However, methionine helped to shorten the lag period in the PG biosynthesis process leading to an increase in its productivity [95]. Williams et al. [96] reported a positive impact of amino acids on the improvement of PG production using four amino acids of casein hydrolysate, including l-glutamic acid, l-proline, dl-aspartic acid, and l-alanine. When supplied individually, the amino acids were found to facilitate the synthesis of pigment [96]. Faraag et al. [97] found that serine supplementation could inhibit PG synthesis, whereas tyrosine showed better stimulation than alanine and proline for PG production. The effect of methionine, leucine, proline, alanine, and their combination on PG production was assessed by Siva et al. [98]. They reported that the combination of proline and methionine supported the highest PG yield, which was a 3-fold increase in PG yield compared to the control group.
- Fatty Acids
Another precursor of PG, 2-methyl-3-n-amylpyrrole, is mainly biosynthesized by the oxidation of fatty acids [94]. The addition of some fatty acids from oils could improve PG productivity. According to the results of Chenqiang et al. [56], supplementation with 10 mL/L−1 olive oil can enhance PG yield by 9.3-fold compared to the initial medium. In addition, this was the first study to note that 60% PG in pellet form is easier to extract. Giri et al. [61] studied fatty acids in seeds and oils and their effects on improving PG productivity. Their results showed that peanut medium led to a 40-fold increase in PG yield, and three oils (sesame oil, peanut oil, and coconut oil) also improved PG yield, as compared to the commercial medium. The effect of some crude fatty acid sources from groundnut, peanut, sesame, castor, and sunflower seeds on PG yield was explored by Picha et al. [99]. In their tests, the highest PG yield was achieved in media containing crude fatty acid sources from peanuts or sunflowers. A report by Parani et al. [100] noted higher PG productivity when the commercial medium was supplemented with the 4% vegetative oil mixture (coconut, sunflower, and olive oil). Another study by Wei et al. [53], on the effect of the oil-supplement strategy for enhancing PG yield, found that PG productivity improved with oil supplements used at 2–6% concentrations. Moreover, sunflower oil increased the PG content up to 14-fold in comparison to LB broth [53].
- Microbial Cells
In nature, microbes exist in a complex population, and their interactions are also a factor to be considered for enhancing the production of secondary metabolites [101,102,103]. Some pigment-producing bacterial strains may increase PG production through their interactions with the living/dead cells of other microbes. PG is anti-bacterial; thus, it can protect the host bacteria from other symbiotic bacteria in culture media. The dead cells of those symbiotic bacteria do not consume nutrients, and so also have no effect on the pigment-producing strain. According to Khalid et al. [51], in a medium with live cells of Escherichia coli, B. subtili, or Saccharomyces cerevisiae, there was a 1.8–2.3-fold increase in PG yield compared to the control medium. Moreover, the heat-killed cells of these microbes also achieved a 1.1–2.1-fold increase in PG yield. Mahmoud et al. [104] reported enhanced PG yield after adding the following bacteria to culture media: E. coli cells achieved a 3.8–9.1-fold increase in PG yield, B. subtilis cells achieved a 6.7–7 fold increase in PG yield, and S. cerevisiae cells achieved about a 2.2–9-fold increase in PG yield. Huy et al. [105] used dead cells of Lactobacillus rhamnosu as substrate at different temperatures (70 °C and 100 °C) for PG production. They found that adding 50 µL of heat-killed L. rhamnosu cells at 70 °C harvested the best pigment content of 9.79 mg/L−1, which was enhanced by 6-fold compared to the system without L. rhamnosu. However, killing the L. rhamnosu cells at 100 °C tended to reduce pigment production [105]. The mechanisms of these impacts are still unclear. It is possible that the addition of microbes led to the release of some compounds that are effective in enhancing PG production but that have not been identified yet. Alternatively, the direct contact between the PG-producing strain and additional strains could have stimulated PG biosynthesis [51]. One of the reasons for the enhanced PG production associated with the addition of bacterial cells might be related to the adsorption ability, as reported by Wang et al. [106]. They assessed the effect of four Lactobacilli strains (TKU 010, TKU 012, BCRC 12193, and BCRC 14011) on the ability of Serratia marcescens TKU011 to produce PG in media containing squid pen powder. Among them, Lactobacillus paracasei TKU 012 cells showed the highest adsorption rate of up to 84% for the hydrophobic pigment. The PG yield also increased proportionately with the concentration of added TKU 012 cells.
- Mineral Salts
Mineral salts also affect PG productivity. Sulfate and phosphate salts are popularly used in PG-producing media. In a study that used a factorial design to examine the role of four mineral salts in PG biosynthesis by Serratia Marcescens BS303, the results showed that NaCl decreased PG content to nearly 20%, whereas a mixture of ammonium iron (III) citrate and anhydrous copper showed the highest PG yield, which was enhanced by 1.8-fold [107]. Suryawanshi et al. [23] explored the effect of some inorganic salts on PG production. Changes in the concentration of NaCl and CaCl2 did not affect PG content, but there was evidence that these salts possibly caused some inhibition of PG biosynthesis at concentrations greater than 2 g/L−1. This study also found the following: (1) CaCO3 salt did not change the PG yield at concentrations of 10–100 mg/L−1; (2) KH2PO4 salt showed a maximum PG yield at 80 mg/L−1 but reduced PG production if the concentration continued to increase; (3) FeSO4 salt produced a stable amount of PG at 30–50 mg/L−1, but the production was significantly reduced at concentrations over 50 mg/L−1; and (4) MgSO4 salt at 10–200 mg/L−1 led to an increase in PG productivity [23]. Iron is an essential factor for PG production but not for the growth of bacteria [108]. Allen et al. [109] confirmed that the concentration of Fe2+ at 0.14 mmol was the most suitable for developing and producing PG by Vibrio gazogenes fermentation. Several studies by Nguyen et al. [2,24,29,30,31,32] showed a 1.3-fold higher yield of PG in media supplemented with various types of sulfate and phosphate salts, and the optimal concentration was as little as 0.002–0.05% for sulfate and 0.025–0.1% for phosphate. According to these results, some new formulations of sulfate and phosphate salts were recorded for enhancing PG yield [2,24,30,31,32]. Nguyen et al. also determined the different optimal formulations of salt using the same bacterial strain (Table 3). Disparities in the optimal salt formulations are possibly due to the difference in nutrient composition of the main substrate used. Research using POC as the main C/N source showed that the supplementation of other ingredients is not necessary when the POC substrate contains enough essential nutrients for fermentation [24].
However, some stimulative salts can inhibit PG production when present in excessive concentrations. Silverman et al. [108] reported that PG production using both the condensation reaction and synthesis reaction of MBC was sensitive to NaCl and that high concentrations of NaCl (3%) inhibited PG biosynthesis. The same result was observed when Na2SO4, KCl, and K2SO4 were used at the same concentration. In some reports, the use of phosphate at high concentrations also reduced PG productivity. Witney et al. [110] found that phosphate concentrations between 10 and 250 mM significantly reduced PG yield. Allen et al. [109] reported a similar decrease in PG yield in the presence of KH2PO4 at concentrations greater than 0.4 mM.
- Other Factors
The supplementation of 5% mineral oil enhanced pigment content by 1.12-fold, as observed by Ulises et al. [62]. The role of α-chitin from shrimp shells and β-chitin from squid pens for PG biosynthesis was reported by Nguyen et al. [24]. They found that α-chitin provided the greatest PG yield (3230 mg/L−1), which was higher than the yield produced when supplemented with β-chitin (2730 mg/L−1) and control squid pens (2450 mg/L−1) [24]. The addition of several sugars into the medium also affects PG production. Lactose supplementation enhanced PG productivity by 18.67% [57]. In another report, sucrose was found to increase PG yield from 8.567 g/L−1 to 9.632 g/L−1 [69]. According to Giri et al. [61], the addition of maltose or glucose to nutrient broth led to only a 2-fold increase in PG yield, whereas no change was observed for the sesame seed medium. However, in some reports, glucose was confirmed as an inhibitory factor for PG biosynthesis [111,112,113]. These differences in glucose results may depend on the strain and ingredients of the culture medium.
Overall, the supplement factors, such as amino acids, fatty acids, microorganism cells, mineral salts, and other factors, assist in improving PG productivity from 1.1 to 9.2-fold (Table 3). Thus, besides finding suitable substrates for fermentation, the determination of additional factors is also considered an essential strategy for efficient PG production via fermentation.

{kind=link}
Table 3.
Some supplement factors that assist in enhancing prodigiosin yield.
| Supplement Ingredient | PG Yield | Unit | Enhancing Yield (Fold) | Reference | |
|---|---|---|---|---|---|
| Initial Medium | Modified Medium | ||||
| dL-alanine (5 mg/L−1) | 12.7 | 19.2 | mg/L−1 | 1.5 | [96] |
| l-glutamic acid, l-proline, dl-aspartic acid, and l-alanine (5 mg/L−1) | 20 | 1.6 | |||
| l-glutamic acid, l-proline, dl-aspartic acid, and l-alanine (10 mg/L−1) | 22.1 | 1.7 | |||
| l-tyrosine (10 mg/L−1) | 2.78 | 8.87 | mg/L−1 | 3.2 | [97] |
| Soybean oil (10 mg/L−1) | 1735 | 9283 | mg/L−1 | 5.4 | [56] |
| Canola oil (10 mg/L−1) | 7142 | 4.1 | |||
| Olive oil (10 mg/L−1) | 11,366 | 6.6 | |||
| Maize oil (10 mg/L−1) | 9119 | 5.3 | |||
| Peanut oil (10 mg/L−1) | 9539 | 5.5 | |||
| Tea oil (10 mg/L−1) | 10,348 | 6.0 | |||
| Soybean oil (4%) | 152 | 525 | mg/L−1 | 3.5 | [53] |
| Sunflower oil (6%) | 790 | 5.2 | |||
| Olive oil (4%) | 579 | 3.8 | |||
| Live cells of E. coli (0.4 mL/g−1) | 100.47 | 225 | mg/g−1 | 2.2 | [51] |
| Live cells of B. subtili (0.4 mL/g−1) | 240 | 2.4 | |||
| Live cells of S. cerevisiae (0.4 mL/g−1) | 170 | 1.7 | |||
| Dead cells of E. coli (0.4 mL/g−1) | 104.47 | 170 | 1.6 | ||
| Dead cells of B. subtili (0.4 mL/g−1) | 203 | 1.9 | |||
| Dead cells of S. cerevisiae (0.4 mL/g−1) | 120 | 1.1 | |||
| Dead cells of L. rhamnosus | 1.43 | 9.79 | mg/L−1 | 6.8 | [105] |
| Live cells of E. coli | 450 | 2500 | mg/L−1 | 5.6 | [104] |
| Live cells of B. subtili | 600 | 1.3 | |||
| Live cells of S. cerevisiae | 2800 | 6.2 | |||
| Dead cells of E. coli | 4100 | 9.2 | |||
| Dead cells of B. subtili | 3500 | 7.8 | |||
| Dead cells of S. cerevisiae | 4100 | 9.1 | |||
| 0.05% MgSO4, 0.03% K2HPO4, | 2450 | 2980 | mg/L−1 | 1.2 | [29] |
| 0.02% K2SO4, 0.05% K2HPO4 | 3980 | 5200 | mg/L−1 | 1.3 | [30] |
| 0.02% K2SO4, 0.025% Ca3(PO4)2 | 3862 | 4500 | mg/L−1 | 1.2 | [31] |
| 0.02% (NH4)2SO4, 0.1% K2HPO4 | 3010 | 4000 | mg/L−1 | 1.3 | [2] |
| 0.05% MgSO4, 0.1% K2HPO4 | 3981 | 5202 | mg/L−1 | 1.3 | [32] |
| 0.05% K2HPO4, 0.1% CaSO4 | 3230 | 4320 | mg/L−1 | 1.3 | [24] |
| α-chitin (5/3 w/w) | 2450 | 3230 | mg/L−1 | 1.3 | [24] |
| β-chitin (2/6 w/w) | 2450 | 2730 | mg/L−1 | 1.1 | [24] |
| Lactose | 15 | 17.8 | mg/L−1 | 1.2 | [57] |
| 0.864% Sucrose | 8.567 | 9.632 | g/L−1 | 1.1 | [69] |
| 0.5% Maltose | 520 | 1836 | mg/L−1 | 3.5 | [61] |
| 0.5% Glucose | 1689 | 3.2 | |||
2.4. Scaling-Up Production of Prodigiosin
Bioreactors are used for the large-scale production of secondary metabolites [114]. In large-scale PG production, the bioreactor system has been used. Some fermentation strategies in bioreactors include batch processes, fed-batch processes, and continuous cultivation [29,115,116]. Among those, batch processes are mainly used for research on PG production [29,30,31,32,33,67,68,72,117,118,119,120] because they provide several advantages including a short duration, unlikely contamination as no nutrients are added, and an easily managed culture process [121]. The fed-batch process is a partly open system, also called a semi-continuous bioreactor, characterized by the continuous or intermittent addition of the required nutrients to the initial medium after the start of fermentation or from the mid-period of the cultivation process. The advantage of this process is that it achieves a higher overall product content. Tao et al. [115] designed a two-step culture process, wherein the first stage used glucose as the main carbon source for cell growth, and in the next stage, PG was produced by switching glucose to glycerol in the medium. A 5-L bioreactor yielded a PG production efficiency that was 7.8 times higher (583 mg/L−1) than the original cultivation mode (75 mg/L−1) with glycerol as the sole carbon source [115]. After a batch growth phase, an equilibrium is established with respect to a particular component (also called a steady state). The fresh medium is added into the batch system along with a corresponding withdrawal of the medium containing the product at the exponential phase of microbial growth. These bioprocesses are referred to as continuous cultures. Continuous cultivation gives a near-balanced growth, reduces product inhibition, and improves space-time yield [121]. However, the long cultivation period also increases the risk of contamination and may lead to long-term changes in the cultures, including the impact on the product yield. In a study, PG was biosynthesized by continuous fermentation of Hahella chejuensis in 10 L and 200 L bioreactors with glucose (2.5 g/L−1) as the most suitable carbon source. The 10 L bioreactor achieved 2280 mg/L−1 of PG yield in 49 h, whereas the 200 L system only recorded a PF yield of 1305 mg/L−1 after 10 days of fermentation [116]. Luis et al. [122] performed a continuous fermentation, and the PG content was 1.2-fold higher compared to that obtained in the batch mode. A continuous fermentation was performed with a 5-fold higher nutrient concentration than that of the initial media to take advantage of a large amount of foam from fermentation. The result was a 2.7-fold increase in PG yield compared to that obtained by batch fermentation [117].
Several bioreactor systems have been examined for PG production. Therein, nearly all research systems have used laboratory-scale bioreactor systems with small and medium capacities from 2.5 to 20 L [29,30,31,32,33,66,67,68,72,115,118,119,120,122,123]. On a larger scale, Qi et al. [117] performed an experiment in a 50 L bioreactor using 47.8 L of a medium. PG was also produced on an industrial scale [116], wherein continuous fermentation was carried out in two different fermenters of sizes 10 L and 200 L. Similar to flask fermentation, S. marcescens is the most popular and widely used strain for fermentation in a bioreactor [29,30,31,32,33,66,67,68,72,115,117,118,119,122,123]. Although, Jeong et al. [116] used Hahella chejuensis for PG production.
To achieve a large-scale, lower cost PG production and reduce environmental pollution, some studies utilized byproducts from food processing or agriculture for fermentation [29,30,31,32,33,67,68,72,123]. In addition to several commercial substrate sources popularly used in fermentation, such as nutrient broth, maltose, peptone, casein, sucrose, glycerol, and glucose [115,116,117,118,119,122], PG was also biosynthesized by utilizing inexpensive byproduct/waste [29,30,31,32,33,67,68,72,123,124]. Nguyen et al. utilized some byproducts and wastes, such as squid pens [29], shrimp shells [30], shrimp heads [31], crab shells [2], and agriculture processing byproducts, including POC [33], cassava wastewater [32] for PG production by fermentation. Vijayalakshmi et al. [67] and Naik et al. [68] used POC as a C/N source for PG production. Aruldass et al. [72] designed a medium containing brown sugar, a kind of byproduct from sugar processing. Compared to commercial substrates, the efficiency of creating PG from these low-cost materials was relatively better and sometimes higher. The yield of PG increased from 583 to 13,100 mg/L−1 when using commercial medium, whereas using the byproduct substrate for fermentation showed an average PG yield of about 6000 mg/L−1 and a maximum of 40,000 mg/L−1 (based on the results synthesized in Table 4).
Fermentation in a bioreactor is also efficient in terms of productivity and shorter time compared to flask cultures. Nearly all research on PG production in bioreactors reported a 1–76.7-fold increase in the yield of PG. Nguyen et al. [2,29,30,31,32,33] reported that fermentation in a bioreactor only required a short time to reach the maximum level of PG expression. Normally, a small-scale fermentation (in the flask) takes up to 2 days to reach the maximal yield, whereas a large-scale fermentation (in a bioreactor) requires only 8–10 h.
Overall, when PG is produced on a large scale, efficiency is achieved in terms of both yield and production time. To date, there have been several reports on PG production; however, almost all have been conducted on a small scale, and there are only a few reports on large-scale PG production. Moreover, nearly all studies have used the bioreactor system on a laboratory scale, and more research is still needed to assess the potential for industrial scale PG production. In particular, other kinds of byproducts or waste for fermentation should be screened for better biosynthesis efficiency and cost effectiveness.
3. The Potential Applications of Prodigiosin in Agriculture
In this section, the bioactivities related to the potential applications of PG in agriculture are presented and discussed. Focus is given to the anti-fungal, anti-bacterial, anti-insect, and anti-nematode effects via in vitro tests, in the greenhouse, and in the field.
3.1. The Potential Applications in Agriculture: In Vitro Tests
3.1.1. Anti-Fungal Activity of Prodigiosin
Many harmful fungi have been reported on crops [125,126]. The anti-fungal activity of PG makes it a potential candidate for use in the agricultural industry to protect crops from fungal harm (Table 5) [23,74,100,124,127,128,129,130,131,132,133].
Most studies focused on the activity of PG in its purified form. Hiroshi et al. [131] reported the inhibitory capacity of PG against 20 species of plant-pathogenic fungi belonging to six genera. At a PG concentration of 10 µg/mL−1, the growth of all tested fungal species was inhibited, some by more than 80% [131]. Nobutaka et al. [129] observed that PG controlled the growth rate of Botrytis cinerea, a gray mold fungus of cyclamen. The spore germination rate was reduced by one-third at 1 µg/mL−1 of PG compared to the treatment using an enzyme mixture. Duzhak et al. [127] assessed the inhibitory ability of PG against a pathogen fungus of raspberries (Didymella applanata) and found that PG was the main factor that suppressed fungal growth at an IC50 value of 2.5 nmol/mL−1. The role of PG and chitinase from Serratia marcescens against the fungus D. applanata was also assessed, and they found that PG was a key anti-fungal agent, with an IC50 value of 2.5 nmol/mL. In contrast, the chitinase production did not provide the bacterium with any competition for the fungus [127]. Suryawanshi et al. [23] studied the potential activity of PG against two pathogenic fungi, including Aspergillus flavus and Fusarium oxysporum, and observed that their minimum inhibitory concentrations (MIC) were 10 µg/mL−1 and 8 µg/mL−1, respectively. Another report studied the inhibitory activity of PG against A. niger and F. moniliforme and found their MIC values to be 230 µg/mL and 210 µg/mL, respectively [74]. In the work of Martha Ingrid et al. [130], PG showed an efficient inhibitory activity against Mycosphaerella fijiensis, a fungus that causes black Sigatoka disease in bananas. At a concentration of 14.3 µg/mL−1, this pigment inhibited 60% of the growth of germ tubes, and the IC50 value was 996 µg/mL−1 [130]. Sagar et al. [124] tested the bioactivity of PG against two popular fungi, Alternaria and Fusarium, at MIC values of 80 μg and 160 μg, respectively. The red pigment was also tested by Samer et al. [128] for the inhibition of two types of fungi: the agent of blackleg disease and white mold on crops. They observed that 50 µM PG significantly inhibited the growth of Phoma lingam with nearly 25% hyphal growth, whereas its effect on Sclerotinia sclerotiorum w relatively insignificant [128]. Most recently, in 2022, there was a report of antifungal activity by purified PG from Serratia rubidaea Mar61-01 against Colletotrichum nymphaeae, the agent that causes anthracnose disease in strawberries. PG showed an ability to inhibit the germination of this fungi by up to 100% [133].
In some studies, the crude extract of the PG pigment was used to evaluate its anti-fungal activities. Parani et al. [100] evaluated the effect of crude PG from Serratia marcescens SR1 on the inhibition of some pathogen fungi using the well-diffusion method. Therein, crude PG showed a maximum inhibitory zone (42 nm) against Helminthosporium sativum and five other types of fungi including, in decreasing order, Curvularia lunata (40 mm), Alternaria alternate (40 mm), Fusarium oxysporum (30 mm), Cercospora apii (24 mm), and Rhizoctonia solani (11 mm) [100]. Jimtha et al. [132] evaluated the anti-phytopathogenic activity of crude extract from S. marcescens Bm1 and observed significant growth inhibition of Pythium myriotylum (71.33%), whereas the inhibitory activity against other fungal strains, such as Phytophthora infestans, Sclerotium rolfsii, and Rhizoctonia solani, was less than 62%. The crude pigment was also found to be effective against Fusarium oxysporum (31% inhibition), although the effect was non-significant [132].
PG is an effective, promising candidate against the inhibition of many fungal species. Many strains, including those of Fusarium sp. and Phytophthora sp., were tested. However, further research is needed to assess the impact of this pigment against other harmful phytopathogens. To date, almost all studies have used purified PG for tests, whereas only a few have used the crude extract to assess the potential activity of PG. Furthermore, the direct treatment by crude extract may possibly have more advantages. Thus, the agricultural bioactivity of the crude pigment should be more exhaustively evaluated.
3.1.2. Prodigiosin Inhibits Other Organisms That are Harmful to Crops
In addition to its inhibitory effect against phytopathogenic fungi, PG is also active against other pathogens, such as bacteria, nematodes, and insects, that are harmful to crops. Several studies showing the bioactivity of PG against these pathogens are summarized in Table 6 [33,65,128,131,134,135,136,137,138,139].
Hiroshi et al. [131] tested the antibacterial activity of PG against plant pathogenic bacteria belonging to 19 species in six genera. PG was the most active against Clavibacter michiganensis subsp. michiganensis with the maximal allowable concentration (MAC) of 6.3 µg/mL−1 [131]. PG was also found to be active against the following bacteria: Erwinia carotovora subsp. carotovora and Xanthomonas campetris pv. oryzae with MAC values of 25 µg/mL−1 and 25.5 µg/mL−1, respectively; and four strains including Acidovonax avenae, Agrobacterium tumefaciens, Erwinia herbicola, and X. campetris pv. carotae that were inhibited at similar MAC values of 50 µg/mL−1. PG displayed a weaker activity against the other tested strains with MAC values greater than 100 µg/mL [131]. Research related to the objective of using PG against plant pathogenic bacteria is still very limited despite the fact that plant bacterial pathogens are spreading widely across the globe [140,141]. PG is also known as a potential inhibitor for many gram-negative and gram-positive bacteria; thus, its potential anti-bacterial role is relatively important.
Plant-damaging nematodes can seriously damage important crops across the world [142]. They can reduce the productivity of crops by 5–10% in developed countries [143] and reduce the quality of agricultural products [144]. Therefore, finding a solution to manage these disease nematodes is necessary [145]. PG has the potential to inhibit some of these crop-damaging nematode species. Samer et al. [128] assessed the impact of the Prodiginine group on the plant parasitic nematode Heterodera schachtii; only PG caused considerable paralysis of the second-stage juveniles (J2s) with an IC50 value of 13.3 μM, whereas the remaining compounds were less effective against J2s [128]. This pigment was also effective against the juvenile stage of Meloidogyne javanica and Radopholus similis at low concentrations, with IC50 values of 79 and 83 µg/mL−1, respectively [134]. At the same tested concentrations, the PG pigment could completely inhibit egg-hatching, and no juveniles were observed [134]. Some reports were conducted to assess the activity of PG against Meloidogyne incognita. The culture filtrate of Serratia marcescens was used for testing the activity of newly hatched juveniles of Meloidogyne incognita after 72 h of treatment [135]. The crude pigment was effective against this nematode even at a low concentration, with an IC50 value of 31.9 mg/mL−1 [135]. This proved the effectiveness of using the microbial secondary metabolites, in comparison with the use of whole organisms. Recently, Nguyen et al. [33] studied the activity of purified PG for Meloidogyne incognita on both the J2s and egg-hatch. They found that the purified compound showed a stronger inhibition compared to the crude pigment, and the IC50 was as low as 0.2 mg/mL−1. These results showed the novel inhibitory effect of PG against the egg-hatching of this nematode with an IC50 value of 0.32 mg/mL−1 [33].
Some studies used the inhibition of PG to target harmful plant insects. Wei et al. [136] evaluated the potential activity of PG against Asian Citrus Psyllid. The toxicity of PG against nymphs depended on temperature. Based on the test results, the most suitable temperature was confirmed to be 30 °C, which was used for follow-up experiments. PG was significantly effective against the oviposition and egg-hatching of Diaphorina citri with an inhibitory rate of 42%. PG also exhibited a moderate inhibitory rate against egg-hatching at a rate of 26%. Moreover, treatment against adult hoppers with an LC20 and LC50 solution of purified PG at 30 °C was recorded to excrete less honeydew, as compared to the control [136]. This pigment also demonstrated insecticidal effects against Helicoverpa armigera and Spodoptera litura larvae, with mortality rates of 70 and 100% at 20 and 30 mg/mL−1, respectively [137]. The activity of pure PG against Drosophila larvae was reported in some studies [65,138]. According to Wang et al. [65], after 5 days of treatment with pure PG, the IC50 value was recorded at 0.23 g/L−1. Furthermore, to confirm the role of PG against Drosophila, the PG concentration in each test formula showed high insecticidal activity, which corresponded to a high PG concentration [65]. In a study by Liang et al. [138], PG presented effective toxicity against Drosophila with an IC50 value of 230 ppm compared to other commercial food colorants. Asano et al. [139] accessed the anti-insect ability of PG against three kinds of insects, including Spodoptera litura (a common cutworm), Plutella xylostella (the diamondback moth), and Adoxophyes honmai (the tea tortrix). PG showed the highest activity against Plutella xylostella with high mortality of up to 96% at an 8 µg/g−1 diet [139]. Spodoptera litura and Adoxophyes honmai were less sensitive to PG, with the mortality only reaching 3% and 12%, respectively [139].
These summarised study results show that the anti-fungal effect of PG was frequently confirmed; however, only a few studies mentioned other bioactivities. Thus, more studies are needed to explore the activities of PG on many other objects. This review enriches the existing information about the future bioactive and potential applications of PG. Recently, there has been a push to develop green and sustainable agriculture, whereby natural biological inhibitory agents are considered effective and safe solutions for plant disease management. PG possesses the ability to inhibit many harmful agents and may become a potential candidate for controlling various plant diseases in the future of agriculture production.
3.2. The Potential Applications of Prodigiosin via Studies in Greenhouses and in the Field
The effect of the culture fluid from Serratia rubidaea Mar61-01 on the growth of Colletotrichum nymphaeae (the causal fungi of strawberry anthracnose) was tested in vivo and in greenhouse conditions [133]. In the in vivo condition, the culture fluid containing pigment significantly decreased fruit decay with a bio-control efficacy of 48.60%. The tested sample also showed a potential effect in the greenhouse test for strawberry anthracnose reduction. The spraying method of culture fluid was highly effective, with a 72.22% decrease in strawberry anthracnose disease compared to the drenching soil method, which only reduced the disease by 44.44% [133]. In a report by Roberts et al. [146], the purified PG from Serratia rubidaea was used to determine the damping-off suppression of cucumber caused by Pythium ultimum fungus. In the presence of Pythium ultimum inoculum, the cucumber plants with seeds that were treated by PG showed a greater development compared to the nontreated plants. In addition, the ethanol extract of mutant strain Tn246 non-producing PG could not control the disease caused by this pathogen. This study helped to clearly understand the role of PG in the management of these phytopathogens [146].
Samer et al. [128] evaluated the bioactivity of prodiginines against the first stage of plant parasitic nematodes (Heterodera schachtii) during the growth of a plant using Arabidopsis thaliana as the model plant. The infection results revealed that treatment with PG reduced the total number of H. schachtii individuals that developed in the Arabidopsis thaliana plant by up to 50%. Furthermore, the size of female nematodes and their associated syncytia were smaller after processing. PG also promoted the growth of the plant depending on treatment concentration. Some reports assessed the role of PG against the knot-root nematode Meloidogyne incognita in greenhouse conditions [135,147]. Omnia et al. [135] used the culture broth and culture filtrate of Serratia marcescens for testing Meloidogyne incognita inhibition in vivo on tomato seedlings. Both applications effectively suppressed the nematode population, both in the soil and root, and promoted shoot and root lengths and plant biomass, compared to that in the untreated plants. Similar results were found in the developmental stages, females, and eggs within tomato roots. Nguyen et al. [147] used the fermented product of Serratia marcescens TNU02 which is rich in PG content to evaluate the anti-nematode Meloidogyne incognita on the black pepper plant model in greenhouse conditions. The fermented product of the TNU02 strain (at the treatment dose of 80 mL) exhibited potent activity against nematodes on pepper root and soil with up to 70% and 85% mortality rates, respectively. It especially reduced the number of nematodes by 4-folds in the root nodules. In addition, the influence of the fermented product on growth targets was determined via growth parameters and chlorophyll content. Among various treatment concentrations, the use of 40 mL and 80 mL of the fermented product was more potent than other treatments at other concentrations. This seems to be a promising application of PG for the management of black pepper nematodes in agriculture. In another study, the purified PG from Serratia Marcescens KH-001 was applied to determine whether it could protect orange orchards against Asian Citrus psyllid [136]. The field experiments showed that a 10% PG emulsifiable concentration was highly effective (up to 70–100%) compared to the other concentrations studied. Furthermore, the results recorded in July and August were better than in October [136].
In addition to its potential activity in vitro, PG also showed promising impacts during tests conducted in vitro, in the greenhouse, and in the field. However, few current studies have assessed the bioactivities of this pigment on a larger scale, especially in greenhouse conditions or the field. Thus, further studies are needed on the application of PG in agriculture. To date, the evaluation of PG activity has been mainly reported in the pure compound form, with only a few studies examining its bioactivities at the nanoscale. Furthermore, nano-PG has been mainly used to assess activities in the medical and dye industries [148,149,150,151,152,153,154,155]. However, few studies have assessed PG nanoparticle bioactivities for potential use in agriculture. Currently, green synthesis by nanotechnology combined with natural active ingredients is being explored with the objective to improve the formulations of pesticides in terms of effectiveness, safety, and environmental protection [156,157,158,159,160]. Thus, investigation of the potential uses of PG in nano forms may find new ways to enhance the bioactivity and stability of PG.
3.3. Mechanisms of Potential Bioactivities of Prodigiosin
The mechanism of PG for bioactivities in medicine has been demonstrated in many reports [18,161,162,163,164,165,166,167,168,169]. The anti-cancer activity of PG was studied the most, and some mechanisms were clarified according to each type of cancer [164,165,166,167,168,169]. PG is known as a potential antibiotic for many gram-negative and gram-positive bacterial strains. The anti-bacterial capacity of prodigiosin is based on its pH modification, inhibition of cell cycles, and bacterial genetic machinery cleavage [18]. Darshan et al. [161] mentioned other mechanisms, including the formation of reactive oxygen species (ROS), whereby PG infiltrates and is localized on the membrane and the nucleus of bacteria cells. The PG then promotes a ROS reaction that damages cells and causes the death of the bacterial cells. In other reports, the anti-bacterial role of PG was proved via an induction process with autolysins, enzymes attending to hydrolysis, and the restructuring of peptidoglycan on bacteria cell walls [162]. PG causes an increase in the permeability of cell walls. It also plays a role in proton transport and ATP synthesis. Therefore, PG has become an ideal factor for the induction of autolysins, causing bacterial death [162]. Moreover, PG causes bacterial inhibition by reducing the respiration of cells and inhibiting protein and RNA synthesis [163]. Yip et al. [18] suggested that PG also inhibits two strategies that harmful bacteria use when they compete with others in the medium. PG can inhibit the growth of bacteria and virulence factors such as hemolysin, protease production, and biofilm formation [18]. The mechanism of PG differs depending on each type of microbe [162,163].
Considering its role in agriculture, the action mechanism of PG is still rarely discussed. Several reports proved the roles of PG against zygomycete and ascomycete fungi [170] and the Meloidogyne incognita nematode [33]. A study on the effect of Serratia marcescens in the inhibition of zygomycete and ascomycete fungi showed that bacteria could not infiltrate inside cell membranes and cause fungal death. However, the activity of PG increased the permeability of cell membranes, and the Serratia marcescens then easily infiltrated inside the fungal hyphae [170]. Some molecular mechanisms of anti-nematode activity were proved by the inhibition of enzyme or protein targets by virtual screening assays [171,172,173,174,175]. Nguyen et al. [33] suggested the anti-nematode role of PG (Meloidogyne incognita) via the inhibition of the enzyme acetylcholinesterase (AChE). This enzyme is often used to evaluate anti-nematode ability [172]. Based on the previous report by Nguyen et al. [31], a docking study was conducted to assess the interaction and inhibitory efficacy of the ligand toward the target enzyme. PG showed strong binding energy (−12.3 kcal/mol), and up to six linkages (interactions with amino acids) in the binding sites were found on AChE.
Overall, reports on the molecular mechanism of PG related to its bioactivities for potential application in agriculture are still very limited. Although PG was tested and promising inhibition has been recorded on many organisms harmful to crops, its mechanism of action on each object is still not explored fully. Thus, understanding the mechanism of action is needed.
3.4. The Toxicology of Prodigiosin
The safety of PG was accessed via experimental studies [23,24,25,26,27,28]. PG was found to provide efficient inhibition against various cancer cell lines but was not toxic to normal cells [24,25]. The purified PG has been shown to be non-genotoxic in biological assessments used to determine mutagenic potential, including the Ames test and in vivo micronucleus [26]. A previous report also confirmed the safety of PG and its benefits to soil biodiversity at a high dose of 500 µg/mL [23]. PG is also safe when used to evaluate toxicity in eukaryotic models, such as the nematode Caenorhabditis elegans [27] and mice [28]. Its safety for mice is a valuable demonstration of the potential application of PG in higher organisms.
A few studies have discussed the virulence of PG in certain cases. However, the results require further scientific evidence to clarify the virulence issue. Various extracts of PG in organic solvents including kerosene ether, chloroform, acetone, ethanol, and methanol have been identified as potentially toxic to chicken embryos. A previous study warned about the choice of a conditioning solvent of suitable modulation for the application potential of PG [176]. PG-induced genomic damage was initially observed in a cell line that is normally extracted from monkey liver [177]. The combination of PG and Doxorubicin (a cancer chemotherapeutic drug) may slightly increase the cytotoxicity of Doxorubicin in tests of normal cells. However, determining the dose of PG/Doxorubicin that causes cytotoxic elongation in normal cells should be considered comprehensively [178]. Lazic et al. [120] proved that PG and its dibrominated derivatives (PG-Br and PG-Br2) did not affect Caenorhabditis elegans at a concentration of up to 50 µg/mL. Furthermore, the toxicity of the new bromination derivatives of PG on the zebrafish model also improved compared to parent PG [120]. Although the safety and toxicity of PG have been explored via experiments, further studies are still needed to evaluate comprehensively the toxicity of PG in each specific field.
4. Conclusions and Perspectives
This review summarized and updated eco-friendly substrates for fermentation to produce PG, and the benefit of PG production on a large scale (bioreactor system) was also clarified. Moreover, the potential uses of PG in agricultural applications were presented in detail. Based on the existing literature, PG has a promising use in large-scale production, even on an industrial scale. It is also considered as a potential candidate in management of multi-agents that are harmful to crops. This topic is interesting and important for the scientific community and must be further studied due to its applicability in practical agriculture in the future.
Although PG was produced using many types of byproducts/wastes, most of these materials are from fishery or agro-industrial sources. Nonetheless, there is still a need to find other byproducts in abundant amounts with nutrients, such as food processing or agricultural byproducts. To date, research related to PG production on a large scale is still very limited; hence, more studies are needed, especially on an industrial scale, to assess the exact PG productivity in industrial conditions in the future. Reports on the activities of PG in agricultural practices mostly focused on the inhibition of pathogenic fungi, whereas the tests on other harmful organisms are still very limited. Thus, subsequent studies should extend the scope of the bioactivity of this pigment to other objects, especially those that harm popular and economic crops. Furthermore, there is a need to increase greenhouse and field tests for potential bioactivities to obtain the optimum application of PG. In agricultural practices, potential bioactivities of PG in nanoscales are still not exploited, making it a promising research direction. Finally, the mechanisms of PG related to respective bioactivity in agriculture should be clarified and combined with other experimental findings to make a robust scientific basis for its application in the near future.
Author Contributions
Conceptualization, T.H.N., S.-L.W. and V.B.N.; writing—original T.H.N.; writing—review and editing, S.-L.W. and V.B.N.; supervision, S.-L.W. and V.B.N.; project administration, S.-L.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported in part by the Ministry of Science and Technology, Taiwan (MOST 111-2320-B-032-001; MOST 111-2923-B-032-001) and a grant from the Ministry of Education and Training, Vietnam (B2022-TTN-06).
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Darshan, N.; Manonmani, H.K. Prodigiosin and its potential applications. J. Food Sci. Technol. 2015, 52, 5393–5407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, V.B.; Nguyen, D.N.; Nguyen, A.D.; Ngo, V.A.; Ton, T.Q.; Doan, C.T.; Pham, T.P.; Tran, T.P.H.; Wang, S.L. Utilization of crab waste for cost-effective bioproduction of prodigiosin. Mar. Drugs 2020, 18, 523. [Google Scholar] [CrossRef]
- Rafael, G.A.; Natalia, R.Z.; Carlos, C.Z.; Mario, E.B.; Enrique, H.V.; Lizeth, P.A.; Jesús, A.R.H.; María, A.M.P.; Juan, E.S.H.; Manuel, M.R.; et al. Recent advances in prodigiosin as a bioactive compound in nanocomposite applications. Molecules 2022, 27, 4982. [Google Scholar]
- Wang, S.L.; Nguyen, V.B.; Doan, C.T.; Tran, T.N.; Nguyen, M.T.; Nguyen, A.D. Production and potential applications of bioconversion of chitin and protein-containing fishery byproducts into prodigiosin: A review. Molecules 2020, 25, 2744. [Google Scholar] [CrossRef] [PubMed]
- Pérez, T.R.; Montaner, B.; Llagostera, E.; Soto-Cerrato, V. The prodigiosins, proapoptotic drugs with anticancer properties. Biochem. Pharmacol. 2003, 66, 1447–1452. [Google Scholar] [CrossRef]
- Ravindran, A.; Anishetty, S.; Pennathur, G. Molecular dynamics of the membrane interaction and localisation of prodigiosin. J. Mol. Graph. Model. 2020, 98, 107614. [Google Scholar] [CrossRef]
- Lapenda, J.C.; Silva, P.A.; Vicalvi, M.C.; Sena, K.X.F.R.; Nascimento, S.C. Antimicrobial activity of prodigiosin isolated from Serratia marcescens UFPEDA 398. World J. Microbiol. Biotechnol. 2014, 31, 399–406. [Google Scholar] [CrossRef]
- Gulani, C.; Bhattacharya, S.; Das, A. Assessment of process parameters influencing the enhanced production of prodigiosin from Serratia marcescens and evaluation of its antimicrobial, antioxidant and dyeing potentials. Malays. J. Microbiol. 2012, 8, 116–122. [Google Scholar]
- Suryawanshi, R.K.; Patil, C.D.; Koli, S.H.; Hallsworth, J.E.; Patil, S.V. Antimicrobial activity of prodigiosin is attributable to plasma-membrane damage. Nat. Prod. Res. 2017, 31, 572–577. [Google Scholar] [CrossRef]
- Shaikh, Z. Biosynthesis of prodigiosin and its applications. IOSR J. Pharm. Biol. Sci. 2016, 11, 1–28. [Google Scholar]
- Han, S.B.; Kim, H.M.; Kim, Y.H.; Lee, C.W.; Jang, E.S.; Son, K.H.; Kim, S.U.; Kim, Y.K. T-cell specific immunosuppression by prodigiosin isolated from Serratia marcescens. Int. J. Immunopharmacol. 1998, 20, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Han, S.B.; Park, S.H.; Jeon, Y.J.; Kim, Y.K.; Kim, H.M.; Yang, K.H. Prodigiosin blocks T cell activation by inhibiting interleukin-2Ralpha expression and delays progression of autoimmune diabetes and collagen-induced arthritis. J. Pharmacol. Exp. Ther. 2001, 299, 415–425. [Google Scholar] [PubMed]
- Jung, E.H.; Hyun, J.K.; Kyung, H.K.; Joung, H.Y.; Hong, K.L.; Eun, W.S.; Suhkneung, P. Immunosuppressive effect of prodigiosin on murine splenocyte and macrophages. Biomol. Ther. 2008, 16, 351–355. [Google Scholar]
- Papireddy, K.; Smilkstein, M.; Kelly, J.X.; Shweta, S.S.M.; Alhamadsheh, M.; Haynes, S.W.; Challis, G.L.; Reynolds, K.A. Antimalarial activity of natural and synthetic prodiginines. J. Med. Chem. 2011, 54, 5296–5306. [Google Scholar] [CrossRef] [PubMed]
- Patil, C.D.; Patil, S.V.; Salunke, B.K.; Salunkhe, R.B. Prodigiosin produced by Serratia marcescens NMCC46 as a mosquito larvicidal agent against Aedes aegypti and Anopheles stephensi. Parasitol. Res. 2011, 109, 1179–1187. [Google Scholar] [CrossRef]
- Rocío, H.; Anna, M.; Alexandra, M.; Miguel, V.; Teresa, V. Using prodigiosin against some gram-positive and gram-negative bacteria and Trypanosoma cruzi. J. Venom. Anim. Toxins Incl. Trop. Dis. 2019, 25, e20190001. [Google Scholar]
- Genes, C.; Baquero, E.; Echeverri, F.; Maya, J.D.; Triana, O. Mitochondrial dysfunction in Trypanosoma cruzi: The role of Serratia marcescens prodigiosin in the alternative treatment of Chagas disease. Parasit Vectors 2011, 4, 66. [Google Scholar] [CrossRef] [Green Version]
- Yip, C.H.; Mahalingam, S.; Wan, K.L.; Nathan, S. Prodigiosin inhibits bacterial growth and virulence factors as a potential physiological response to interspecies competition. PLoS ONE 2021, 16, e0253445. [Google Scholar] [CrossRef]
- Taif, H.H.; Sawsan, H.A. Antimicrobial and antiadhesive properties of prodigiosin extracted from Serratia marcescens against bacteria isolated from patients with leukemia. World J. Pharm. Res. 2015, 4, 136–153. [Google Scholar]
- Arivizhivendhan, V.; Mahesh, M.; Boopathy, R.; Swarnalatha, S.; Regina, M.R.; Sekaran, G. Antioxidant and antimicrobial activity of bioactive prodigiosin produces from Serratia marcescens using agricultural waste as a substrate. J. Food Sci. Technol. 2018, 55, 2661–2670. [Google Scholar] [CrossRef]
- Muthukumar, A.; Pradeep, P.; Thigale, I.; Mohanasrinivasan, V.; Jemimah, N.S.; Devi, C.S. Exploring the bioactive potential of Serratia marcescens VITAPI (Acc: 1933637) isolated from soil. Front. Biol. 2016, 11, 476–480. [Google Scholar] [CrossRef]
- Borić, M.; Danevčič, T.; Stopar, D. Prodigiosin from Vibrio sp. DSM 14379; a new UV-protective pigment. Microb. Ecol. 2011, 62, 528. [Google Scholar] [CrossRef] [PubMed]
- Suryawanshi, R.K.; Patil, C.D.; Borase, H.P.; Salunke, B.K.; Patil, S.V. Studies on production and biological potential of prodigiosin by Serratia marcescens. Appl. Biochem. Biotechnol. 2014, 173, 1209–1221. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, V.B.; Chen, S.P.; Nguyen, T.H.; Nguyen, M.T.; Tran, T.T.T.; Doan, C.T.; Tran, T.N.; Nguyen, A.D.; Kuo, Y.H.; Wang, S.L. Novel efficient bioprocessing of marine chitins into active anticancer prodigiosin. Mar. Drugs 2020, 18, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomas, R.P.; Vinas, M. New insights on the antitumoral properties of prodiginines. Curr. Med. Chem. 2010, 17, 2222–2231. [Google Scholar] [CrossRef]
- Guryanova, I.D.; Karamovab, N.S.; Yusupovab, D.V.; Gnezdilovc, O.I.; Koshkarova, L.A. Bacterial pigment prodigiosin and its genotoxic effect. Russ. J. Bioorg. Chem. 2013, 39, 106–111. [Google Scholar] [CrossRef]
- Siew, W.S.; Sheila, N.; Kiew, L.W. Toxicity evaluation of prodigiosin from Serratia marcescens in a Caenorhabditis elegans model. AIP Conf. Proc. 2016, 1784, 020015. [Google Scholar]
- Li, X.; Tan, X.; Chen, Q.; Zhu, X.; Zhang, J.; Zhang, J.; Jia, B. Prodigiosin of Serratia marcescens ZPG19 alters the gut microbiota composition of Kunming mice. Molecules 2021, 26, 2156. [Google Scholar] [CrossRef]
- Nguyen, V.B.; Nguyen, D.N.; Wang, S.L. Microbial reclamation of chitin and protein-containing marine by-products for the production of prodigiosin and the evaluation of its bioactivities. Polymers 2020, 12, 1328. [Google Scholar] [CrossRef]
- Nguyen, T.H.; Wang, S.L.; Nguyen, D.N.; Nguyen, A.D.; Nguyen, T.H.; Doan, M.D.; Ngo, V.A.; Doan, C.T.; Kuo, Y.H.; Nguyen, V.B. Bioprocessing of marine chitinous wastes for the production of bioactive prodigiosin. Molecules 2021, 26, 3138. [Google Scholar] [CrossRef]
- Nguyen, V.B.; Wang, S.L.; Nguyen, A.D.; Phan, T.Q.; Techato, K.; Pradit, S. Bioproduction of prodigiosin from fishery processing waste shrimp heads and evaluation of its potential bioactivities. Fishes 2021, 6, 30. [Google Scholar] [CrossRef]
- Tran, L.T.; Techato, K.; Nguyen, V.B.; Wang, S.L.; Nguyen, A.D.; Phan, T.Q.; Doan, M.D.; Phoungthong, K. Utilization of cassava wastewater for low-cost production of prodigiosin via Serratia marcescens TNU01 fermentation and its novel potent α-glucosidase inhibitory effect. Molecules 2021, 26, 6270. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.H.; Wang, S.L.; Doan, M.D.; Nguyen, T.H.; Tran, T.H.T.; Tran, T.N.; Doan, C.T.; Ngo, V.A.; Ho, N.D.; Do, V.C.; et al. Utilization of by-product of groundnut oil processing for production of prodigiosin by microbial fermentation and its novel potent anti-nematodes effect. Agronomy 2022, 12, 41. [Google Scholar] [CrossRef]
- Atim, A.; Maurice, E.; Ubong, B.; Richard, A.; Nkpa, O.; Anitha, R.; Anthony, A.; Nsikak, B.; Joseph, E.; Sylvester, A. Stochastic modeling and meta-heuristic multivariate optimization of bioprocess conditions for co-valorization of feather and waste frying oil toward prodigiosin production. Prep. Biochem. Biotechnol. 2022, 1–14. [Google Scholar] [CrossRef]
- Islan, G.A.; Rodenak, K.B.; Noacco, N.; Duran, N.; Castro, G.R. Prodigiosin: A promising biomolecule with many potential biomedical applications. Bioengineered 2022, 13, 14227–14258. [Google Scholar] [CrossRef] [PubMed]
- Anita, K.; Mahnaz, M.A.; Fatemeh, A.F. Review of prodigiosin, pigmentation in Serratia marcescens. J. Biol. Sci. 2006, 6, 1–13. [Google Scholar]
- Han, R.; Xiang, R.; Li, J.; Wang, F.; Wang, C. High-level production of microbial prodigiosin: A review. J. Basic Microbiol. 2021, 61, 506–523. [Google Scholar] [CrossRef] [PubMed]
- Mnif, S.; Jardak, M.; Bouizgarne, B.; Aifa, S. Prodigiosin from Serratia: Synthesis and potential applications. Asian Pac. J. Trop. Biomed. 2022, 12, 233–242. [Google Scholar] [CrossRef]
- Venil, C.K.; Dufossé, L.; Renuka, D.P. Bacterial pigments: Sustainable compounds with market potential for pharma and food industry. Front. Sustain. Food Syst. 2020, 4, 100. [Google Scholar] [CrossRef]
- Choo, W.S.; Dufossé, L.; Morales, O.L. Editorial: Sustainable production of bioactive pigments. Front. Sustain. Food Syst. 2021, 5, 674311. [Google Scholar] [CrossRef]
- Yip, C.H.; Yarkoni, O.; Ajioka, J.; Wan, K.L.; Nathan, S. Recent advancements in high-level synthesis of the promising clinical drug, prodigiosin. Appl. Microbiol. Biotechnol. 2019, 103, 1667–1680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Numan, M.; Bashir, S.; Mumtaz, R.; Tayyab, S.; Rehman, N.U.; Khan, A.L.; Shinwari, Z.K.; Al-Harrasi, A. Therapeutic applications of bacterial pigments: A review of current status and future opportunities. 3 Biotech 2018, 8, 207. [Google Scholar] [CrossRef] [PubMed]
- Sen, T.; Barrow, C.J.; Deshmukh, S.K. Microbial pigments in the food industry-Challenges and the way forward. Front. Nutr. 2019, 6, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panesar, R.; Kaur, S.; Panesar, P.S. Production of microbial pigments utilizing agro-industrial waste: A review. Curr. Opin. Food Sci. 2015, 1, 70–76. [Google Scholar] [CrossRef]
- Lopes, F.C. Agro-industrial residues: Eco-friendly and inexpensive substrates for microbial pigments production. Front. Sustain. Food Syst. 2021, 5, 589414. [Google Scholar] [CrossRef]
- Nguyen, V.B.; Wang, S.L. New novel α-glucosidase inhibitors produced by microbial conversion. Process. Biochem. 2018, 65, 228–232. [Google Scholar] [CrossRef]
- Nguyen, T.H.; Wang, S.-L.; Nguyen, A.D.; Doan, M.D.; Tran, T.N.; Doan, C.T.; Nguyen, V.B. Novel α-amylase inhibitor hemi-pyocyanin produced by microbial conversion of chitinous discards. Mar. Drugs 2022, 20, 283. [Google Scholar] [CrossRef]
- Minaxi, S.; Zeba, U.; Vijai, K.G.; Rajeev, B. Valorization of fruits and vegetable wastes and by-products to produce natural pigments. Crit. Rev. Biotechnol. 2021, 41, 535–563. [Google Scholar]
- Faiqa, A.W.; Rukhsana, R.; Abida, J.; Bethania, B.; Shashikant, Y.; Thameed, A.; Makroo, H.A.; Dar, B.N. Valorisation of food wastes to produce natural pigments using non-thermal novel extraction methods: A review. Int. J. Food Sci. Technol. 2021, 56, 4823–4833. [Google Scholar]
- Usmani, Z.; Sharma, M.; Sudheer, S.; Gupta, V.K.; Bhat, R. Engineered microbes for pigment production using waste biomass. Curr. Genom. 2020, 21, 80–95. [Google Scholar] [CrossRef]
- Khalid, J.K.L.; Reem, W.Y.; Samer, T.M. An application of solid-state fermentation and elicitation with some microbial cells for the enhancement of prodigiosin production by Serratia marcescens. Al-Nahrain J. Sci. 2018, 21, 98–105. [Google Scholar]
- Filer, K. Production of enzymes for the feed industry using solid substrate fermentation. Engormix 2007. Available online: https://en.engormix.com/feed-machinery/articles/enzymes-feed-industry-t33575.htm (accessed on 25 November 2022).
- Wei, Y.H.; Chen, W.C. Enhanced production of prodigiosin-like pigment from Serratia marcescens SMdeltaR by medium improvement and oil-supplementation strategies. J. Biosci. Bioeng. 2005, 99, 616–622. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.H.; Yu, W.J.; Chen, W.C. Enhanced undecyl prodigiosin production from Serratia marcescens SS-1 by medium formulation and amino-acid supplementation. J. Biosci. Bioeng. 2005, 100, 466–471. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Tao, J.; Wei, D.; Shen, Y.; Tong, W. Development of an adsorption procedure for the direct separation and purification of prodigiosin from culture broth. Biotechnol. Appl. Biochem. 2005, 40, 277–280. [Google Scholar]
- Lin, C.; Jia, X.; Fang, Y.; Chen, L.; Zhang, H.; Lin, R.; Chen, J. Enhanced production of prodigiosin by Serratia marcescens FZSF02 in the form of pigment pellets. Electron. J. Biotechnol. 2019, 40, 58–64. [Google Scholar] [CrossRef]
- Paul, T.; Abhijit, M.; Tarun, K.B.; Biswanath, B. Prodigiosin production and recovery from Serratia marcescens: Process development and cost–benefit analysis. Biomass Convers. Biorefin. 2022. [Google Scholar] [CrossRef]
- Shahitha, S.; Poornima, K. Enhanced production of prodigiosin production in Serratia marcescens. J. Appl. Pharm. Sci. 2012, 2, 138–140. [Google Scholar] [CrossRef] [Green Version]
- Domröse, A.; Klein, A.S.; Hage, H.J.; Thies, S.; Svensson, V.; Classen, T.; Pietruszka, J.; Jaeger, K.E.; Drepper, T.; Loeschcke, A. Efficient recombinant production of prodigiosin in Pseudomonas putida. Front. Microbiol. 2015, 6, 972. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Wang, L.; Pan, X.; Osire, T.; Fang, H.; Zhang, H.; Yang, S.T.; Yang, T.; Rao, Z. Improved prodigiosin production by relieving CpxR temperature-sensitive inhibition. Front. Bioeng. Biotechnol. 2020, 8, 344. [Google Scholar] [CrossRef]
- Giri, A.V.; Anandkumar, N.; Muthukumaran, G.; Pennathur, G. A novel medium for the enhanced cell growth and production of prodigiosin from Serratia marcescens isolated from soil. BMC Microbiol. 2004, 4, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ulises, A.S.V.; Arlette, S.; Josefina, C.R.; Oscar, A. An in-situ approach based in mineral oil to decrease end-product inhibition in prodigiosin production by Serratia marcescens. Food Bioprod. Process. 2022, 135, 217–226. [Google Scholar]
- Liu, W.; Yang, J.; Tian, Y.; Zhou, X.; Wang, S.; Zhu, J.; Sun, D.; Liu, C. An in situ extractive fermentation strategy for enhancing prodigiosin production from Serratia marcescens BWL1001 and its application to inhibiting the growth of Microcystis aeruginosa. Biochem. Eng. J. 2021, 166, 107836. [Google Scholar] [CrossRef]
- Ahmed, M.A.E.; Mervat, M.A.E.G.; Rasha, H.B. Over production and biological activity of prodigiosin-like pigments from recombinant fusant of endophytic marine Streptomyces species. Antonie Van Leeuwenhoek 2012, 102, 719–734. [Google Scholar]
- Wang, S.L.; Wang, C.Y.; Yen, Y.H.; Liang, T.W.; Chen, S.Y.; Chen, C.H. Enhanced production of insecticidal prodigiosin from Serratia marcescens TKU011 in media containing squid pen. Process Biochem. 2012, 47, 1684–1690. [Google Scholar] [CrossRef]
- de Araújo, H.W.; Fukushima, K.; Takaki, G.M. Prodigiosin production by Serratia marcescens UCP 1549 using renewable resources as a low cost substrate. Molecules 2010, 15, 6931–6940. [Google Scholar] [CrossRef]
- Vijayalakshmi, K.; Jagathy, K. Production of prodigiosin from Serratia marcescens and its antioxidant and anticancer potential. Int. J. Adv. Res. Biol. Sci. 2016, 3, 75–88. [Google Scholar]
- Naik, C.; Srisevita, J.M.; Shushma, K.N.; Farah, N.; Shilpa, A.C.; Muttanna, C.D.; Darshan, N.; Sannadurgappa, D. Peanut oil cake: A novel substrate for enhanced cell growth and prodigiosin production from Serratia marcescens CF-53. J. Res. Biol. 2012, 2, 549–557. [Google Scholar]
- Nisarg, G.; Gargi, B.; Ravi, K.; Vikrant, P.; Khushal, K.; Jigresh, G.; Khalid, J.A.; Show, P.L.; Rupesh, M.; Vijai, S. Biovalorization of agro-industrial waste soybean meal for the production of prodigiosin by Serratia marcescens. Biomass Convers. Biorefin. 2021. [Google Scholar] [CrossRef]
- Silva, L.A.R.; Santos, R.A.; Mendonça, R.S.; Andrade, R.F.S.; Rodríguez, D.M.; Campos-Takaki, G.M. Cost-effective production of stable prodigiosin by Serratia marcescens UCP 1549 and application in soap coloring. Res. Soc. Dev. 2022, 11, e9711427078. [Google Scholar] [CrossRef]
- Ozdal, M. Enhanced production of prodigiosin by Serratia marcescens MO-1 using ram horn peptone. Braz. J. Microbiol. 2015, 46, 631–637. [Google Scholar]
- Aruldass, C.A.; Venil, C.K.; Zakaria, Z.A.; Ahmad, W.A. Brown sugar as a low-cost medium for the production of prodigiosin by locally isolated Serratia marcescens UTM1. Int. Biodeterior. Biodegrad. 2014, 95, 19–24. [Google Scholar] [CrossRef]
- Nittu, B.A.; Hitha, K.; Shirly, K.T. Solid state fermentation for prodigiosin production using Serratia marcescens. Int. Res. J. Eng. Technol. 2019, 6, 330–335. [Google Scholar]
- Sumathi, C.; Mohana, P.D.; Swarnalatha, S.; Dinesh, M.G.; Sekaran, G. Production of prodigiosin using tannery fleshing and evaluating its pharmacological effects. Sci. World J. 2014, 2014, 290327. [Google Scholar] [CrossRef]
- Xia, S.; Veony, E.; Yang, Q. Kitchen waste as a novel available substrate for prodigiosin production by Serratia marcescense. IOP Conf. Ser. Earth Environ. Sci. 2018, 171, 01203749. [Google Scholar] [CrossRef]
- FAO. The State of World Fisheries and Aquaculture-Sustainability in Action; FAO: Rome, Italy, 2020; 224p. [Google Scholar]
- Grewal, J.; Pyter, W.; Joshi, N.; Drewniak, L.; Pranaw, K. Colorful treasure from agro-industrial wastes: A sustainable chassis for microbial pigment production. Front. Microbiol. 2022, 13, 832918. [Google Scholar] [CrossRef]
- Madeira, J.; Contesini, F.J.; Calzado, F.; Rubio, M.V.; Zubieta, M.P.; Lopes, D.B.; de Melo, R.R. Agro-Industrial Residues and Microbial Enzymes: An Overview on the Eco-Friendly Bioconversion into High Value-Added Products. In Biotechnology of Microbial Enzymes; Brahmachari, G., Ed.; Elsevier: London, UK, 2017; pp. 475–511. [Google Scholar]
- Sodhi, A.S.; Sharma, N.; Bhatia, S.; Verma, A.; Soni, S.; Batra, N. Insights on sustainable approaches for production and applications of value added products. Chemosphere 2021, 286, 131623. [Google Scholar] [CrossRef]
- Siddhartha, G.V.A.O.C.; François, L.; Sylvain, M.; Eric, D.; Marcia, N.; Jonas, C. Cassava wastewater as a substrate for the simultaneous production of rhamnolipids and polyhydroxy alkanoates by Pseudomonas aeruginosa. J. Ind. Microbiol. Biotechnol. 2009, 36, 1063–1072. [Google Scholar]
- Sánchez, A.S.; Silva, Y.L.; Kalid, R.A.; Cohim, E.; Torres, E.A. Waste bio-refineries for the cassava starch industry: New trends and review of alternatives. Renew. Sustain. Energy Rev. 2017, 73, 1265–1275. [Google Scholar] [CrossRef]
- Dos Santos, R.A.; Rodríguez, D.M.; da Silva, L.A.R.; de Almeida, S.M.; de Campos, T.G.M.; de Lima, M.A.B. Enhanced production of prodigiosin by Serratia marcescens UCP 1549 using agrosubstrates in solid-state fermentation. Arch. Microbiol. 2021, 203, 4091–4100. [Google Scholar] [CrossRef]
- Xia, Y.; Wang, G.; Lin, X.; Xin, S.; Lianzhong, A. Solid-state fermentation with Serratia marcescens Xd-1 enhanced production of prodigiosin by using bagasse as an inertia matrix. Ann. Microbiol. 2016, 66, 1239–1247. [Google Scholar] [CrossRef]
- Arivizhivendhan, K.V.; Mahesh, M.; Regina, M.R.; Sekaran, G. Bioactive prodigiosin isolated from Serratia marcescens using solid state fermenter and its bactericidal activity compared with conventional antibiotics. J. Microb. Biochem. Technol. 2015, 7, 305–312. [Google Scholar]
- Lapenda, L.J.C.; Maciel, C.C.S.; Xavier, H.S.; Alves, S.C.A.; Campos, T.G. Production and toxicological evaluation of prodigiosin from Serratia marcescens UCP/WFCC1549 on mannitol solid medium. Int. J. Appl. Res. Nat. Prod. 2014, 7, 32–38. [Google Scholar]
- Kamlesh, K.M.; Abhishek, D.T.; Deepak, K.; Suresh, K.S. Production, purification and characterization of prodigiosin by Serratia nematodiphilia (NCIM 5606) using solid-state fermentation with various substrate. Ann. Phytomed. 2020, 9, 302–306. [Google Scholar]
- Robinson, T.; Singh, D.; Nigam, P. Solid-state fermentation: A promising microbial technology for secondary metabolite production. Appl. Microbiol. Biotechnol. 2001, 55, 284–289. [Google Scholar] [CrossRef]
- Mudgett, R.E.; Nash, J.; Ruther, R. Controlled gas environments in solid state fermentations. Dev. Ind. Microbiol. 1992, 34, 1217–1233. [Google Scholar]
- Yang, S.S.; Yuan, S.S. Oxytetracycline production by Streptomyces rimosus in solid state fermentation of sweet potato residue. World J. Microbiol. Biotechnol. 1990, 6, 236–244. [Google Scholar] [CrossRef]
- Nguyen, S.L.T.; Nguyen, T.C.; Do, T.T.; Vu, T.L.; Nguyen, T.T.; Do, T.T.; Nguyen, T.H.T.; Le, T.H.; Trinh, D.K.; Nguyen, T.A.T. Study on the anticancer activity of prodigiosin from variants of Serratia marcescens QBN VTCC 910026. Biomed Res. Int. 2022, 2022, 11. [Google Scholar] [CrossRef]
- Pardeep, K.S.; Suresh, K.; Prince, C.; Joginder, S.D. Fermentation: A boon for production of bioactive compounds by processing of food industries wastes (by-products). Molecules 2018, 23, 2560. [Google Scholar]
- Lizardi, J.M.A.; Hernández, M.R. Solid state fermentation (SSF): Diversity of applications to valorize waste and biomass. 3 Biotech 2017, 7, 44. [Google Scholar] [CrossRef]
- Harris, A.K.P.; Williamson, N.R.; Slater, H.; Cox, A.; Abbasi, S.; Foulds, I. The Serratia gene cluster encoding biosynthesis of the red antibiotic, prodigiosin, shows species- and strain-dependent genome context variation. Microbiol. 2004, 150, 3547–3560. [Google Scholar] [CrossRef] [PubMed]
- Williamson, N.R.; Simonsen, H.T.; Ahmed, R.A.; Goldet, G.; Slater, H.; Woodley, L. Biosynthesis of the red antibiotic, prodigiosin, in Serratia: Identification of a novel 2-methyl-3- n-amyl-pyrrole (MAP) assembly pathway, definition of the terminal condensing enzyme, and implications for undecylprodigiosin biosynthesis in Streptomyces. Mol. Microbiol. 2005, 56, 971–989. [Google Scholar] [CrossRef]
- Qadri, S.M.H.; Williams, R.P. Role of methionine in biosynthesis of prodigiosin by Serratia marcescens. J. Bacteriol. 1973, 116, 1191–1198. [Google Scholar] [CrossRef] [Green Version]
- Williams, R.P.; Gott, C.L.; Qadri, S.M. Induction of pigmentation in nonproliferating cells of Serratia marcescens by addition of single amino acids. J. Bacteriol. 1971, 106, 444–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faraag, A.; El-batal, A.; El-Hendawy, H. Characterization of prodigiosin produced by Serratia marcescens strain isolated from irrigation water in Egypt. Nat. Sci. 2017, 15, 55–68. [Google Scholar]
- Siva, R.; Subha, K.; Bhakta, D.; Ghosh, A.R.; Babu, S. Characterization and enhanced production of prodigiosin from the spoiled coconut. Appl. Biochem. Biotechnol. 2012, 166, 187–196. [Google Scholar] [CrossRef]
- Picha, P.; Kale, D.; Dave, I.; Pardeshi, S. Comparative studies on prodigiosin production by Serratia marcescens using various crude fatty acid sources—Its characterization and applications. J. Int. J. Curr. Microbiol. Appl. Sci. 2015, 2, 254–267. [Google Scholar]
- Parani, K.; Saha, B.K. Optimization of prodigiosin production from a strain of Serratia marcescens SRI and screening for antifungal activity. J. Biol. Control 2008, 22, 73–79. [Google Scholar]
- Oh, D.C.; Jensen, P.R.; Kauffman, C.A.; Fenical, W. Libertellenones A–D: Induction of cytotoxic diterpenoid biosynthesis by marine microbial competition. Bioorg. Med. Chem. 2005, 13, 5267–5273. [Google Scholar] [CrossRef]
- Abdel, G.T.M. Eco-friendly and safe role of juniperus procera in controlling of fungal growth and secondary metabolites. J. Plant Pathol. Microbiol. 2014, 5, 231. [Google Scholar]
- Abdelghany, T.M.; El-Naggar, M.A.; Ganash, M.A.; Mohamed, A.A.A. PCR Identification of Aspergillus niger with using natural additives for controlling and detection of malformins and maltoryzine production by HPLC. BioNanoScience 2017, 7, 588–596. [Google Scholar] [CrossRef]
- Mahmoud, S.T.; Luti, K.J.K.; Yonis, R.W. Enhancement of prodigiosin production by Serratia marcescens S23 via introducing microbial elicitor cells into culture medium. Iraqi J. Sci. 2015, 56, 1938–1951. [Google Scholar]
- Huy, N.A.D.; Nguyen, T.H.K. Studies on the prodigiosin production from Streptomyces coelicolor in liquid media by using heated Lactobacillus rhamnosus. J. Appl. Pharm. Sci. 2014, 4, 21–24. [Google Scholar]
- Wang, S.L.; Chen, S.Y.; Yen, Y.H.; Liang, T.W. Utilization of chitinous materials in pigment adsorption. Food Chem. 2012, 135, 1134–1140. [Google Scholar] [CrossRef]
- Chavez, C.L.; Aguilar, O. Effect of mineral supplementation and type of starch on the production of prodigiosin from a culture of Serratia marcescens BS303. Rev. Mex. Ing. Quim. 2015, 14, 641–652. [Google Scholar]
- Silverman, M.; Munoz, E. Effect of iron and salt on prodigiosin synthesis in Serratia marcescens. J. Bacteriol. 1973, 114, 999–1006. [Google Scholar] [CrossRef] [Green Version]
- Allen, G.; Reichelt, J.; Grayv, P. Influence of environmental factors and medium composition on Vibrio gazogenes growth and prodigiosin production. Appl. Environ. Microbiol. 1983, 45, 1727–1732. [Google Scholar] [CrossRef] [Green Version]
- Witney, R.F.; Failla, M.L.; Weinberg, E.D. Phosphate inhibition of secondary metabolism in Serratia marcescens. Appl. Environ. Microbiol. 1977, 33, 1042–1046. [Google Scholar] [CrossRef] [Green Version]
- Stella, N.A.; Lahr, R.M.; Kalivoda, E.J.; Shanks, R.M.Q. Serratia marcescens quinoprotein glucose dehydrogenase activity mediates acidification and inhibition of prodigiosin production by glucose. Appl. Environ. Microbiol. 2012, 78, 6225–6235. [Google Scholar]
- Solé, M.; Rius, N.; Francia, A.; Lorén, J.G. The effect of pH on prodigiosin production by non-proliferating cells of Serratia marcescens. Lett. Appl. Microbiol. 1994, 19, 341–344. [Google Scholar] [CrossRef]
- Clements, J.S. The reversal of glucose repressed prodigiosin production in Serratia marcescens by the cyclic 3′5′- adenosine monophosphate inhibitor theophylline. Experientia 1976, 32, 421–422. [Google Scholar] [CrossRef] [PubMed]
- Basit, G.; Mujib, A.; Moien, Q.M.; Jyoti, M.; Rukaya, S.; Nadia, Z. Plant tissue culture: Agriculture and industrial applications. In Transgenic Technology Based Value Addition in Plant Biotechnology, 1st ed.; Usha, K., Malik, A., Kamaluddin, Eds.; Academic Press: New York, NY, USA, 2020; pp. 25–49. [Google Scholar]
- Tao, J.L.; Wang, X.D.; Shen, Y.L.; Wei, D.Z. Strategy for the improvement of prodigiosin production by a Serratia marcescens mutant through fed-batch fermentation. World J. Microbiol. Biotechnol. 2005, 21, 969–972. [Google Scholar] [CrossRef]
- Jeong, Y.; Kim, H.J.; Kim, S.; Park, S.Y.; Kim, H.R.; Jeong, S.; Lee, S.J.; Lee, M.S. Enhanced large-scale production of Hahella chejuensis -derived prodigiosin and evaluation of its bioactivity. J. Microbiol. Biotechnol. 2021, 31, 1624–1631. [Google Scholar] [CrossRef] [PubMed]
- Qi, F.; Xiao, Y.; Duan, X.; Huang, H.; Zhuang, Z.; Shen, J.; Pei, Y.; Xu, H.; Gan, M. Continuous fermentation of a prodigiosin-producing Serratia marcescens strain isolated from soil. Adv. Biosci. Biotechnol. 2019, 10, 98–108. [Google Scholar]
- Bae, J.; Moon, H.; Oh, K.K.; Kim, C.H.; Lee, D.S.; Kim, S.W.; Hong, S.I. A novel bioreactor with an internal adsorbent for integrated fermentation and recovery of prodigiosin-like pigment produced from Serratia sp. KH-95. Biotechnol. Lett. 2001, 23, 1315–1319. [Google Scholar] [CrossRef]
- Sura, J.M.; Khalid, J.K.L. A kinetic model for prodigiosin production by Serratia marcescens as a bio-colorant in bioreactor. AIP Conf. Proc. 2020, 2213, 020027. [Google Scholar]
- Lazic, J.; Skaro, B.S.; Vojnovic, S.; Aleksic, I.; Milivojevic, D.; Kretzschmar, M.; Gulder, T.; Petkovic, M.; Nikodinovic, R.J. Synthesis, anticancer potential and comprehensive toxicity studies of novel brominated derivatives of bacterial biopigment prodigiosin from Serratia marcescens ATCC 27117. Molecules 2022, 27, 3729. [Google Scholar] [CrossRef] [PubMed]
- Michele, R.S.; Luciana, V.; Adriane, B.P.M.; Carlos, S. Application of different types of bioreactors in bioprocesses. In Bioreactors: Design, Properties and Applications, UK ed.; Paolo, G.A., Liu, Z., Eds.; Nova Science Publishers Inc.: New York, NY, USA, 2011; pp. 55–90. [Google Scholar]
- Luis, C.C.R.; Aguilar, O. An integrated process for the in situ recovery of prodigiosin using micellar ATPS from a culture of Serratia marcescens. J. Chem. Technol. Biotechnol. 2016, 91, 2896–2903. [Google Scholar]
- Amera, E.; Ahmed, A.E.; Amr, M.M.; Yehia, O. Optimization of bioreactor cultivation parameters by Taguchi Orthogonal array design for enhanced prodigiosin production. Iran J. Chem. Chem. Eng. 2020, 39, 319–330. [Google Scholar]
- Sagar, B.S.V.; Deepak, B.S.; Tejaswini, G.S.; Aparna, Y.; Sarada, J. Evaluation of prodigiosin pigment for antimicrobial and insecticidal activities on selected bacterial pathogens & household pests. Int. J. Res. Biol. Sci. 2019, 6, 96–102. [Google Scholar]
- Jain, A.; Sarsaiya, S.; Wu, Q.; Lu, Y.; Shi, J. A review of plant leaf fungal diseases and its environment speciation. Bioengineered 2019, 10, 409–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iqbal, Z.; Khan, M.A.; Sharif, M.; Jamal, H.S.; Muhammad, H.R.; Kashif, J. An automated detection and classification of citrus plant diseases using image processing techniques: A review. Comput. Electron. Agric. 2018, 153, 12–32. [Google Scholar] [CrossRef]
- Duzhak, A.B.; Panfilova, Z.I.; Duzhak, T.G.; Vasyunina, E.A.; Shternshis, M.V. Role of prodigiosin and chitinases in antagonistic activity of the bacterium Serratia marcescens against the fungus Didymella applanata. Biochemistry 2012, 77, 910–916. [Google Scholar] [CrossRef]
- Samer, S.H.; Hannah, U.C.B.; Andreas, S.K.; David, P.K.; Tim, M.W.; Thomas, C.; Jörg, P.; Florian, M.W.G.; Sylvia, A.S.S. Novel prodiginine derivatives demonstrate bioactivities on plants, nematodes, and fungi. Front. Plant Sci. 2020, 11, 579807. [Google Scholar]
- Nobutaka, S.; Masami, N.; Kazuyuki, H.; Tadaaki, H.; Katsumi, A. Synergistic antifungal activity of chitinolytic enzymes and prodigiosin produced by biocontrol bacterium, Serratia marcescens strain B2 against gray mold pathogen, Botrytis cinerea. J. Gen. Plant Pathol. 2001, 67, 312–317. [Google Scholar]
- Ingrid, G.R.M.; Francisco, H.M.; Dunn, M.F.; Karina, G.N.; Graciela, H.P. Antifungal activity of Serratia marcescens CFFSUR-B2 purified chitinolytic enzymes and prodigiosin against Mycosphaerella fijiensis, causal agent of black Sigatoka in banana (Musa spp.). BioControl 2015, 60, 565–572. [Google Scholar]
- Hiroshi, O.; Sato, Z.; Sato, M.; Koiso, Y.; Iwasaki, S.; Isaka, M. Identification of antibiotic red pigments of Serratia marcescens F-1-1, a biocontrol agent of damping-off of cucumber, and antimicrobial activity against other plant pathogens. Jpn. J. Phytopathol. 1998, 64, 294–298. [Google Scholar]
- Jimtha, J.C.; Jishma, P.; Sreelekha, S.; Chithra, S.; Radhakrishnan, E.K. Antifungal properties of prodigiosin producing rhizospheric Serratia sp. Rhizosphere 2017, 3, 105–108. [Google Scholar] [CrossRef]
- Alijani, Z.; Amini, J.; Ashengroph, M.; Bahman, B. Antifungal activity of Serratia rubidaea Mar61-01 purifed prodigiosin against Colletotrichum nymphaeae, the causal agent of strawberry anthracnose. J. Plant Growth Regul. 2022, 41, 585–595. [Google Scholar] [CrossRef]
- Rahul, S.; Patil, C.; Hemant, B.; Chandrakant, N.; Laxmikant, S.; Satish, P. Nematicidal activity of microbial pigment from Serratia marcescens. Nat. Prod. Res. 2014, 28, 1399–1404. [Google Scholar] [CrossRef]
- Omnia, M.M.; Rania, A.A.H.; Dina, S.S.I.; Mona, H.B.; Hussien, E.M. Effects of Serratia marcescens and prodigiosin pigment on the root-knot nematode Meloidogyne incognita. Middle East J. Agric. Res. 2020, 9, 243–252. [Google Scholar]
- Wei, H.; Zheng, R.; Liao, Y.; Fan, K.; Yang, Z.; Chen, T.; Zhang, N. Evaluating the biological potential of prodigiosin from Serratia Marcescens KH-001 against Asian citrus psyllid. J. Econ. Entomol. 2021, 114, 1219–1225. [Google Scholar]
- Patil, N.G.; Kadam, M.S.; Patil, V.R.; Chincholkar, S.B. Insecticidal properties of water diffusible prodigiosin produced by Serratia nematodiphila 213C. Curr. Trends Biotechnol. Pharm. 2013, 7, 773–781. [Google Scholar]
- Liang, T.W.; Chen, S.Y.; Chen, Y.C.; Chen, C.H.; Yen, Y.H.; Wang, S.L. Enhancement of prodigiosin production by Serratia marcescens TKU011 and its insecticidal activity relative to food colorants. J. Food Sci. 2013, 78, M1743-51. [Google Scholar] [CrossRef] [PubMed]
- Asano, S.; Ogiwara, K.; Nakagawa, Y.; Suzuki, K.; Hori, H.; Watanabe, T. Prodigiosin produced by Serratia marcescens enhances the insecticidal activity of Bacillus thuringiensis delta-endotoxin (Cry 1C) against common cutworm, Spodoptera litura. J. Pestic. Sci. 1999, 24, 381–385. [Google Scholar] [CrossRef]
- Bar-On, Y.M.; Philips, R.; Milo, R. The biomass distribution on earth. Proc. Natl. Acad. Sci. USA 2018, 115, 6506–6511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajest, K.V.; Kutulus, B.K.; Antony, R. Plant pathogenic bacteria: An overview. In Sustanable Approaches to Controlling Plant Pathogenic Bacteria, 1st ed.; Kannan, V.R., Bastas, K.K., Eds.; CRC Press: Boca Raton, FL, USA, 2016; pp. 1–15. [Google Scholar]
- Jorge, P.; Patricia, A.U.; Carolina, E. Biological control of plant-parasitic nematodes by Filamentous fungi inducers of resistance: Trichoderma, Mycorrhizal and Endophytic fungi. Front. Microbiol. 2020, 11, 992. [Google Scholar]
- Mitiku, M. Plant-parasitic nematodes and their management: A review. J. Biol. Agric. Healthc. 2018, 16, 555980. [Google Scholar] [CrossRef]
- Palomares, R.J.E.; Escobar, C.; Cabrera, J.; Vovlas, A.; Castillo, P. Anatomical alterations in plant tissues induced by plant-parasitic nematodes. Front. Plant Sci. 2017, 8, 1987. [Google Scholar] [CrossRef] [Green Version]
- Vina, M.; Loekas, S.S.; Khairan, K.; Takahiro, H.; Koichi, H.; Rina, S. Secondary metabolites produced by endophytic bacteria against the root-knot nematode (Meloidogyne sp.). Biodiversitas 2020, 21, 5270–5275. [Google Scholar]
- Roberts, D.P.; Selmer, K.; Lupitskyy, R.; Rice, C.; Buyer, J.S.; Maul, J.E.; Lakshman, D.K.; DeSouza, J. Seed treatment with prodigiosin controls damping-off of cucumber caused by Pythium ultimum. AMB Express 2021, 11, 10. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.N.; Do, V.C.; Nguyen, V.B. Selection of baterial strain processing ability to ferment shrimp shell powder for the production of anti-nematode Meloidogyne incognita affecting black pepper plants. Tay Nguyen J. Sci. 2020, 42, 18–25. [Google Scholar]
- Ahmed, E.; Hoda, E.; Ahmed, I.F. In silico and in vitro cytotoxic effect of prodigiosin-conjugated silver nanoparticles on liver cancer cells (HepG2). Biotechnologia 2017, 98, 225–243. [Google Scholar]
- Karthika, D.; Vadakkan, K.; Ashwini, R.; Shyamala, A.; Hemapriya, J.; Vijayanand, S. Prodigiosin mediated biosynthesis of silver nanoparticles and evaluation of its antibacterial activity. Int. J. Curr. Microbiol. Appl. Sci. 2015, 3, 868–874. [Google Scholar]
- Dozie, S.O.N.; Obayemi, J.D.; Danyuo, Y.T.; Etuk, U.G.; Chi, Y.; Hu, J.; Anuku, N.; Odusanya, O.S.; Malatesta, K.; Soboyejo, W.O. Biosynthesis of gold nanoparticles and gold/prodigiosin nanoparticles with Serratia marcescens bacteria. Waste Biomass Valor. 2017, 8, 2045–2059. [Google Scholar] [CrossRef]
- Suryawanshi, R.; Patil, C.; Borase, H.; Salunke, I.; Elisabeth, M.; Philippe, G.; Patil, S. In vitro antiparasitic activity of microbial pigments and their combination with phytosynthesized metal nanoparticles. Parasitol. Int. 2015, 64, 353–356. [Google Scholar]
- John, D.O.; Karen, A.M.; Olushola, S.O.; Danyuo, Y.; Wei, Y.; Kathryn, E.U.; Winston, O.S. Injectable, biodegradable micro- and nano-particles loaded with prodigiosin-based drug for localized anticancer drug delivery. Cancer Epidemiol. Biomark. Prev. 2016, 25, C60. [Google Scholar]
- Ren, Y.; Gong, J.; Fu, R.; Li, Z.; Yu, Z.; Lou, J.; Wang, F.; Zhang, J. Dyeing and functional properties of polyester fabric dyed with prodigiosins nanomicelles produced by microbial fermentation. J. Clean. Prod. 2017, 148, 375–385. [Google Scholar] [CrossRef]
- Gong, J.; Liu, J.; Tan, X.; Li, Z.; Li, Q.; Zhang, J. Bio-preparation and regulation of pyrrole structure nano-pigment based on biomimetic membrane. Nanomaterial 2019, 9, 114. [Google Scholar] [CrossRef] [Green Version]
- Shahla, N. Spray-dried prodigiosin from Serratia marcescens as a colorant. Biosci. Biotechnol. Res. Asia 2013, 10, 69–76. [Google Scholar]
- Abd, E.; Kamel, A.; Prasad, R. Role of nanotechnology applications in plant-parasitic nematode control. In Nanobiotechnology Applications in Plant Protection; Abd-Elsalam, K.A., Prasad, R., Eds.; Springer: New York, NY, USA, 2019; pp. 223–240. [Google Scholar]
- Abdelghany, T.M.; Al-Rajhi, A.M.H.; Al Abboud, M.A.; Alawlaqi, M.M.; Magdah, A.G.; Eman, A.M.H.; Ahmed, S.M. Recent advances in green synthesis of silver nanoparticles and their applications: About future directions. A review. BioNanoScience 2018, 8, 5–16. [Google Scholar] [CrossRef]
- Hasanin, M.; Al Abboud, M.A.; Alawlaqi, M.M.; Tarek, M.A.; Amr, H.H. Ecofriendly synthesis of biosynthesized copper nanoparticles with starch-based nanocomposite: Antimicrobial, antioxidant, and anticancer activities. Biol. Trace Elem. Res. 2022, 200, 2099–2112. [Google Scholar] [CrossRef]
- Amr, H.H.; Mohamed, A.A.A.; Mohamed, M.A.; Tarek, M.A.; Mohamed, H. Synthesis of nanocapsules based on biosynthesized nickel nanoparticles and potato starch: Antimicrobial, antioxidant, and anticancer activity. Starch 2021, 74, 2100165. [Google Scholar]
- Abdelghany, T.M.; Al-Rajhi, A.M.H.; Almuhayawi, M.S.; Emad, A.; Mohamed, A.A.A.; Hanan, M.; Reham, Y.; Samy, S. Green fabrication of nanocomposite doped with selenium nanoparticle–based starch and glycogen with its therapeutic activity: Antimicrobial, antioxidant, and anti-inflammatory in vitro. Biomass Conv. Biore. 2022. [Google Scholar] [CrossRef]
- Darshan, N.; Manonmani, H.K. Prodigiosin inhibits motility and activates bacterial cell death revealing molecular biomarkers of programmed cell death. AMB Express 2016, 6, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danevčič, T.; Borić, V.M.; Tabor, M.; Zorec, M.; Stopar, D. Prodigiosin induces autolysins in actively grown Bacillus subtilis cells. Front. Microbiol. 2016, 7, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danevčič, T.; Borić, V.M.; Zorec, M.; Stopar, D. Prodigiosin—A multifaceted Escherichia coli antimicrobial agent. PLoS ONE 2016, 11, e0162412. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Liu, J.; Wang, X.; Kong, D.; Du, W.; Li, H.; Hse, C.Y.; Shupe, T.; Zhou, D.; Zhao, K. Biological potential and mechanism of prodigiosin from Serratia marcescens Subsp. lawsoniana in human choriocarcinoma and prostate cancer cell lines. Int. J. Mol. Sci. 2018, 19, 3465. [Google Scholar]
- Francisco, R.; Pérez, T.R.; Gimènez, B.P.; Soto, C.V.; Giménez, X.P.; Ambrosio, S. Mechanisms of prodigiosin cytotoxicity in human neuroblastoma cell lines. Eur. J. Pharmacol. 2007, 572, 111–119. [Google Scholar] [CrossRef]
- Llagostera, E.; Soto, C.V.; Montaner, B.; Pérez, T.R. Prodigiosin induces apoptosis by acting on mitochondria in human lung cancer cells. Ann. N. Y. Acad. Sci. 2003, 1010, 178–181. [Google Scholar] [CrossRef]
- Baldino, C.M.; Parr, J.; Wilson, C.J.; Ng, S.C.; Yohannes, D.; Wasserman, H.H. Indoloprodigiosins from the C-10 bipyrrolic precursor: New antiproliferative prodigiosin analogs. Bioorg. Med. Chem. Lett. 2006, 16, 701–704. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Li, B.; Zhou, L.; Yu, S.; Su, Z.; Song, J.; Sun, Q.; Sha, O.; Wang, X.; Jiang, W.; et al. Prodigiosin inhibits Wnt/β-catenin signaling and exerts anticancer activity in breast cancer cells. Proc. Natl. Acad. Sci. USA 2016, 113, 13150–13155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Zhou, H.; Ma, X.; Lin, C.; Lu, L.; Liu, D.; Ma, D.; Gao, X.; Qian, X.Y. Prodigiosin inhibits proliferation, migration, and invasion of nasopharyngeal cancer cells. Cell Physiol. Biochem. 2018, 48, 1556–1562. [Google Scholar] [CrossRef] [PubMed]
- Hazarika, D.J.; Gautom, T.; Parveen, A.; Goswami, G.; Barooah, M.; Modi, M.K.; Robin, C.B. Mechanism of interaction of an endofungal bacterium Serratia marcescens D1 with its host and non-host fungi. PLoS ONE 2020, 15, e0224051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, Z.; Shen, Z.H.; Xing, J.H.; Guo, Y.; Xu, T.M.; Peng, W.L.; Liu, X.H. Synthesis and nematocidal activity of novel 1-(3-chloropyridin-2-yl)-3-(trifluoromethyl)-1H-pyrazole-4-carboxamide derivatives. Chem. Pap. 2017, 71, 921–928. [Google Scholar]
- Keerthiraj, M.; Mandal, A.; Dutta, T.K.; Saha, S.; Dutta, A.; Singh, A.; Kundu, A. Nematicidal and molecular docking investigation of essential oils from Pogostemon cablin ecotypes against Meloidogyne incognita. Chem. Biodivers. 2021, 18, e2100320. [Google Scholar] [CrossRef]
- Babu, R.O.; Moorkoth, D.; Azeez, S.; Eapen, S.J. Virtual screening and in vitro assay of potential drug like inhibitors from spices against Glutathione-S-Transferase of Meloidogyne incognita. Bioinformation 2012, 8, 319–325. [Google Scholar] [CrossRef] [Green Version]
- Kundu, A.; Dutta, A.; Mandal, A.; Negi, L.; Malik, M.; Puramchatwad, R.; Antil, J.; Singh, A.; Rao, U.; Saha, S.; et al. A comprehensive in vitro and in silico analysis of nematicidal action of essential oils. Front. Plant Sci. 2021, 11, 614143. [Google Scholar] [CrossRef]
- Emmanuel, B.; Diana, F.; Deisy, X.A.; Philippe, N.; Petitot, A.S.; Lisei, S.M.E.; Erika, V.S.A.; Etienn, G.J.D.; Bernard, M.; Natália, F.M. A chemosensory GPCR as a potential target to control the root-knot nematode Meloidogyne incognita parasitism in plants. Molecules 2019, 24, 3798. [Google Scholar]
- Kalesperis, G.S.; Prahlad, K.V.; Lynch, D.L. Toxigenic studies with the antibiotic pigments from Serratia marcescens. Can. J. Microbiol. 1975, 21, 213–220. [Google Scholar] [CrossRef]
- Jeanne, C.L.L.; Maria, E.B.D.M.; Silene, C.D.N.; Monica, L.A. Differential genomic damage in different tumor lines induced by prodigiosin. Anticancer Res. 2015, 35, 3325–3332. [Google Scholar]
- Lin, S.R.; Lin, C.S.; Chen, C.C.; Tseng, F.J.; Wu, T.J.; Weng, L.; Weng, C.F. Doxorubicin metabolism moderately attributes to putative toxicity in prodigiosin/doxorubicin synergism in vitro cells. Mol. Cell Biochem. 2020, 475, 119–126. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The structure and physicochemical properties of prodigiosin.

Table 1.
The substrate sources for prodigiosin production by liquid-state fermentation.
| Bacteria | Main Substrate | Yield (mg/L−1) | Reference |
|---|---|---|---|
| Commercial media | |||
| S.marcescens SMΔR | Luria–Bertani broth | 50 | [53] |
| S. marcescens SS-1 | Luria–Bertani broth | 32 | [54] |
| Yeast extract | 690 | ||
| Yeast extract, axit aspartic | 1400 | ||
| Yeast extract, histidine | 1400 | ||
| Yeast extract, proline | 2500 | ||
| S. marcescens | Tryptone, glycerol | 123 | [55] |
| S. marcescens FZSF02 | Soya peptone | 1774 | [56] |
| Beef extract | 1699 | ||
| Tryptone | 353 | ||
| Yeast extract | 380 | ||
| S. marcescens | Nutrient broth | 15 | [57] |
| Peptone glycerol | 12 | ||
| Tryptone soy broth | 11 | ||
| Luria–Bertani broth | 10 | ||
| Glycerol beef broth | 8 | ||
| S. marcescens | Nutrient broth | 510 | [58] |
| Peptone glycerol broth | 300 | ||
| Nutrient broth, maltose | 1820 | ||
| Pseudomonas putida | Luria–Bertani broth | 17 | [59] |
| Terrific broth | 94 | ||
| S. marcescens JNB 5-1 | Luria–Bertani broth | 5830 | [60] |
| Low-cost organic materials | |||
| S.marcescens SMΔR | 4% soybean oil, Luria–Bertani broth | 525 | [53] |
| 4% olive oil, Luria–Bertani broth | 579 | ||
| 6% sunflower oil, Luria–Bertani broth | 790 | ||
| S. marcescens FZSF02 | Peanut powder | 3762 | [56] |
| Peanut powder, soya peptone | 1588 | ||
| Peanut powder, beef extract | 5062 | ||
| Fish meal | 0 | ||
| Soybean powder | 0 | ||
| Corn steep liquor | 0 | ||
| Peanut powder, olive oil, beef extract | 13,622 | ||
| S. marcescens | Peanut seed broth | 37,600 | [58] |
| Sesame seed broth | 16,500 | ||
| S. marcescens | Peanut seed broth | 38,750 | [61] |
| Peanut oil broth | 2890 | ||
| Sesame seed broth | 16,680 | ||
| Sesame oil broth | 767 | ||
| Copra seed broth | 1940 | ||
| Coconut oil broth | 1420 | ||
| S. marcescens | Peanut powder, defatted soybean flour | 1282 | [62] |
| S. marcescens BWL1001 | Soybean oil, peptone (100/10) | 27,650 | [63] |
| Streptomyces fusant NRCF69 | Peanut seed broth | 42,030 | [64] |
| Sunflower oil broth | 40,110 | ||
| Organic byproduct/waste source | |||
| Fishery processing byproducts | |||
| S. marcescens TNU02 | Demineralized crab shell powder | 4514 | [2] |
| S. marcescens TNU01 | Squid pen powder | 3790 | [29] |
| S. marcescens TNU01 | Demineralized shrimp shell powder | 5910 | [30] |
| S. marcescens CC17 | Shrimp head powder | 5355 | [31] |
| S. marcescens TKU011 | Squid pen powder | 978 | [65] |
| Agro-industrial byproducts | |||
| S. marcescens TNU01 | Cassava wastewater | 5202 | [32] |
| S. marcescens TUN02 | Peanut oil cake | 5380 | [33] |
| S. marcescens UCP 1549 | Cassava wastewater | 49,500 | [66] |
| S. marcescens | Peanut oil cake | 40.9 | [67] |
| S. marcescens CF-53 | Peanut oil cake | 39,800 | [68] |
| S. marcescens | Soybean meal | 9632 | [69] |
| S. marcescens UCP 1549 | Corn bran | 1680 | [70] |
| S. marcescens MO-1 | Ram horn peptone | 277.74 | [71] |
| S. marcescens UTM1 | Brown sugar | 8109 | [72] |
| Streptomyces fusant NRCF69 | Dairy processing wastewater broth, | 36,700 | [64] |
| Dairy processing wastewater broth, 0.5% mannitol | 47,000 | ||
| Other byproducts/wastes | |||
| S. marcescens UCCM 00009 | Feather and waste frying oil | 9660 | [34] |
| S. marcescens | Food waste, Rice husk | 7890 | [73] |
| S. marcescens NPLR1 | Tannery solid waste fleshing | 33,000 | [74] |
| S. marcescens | Kitchen waste, peptone, proline | 890 | [75] |
Table 2.
A summary of the substrate sources for prodigiosin production in solid-state fermentation.
| Bacteria | Main Substrate | Yield (mg/g−1) | Reference |
|---|---|---|---|
| S. marcescens | Ground corn | 3.4 | [51] |
| Wheat bran | 47.5 | ||
| Rice husk | 11.7 | ||
| Soya bean ground | 10.2 | ||
| Wheat bran, sunflower oil, live cells of Bacillus subtilis | 240 | ||
| S. marcescens UCP 1549 | Soybean oil waste, wheat bran | 119.8 | [82] |
| Wheat bran | 119.8 | ||
| Sugarcane bagasse | 1.8 | ||
| Instant noodle waste | 66.2 | ||
| Tangerine peels | 22.1 | ||
| Pineapple peels | 18.57 | ||
| Pineapple crown | 31.47 | ||
| S. marcescens Xd-1 | Bagasse | 20.13 | [83] |
| Wood powder | 15 | ||
| Wheat straw powder | 10.6 | ||
| Bagasse, glycerol, bagasse soy peptone | 40.86 | ||
| S. marcescens | Tannery fleshing waste | 70.4 | [84] |
| S. marcescens UCP/WFCC1549 | Mannitol | 3.2 | [85] |
| S. matodiphilia NCIM 5606 | Wheat bran | 1.307 mg/L−1 | [86] |
| Sweet lemon peel | 0.169 mg/L−1 | ||
| Orange peel | 0.149 mg/L−1 | ||
| Pigeon pea peel | 0.008 mg/L−1 | ||
| Rice bran | 0.066 mg/L−1 | ||
| S. marcescens wild | Peanut powder | 858 | [90] |
| S. marcescens EMS5 | 1304 |
Table 4.
The efficiency of prodigiosin production in large-scale (bioreactor system).
| Strain | Substrate | Max Prodigiosin Yield (mg/L−1) in Fermentation Time (hours) | Culture Volume (L)/Bioreactor (L) | Enhancing Yield (Fold) | Reference | |
|---|---|---|---|---|---|---|
| In Flask | Bioreactor | |||||
| S. marcescens TUN02 | demineralized crab shell powder, casein, (NH4)2SO4, K2HPO4 | 4514 (36 h) | 5100 (8 h) | 4.5 L (15 L) | 1.1 | [2] |
| S. marcescens TNU01 | Squid pens powder, K2HPO4, MgSO4 | 3790 (48 h) | 3450 (12 h) | 3 L (10 L) | - | [29] |
| S. marcescens TUN02 | Demineralized shrimp shell powder, casein, K2SO4, K2HPO4 | 5910 (36 h) | 6200 (8 h) | 5 L (15 L) | 1 | [30] |
| S. marcescens CC17 | Shrimp head powder, casein, K2SO4, Ca3(PO4)2 | 5355 (60 h) | 6310 (8 h) | 6.75 L (12 L) | 1.2 | [31] |
| S. marcescens TNU01 | Cassava wastewater, casein, MgSO4, K2HPO4 | 5202 (48 h) | 6150 (8 h) | 7 L (14 L) | 1.1 | [32] |
| S. marcescens TUN02 | Peanut oil cake | 5380 (48 h) | 6886 (10 h) | 4 L (14 L) | 1.3 | [33] |
| S. marcescens | Peanut oil cake | 40.9 (30 h) | 50 (30 h) | 1.5 L (3 L) | 1.2 | [67] |
| S. marcescens CF-53 | Peanut oil cake | 39,800 (42 h) | 40,000 (42 h) | 1 L (2 L) | 1 | [68] |
| S. marcescens UTM1 | Brown sugar | 237 (24 h) | 8109 (24 h) | 5L (5L) | 34.2 | [72] |
| S. marcescens B6 | Two-step feeding strategy with glycerol | ND | 583 (30 h) | 2.5 L (5 L) | ND | [115] |
| Hahella chejuensis | Glucose (Continuous fermentation) | 448.1 (24 h) | 2280 (49 h) | 5 L (10 L) | 5 | [116] |
| 1305 (240 h) | 100 L (200 L) | 2.9 | ||||
| S. marcescens NS-17 | Maltose, peptone, Tween-80, soybean oil, NaCl, KCl | 60.5 (56 h) | 4644.6 (56 h) | 47.8 L (50 L) | 76.7 | [117] |
| Serratia sp. KH-95 | HP-20 resin, casein, K2HPO4, MgSO4, NaCl | ND | 13,100 (30 h) | 1 L (2.5 L) | ND | [118] |
| S. marcescens | Sucrose, peptone | 391.1 (48 h) | 595 (48 h) | 6.5 L (7 L) | 1.5 | [119] |
| S. marcescens ATCC 27117 | Nutrient broth | 13,600 (24 h) | 7800 (20 h) | 3 L (4.5 L) | - | [120] |
| S. marcescens BS 303 | Peptone, glycerol, mineral broth, TritonX-114 | 540 (24 h) | 872 (62 h) | 0.935 L (1.5 L) | ND | [122] |
| Serratia AM8887 | Fertilizer waste, sucrose, glycerol, NaCl | ND | 7316 (19 h) | 17.8 L (20 L) | ND | [123] |
ND: No determine.
Table 5.
Anti-fungal activity of prodigiosin for agriculture.
| Fungi | The Unit of Antifungal Activity | Value | Reference |
|---|---|---|---|
| Aspergillus flavus | MIC—Minimum inhibitory concentration (µg/mL) | 10 | [23] |
| Fusarium oxysporum | 8 | ||
| Aspergillus niger | 230 | [74] | |
| Fusarium moniliforme | 210 | ||
| Helminthosporium sativum | Diameter of inhibition zone (mm) | 42 | [100] |
| Curvularia lunata | 40 | ||
| Alternaria alternate | 40 | ||
| Fusarium oxysporum | 30 | ||
| Cercospora apii | 24 | ||
| Rhizoctonia solani | 11 | ||
| Alternaria sp. | MIC—Minimum inhibitory concentration (µg/mL) | 80 | [124] |
| Fusarium sp. | 160 | ||
| Didymella applanata | IC50—Concentration to inhibit 50% fungal (nmol/mL) | 2.5 | [127] |
| Phoma lingam | Hyphal growth diameter (%) | 25 | [128] |
| Sclerotinia sclerotiorum | 60 | ||
| Botrytis cinerea | Inhibition of spore germination (%) | 0–80 | [129] |
| Mycosphaerella fijiensis | IC50—Concentration to inhibit 50% fungal (µg/mL) | 996 | [130] |
| Inhibits growing germ tubes (%) | 63 | ||
| Cochliobolus miyabeanus | Growth inhibition (%) | 83.3 | [131] |
| Fusarium moniliforme | 5.9 | ||
| F. oxysporum f. sp. allii | 17.6 | ||
| F. oxysporum f. sp. raphani | 16.3 | ||
| F. oxysporum f. sp. cucumerinum | 23.5 | ||
| F. oxysporum f. sp. spinaciae | 1.2 | ||
| F. oxysporum f. sp. cepae | 17.8 | ||
| F. roseum | 5.8 | ||
| F. solani var. coeruleum | 14.3 | ||
| F. ventricosum | 11.1 | ||
| Phytophthora cactorum | 89.7 | ||
| P. capsici | 63.5 | ||
| P. castaneae | 74.5 | ||
| P. citrophthora | 85.1 | ||
| P. infestans sp. | 80.5–83.2 | ||
| P. melonis | 93 | ||
| Pyricularia oryzae | 28.8 | ||
| Pythium spinosum | 66.4 | ||
| Pythium ultimum | 44.2 | ||
| Rhizoctonia solani sp. | 17.6–52.9 | ||
| Pythium myriotylum | Growth inhibition (%) | 71.33 | [132] |
| Rhizoctonia solani | 61.33 | ||
| Sclerotium rolfsii | 49.33 | ||
| Phytophthora infestans | 48.66 | ||
| Fusarium oxysporum | 31 | ||
| Colletotrichum nymphaeae | Inhibits germination (%) | 100 | [133] |
Table 6.
The bioactivity of prodigiosin against bacteria, nematodes, and insects harmful to crops.
| Object | The Unit of Activity | Value | Reference |
|---|---|---|---|
| Bacteria | |||
| Acidovonax avenae | MAC—The maximal allowable concentration (µg/mL−1) | 50 | [131] |
| Agrobacterium tumefaciens | 50 | ||
| Clavibacter michiganensis subsp. michiganesis | 6.3 | ||
| Erwinia carotovora subsp. carotovora | 25 | ||
| E. herbicola | 50 | ||
| Pseudomonas cichorii | >100 | ||
| P. fluorecens | >100 | ||
| P. gladioli pv. gladioli | >100 | ||
| P. glumae | >100 | ||
| P. mariginalis pv. marginalis | >100 | ||
| P. syringae pv. lachrymans | >100 | ||
| P. syringae pv. mori | >100 | ||
| P. syringae pv. phaseolicola | >100 | ||
| P. syringae pv. pisi | >100 | ||
| Ralstonia solanacearum sp. | >100 | ||
| Xanthomonas campetris pv. campestris | 25 | ||
| X. campetris pv. carotae | 50 | ||
| X. campetris pv. oryzae | 25.5 | ||
| Nematode | |||
| Heterodera schachtii | IC50 (μM) | 13.3 | [128] |
| Radopholus similis | IC50 (µg/mL−1) | 83 | [134] |
| Meloidogyne javanica | 79 | ||
| Meloidogyne incognita | IC50—Anti juvenile (mg/mL−1) | 0.2 | [33] |
| IC50—Anti egg-hatching (mg/mL−1) | 0.32 | ||
| IC50—Anti juvenile (mg/mL−1) | 31.9 | [135] | |
| Insect | |||
| Diaphorina citri | The inhibitory rate of oviposition (%) | 42 | [136] |
| The moderate inhibitory rate of egg hatch (%) | 26 | ||
| Helicoverpa armigera | Larval mortality rate (%) | 70–100 | [137] |
| Spodoptera litura | |||
| Drosophila | IC50—Anti larval (ppm) | 230 | [138] |
| IC50—Anti larval (g/L−1) | 0.23 | [65] | |
| Spodoptera litura | The mortality at 8µg/g diet (%) | 3 | [139] |
| Plutella xylostella | 96 | ||
| Adoxophyes honmai | 12 | ||
IC50—Concentration to inhibit 50%.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Nguyen, T.H.; Wang, S.-L.; Nguyen, V.B. Recent Advances in Eco-Friendly and Scaling-Up Bioproduction of Prodigiosin and Its Potential Applications in Agriculture. Agronomy 2022, 12, 3099. https://doi.org/10.3390/agronomy12123099
AMA Style
Nguyen TH, Wang S-L, Nguyen VB. Recent Advances in Eco-Friendly and Scaling-Up Bioproduction of Prodigiosin and Its Potential Applications in Agriculture. Agronomy. 2022; 12(12):3099. https://doi.org/10.3390/agronomy12123099
Chicago/Turabian StyleNguyen, Thi Hanh, San-Lang Wang, and Van Bon Nguyen. 2022. "Recent Advances in Eco-Friendly and Scaling-Up Bioproduction of Prodigiosin and Its Potential Applications in Agriculture" Agronomy 12, no. 12: 3099. https://doi.org/10.3390/agronomy12123099
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.