


Soil Microbial Response to Cover Crop Termination Methods under Two Water Levels
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design and Setup of the Experiment
2.2. Sampling and Measurements
2.3. Statistical Analysis
3. Results
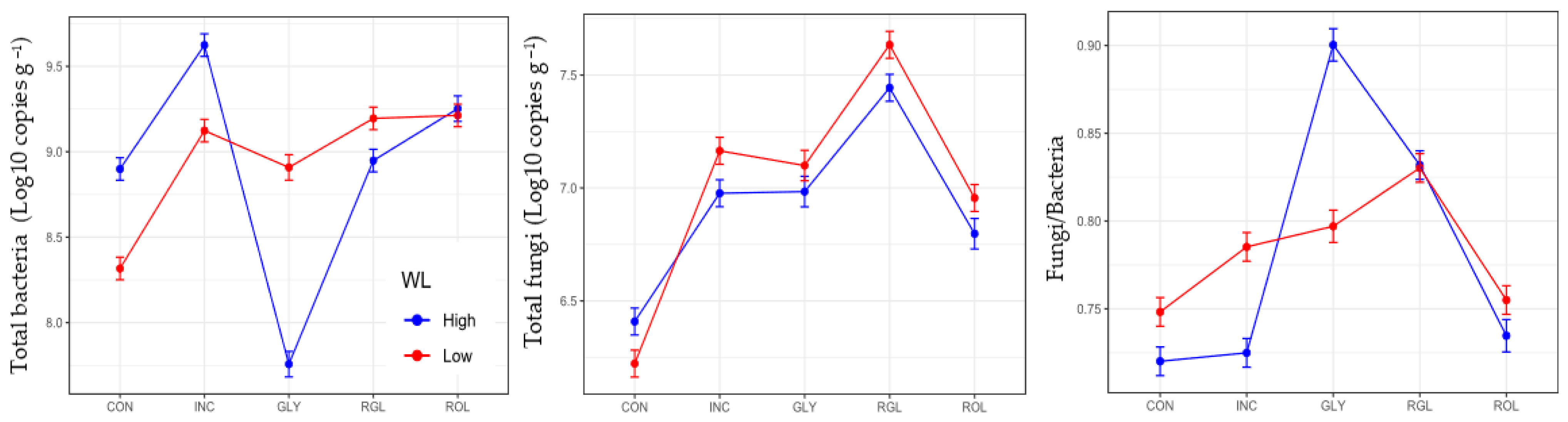
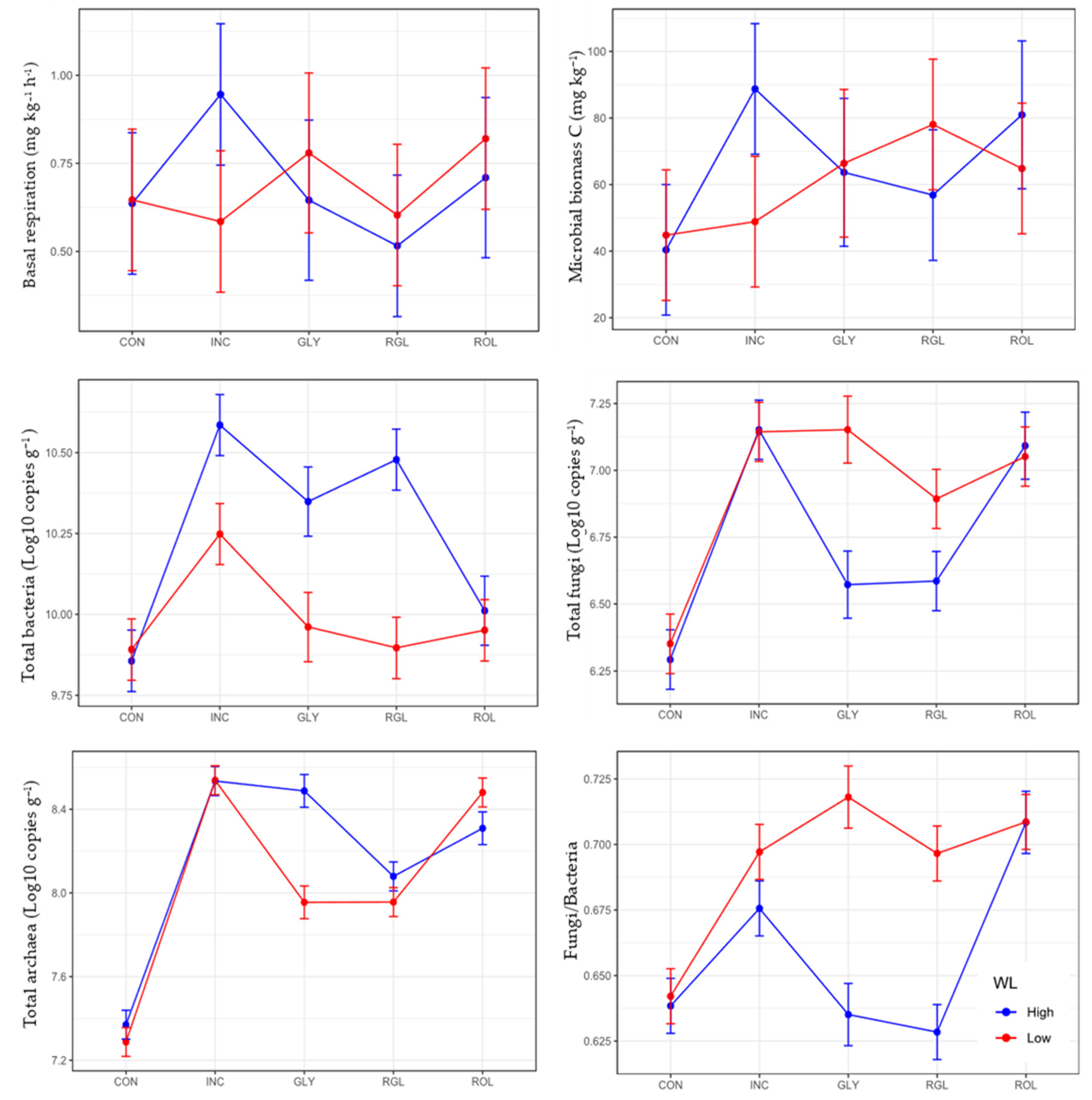
3.1. Soil Microbial Variables at Maize Pre-Emergence and 57 Days after Sowing Maize
3.2. Soil Physicochemical Variables at 57 Days after Sowing Maize
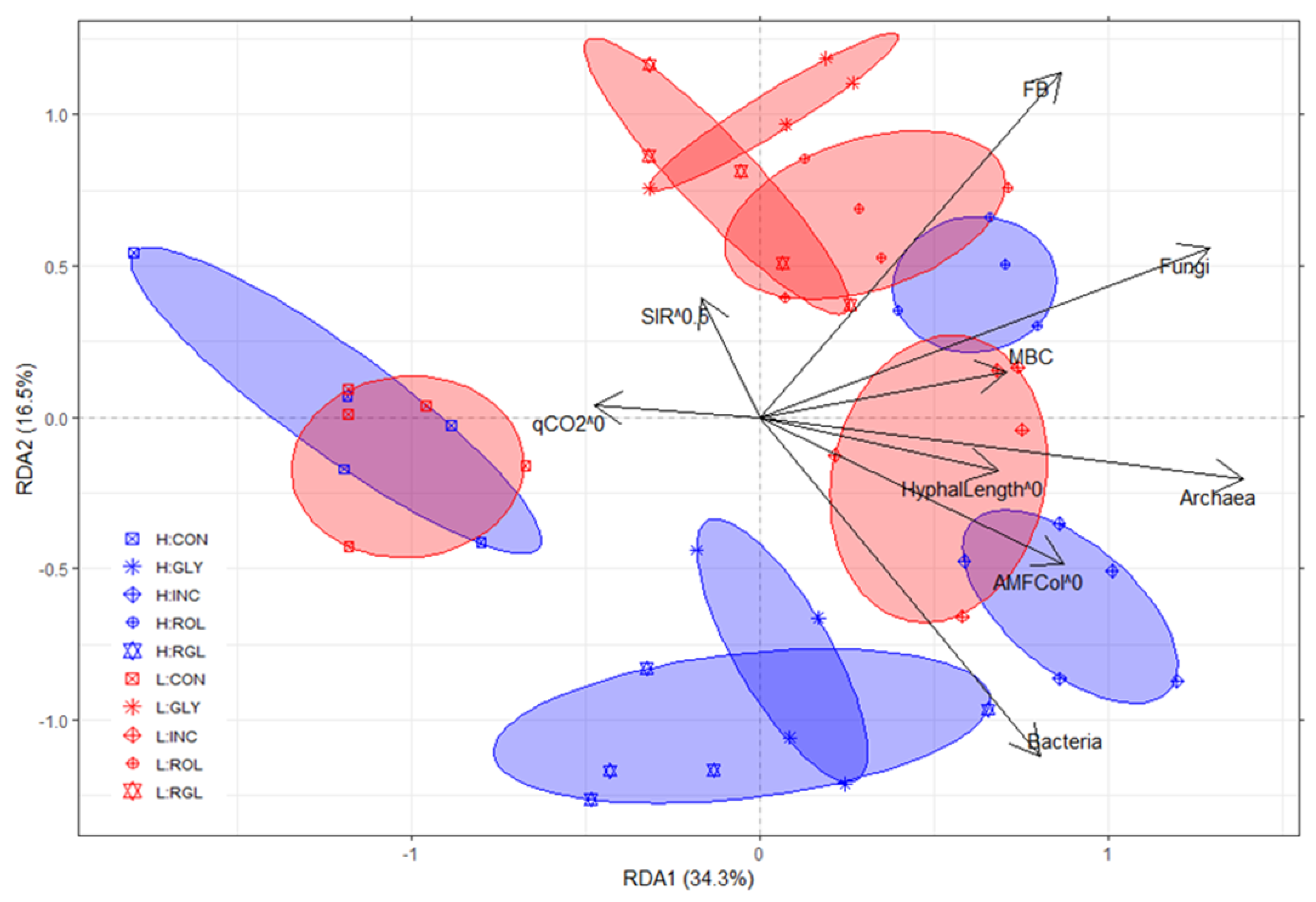
3.3. Redundancy Analysis to Explain Soil Microbial Variables at 57 Days after Sowing Maize from Treatments and Soil Physicochemical Variables
4. Discussion
4.1. Effects of CC Incorporation and Rolling on Soil Microbial Attributes at 57 DAS Maize
4.2. Effects of Glyphosate with and without Rolling on Soil Microbial Attributes at 57 DAS Maize
4.3. Physicochemical Soil Properties and Microbial Attributes as Affected by CC Termination Methods
4.4. Legacy Effects of CC Termination Methods over Time
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schipanski, M.E.; Barbercheck, M.; Douglas, M.R.; Finney, D.M.; Haider, K.; Kaye, J.P.; Kemanian, A.R.; Mortensen, D.A.; Ryan, M.R.; Tooker, J.; et al. A framework for evaluating ecosystem services provided by cover crops in agroecosystems. Agric. Syst. 2014, 125, 12–22. [Google Scholar] [CrossRef]
- Kaye, J.P.; Quemada, M. Using cover crops to mitigate and adapt to climate change. A review. Agron. Sustain. Dev. 2017, 37, 4. [Google Scholar] [CrossRef] [Green Version]
- Abdalla, M.; Hastings, A.; Cheng, K.; Yue, Q.; Chadwick, D.; Espenberg, M.; Truu, J.; Rees, R.M.; Smith, P. A critical review of the impacts of cover crops on nitrogen leaching, net greenhouse gas balance and crop productivity. Glob. Change Biol. 2019, 25, 2530–2543. [Google Scholar] [CrossRef] [Green Version]
- Jian, S.; Li, J.; Wang, G.; Kluber, L.A.; Schadt, C.W.; Liang, J.; Mayes, M.A. Multi-year incubation experiments boost confidence in model projections of long-term soil carbon dynamics. Nat. Commun. 2020, 11, 5864. [Google Scholar] [CrossRef]
- Garba, I.I.; Bell, L.W.; Williams, A. Cover crop legacy impacts on soil water and nitrogen dynamics, and on subsequent crop yields in drylands: A meta-analysis. Agron. Sustain. Dev. 2022, 42, 34. [Google Scholar] [CrossRef]
- Vukicevich, E.; Lowery, T.; Bowen, P.; Úrbez-Torres, J.R.; Hart, M. Cover crops to increase soil microbial diversity and mitigate decline in perennial agriculture. A review. Agron. Sustain. Dev. 2016, 36, 48. [Google Scholar] [CrossRef] [Green Version]
- Adetunji, A.T.; Ncube, B.; Mulidzi, R.; Lewu, F.B. Management impact and benefit of cover crops on soil quality: A review. Soil Tillage Res. 2020, 204, 104717. [Google Scholar] [CrossRef]
- Thapa, V.R.; Ghimire, R.; Acosta-Martínez, V.; Marsalis, M.A.; Schipanski, M.E. Cover crop biomass and species composition affect soil microbial community structure and enzyme activities in semiarid cropping systems. Appl. Soil Ecol. 2021, 157, 103735. [Google Scholar] [CrossRef]
- Kim, N.; Zabaloy, M.C.; Guan, K.; Villamil, M.B. Do cover crops benefit soil microbiome? A meta-analysis of current research. Soil Biol. Biochem. 2020, 142, 107701. [Google Scholar] [CrossRef]
- Muhammad, I.; Wang, J.; Sainju, U.M.; Zhang, S.; Zhao, F.; Khan, A. Cover cropping enhances soil microbial biomass and affects microbial community structure: A meta-analysis. Geoderma 2021, 381, 114696. [Google Scholar] [CrossRef]
- Bowles, T.M.; Jackson, L.E.; Loeher, M.; Cavagnaro, T.R. Ecological intensification and arbuscular mycorrhizas: A meta-analysis of tillage and cover crop effects. J. Appl. Ecol. 2017, 54, 1785–1793. [Google Scholar] [CrossRef] [Green Version]
- García-González, I.; Hontoria, C.; Gabriel, J.L.; Alonso-Ayuso, M.; Quemada, M. Cover crops to mitigate soil degradation and enhance soil functionality in irrigated land. Geoderma 2018, 322, 81–88. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Elsevier Ltd.: NewYork, NY, USA, 2008. [Google Scholar]
- Liang, S.; Grossman, J.; Shi, W. Soil microbial responses to winter legume cover crop management during organic transition. Eur. J. Soil Biol. 2014, 65, 15–22. [Google Scholar] [CrossRef]
- Nevins, C.J.; Nakatsu, C.; Armstrong, S. Characterization of microbial community response to cover crop residue decomposition. Soil Biol. Biochem. 2018, 127, 39–49. [Google Scholar] [CrossRef]
- Schmidt, R.; Gravuer, K.; Bossange, A.V.; Mitchell, J.; Scow, K. Long-term use of cover crops and no-till shift soil microbial community life strategies in agricultural soil. PLoS ONE 2018, 13, e0192953. [Google Scholar] [CrossRef] [PubMed]
- Hontoria, C.; García-González, I.; Quemada, M.; Roldán, A.; Alguacil, M.M. The cover crop determines the AMF community composition in soil and in roots of maize after a ten-year continuous crop rotation. Sci. Total Environ. 2019, 660, 913–922. [Google Scholar] [CrossRef] [PubMed]
- Romdhane, S.; Spor, A.; Busset, H.; Falchetto, L.; Martin, J.; Bizouard, F.; Bru, D.; Breuil, M.; Philippot, L.; Cordeau, S. Cover crop management practices rather than composition of cover crop mixtures affect bacterial communities in no-till agroecosystems. Front. Microbiol. 2019, 10, 1618. [Google Scholar] [CrossRef]
- Jani, A.D.; Grossman, J.M.; Smyth, T.J.; Hu, S. Influence of soil inorganic nitrogen and root diameter size on legume cover crop root decomposition and nitrogen release. Plant Soil 2015, 393, 57–68. [Google Scholar] [CrossRef]
- Daryanto, S.; Fu, B.; Wang, L.; Jacinthe, P.; Zhao, W. Quantitative synthesis on the ecosystem services of cover crops. Earth-Sci. Rev. 2018, 185, 357–373. [Google Scholar] [CrossRef]
- Alonso-Ayuso, M.; Gabriel, J.L.; Hontoria, C.; Ibáñez, M.Á.; Quemada, M. The cover crop termination choice to designing sustainable cropping systems. Eur. J. Agron. 2020, 114, 126000. [Google Scholar] [CrossRef]
- Turmel, M.; Speratti, A.; Baudron, F.; Verhulst, N.; Govaerts, B. Crop residue management and soil health: A systems analysis. Agric. Syst. 2015, 134, 6–16. [Google Scholar] [CrossRef]
- Mbuthia, L.W.; Acosta-Martínez, V.; DeBruyn, J.; Schaeffer, S.; Tyler, D.; Odoi, E.; Mpheshea, M.; Walker, F.; Eash, N. Long term tillage, cover crop, and fertilization effects on microbial community structure, activity: Implications for soil quality. Soil Biol. Biochem. 2015, 89, 24–34. [Google Scholar] [CrossRef]
- Chen, H.; Dai, Z.; Veach, A.M.; Zheng, J.; Xu, J.; Schadt, C.W. Global meta-analyses show that conservation tillage practices promote soil fungal and bacterial biomass. Agric. Ecosyst. Environ. 2020, 293, 106841. [Google Scholar] [CrossRef]
- Hage-Ahmed, K.; Rosner, K.; Steinkellner, S. Arbuscular mycorrhizal fungi and their response to pesticides. Pest Manag. Sci. 2019, 75, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Six, J.; Frey, S.D.; Thiet, R.K.; Batten, K.M. Bacterial and fungal contributions to carbon sequestration in agroecosystems. Soil Sci. Soc. Am. J. 2006, 70, 555–569. [Google Scholar] [CrossRef]
- Van Bruggen, A.H.; He, M.M.; Shin, K.; Mai, V.; Jeong, K.C.; Finckh, M.R.; Morris, J.G., Jr. Environmental and health effects of the herbicide glyphosate. Sci. Total Environ. 2018, 616, 255–268. [Google Scholar] [CrossRef]
- Zabaloy, M.C.; Allegrini, M.; Hernandez Guijarro, K.; Behrends Kraemer, F.; Morrás, H.; Erijman, L. Microbiomes and glyphosate biodegradation in edaphic and aquatic environments: Recent issues and trends. World J. Microbiol. Biotechnol. 2022, 38, 98. [Google Scholar] [CrossRef]
- Van Bruggen, A.H.; Finckh, M.R.; He, M.; Ritsema, C.J.; Harkes, P.; Knuth, D.; Geissen, V. Indirect effects of the herbicide glyphosate on plant, animal and human health through its effects on microbial communities. Front. Environ. Sci. 2021, 9, 464. [Google Scholar] [CrossRef]
- Dennis, P.G.; Kukulies, T.; Forstner, C.; Orton, T.G.; Pattison, A.B. The effects of glyphosate, glufosinate, paraquat and paraquat-diquat on soil microbial activity and bacterial, archaeal and nematode diversity. Sci. Rep. 2018, 8, 2119. [Google Scholar] [CrossRef] [Green Version]
- Druille, M.; Omacini, M.; Golluscio, R.A.; Cabello, M.N. Arbuscular mycorrhizal fungi are directly and indirectly affected by glyphosate application. Appl. Soil Ecol. 2013, 72, 143–149. [Google Scholar] [CrossRef]
- Imparato, V.; Santos, S.S.; Johansen, A.; Geisen, S.; Winding, A. Stimulation of bacteria and protists in rhizosphere of glyphosate-treated barley. Appl. Soil Ecol. 2016, 98, 47–55. [Google Scholar] [CrossRef]
- Navarro-Miró, D.; Blanco-Moreno, J.M.; Ciaccia, C.; Testani, E.; Iocola, I.; Depalo, L.; Burgio, G.; Kristensen, H.L.; Hefner, M.; Tamm, K. The concurrent assessment of agronomic, ecological and environmental variables enables better choice of agroecological service crop termination management. J. Appl. Ecol. 2022, 59, 1026–1037. [Google Scholar] [CrossRef]
- Mirsky, S.B.; Curran, W.S.; Mortensen, D.A.; Ryan, M.R.; Shumway, D.L. Control of cereal rye with a roller/crimper as influenced by cover crop phenology. Agron. J. 2009, 101, 1589–1596. [Google Scholar] [CrossRef] [Green Version]
- Manici, L.M.; Caputo, F.; Nicoletti, F.; Leteo, F.; Campanelli, G. The impact of legume and cereal cover crops on rhizosphere microbial communities of subsequent vegetable crops for contrasting crop decline. Biol. Control 2018, 120, 17–25. [Google Scholar] [CrossRef]
- Navarro-Miró, D.; Blanco-Moreno, J.M.; Ciaccia, C.; Chamorro, L.; Testani, E.; Kristensen, H.L.; Hefner, M.; Tamm, K.; Bender, I.; Jakop, M. Agroecological service crops managed with roller crimper reduce weed density and weed species richness in organic vegetable systems across Europe. Agron. Sustain. Dev. 2019, 39, 55. [Google Scholar] [CrossRef]
- Bloszies, S.A.; Reberg-Horton, S.C.; Heitman, J.L.; Woodley, A.L.; Grossman, J.M.; Hu, S. Legume cover crop type and termination method effects on labile soil carbon and nitrogen and aggregation. Agron. J. 2022, 114, 1817–1832. [Google Scholar] [CrossRef]
- Kornecki, T.S.; Kichler, C.M. Effectiveness of Cover Crop Termination Methods on No-Till Cantaloupe. Agriculture 2022, 12, 66. [Google Scholar] [CrossRef]
- Williams, A.; de Vries, F.T. Plant root exudation under drought: Implications for ecosystem functioning. New Phytol. 2020, 225, 1899–1905. [Google Scholar] [CrossRef] [Green Version]
- Manzoni, S.; Schimel, J.P.; Porporato, A. Responses of soil microbial communities to water stress: Results from a meta-analysis. Ecology 2012, 93, 930–938. [Google Scholar] [CrossRef] [PubMed]
- Schimel, J.P. Life in dry soils: Effects of drought on soil microbial communities and processes. Annu. Rev. Ecol. Evol. Syst. 2018, 49, 409–432. [Google Scholar] [CrossRef]
- Mollaee, M.; Matloob, A.; Mobli, A.; Thompson, M.; Chauhan, B.S. Response of glyphosate-resistant and susceptible biotypes of Echinochloa colona to low doses of glyphosate in different soil moisture conditions. PLoS ONE 2020, 15, e0233428. [Google Scholar] [CrossRef]
- Bento, C.P.M.; Yang, X.; Gort, G.; Xue, S.; van Dam, R.; Zomer, P.; Mol, H.G.J.; Ritsema, C.J.; Geissen, V. Persistence of glyphosate and aminomethylphosphonic acid in loess soil under different combinations of temperature, soil moisture and light/darkness. Sci. Total Environ. 2016, 572, 301–311. [Google Scholar] [CrossRef]
- Köppen, W.; Geiger, R. Handbuch Der Klimatologie; Gebrüder Borntraeger: Berlin, Germany, 1930. [Google Scholar]
- Gabriel, J.L.; García-González, I.; Quemada, M.; Martin-Lammerding, D.; Alonso-Ayuso, M.; Hontoria, C. Cover crops reduce soil resistance to penetration by preserving soil surface water content. Geoderma 2021, 386, 114911. [Google Scholar] [CrossRef]
- Vierheilig, H.; Coughlan, A.P.; Wyss, U.; Piché, Y. Ink and vinegar, a simple staining technique for arbuscular-mycorrhizal fungi. Appl. Environ. Microbiol. 1998, 64, 5004–5007. [Google Scholar] [CrossRef] [Green Version]
- McGonigle, T.P.; Miller, M.H.; Evans, D.G.; Fairchild, G.L.; Swan, J.A. A new method which gives an objective measure of colonization of roots by vesicular—Arbuscular mycorrhizal fungi. New Phytol. 1990, 115, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.W.; Sommers, L.E. Total carbon, organic carbon, and organic matter. In Methods of Soil Analysis: Part 3 Chemical Methods; Soil Science Society of America and American Society of Agronomy: Madison, WI, USA, 1996; Volume 5, pp. 961–1010. [Google Scholar] [CrossRef]
- Bremner, J.M.; Mulvaney, C.S. Total nitrogen. In Methods of Soil Analysis; Page, A.L., Miller, R.H., Keeney, D.R., Eds.; Soil Science Society of America and American Society of Agronomy: Madison, WI, USA, 1982; Part 2; pp. 595–624. [Google Scholar]
- Yakovchenko, V.P.; Sikora, L.J. Modified dichromate method for determining low concentrations of extractable organic carbon in soil. Commun. Soil Sci. Plant Anal. 1998, 29, 421–433. [Google Scholar] [CrossRef]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Alef, K.; Nannipieri, P. Methods in Applied Soil Microbiology and Biochemistry; Academic Press: Cambridge, MA, USA, 1995. [Google Scholar]
- Anderson, J.P.; Domsch, K.H. A physiological method for the quantitative measurement of microbial biomass in soils. Soil Biol. Biochem. 1978, 10, 215–221. [Google Scholar] [CrossRef]
- Anderson, T.; Domsch, A.K. The metabolic quotient for CO2 (qCO2) as a specific activity parameter to assess the effects of environmental conditions, such as pH, on the microbial biomass of forest soils. Soil Biol. Biochem. 1993, 25, 393–395. [Google Scholar] [CrossRef]
- Sparling, G.P. Ratio of microbial biomass carbon to soil organic carbon as a sensitive indicator of changes in soil organic matter. Soil Res. 1992, 30, 195–207. [Google Scholar] [CrossRef]
- García-González, I.; Quemada, M.; Gabriel, J.L.; Hontoria, C. Arbuscular mycorrhizal fungal activity responses to winter cover crops in a sunflower and maize cropping system. Appl. Soil Ecol. 2016, 102, 10–18. [Google Scholar] [CrossRef]
- Jakobsen, I.; Abbott, L.K.; Robson, A.D. External hyphae of vesicular-arbuscular mycorrhizal fungi associated with Trifolium subterraneum L. 1. Spread of hyphae and phosphorus inflow into roots. New Phytol. 1992, 120, 371–380. [Google Scholar] [CrossRef]
- Tennant, D. A test of a modified line intersect method of estimating root length. J. Ecol. 1975, 63, 995–1001. [Google Scholar] [CrossRef]
- RStudio Team. RStudio: Integrated Development Environment for R; RStudio, PBC: Boston, MA, USA, 2022; Available online: http://www.rstudio.com/ (accessed on 10 May 2022).
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P. Vegan: Community Ecology Package, R package version 2.5; Viena, Austria, 2020. Available online: https://www.researchgate.net/publication/346579465_vegan_community_ecology_package_version_25-7_November_2020(accessed on 15 July 2022).
- Wickham, H. Data Analysis, in Anonymous Ggplot2; Springer: Berlin/Heidelberg, Germany, 2016; pp. 189–201. [Google Scholar]
- Dilly, O. Microbial energetics in soils. In Anonymous Microorganisms in Soils: Roles in Genesis and Functions; Springer: Berlin/Heidelberg, Germany, 2005; pp. 123–138. [Google Scholar]
- Sasse, J.; Martinoia, E.; Northen, T. Feed your friends: Do plant exudates shape the root microbiome? Trends Plant Sci. 2018, 23, 25–41. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Yuan, P.; Pozsgai, G.; Chen, P.; Zhu, H.; You, M. The impact of cover crops on the predatory mite Anystis baccarum (Acari, Anystidae) and the leafhopper pest Empoasca onukii (Hemiptera, Cicadellidae) in a tea plantation. Pest Manag. Sci. 2019, 75, 3371–3380. [Google Scholar] [CrossRef] [PubMed]
- Drost, S.M.; Rutgers, M.; Wouterse, M.; De Boer, W.; Bodelier, P.L. Decomposition of mixtures of cover crop residues increases microbial functional diversity. Geoderma 2020, 361, 114060. [Google Scholar] [CrossRef]
- Cordeiro, C.F.D.S.; Rodrigues, D.R.; Rocha, C.H.; Araujo, F.F.; Echer, F.R. Glomalin and microbial activity affected by cover crops and nitrogen management in sandy soil with cotton cultivation. Appl. Soil Ecol. 2021, 167, 104026. [Google Scholar] [CrossRef]
- Valentine, D.L. Adaptations to energy stress dictate the ecology and evolution of the Archaea. Nat. Rev. Microbiol. 2007, 5, 316–323. [Google Scholar] [CrossRef] [PubMed]
- Vincent-Caboud, L.; Casagrande, M.; David, C.; Ryan, M.R.; Silva, E.M.; Peigne, J. Using mulch from cover crops to facilitate organic no-till soybean and maize production. A review. Agron. Sustain. Dev. 2019, 39, 45. [Google Scholar] [CrossRef]
- Elfstrand, S. Impact of Green Manure on Soil Organisms; Swedish University of Agricultural Sciences: Upsala, Sweden, 2007. [Google Scholar]
- Davidson, E.A.; Belk, E.; Boone, R.D. Soil water content and temperature as independent or confounded factors controlling soil respiration in a temperate mixed hardwood forest. Glob. Change Biol. 1998, 4, 217–227. [Google Scholar] [CrossRef] [Green Version]
- Kaurin, A.; Mihelič, R.; Kastelec, D.; Grčman, H.; Bru, D.; Philippot, L.; Suhadolc, M. Resilience of bacteria, archaea, fungi and N-cycling microbial guilds under plough and conservation tillage, to agricultural drought. Soil Biol. Biochem. 2018, 120, 233–245. [Google Scholar] [CrossRef]
- Teasdale, J.R.; Mohler, C.L. Light transmittance, soil temperature, and soil moisture under residue of hairy vetch and rye. Agron. J. 1993, 85, 673–680. [Google Scholar] [CrossRef]
- Sheng, M.; Hamel, C.; Fernandez, M.R. Cropping practices modulate the impact of glyphosate on arbuscular mycorrhizal fungi and rhizosphere bacteria in agroecosystems of the semiarid prairie. Can. J. Microbiol. 2012, 58, 990–1001. [Google Scholar] [CrossRef]
- Zhelezova, A.D.; Manucharova, N.A.; Gorlenko, M.V. Structural and functional characteristics of the prokaryotic community of soddy-podzolic soil influenced by the herbicide glyphosate. Mosc. Univ. Soil Sci. Bull. 2018, 73, 89–94. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, W.; Huang, Y.; Li, J.; Zhong, J.; Zhang, W.; Zou, Y.; Mishra, S.; Bhatt, P.; Chen, S. Insights into the microbial degradation and resistance mechanisms of glyphosate. Environ. Res. 2022, 215, 114153. [Google Scholar] [CrossRef]
- Means, N.E.; Kremer, R.J. Influence of soil moisture on root colonization of glyphosate-treated soybean by Fusarium species. Commun. Soil Sci. Plant Anal. 2007, 38, 1713–1720. [Google Scholar] [CrossRef]
- Jenkins, M.; Locke, M.; Reddy, K.; McChesney, D.S.; Steinriede, R. Glyphosate applications, glyphosate resistant corn, and tillage on nitrification rates and distribution of nitrifying microbial communities. Soil Sci. Soc. Am. J. 2017, 81, 1371–1380. [Google Scholar] [CrossRef] [Green Version]
- Zabaloy, M.C.; Carné, I.; Viassolo, R.; Gómez, M.A.; Gomez, E. Soil ecotoxicity assessment of glyphosate use under field conditions: Microbial activity and community structure of Eubacteria and ammonia-oxidising bacteria. Pest Manag. Sci. 2016, 72, 684–691. [Google Scholar] [CrossRef] [PubMed]
- Ratcliff, A.W.; Busse, M.D.; Shestak, C.J. Changes in microbial community structure following herbicide (glyphosate) additions to forest soils. Appl. Soil Ecol. 2006, 34, 114–124. [Google Scholar] [CrossRef]
- Bruckner, A.; Schmerbauch, A.; Ruess, L.; Heigl, F.; Zaller, J. Foliar Roundup application has minor effects on the compositional and functional diversity of soil microorganisms in a short-term greenhouse experiment. Ecotoxicol. Environ. Saf. 2019, 174, 506–513. [Google Scholar] [CrossRef] [PubMed]
- Kepler, R.M.; Epp Schmidt, D.J.; Yarwood, S.A.; Cavigelli, M.A.; Reddy, K.N.; Duke, S.O.; Bradley, C.A.; Williams, M.M., Jr.; Buyer, J.S.; Maul, J.E. Soil microbial communities in diverse agroecosystems exposed to the herbicide glyphosate. Appl. Environ. Microbiol. 2020, 86, 1744. [Google Scholar] [CrossRef]
- Schlatter, D.C.; Yin, C.; Burke, I.; Hulbert, S.; Paulitz, T. Location, root proximity, and glyphosate-use history modulate the effects of glyphosate on fungal community networks of wheat. Microb. Ecol. 2018, 76, 240–257. [Google Scholar] [CrossRef]
- Vázquez, M.B.; Moreno, M.V.; Amodeo, M.R.; Bianchinotti, M.V. Efecto del glifosato en las comunidades fúngicas del suelo: Estudio a campo. Rev. Argent. Microbiol. 2021, 53, 11–20. [Google Scholar]
- Nguyen, D.B.; Rose, M.T.; Rose, T.J.; Van Zwieten, L. Effect of glyphosate and a commercial formulation on soil functionality assessed by substrate induced respiration and enzyme activity. Eur. J. Soil Biol. 2018, 85, 64–72. [Google Scholar] [CrossRef]
- Schimel, J.P.; Bennett, J. Nitrogen mineralization: Challenges of a changing paradigm. Ecology 2004, 85, 591–602. [Google Scholar] [CrossRef]
- Zaller, J.G.; Heigl, F.; Ruess, L.; Grabmaier, A. Glyphosate herbicide affects belowground interactions between earthworms and symbiotic mycorrhizal fungi in a model ecosystem. Sci. Rep. 2014, 4, 5634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkes, T.I.; Warner, D.J.; Davies, K.G.; Edmonds-Brown, V. Tillage, glyphosate and beneficial arbuscular mycorrhizal fungi: Optimising crop management for plant–fungal symbiosis. Agriculture 2020, 10, 520. [Google Scholar] [CrossRef]
- Helander, M.; Pauna, A.; Saikkonen, K.; Saloniemi, I. Glyphosate residues in soil affect crop plant germination and growth. Sci. Rep. 2019, 9, 19653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, D.B.; Rose, M.T.; Rose, T.J.; Morris, S.G.; Van Zwieten, L. Impact of glyphosate on soil microbial biomass and respiration: A meta-analysis. Soil Biol. Biochem. 2016, 92, 50–57. [Google Scholar] [CrossRef]
- Alan, M.Z.; Fabiano, A.P.; Jlio, C.A.N.; Leandro, P.P.; Francisco, A.N.; Fabrcio, R.A. Microbiological attributes in a Latosol in glyphosate application under water deficit conditions. Afr. J. Agric. Res. 2014, 9, 2495–2505. [Google Scholar] [CrossRef]
- Buyer, J.S.; Teasdale, J.R.; Roberts, D.P.; Zasada, I.A.; Maul, J.E. Factors affecting soil microbial community structure in tomato cropping systems. Soil Biol. Biochem. 2010, 42, 831–841. [Google Scholar] [CrossRef]
- Wolmarans, K.; Swart, W.J. Influence of glyphosate, other herbicides and genetically modified herbicide resistant crops on soil microbiota: A review. S. Afr. J. Plant Soil 2014, 31, 177–186. Available online: https://hdl.handle.net/10520/EJC163053 (accessed on 15 July 2022). [CrossRef]
- Zobiole, L.; Kremer, R.J.; Oliveira, R.S., Jr.; Constantin, J. Glyphosate affects micro-organisms in rhizospheres of glyphosate-resistant soybeans. J. Appl. Microbiol. 2011, 110, 118–127. [Google Scholar] [CrossRef] [PubMed]
- White, T.; Bruns, T.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Gnes for Phylogenetics; Elsevier: Amsterdam, The Netherlands, 1990; pp. 315–322. [Google Scholar]
- López-Gutiérrez, J.; Henry, S.; Hallet, S.; Martin-Laurent, F.; Catroux, G.; Philippot, L. Quantification of a novel group of nitrate-reducing bacteria in the environment by real-time PCR. J. Microbiol. Methods 2004, 57, 399–407. [Google Scholar] [CrossRef] [PubMed]
- Ochsenreiter, T.; Selezi, D.; Quaiser, A.; Bonch-Osmolovskaya, L.; Schleper, C. Diversity and abundance of Crenarchaeota in terrestrial habitats studied by 16S RNA surveys and real time PCR. Environ. Microbiol. 2003, 5, 787–797. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Factors | Levels | Hyphal Length | Total Bacteria | Total Fungi | Fungi/Bacteria | Total Archaea |
|---|---|---|---|---|---|---|
| cm g−1 | Log10 Copies g−1 | Log10 Copies g−1 | Log10 Copies g−1 | |||
| TM | CON | 16.78 a | 8.60 | 6.31 | 0.73 | 6.77 a |
| INC | 26.75 b | 9.37 | 7.07 | 0.75 | 7.54 b | |
| GLY | 23.87 ab | 8.35 | 7.04 | 0.84 | 7.60 bc | |
| RGL | 19.70 ab | 9.07 | 7.53 | 0.83 | 7.74 c | |
| ROL | 22.75 ab | 9.22 | 6.86 | 0.74 | 7.76 c | |
| WL | High | 26.70 b | 8.89 | 6.91 | 0.78 | 7.49 a |
| Low | 17.87 a | 8.95 | 7.01 | 0.78 | 7.47 a | |
| TM | ** | *** | *** | *** | *** | |
| WL | *** | * | *** | ns | ns | |
| TM x WL | ns | *** | *** | *** | ns | |
| Factors | Levels | AMF Col. | Hyphal Length | Basal Resp. | SIR | MBC | qCO2 | qMIC | Total Bacteria | Total Fungi | Fungi/ Bacteria | Total Archaea |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| % | cm g−1 | mg C-CO2 kg−1h−1 | mg C-CO2 kg−1h−1 | mg kg−1 | mg C-CO2 mg MBC h−1 | mg MBC g−1 C | Log10 Copies g−1 | Log10 Copies g−1 | Log10 Copies g−1 | |||
| TM | CON | 10.8 a | 18.13 a | 0.64 a | 1.52 a | 42.61 a | 0.019 b | 8.78 a | 9.87 a | 6.32 a | 0.64 a | 7.32 a |
| INC | 26.6 b | 30.51 b | 0.76 a | 0.93 a | 68.8 ab | 0.011 ab | 13.46 a | 10.41 c | 7.14 c | 0.68 c | 8.53 e | |
| GLY | 16.0 ab | 19.14 a | 0.71 a | 1.67 a | 65.0 ab | 0.011 ab | 12.98 a | 10.15 b | 6.86 b | 0.67 bc | 8.21 c | |
| RGL | 16.6 ab | 16.86 a | 0.55 a | 1.32 a | 67.4 ab | 0.008 a | 13.61 a | 10.18 b | 6.73 b | 0.66 b | 8.01 b | |
| ROL | 22.1 b | 24.27ab | 0.76 a | 1.53 a | 72.8 b | 0.012 ab | 14.83 a | 9.98 a | 7.10 c | 0.70 d | 8.40 d | |
| WL | High | 21.5 b | 24.01 a | 0.69 a | 1.43 a | 66.1 a | 0.012 a | 13.13 a | 10.26 b | 6.73 a | 0.65 a | 8.15 b |
| Low | 15.3 a | 19.76 a | 0.68 a | 1.36 a | 60.5 a | 0.012 a | 12.34 a | 10.00 a | 6.91 b | 0.69 b | 8.04 a | |
| TM | *** | ** | ns | ns | * | * | ns | *** | *** | *** | *** | |
| WL | * | ns | ns | ns | ns | ns | ns | *** | *** | *** | *** | |
| TM x WL | ns | ns | ns | ns | * | ns | * | *** | *** | *** | *** | |
| Factors | Levels | pH1:2.5 | EC1:2.5 | TOC | DOC | %DOC/ TOC | C/N | C/P | N/P | Moisture | T | PR | Db |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| µS cm−1 | g kg−1 | mg kg−1 | %v | °C | kg cm−2 | g cm−3 | |||||||
| TM | CON | 8.35 a | 244 c | 4.90 a | 58.1 a | 1.19 a | 8.14 a | 0.16 a | 0.02 a | 5.12 ab | 24.7 a | 1.82 ab | 1.30 a |
| INC | 8.45 a | 188 ab | 5.09 a | 56.3 a | 1.11 a | 9.24 a | 0.18 ab | 0.03 a | 4.67 a | 24.2 a | 2.13 b | 1.32 a | |
| GLY | 8.48 a | 191 abc | 5.08 a | 54.6 a | 1.08 a | 7.28 a | 0.21 b | 0.02 a | 6.70 ab | 24.7 a | 1.39 a | 1.37 a | |
| RGL | 8.41 a | 217 bc | 5.05 a | 54.6 a | 1.10 a | 8.04 a | 0.17 ab | 0.02 a | 7.70 b | 23.6 a | 1.37 a | 1.33 a | |
| ROL | 8.43 a | 143 a | 4.99 a | 54.0 a | 1.10 a | 6.73 a | 0.18 ab | 0.02 a | 5.57 ab | 23.8 a | 1.75 ab | 1.35 a | |
| WL | High | 8.46 a | 190 a | 5.07 a | 55.8 a | 1.10 a | 8.00 a | 0.19 b | 0.02 a | 8.48 b | 24.0 a | 1.60 a | 1.35 a |
| Low | 8.39 a | 204 a | 4.98 a | 55.3 a | 1.12 a | 7.80 a | 0.16 a | 0.02 a | 3.06 a | 24.5 a | 1.78 b | 1.32 a | |
| TM | ns | *** | ns | ns | ns | ns | * | ns | * | ns | *** | ns | |
| WL | ns | ns | ns | ns | ns | ns | ** | ns | *** | ns | * | ns | |
| TM x WL | ns | ns | ns | ns | ns | * | ns | * | * | ns | ns | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Centurión, N.; Ulcuango, K.; Navas, M.; Mariscal-Sancho, I.; Ibáñez, M.A.; Moliner, A.; Hontoria, C. Soil Microbial Response to Cover Crop Termination Methods under Two Water Levels. Agronomy 2022, 12, 3002. https://doi.org/10.3390/agronomy12123002
Centurión N, Ulcuango K, Navas M, Mariscal-Sancho I, Ibáñez MA, Moliner A, Hontoria C. Soil Microbial Response to Cover Crop Termination Methods under Two Water Levels. Agronomy. 2022; 12(12):3002. https://doi.org/10.3390/agronomy12123002
Chicago/Turabian StyleCenturión, Nelly, Kelly Ulcuango, Mariela Navas, Ignacio Mariscal-Sancho, Miguel A. Ibáñez, Ana Moliner, and Chiquinquirá Hontoria. 2022. "Soil Microbial Response to Cover Crop Termination Methods under Two Water Levels" Agronomy 12, no. 12: 3002. https://doi.org/10.3390/agronomy12123002