
Phenotypic Variation of Sorghum Accessions for Grain Yield and Quality Traits


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Site
2.2. Yield Determination
2.3. Moisture, Protein, and Starch Content Determination
2.4. Tannin Content Determination
2.5. Data Analysis
3. Results
3.1. Protein, Starch, Moisture and Grain Yield under First and Second Growing Seasons
3.2. Analysis of Variance
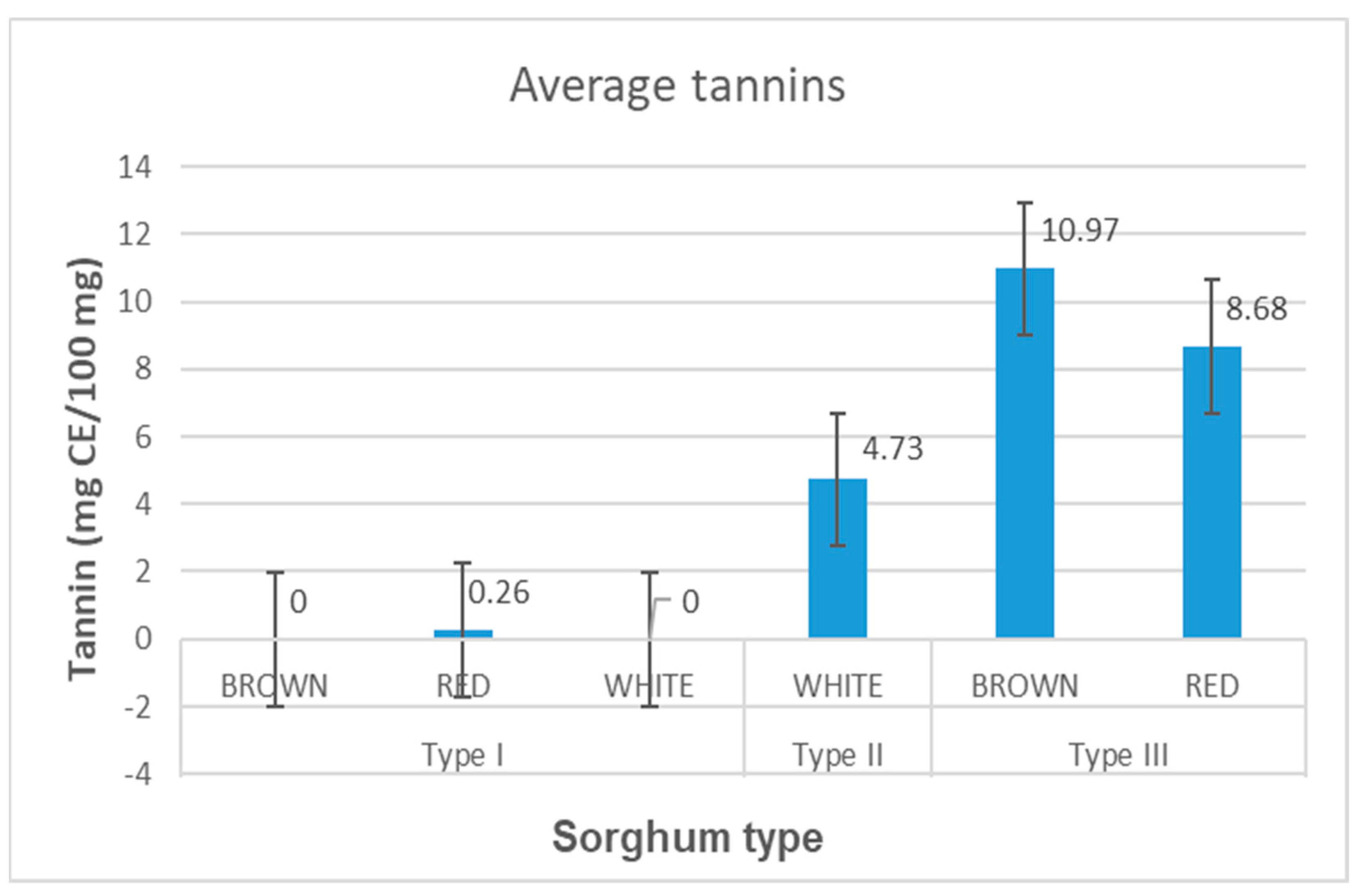
3.3. Tannin Content, Grain Color and Sorghum Type Characterization
3.4. Correlation Coefficients
3.5. Potential Accessions Selected for Future Breeding
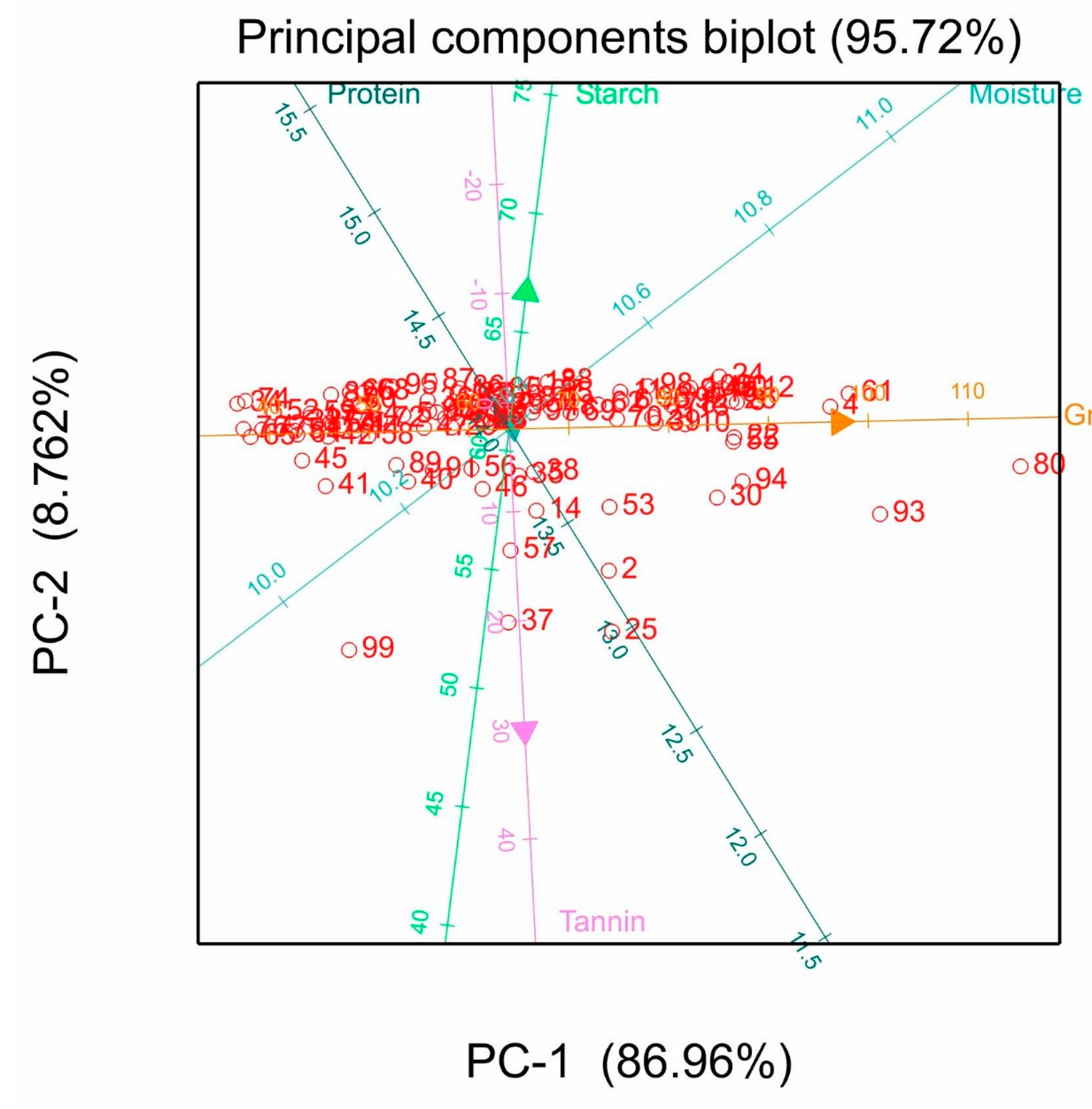
3.6. Principal Component Analysis
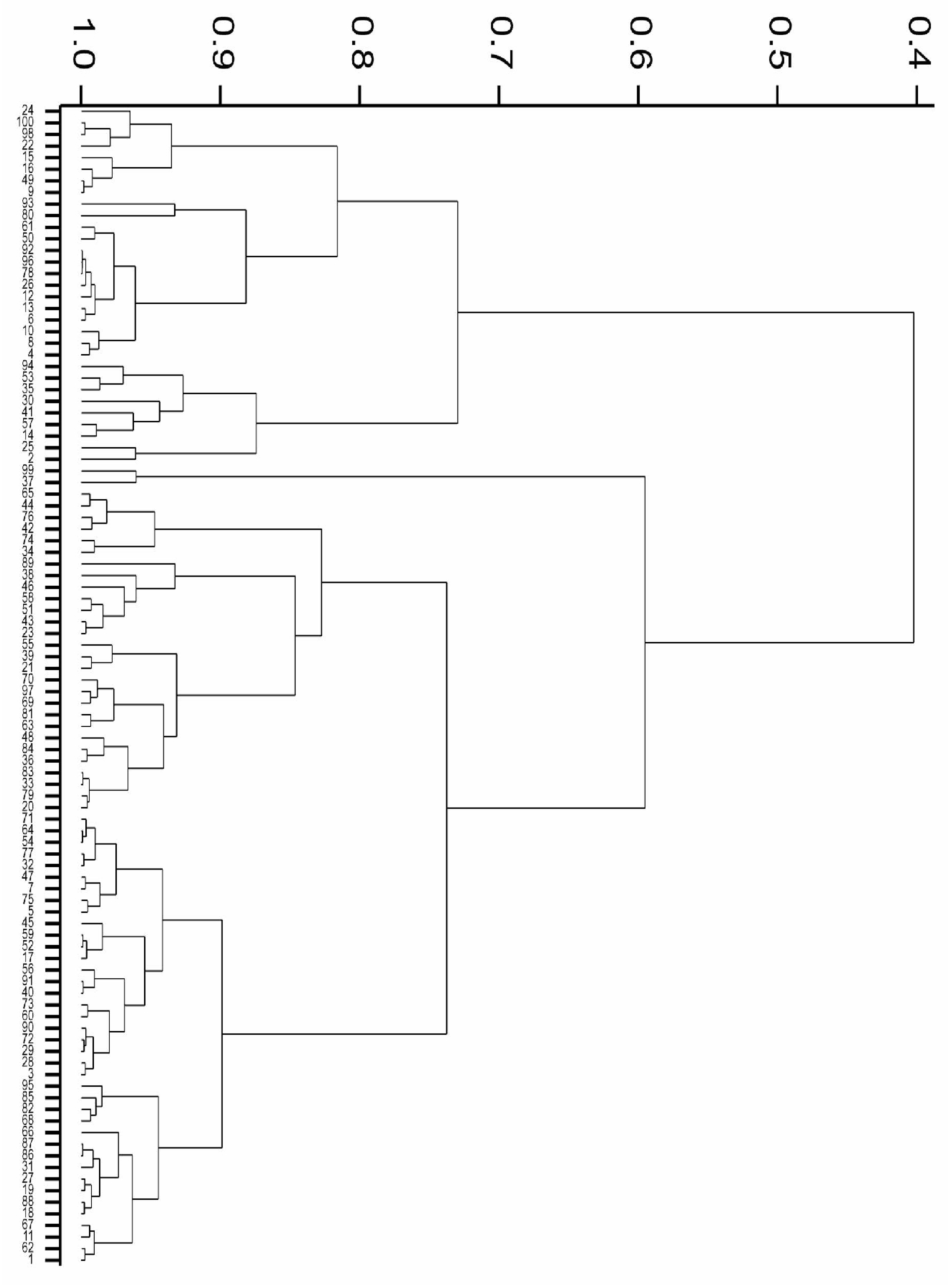
3.7. Cluster Analysis
4. Discussion
4.1. Variation in Grain Yield, Protein and Starch Contents
4.2. Tannin Content
4.3. Heritability Estimates
4.4. Correlations
4.5. Principal Component Analysis
4.6. Cluster Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- United States Grains Council. Sorghum. 2006. Available online: www.grains.org/page.ww?section=Barley%2C+Corn+%26+Sorghum&name=Sorghum (accessed on 29 September 2020).
- Shegro, A.G.; Labuschagne, M.T.; Van Biljon, A.; Shargie, N.G. Genetic variability among sorghum accessions for seed starch and stalk total sugar content. Sci. Agric. 2014, 71, 472–479. [Google Scholar]
- Tripathi, A.D.; Mishra, R.; Maurya, K.K.; Singh, R.B.; Wilson, D.W. Estimates for world population and global food availability for global health. In The Role of Functional Food Security in Global Health; Singh, R.B., Watson, R.R., Takahashi, T., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 3–24. [Google Scholar]
- Carter, P.R.; Hicks, D.R.; Oplinger, E.S.; Doll, J.D.; Bundy, L.G.; Schuler, R.T.; Holmes, B.J. “Grain Sorghum (Milo)”. Alternative Field Crops Manual. 1989. Available online: www.hort.purdue.edu/newcrop/afcm/sorghum.html (accessed on 29 September 2020).
- Crop Plant Resources Sorghum. Sorghum Bicolor. 2000. Available online: http://darwin.nmsu.edu/~molbio/plant/sorghum.html (accessed on 30 September 2020).
- Department of Agriculture, Forestry and Fisheries (DAFF). Sorghum Production Guideline; DAFF: Pretoria, South Africa, 2017; p. 20.
- Food and Agricultural Organization (FAO). International Crops Research Institute for the Semi-Arid Tropics (ICRISAT), The World Sorghum and Millet Economies. Facts, Trends and Outlook. 1996. Available online: http://www.fao.org/ (accessed on 31 September 2020).
- Olack, B.; Burke, H.; Cosmas, L.; Bamrah, S.; Dooling, K.; Feikin, D.R.; Breiman, R.F. Nutritional status of under-five children living in an informal urban settlement in Nairobi, Kenya. J. Health Popul. Nutr. 2011, 29, 357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Valença, A.W.; Bake, A.; Brouwer, I.D.; Giller, K.E. Agronomic biofortification of crops to fight hidden hunger in sub-Saharan Africa. Glob. Food Secur. 2017, 12, 8–14. [Google Scholar] [CrossRef]
- Food and Agriculture Organization Corporate Statistical Database (FAOSTAT) Data Production Yearbook; FAO: Rome, Italy, 2008; p. 49.
- Kim, J.S.; Hyun, T.K.; Kim, M.J. The inhibitory effects of ethanol extracts from sorghum, foxtail millet and proso millet on α-glucosidase and α-amylase activities. Food Chem. 2011, 124, 1647–1651. [Google Scholar] [CrossRef]
- Oh, K.K.; Adnan, M.; Cho, D.H. Network pharmacology of bioactives from Sorghum bicolor with targets related to diabetes mellitus. PLoS ONE 2020, 15, e0240873. [Google Scholar] [CrossRef] [PubMed]
- Hamaker, B.R.; Bugusu, B.A. Overview: Sorghum proteins and food quality. In Proceedings of the Workshop on the Proteins of Sorghum and Millets: Enhancing Nutritional and Functional Properties for Africa, Pretoria, South Africa, 2–4 April 2003. [Google Scholar]
- Zhao, Z. The Africa Biofortified Sorghum Project Applying Biotechnology to Develop Nutritionally Improved Sorghum for Africa. In Biotechnology and Sustainable Agriculture 2006 and Beyond; Springer: Dordrecht, The Netherlands, 2007; pp. 273–277. [Google Scholar]
- Selle, P.H.; Cadoganb, D.J.; Li, X.; Bryden, W.L. Implications of sorghum in broiler chicken nutrition. Anim. Feed. Sci. Technol. 2010, 156, 57–74. [Google Scholar] [CrossRef]
- Mahmood, S.; Ali, H.; Ahmad, F.; Iqbal, Z. Estimation of tannins in different sorghum varieties and their effects on nutrient digestibility and absorption of some minerals in caged White Leghorn layers. Int. J. Agric. Biol. 2014, 16, 217–221. [Google Scholar]
- Awika, J.M.; Rooney, L.W. Sorghum phytochemicals and their potential impact on human health. Phytochemistry 2004, 65, 1199–1221. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.; Sun, Y.; Halgreen, L. The relationships of sorghum kernel pericarp and testa characteristics with tannin content. Asian J. Crop Sci. 2009, 1, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Sejake, T.; Shargie, N.; Christian, R.; Tsilo, T. Assessment of genetic diversity in sorghum germplasm using agro-morphological traits. S. Afr. J. Plant Soil 2020, 37, 376–388. [Google Scholar] [CrossRef]
- Maxson, E.D.; Rooney, L.W. Evaluation of methods of tannin analysis in sorghum grain. Cereal Chem. 1972, 49, 719–729. [Google Scholar]
- Price, M.L.; Van Scoyoc, S.; Butler, L.G. A critical evaluation of the vanillin reaction as an assay for tannin in sorghum grain. J. Agric. Food Chem. 1978, 26, 1214–1218. [Google Scholar] [CrossRef]
- VSN International. GenStat for Windows, 18th ed.; VSN International: Hemel Hempstead, UK, 2015. [Google Scholar]
- SPSS. Institute Statistical Package for Social Sciences-Users Guide; SPSS: Chicago, IL, USA, 2006. [Google Scholar]
- Tsilo, T.J.; Kolmer, J.A.; Anderson, J.A. Molecular mapping and improvement of leaf rust resistance in wheat breeding lines. Phytopathology 2014, 104, 865–870. [Google Scholar] [CrossRef] [PubMed]
- Mofokeng, M.A.; Shimelis, H.; Tongoona, P.; Laing, M.D. Protein content and amino acid composition among selected South African sorghum genotypes. J. Agric. Food Chem. 2018, 3, 27–34. [Google Scholar] [CrossRef] [Green Version]
- Ng’uni, D.; Geleta, M.; Hofvander, P.; Fatih, M.; Bryngelsson, T. Comparative genetic diversity and nutritional quality variation among some important Southern African sorghum accessions (Sorghum bicolor L. Moench). Afr. Crop Sci. J. 2012, 6, 56–64. [Google Scholar]
- Shegro, A.; Shargie, N.G.; Van Biljon, A.; Labuschagne, M.T. Diversity in starch, protein and mineral composition of sorghum landrace accessions from Ethiopia. J. Crop Sci. Biotechnol. 2012, 15, 275–280. [Google Scholar] [CrossRef]
- Perez-Maldonado, R.A.; Rodriguez, H.D. Nutritional Characteristics of 26 Sorghums in Queensland and New South Wale for Chicken Meat Production; (RIRDC Publication 7); RIRDC: Barton, Australia, 2007. [Google Scholar]
- Dicko, M.H.; Gruppen, H.; Traore, A.S.; Alphons, G.J.; Voragen, A.J.; Van Berkel, W.J. Sorghum grain as human food in Africa: Relevance of content of starch and amylase activities. Afr. J. Biotechnol. 2006, 5, 384–395. [Google Scholar]
- Reichert, R.D.; Mwasaru, M.A.; Mukuru, S.Z. Characterization of coloured-grain sorghum lines and identification of high tannin lines with good dehulling characteristics. Cereal Chem. 1988, 65, 156–170. [Google Scholar]
- Dykes, L.; Rooney, L.W. Sorghum and millet phenols and antioxidants. J. Cereal Sci. 2006, 44, 236–251. [Google Scholar] [CrossRef]
- Lu, Q.; Sun, Y. Genetic Improvement of Hybrid Sorghum; China Agricultural Science and Technology Publisher: Beijing, China, 2005. [Google Scholar]
- Chatfield, C.; Collin, A.J. Introduction to Multivariate Analysis; Chapman and Hall in Association with Methuen: New York, NY, USA, 1980. [Google Scholar]
- Hair, J.F.; Anderson, R.E.; Tatham, R.L.; Black, W.C. Multivariate Data Analysis, 5th ed.; Prentice Hall: Hoboken, NJ, USA, 1998; Available online: https://www.amazon.com/Multivariate-Data-Analysis-Joseph-Hair/dp/0138948585 (accessed on 31 October 2022).
- Felix, K.; Cheruiyot, E.; Mwendia, C.M.; Francis, W.N.; Kimani, P. Genetic variation of biochemical characteristics of selected sorghum varieties from East Africa. Afr. J. Food Sci. 2015, 9, 314–321. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Source of Variance | d.f. | Grain Yield and Quality Traits | ||||
|---|---|---|---|---|---|---|
| Protein | Starch | Tannin | Grain Yield | Moisture | ||
| Season | 1 | 11.27 ** | 431.51 ** | - | 20,769.63 ** | 9.79 ** |
| Replication | 2 | 0.19 ns | 4.75 ns | 0.24 ns | 1507.50 ** | 0.02 ns |
| Genotype | 99 | 9.38 ** | 80.43 ** | 78.17 ** | 1330.36 ** | 0.87 ** |
| Genotype × Season | 99 | 4.83 ** | 44.49 ** | - | 395.30 ** | 0.57 ** |
| Residual | 298 | 1.03 | 11.67 | 0.80 | 50.03 | 0.03 |
| h2B | 49 | 45 | - | 70 | 34 | |
| SE | 1.01 | 3.42 | 0.80 | 7.07 | 0.17 | |
| CV (%) | 7.26 | 5.60 | 1.8 | 11.04 | 1.61 | |
| No. | Accession | Protein (%) | Starch (%) | Tannin (mg CE/100 mg) | |
|---|---|---|---|---|---|
| 4 | Lenthate | 14.81 ± 0.4122 hijklmnopqrstu | 62.33 ± 1.404 cdefghijklmnopqr | 0.00 ± 0.4552 a | |
| 6 | Mabele a sesotho | 13.92 ± 0.4122 cdefghijklmnopqrst | 62.40 ± 1.404 cdefghijklmnopqr | 0.34 ± 0.4552 ab | |
| 8 | Mammopane | 13.89 ± 0.4130 cdefghijklmnopqrst | 65.37 ± 1.407 jklmnopqr | 0.48 ± 0.4552 ab | |
| 10 | Manthate | 13.87 ± 0.4122 cdefghijklmnopqrs | 57.74 ± 1.404 bcdefghijklmno | 0.87 ± 0.4552 ab | |
| 11 | Mapimkana | 13.09 ± 0.4122 bcdefghijklmno | 59.92 ± 1.404 bcdefghijklmnopqr | 0.25 ± 0.4552 a | |
| 12 | Maseka a swere | 14.54 ± 0.4122 fghijklmnopqrstu | 65.12 ± 1.404 ijklmnopqr | 0.35 ± 0.4552 a | |
| 13 | SA 0009 | 13.65 ± 0.4132 bcdefghijklmnopqrs | 60.88 ± 1.407 cdefghijklmnopqr | 0.00 ± 0.4552 bcde | |
| 16 | SA 0062 | 13.91 ± 0.4130 cdefghijklmnopqrst | 60.51 ± 1.407 bcdefghijklmnopqr | 0.34 ± 0.4552 a | |
| 22 | SA 0133 | 11.77 ± 0.4122 abcde | 61.98 ± 1.404 cdefghijklmnopqr | 4.27 ± 0.4552 cde | |
| 24 | SA 0213 | 14.40 ± 0.4130 fghijklmnopqrstu | 62.94 ± 1.407 cdefghijklmnopqr | 0.00 ± 0.4552 a | |
| 25 | SA 0223 | 12.02 ± 0.4122 abcdef | 59.72 ± 1.404 bcdefghijklmnopqr | 24.40 ± 0.4552 k | |
| 26 | SA 0260 | 14.53 ± 0.4122 fghijklmnopqrstu | 61.47 ± 1.404 cdefghijklmnopqr | 0.13 ± 0.4552 a | |
| 30 | SA 0390 | 10.41 ± 0.4130 a | 61.05 ± 1.406 cdefghijklmnopqr | 10.4 ± 0.45521 ghi | |
| 38 | SA 0567 | 15.53 ± 0.4122 nopqrstu | 54.83 ± 1.404 abcd | 4.52 ± 0.4552 de | |
| 49 | SA 0672 | 13.75 ± 0.4130 bcdefghijklmnopqrs | 63.94 ± 1.406 efghijklmnopqr | 0.00 ± 0.4552 ab | |
| 50 | SA 0673 | 11.23 ± 0.4130 ab | 67.43 ± 1.406 qr | 0.00 ± 0.4552 a | |
| 53 | SA 0696 | 11.98 ± 0.4122 abcdef | 65.05 ± 1.404 ijklmnopqr | 13.05 ± 0.4552 i | |
| 55 | SA 0718 | 14.20 ± 0.4122 efghijklmnopqrstu | 57.04 ± 1.404 abcdefghijkl | 2.61 ± 0.4552 abcd | |
| 61 | SA 0954 | 12.59 ± 0.4130 abcdefghijk | 64.78 ± 1.406 hijklmnopqr | 0.00 ± 0.4552 a | |
| 73 | SA 1730 | 13.44 ± 0.4122 bcdefghijklmnopq | 61.78 ± 1.404 cdefghijklmnopqr | 1.67 ± 0.4552 abcd | |
| 80 | SA 2097 | 12.00 ± 0.4122 abcdef | 65.33 ± 1.404 jklmnopqr | 9.69 ± 0.4552 gh | |
| 91 | SA 2913 | 13.87 ± 0.4130 cdefghijklmnopqrs | 62.68 ± 1.406 cdefghijklmnopqr | 7.58 ± 0.4552 fg | |
| 93 | SA 3710 | 14.48 ± 0.4122 fghijklmnopqrstu | 57.82 ± 1.404 bcdefghijklmno | 11.32 ± 0.4552 hi | |
| 94 | SA 3711 | 12.39 ± 0.4130 abcdefghi | 60.90 ± 1.407 cdefghijklmnopqr | 8.68 ± 0.4552 fgh | |
| 100 | Wahi | 12.96 ± 0.4130 abcdefghijklm | 64.79 ± 1.407 hijklmnopqr | 0.29 ± 0.4552 a | |
| Mean | 13.95 | 60.91 | 2.24 | ||
| Ranges | 10.41–16.59 | 48.90–67.61 | 0–24.40 | ||
| No. | Accession | Grain yield (g/panicle) | Moisture (%) | Grain Color | Sorghum Type |
| 4 | Lenthate | 96.50 ± 6.065 nop | 10.60 ± 0.1607 fghijklm | Brown | I |
| 6 | Mabele a sesotho | 75.34 ± 6.065 bcdefghijklmno | 10.48 ± 0.1607 bcdefghijklm | Red | I |
| 8 | Mammopane | 84.30 ± 6.066 ghijklmnop | 10.82 ± 0.1611 hijklm | Brown | I |
| 10 | Manthate | 81.98 ± 6.054 fghijklmnop | 10.81 ± 0.1607 hijklm | Red | I |
| 11 | Mapimkana | 96.48 ± 6.065 mnop | 10.75 ± 0.1607 hijklm | Red | I |
| 12 | Maseka a swere | 88.10 ± 6.065 hijklmnop | 10.55 ± 0.1607 cdefghijklm | White | I |
| 13 | SA 0009 | 87.05 ± 6.072 jklmnop | 10.88 ± 0.1611 ijklm | White | I |
| 16 | SA 0062 | 81.73 ± 6.065 efghijklmnop | 10.48 ± 0.1611 bcdefghijklm | Brown | I |
| 22 | SA 0133 | 86.49 ± 6.065 jklmnop | 10.98 ± 0.1607 klm | Brown | III |
| 24 | SA 0213 | 84.51 ± 6.072 ghijklmnop | 11.08 ± 0.1611 lm | Red | I |
| 25 | SA 0223 | 73.38 ± 6.065 abcdefghijklmno | 10.50 ± 0.1607 bcdefghijklm | Brown | III |
| 26 | SA 0260 | 86.50 ± 6.065 ijklmnop | 10.26 ± 0.1607 abcdefghijkl | White | I |
| 30 | SA 0390 | 84.50 ± 6.072 ghijklmnop | 10.88 ± 0.1610 ijklm | Brown | III |
| 38 | SA 0567 | 66.76 ± 6.065 abcdefghijklmno | 9.55 ± 0.1607 abcd | White | II |
| 49 | SA 0672 | 88.13 ± 6.072 lmnop | 10.26 ± 0.1610 abcdefghijkl | Brown | I |
| 50 | SA 0673 | 84.89 ± 6.065 hijklmnop | 11.28 ± 0.1610 m | Brown | I |
| 53 | SA 0696 | 73.38 ± 6.066 abcdefghijklmno | 10.61 ± 0.1607 ghijklm | Brown | III |
| 55 | SA 0718 | 86.78 ± 6.065 ijklmnop | 10.19 ± 0.1607 abcdefghijkl | White | II |
| 61 | SA 0954 | 89.30 ± 6.074 ghijklmnop | 10.65 ± 0.1610 ghijklm | White | I |
| 73 | SA 1730 | 45.96 ± 6.066 abcdefg | 10.85 ± 0.1607 ijklm | White | II |
| 80 | SA 2097 | 114.90 ± 6.065 p | 10.59 ± 0.1607 efghijklm | Brown | III |
| 91 | SA 2913 | 56.00 ± 6.065 abcdefghijkl | 10.58 ± 0.1610 efghijklm | White | II |
| 93 | SA 3710 | 101.12 ± 6.066 op | 10.52 ± 0.1607 bcdefghijklm | White | II |
| 94 | SA 3711 | 87.20 ± 6.072 klmnop | 10.48 ± 0.1611 bcdefghijklm | Red | III |
| 100 | Wahi | 75.09 ± 6.074 abcdefghijklmno | 10.50 ± 0.1611 bcdefghijklm | White | I |
| Mean | 64.03 | 10.37 | |||
| Ranges | 36.7–114.9 | 9.36–11.28 | |||
| Crude Protein | Starch | Tannin | Grain Yield | |
|---|---|---|---|---|
| Starch | −0.356 ** | |||
| Tannin | −0.300 ** | −0.265 ** | ||
| Grain yield | −0.273 ** | 0.209 * | 0.142 | |
| Moisture | −0.609 ** | 0.428 ** | 0.056 | 0.440 ** |
| No. | Accession | Protein (%) | Starch (%) | Tannin (mg CE/100 mg) | Grain Yield (g/Panicle) | Moisture (%) |
|---|---|---|---|---|---|---|
| Selected accessions for starch content | ||||||
| 24 | SA 0213 | 11.23 | 67.43 | 0.00 | 84.89 | 11.28 |
| 98 | SA0139 | 12.13 | 66.10 | 0.30 | 77.75 | 10.71 |
| 49 | SA 0672 | 13.89 | 65.37 | 0.48 | 84.30 | 10.82 |
| 50 | SA 0673 | 14.54 | 65.12 | 0.35 | 85.34 | 10.55 |
| 9 | Mamolokwane | 13.89 | 64.80 | 0.48 | 81.86 | 10.99 |
| 11 | Mapimkana | 12.96 | 64.79 | 0.29 | 75.09 | 10.50 |
| 100 | Wahi | 12.59 | 64.78 | 0.00 | 82.15 | 10.65 |
| Selected accessions for crude protein content | ||||||
| 39 | SA 0572 | 16.02 | 56.10 | 0.00 | 79.17 | 10.13 |
| 70 | SA 1564 | 15.65 | 57.26 | 0.00 | 75.22 | 10.56 |
| 21 | SA 0126 | 14.94 | 57.64 | 0.00 | 78.73 | 10.16 |
| 61 | SA 0954 | 14.81 | 62.33 | 0.00 | 98.26 | 10.60 |
| 26 | SA 0260 | 14.53 | 61.47 | 0.13 | 86.50 | 10.26 |
| 92 | SA 3125 | 14.49 | 61.78 | 0.09 | 81.48 | 10.16 |
| 78 | SA 2080 | 14.21 | 61.54 | 0.46 | 80.43 | 10.22 |
| Selected accessions for tannin content | ||||||
| 93 | SA 3710 | 14.48 | 57.82 | 11.32 | 101.12 | 10.52 |
| 30 | SA 0390 | 10.41 | 61.05 | 10.41 | 84.50 | 10.88 |
| 80 | SA 2097 | 12.00 | 65.33 | 9.69 | 114.90 | 10.59 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sejake, T.; Shargie, N.; Figlan, S.; Mofokeng, A.; Tsilo, T.J. Phenotypic Variation of Sorghum Accessions for Grain Yield and Quality Traits. Agronomy 2022, 12, 3089. https://doi.org/10.3390/agronomy12123089
Sejake T, Shargie N, Figlan S, Mofokeng A, Tsilo TJ. Phenotypic Variation of Sorghum Accessions for Grain Yield and Quality Traits. Agronomy. 2022; 12(12):3089. https://doi.org/10.3390/agronomy12123089
Chicago/Turabian StyleSejake, Thulo, Nemera Shargie, Sandiswa Figlan, Alina Mofokeng, and Toi John Tsilo. 2022. "Phenotypic Variation of Sorghum Accessions for Grain Yield and Quality Traits" Agronomy 12, no. 12: 3089. https://doi.org/10.3390/agronomy12123089