
Potassium Application Increases Cotton (Gossypium hirsutum L.) Fiber Length by Improving K+/Na+ Homeostasis and Potassium Transport Capacity in the Boll-Leaf System under Moderate Salinity


 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material, Experimental Site, and Soil Properties
2.2. Experimental Design
2.3. Sampling and Processing
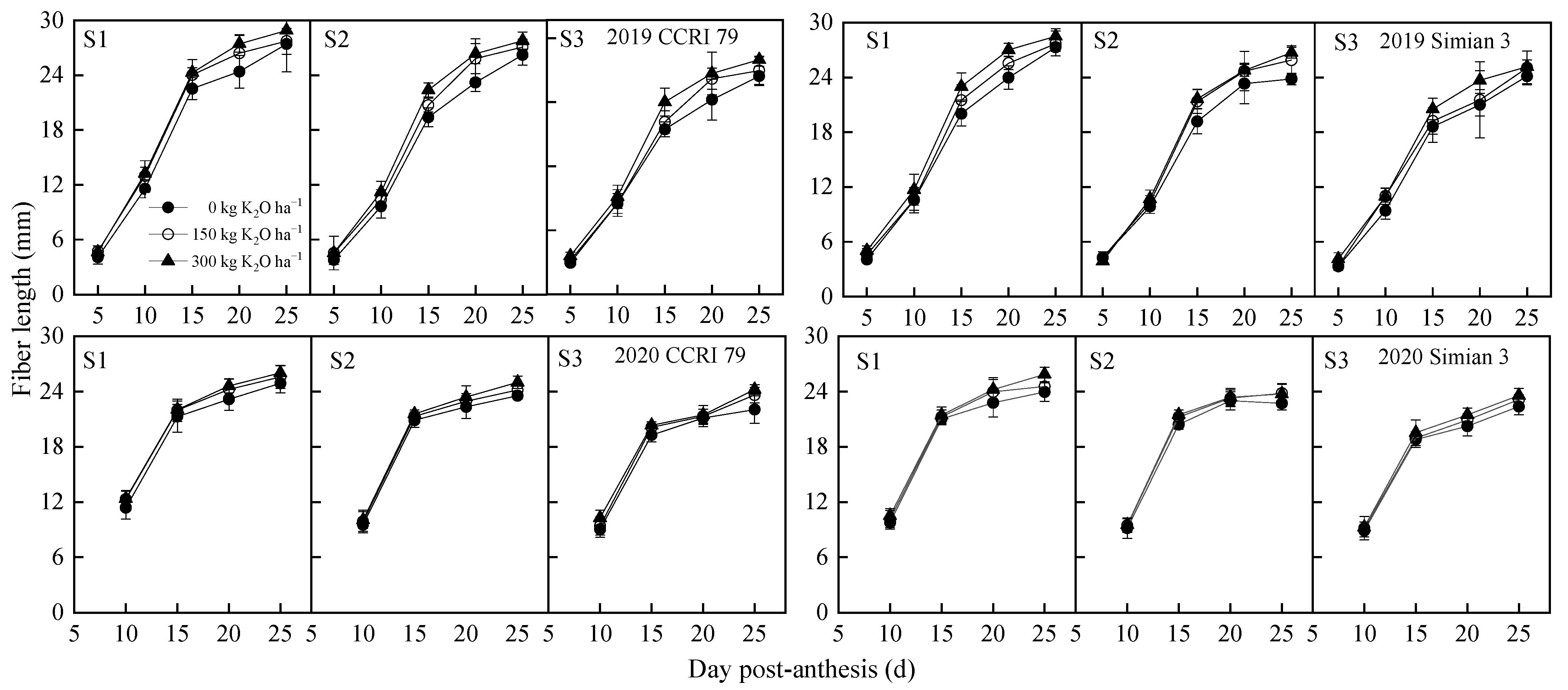
2.4. Cotton Fiber Length
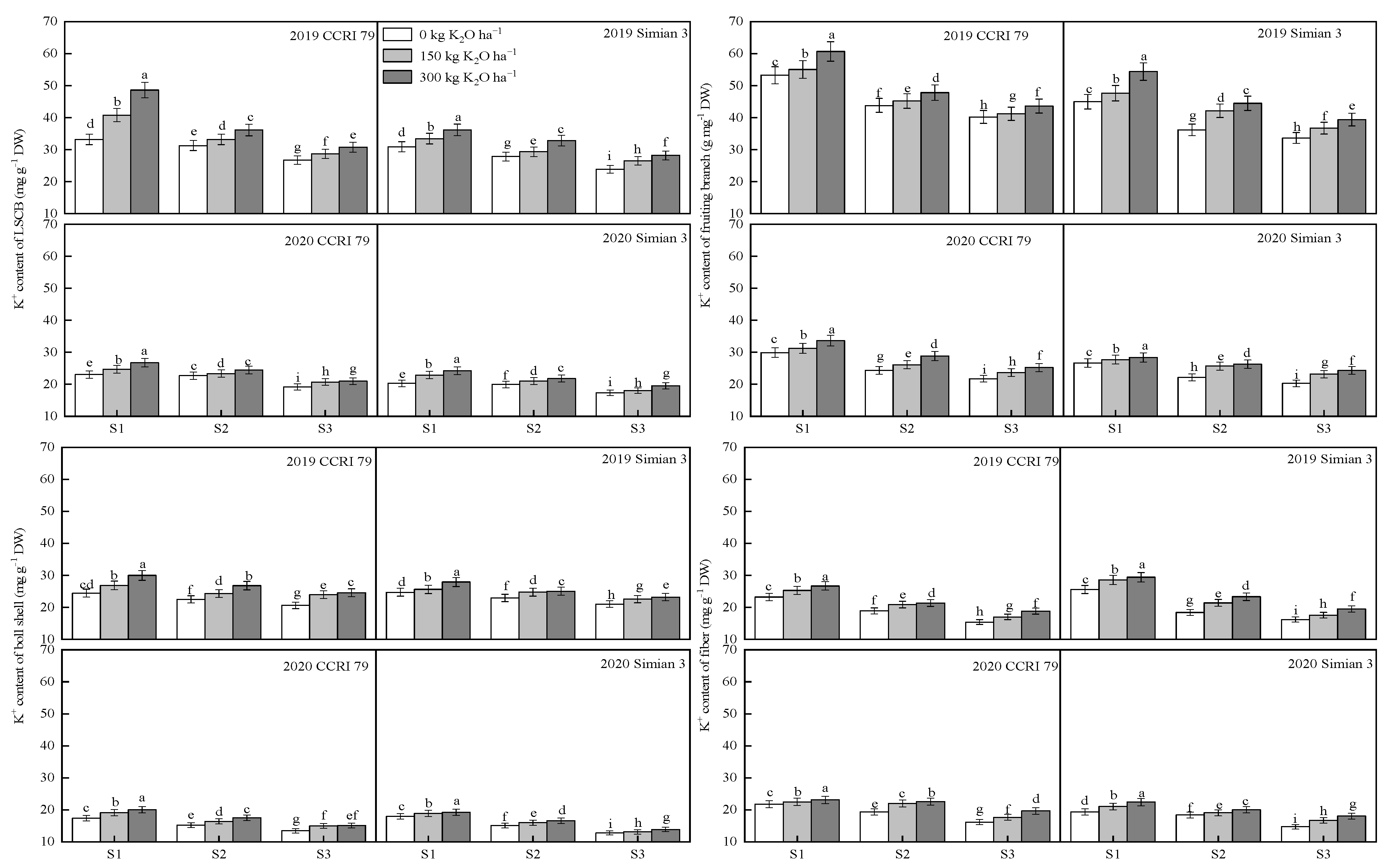
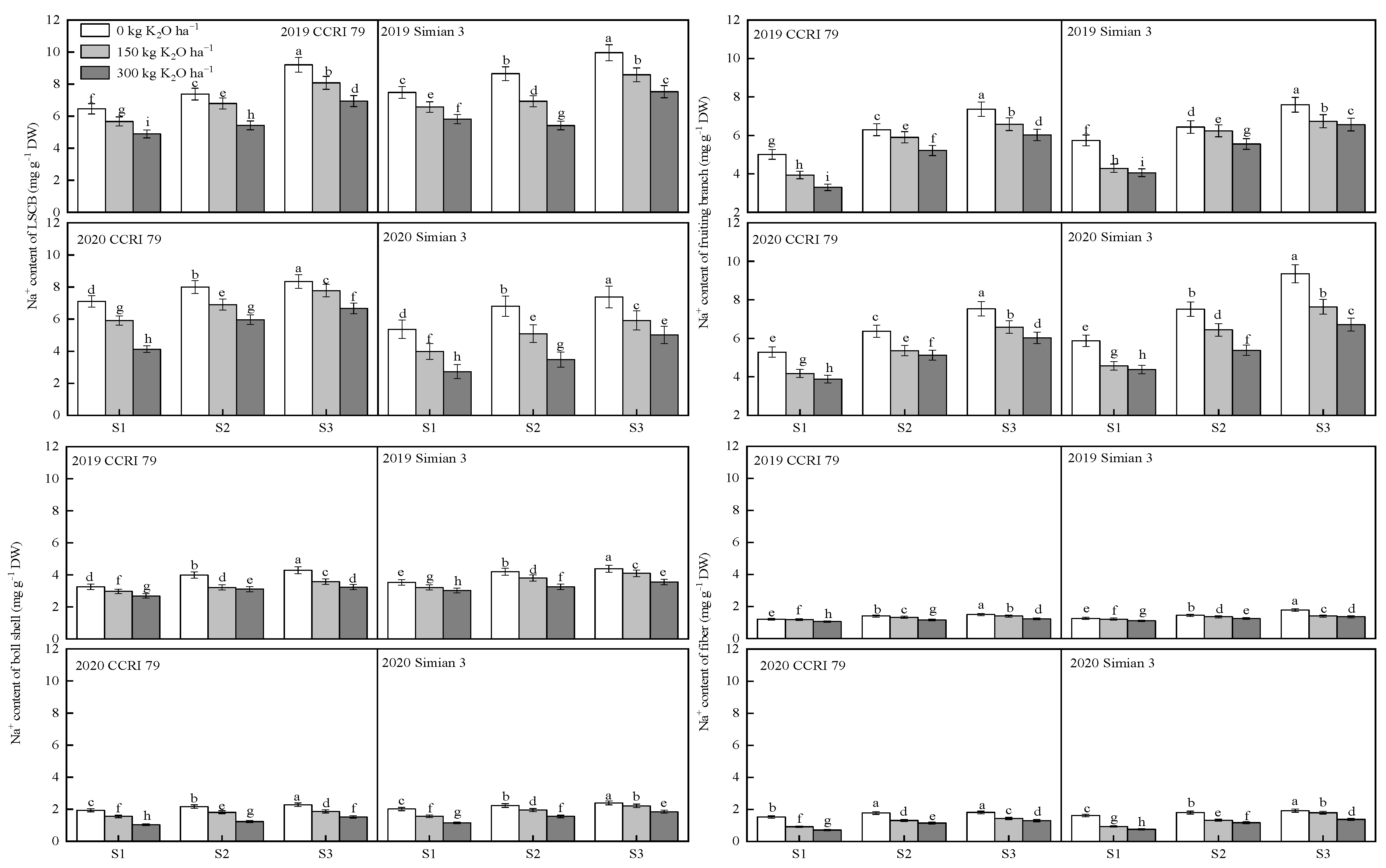
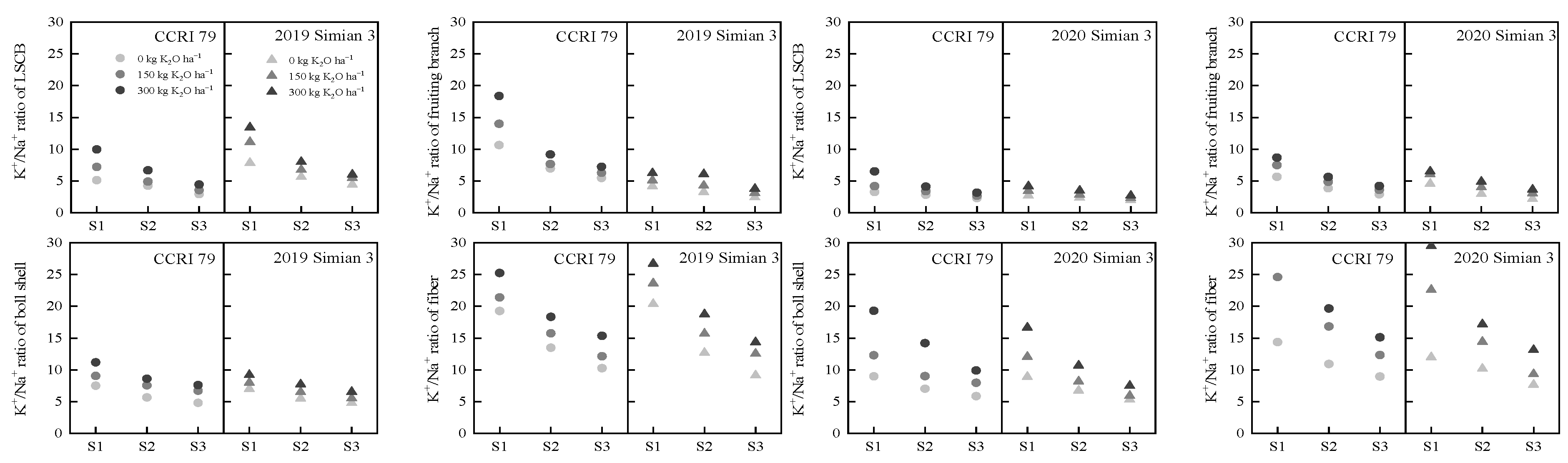
2.5. Concentrations of K+ and Na+ in the Boll-Leaf System
2.6. Statistical Analysis
3. Results
3.1. Effect of the Application of K on Fiber Length and Its Alleviation Coefficient under Salt Stress
3.2. Effect of K Application on the K+ Content in Various Organs of the Boll-Leaf System under Salt Stress
3.3. Effect of K Application on Na+ Content in Boll-Leaf System Organs under Salt Stress
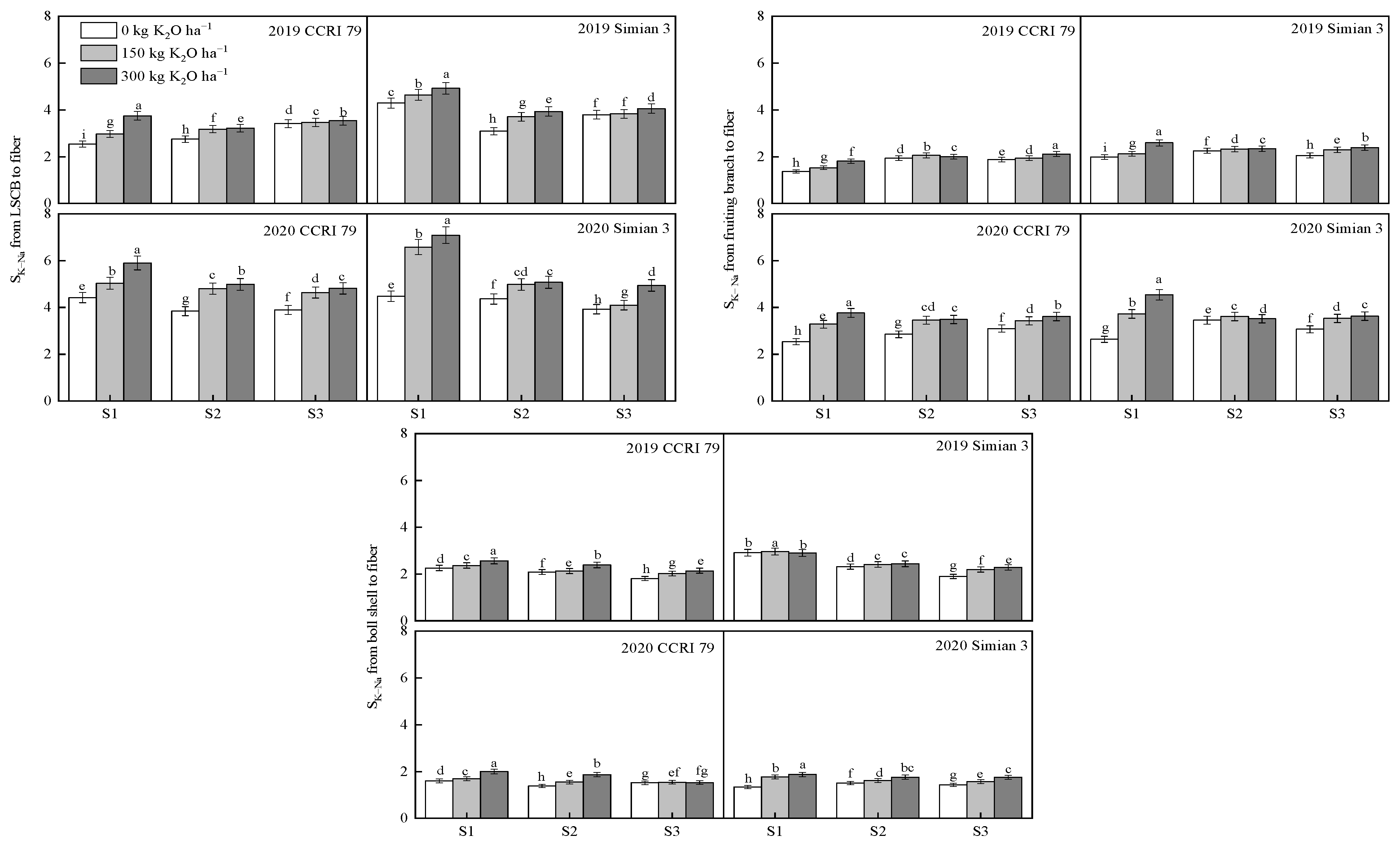
3.4. Effect of K Application on the K+/Na+ Ratio in the Boll-Leaf System under Salt Stress
3.5. Correlations of Cotton Fiber Length with K+ and Na+ Contents in the Boll-Leaf System
3.6. The K Transport Capacity in the Boll-Leaf System under Salt Stress
4. Discussion
4.1. Alleviation Effects of K Application on Cotton Fiber Length under Salt Stress
4.2. Effects of K Application on K+ and Na+ Contents and the K+/Na+ Ratio in the Boll-Leaf System under Salt Stress
4.3. Effect of K Application on K+ Transport Capacity in the Boll-Leaf System under Salt Stress
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bailey-Serres, J.; Parker, J.E.; Ainsworth, E.A.; Oldroyd, G.E.D.; Schroeder, J.I. Genetic strategies for improving crop yields. Nature 2019, 575, 109–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jamil, A.; Riaz, S.; Ashraf, M.; Foolad, M.R. Gene expression profiling of plants under salt stress. Crit. Rev. Plant Sci. 2011, 30, 435–458. [Google Scholar] [CrossRef]
- Wei, Y.; Xu, Y.C.; Lu, P.; Wang, X.X.; Li, Z.Q.; Cai, X.Y.; Zhou, Z.L.; Wang, Y.H.; Zhang, Z.M.; Lin, Z.X.; et al. Salt stress responsiveness of a wild cotton species (Gossypium klotzschianum) based on transcriptomic analysis. PLoS ONE 2017, 12, e0178313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Z.P.; Duan, X.B.; Luo, L.; Dai, S.J.; Ding, Z.J.; Xia, G.M. How plant hormones mediate salt stress responses. Trends Plant Sci. 2020, 25, 1117–1130. [Google Scholar] [CrossRef] [PubMed]
- Munns, R.; Gilliham, M. Salinity tolerance of crops—What is the cost. New Phytol. 2015, 208, 668–673. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.J.; Huang, Z.J.; Li, M.Q.; Hou, Z.N. Growth, ionic homeostasis, and physiological responses of cotton under different salt and alkali stresses. Sci. Rep. 2020, 10, 21844. [Google Scholar] [CrossRef]
- Su, Y.; Guo, A.H.; Huang, Y.; Wang, Y.M.; Hua, J.P. GhCIPK6a increases salt tolerance in transgenic upland cotton by involving in ROS scavenging and MAPK signaling pathways. BMC Plant Biol. 2020, 20, 421. [Google Scholar] [CrossRef]
- Xu, P.; Guo, Q.; Meng, S.; Zhang, X.G.; Xu, Z.Z.; Guo, W.Z.; Shen, X.L. Genome-wide association analysis reveals genetic variations and candidate genes associated with salt tolerance related traits in Gossypium hirsutum. BMC Genom. 2021, 22, 26. [Google Scholar] [CrossRef]
- Ashraf, J.; Zuo, D.; Wang, Q.; Malik, W.; Zhang, Y.; Abid, M.A.; Song, G. Recent insights into cotton functional genomics: Progress and future perspectives. Plant Biotechnol. J. 2018, 16, 699–713. [Google Scholar] [CrossRef] [Green Version]
- Pandey, G.K.; Mahiwal, S. Potassium in abiotic stress. In Role of Potassium in Plants; Pandey, G.K., Mahiwal, S., Eds.; Springer: Cham, Switzerland, 2020; pp. 45–49. [Google Scholar] [CrossRef]
- Peng, J.; Zhang, L.; Liu, J.; Luo, J.; Zhao, X.; Dong, H.; Meng, Y. Effects of Soil Salinity on Sucrose Metabolism in Cotton Fiber. PLoS ONE 2016, 11, e0156398. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Guo, Y. Elucidating the molecular mechanisms mediating plant salt-stress responses. New Phytol. 2017, 217, 523–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, X.L.; Zhou, D.Y.; Guo, P.; Zhang, H.; Dong, J.L.; Ren, J.Y.; Jiang, C.J.; Zhong, C.; Zhao, X.H.; Yu, H.Q. External potassium mediates the response and tolerance to salt stress in peanut at the flowering and needling stages. Photosynthetica 2020, 58, 1141–1149. [Google Scholar] [CrossRef]
- Abbasi, G.H.; Akhtar, J.; Ahmad, R.; Jamil, M.; Anwar-ul-Haq, M.; Ali, S.; Ijaz, M. Potassium application mitigates salt stress differentially at different growth stages in tolerant and sensitive maize hybrids. Plant Growth Regul. 2015, 76, 111–125. [Google Scholar] [CrossRef]
- Ashraf, M.; Ahmad, S. Genetic effects for yield components and fiber characteristics in upland cotton (Gossypium hirsutum L.) cultivated under salinized (NaCl) conditions. Agronomie 2000, 20, 917–926. [Google Scholar] [CrossRef] [Green Version]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [Green Version]
- Aldesuquy, H.S.; Baka, Z.A.; Mickky, B.M. Role of kinetin and spermine in the reversal of seawater stress-induced alteration in growth vigor, water relations and nucleic acids of wheat plants. Phyton-Ann. Rei Bot. 2014, 54, 251–274. [Google Scholar] [CrossRef]
- Jiang, Y.R.; Lv, Y.J.; Zhu, S.J. Advance in studies of the mechanism of salt tolerance and controlling of salt damage in upland cotton. Cotton Sci. 2006, 18, 248–254. [Google Scholar]
- Sanyal, S.K.; Rajasheker, G.; Kishor, P.K.; Kumar, S.A.; Kumari, P.H.; Saritha, K.V.; Rathnagiri, P.; Pandey, G.K. Role of protein phosphatases in signaling, potassium transport, and abiotic stress responses. In Protein Phosphatases and Stress Management in Plants; Pandey, G.K., Ed.; Springer: Cham, Switzerland, 2020; pp. 203–232. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J. Potassium control of plant functions: Ecological and agricultural implications. Plants 2021, 10, 419. [Google Scholar] [CrossRef]
- Isayenkov, S.V.; Maathuis, F.J.M. Plant salinity stress: Many unanswered questions remain. Front. Plant Sci. 2019, 10, 80. [Google Scholar] [CrossRef]
- Parveen; Anwar-Ul-Haq, M.; Aziz, T.; Aziz, O.; Maqsood, L. Potassium induces carbohydrates accumulation by enhancing morpho-physiological and biochemical attributes in soybean under salinity. Arch. Agron. Soil Sci. 2020, 67, 946–959. [Google Scholar] [CrossRef]
- Guo, K.; Tu, L.L.; He, Y.H.; Deng, J.W.; Wang, M.J.; Huang, H.; Zhang, X. Interaction between calcium and potassium modulates elongation rate in cotton fiber cells. J. Exp. Bot. 2017, 68, 5161–5175. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.S.; Hu, W.; Zhao, W.Q.; Chen, B.L.; Wang, Y.H.; Zhou, Z.G.; Meng, Y.L. Fruiting branch K+ level affects cotton fiber elongation through osmoregulation. Front. Plant Sci. 2016, 7, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.H.; Fang, J.P.; Wu, X.B.; Dong, L.Y. Na+/K+ balance and transport regulatory mechanisms in weedy and cultivated rice (Oryza sativa L.) under salt stress. BMC Plant Biol. 2018, 18, 375. [Google Scholar] [CrossRef] [Green Version]
- Li, X.Y.; Mu, C.S.; Lin, J.X.; Wang, Y.; Li, X.J. Effect of alkaline potassium and sodium salts on growth, photosynthesis, ions absorption and solutes synthesis of wheat seedlings. Exp. Agric. 2014, 50, 144–157. [Google Scholar] [CrossRef]
- Rodrigues, J.F.; Junqueira, G.; Gonçalves, C.S.; Carneiro, J.D.; Pinheiro, A.C.; Nunes, C.A. Elaboration of garlic and salt spice with reduced sodium intake. An. Acad. Bras. Ciênc. 2014, 86, 2065–2075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teakle, N.L.; Flowers, T.J.; Real, D.; Colmer, T.D. Lotus tenuis tolerates the interactive effects of salinity and waterlogging by ’excluding’ Na+ and Cl− from the xylem. J. Exp. Bot. 2007, 58, 2169–2180. [Google Scholar] [CrossRef]
- Sun, J.B.; Sun, G.Y.; Liu, X.D.; Hu, Y.B.; Zhao, L.S. Effects of salt stress on mulberry seedlings growth, leaf water status, and ion distribution in various organs. J. Appl. Ecol. 2009, 20, 543–548. (In Chinese) [Google Scholar] [CrossRef]
- Wahid, A.; Bukhari, S.; Rasul, E. Inter-specific differences in cotton for nutrient partitioning from subtending leaves to reproductive parts at various developmental stages, consequences for fruit growth and yield. Biol. Plant 2003, 47, 379–385. [Google Scholar] [CrossRef]
- Samal, D.; Kovar, J.L.; Steingrobe, B.; Sadana, U.S.; Bhadoria, P.S.; Claassen, N. Potassium uptake efficiency and dynamics in the rhizosphere of maize (Zea mays L.), wheat (Triticum aestivum L.), and sugar beet (Beta vulgaris L.) evaluated with a mechanistic model. Plant Soil 2010, 332, 105–121. [Google Scholar] [CrossRef]
- Wang, N.; Hua, H.B.; Eneji, A.E.; Li, Z.H.; Duan, L.S.; Tian, X.L. Genotypic variations in photosynthetic and physiological adjustment to potassium deficiency in cotton (Gossypium hirsutum). J. Photochem. Photobiol. B 2012, 110, 1–8. [Google Scholar] [CrossRef]
- Zhou, G.S.; Lin, Y.; Tong, C.; An, L.L.; Liu, G.J. Effects of nitrogen application amount on growth characteristics, boll development and lint yield of high quality cotton. Agric. Sci. Technol. 2011, 12, 1667–1670. [Google Scholar] [CrossRef]
- Liu, R.X.; Zhou, Z.G.; Guo, W.Q.; Chen, B.L.; Osterbuis, D.M. Effects of N fertilization on root development and activity of water-stressed cotton (Gossypium hirsutum L.) plants. Agric. Water Manag. 2008, 95, 1261–1270. [Google Scholar] [CrossRef]
- Rizwan, Z.; Haoran, D.; Muhammad, A.; Zhao, W.Q.; Wang, Y.H.; Zhou, Z.G. Potassium fertilizer improves drought stress alleviation potential in cotton by enhancing photosynthesis and carbohydrate metabolism. Environ. Exp. Bot. 2017, 137, 73–83. [Google Scholar] [CrossRef]
- Gipson, J.R.; Ray, L.L. Fiber Elongation Rates in Five Varieties of Cotton (Gossypium hirsutum L.) as Influenced by Night Temperature1. Crop Sci. 1969, 9, 339–341. [Google Scholar] [CrossRef]
- Yang, X.Y.; Li, C.L.; Zhang, Q.; Liu, Z.G.; Geng, J.B.; Zhang, M. Effects of polymer-coated potassium chloride on cotton yield, leaf senescence and soil potassium. Field Crops Res. 2017, 212, 145–152. [Google Scholar] [CrossRef]
- Flowers, T.J.; Yeo, A.R. Ion relation of salt tolerance. In Solute Transport in Plant Cells and Tissues; Baker, D.A., Hall, J.L., Eds.; John Wiley and Sons, Inc.: New York, NY, USA, 1988; pp. 392–416. [Google Scholar] [CrossRef]
- Pettigrew, W.T.; Meredith, W.R.; Young, L.D. Potassium fertilization effects on cotton lint yield, yield components, and reniform nematode populations. Agron. J. 2005, 97, 1245–1251. [Google Scholar] [CrossRef] [Green Version]
- Adhikari, B.; Dhungana, S.K.; Kim, I.D.; Shin, D.H. Effect of foliar application of potassium fertilizers on soybean plants under salinity stress. J. Saudi Soc. Agric. Sci. 2020, 19, 261–269. [Google Scholar] [CrossRef]
- Queirós, F.; Fontes, N.; Silva, P.; Almeida, D.; Maeshima, M.; Gerós, H.; Fidalgo, F. Activity of tonoplast proton pumps and Na+/H+ exchange in potato cell cultures is modulated by salt. J. Exp. Bot. 2009, 60, 1363–1374. [Google Scholar] [CrossRef]
- Armengaud, P.; Sulpice, R.; Miller, A.J.; Stitt, M.; Amtmann, A.; Gibon, Y. Multilevel analysis of primary metabolism provides new insights into the role of potassium nutrition for glycolysis and nitrogen assimilation in Arabidopsis roots. Plant Physiol. 2009, 150, 772–785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, T.; Blumwald, E. Developing salt-tolerant crop plants: Challenges and opportunities. Trends Plant Sci. 2005, 10, 615–620. [Google Scholar] [CrossRef]
- Lauter, D.; Meiri, A.; Shuali, M. Isoosmotic regulation of cotton and peanut at saline concentrations of K and Na. Plant Physiol. 1988, 87, 911–916. [Google Scholar] [CrossRef] [PubMed]
- Leonova, T.G.; Goncharova, E.A.; Khodorenko, A.V.; Babakov, A.V. Characteristics of salt-tolerant and salt-susceptible cultivars of barley. Russ. J. Plant Physiol. 2005, 52, 774–778. [Google Scholar] [CrossRef]
- Li, N.Y.; Chen, S.L.; Zhou, X.Y.; Li, C.Y.; Shao, J.; Wang, R.G.; Fritz, E.; Huettermann, A.; Polle, A. Effect of NaCl on photosynthesis, salt accumulation and ion compartmentation in two mangrove species, Kandelia candel and Bruguiera gymnorhiza. Aquat. Bot. 2008, 88, 303–310. [Google Scholar] [CrossRef]
- Ju, F.Y.; Pang, J.L.; Huo, Y.Y.; Zhu, J.J.; Yu, K.; Sun, L.Y.; Loca, D.A.; Hu, W.; Zhou, Z.G.; Wang, S.S.; et al. Potassium application alleviates the negative effects of salt stress on cotton (Gossypium hirsutum L.) yield by improving the ionic homeostasis, photosynthetic capacity and carbohydrate metabolism of the leaf subtending the cotton boll. Field Crops Res. 2021, 272, 108288. [Google Scholar] [CrossRef]
- Ashraf, M.; Rahmatullah; Ahmad, R.; Bhatti, A.S.; Afzal, M.; Sarwar, A.; Maqsood, M.A.; Kanwal, S. Amelioration of salt stress in sugarcane (Saccharum officinarum L.) by supplying potassium and silicon in hydroponics. Pedosphere 2010, 20, 153–162. [Google Scholar] [CrossRef]
- Zhao, D.; Oosterhuis, D.M. Dynamics of non-structural carbohydrates in developing leaves, bracts and floral buds of cotton. Environ. Exp. Bot. 2000, 43, 185–195. [Google Scholar] [CrossRef]
- Liu, Z.X.; Zhang, H.X.; Yang, X.Y.; Liu, T.; Di, W.B. Growth, and cationic absorption, transportation and allocation of Elaeagnus angustifolia seedlings under NaCl stress. Acta Ecol. Sin. 2014, 34, 326–336. [Google Scholar] [CrossRef] [Green Version]
- Wu, F.Z.; Zhu, X.W.; Hu, R.F.; Wang, H.X.; Chen, Y.M. Effects of NaCl stress on growth ion uptake, transportation and distribution of two blueberry (Vaccinium corymbosum) cultivars seedlings. Sci. Silvae Sin. 2017, 53, 40–49. [Google Scholar] [CrossRef]
- Ran, X.; Huang, X.; Wang, X.; Liang, H.; Wang, Y.; Li, J.; Huo, Z.; Liu, B.; Ma, C. Ion absorption, distribution and salt tolerance threshold of three willow species under salt stress. Front. Plant Sci. 2022, 13, 969896. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Salinity Level | EC (dS m−1) | NaCl (g kg−1) | pH | ρb (g cm−3) | TN (g kg−1) | Ah-N (mg kg−1) | Av-P (mg kg−1) | Av-K (mg kg−1) |
|---|---|---|---|---|---|---|---|---|---|
| 2019 | S1 | 1.7 | 2.3 | 8.2 | 1.5 | 1.6 | 74.5 | 34.4 | 116.9 |
| S2 | 6.4 | 3.8 | 8.2 | 1.5 | 1.6 | 75.6 | 34.8 | 129.5 | |
| S3 | 11.8 | 4.8 | 8.8 | 1.5 | 1.6 | 73.2 | 33.4 | 134.0 | |
| 2020 | S1 | 1.8 | 1.8 | 8.1 | 1.5 | 1.7 | 79.3 | 37.0 | 160.0 |
| S2 | 6.9 | 3.6 | 8.1 | 1.5 | 1.7 | 77.5 | 37.5 | 136.4 | |
| S3 | 10.6 | 4.3 | 8.4 | 1.4 | 1.7 | 82.5 | 39.0 | 170.2 |
| Salinity Level | K Rate (kg K2O ha−1) | K+/Na+ of 2019 | K+/Na+ of 2020 | Average of K+/Na+ |
|---|---|---|---|---|
| S1 | 0 | 1:13 | 1:8 | 1:10 |
| 150 | 1:9 | 1:5 | 1:7 | |
| 300 | 1:7 | 1:4 | 1:5 | |
| S2 | 0 | 1:20 | 1:18 | 1:19 |
| 150 | 1:13 | 1:12 | 1:13 | |
| 300 | 1:10 | 1:9 | 1:10 | |
| S3 | 0 | 1:24 | 1:17 | 1:20 |
| 150 | 1:17 | 1:13 | 1:15 | |
| 300 | 1:13 | 1:10 | 1:11 |
| Cultivar | Salinity Level | K Rate (kg K2O ha−1) | 2019 (mm) | 2020 (mm) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Upper | Middle | Lower | Whole | Upper | Middle | Lower | Whole | |||
| CCRI 79 | S1 | 0 | 29.1 b | 29.2 c | 28.2 bc | 28.8 b | 29.5 b | 29.2 bc | 28.3 bc | 29.0 b |
| 150 | 29.9 a | 29.8 a | 28.8 ab | 29.5 a | 29.9 a | 30.0 ab | 29.0 ab | 29.6 a | ||
| 300 | 29.9 a | 30.0 a | 29.0 a | 29.6 a | 30.2 a | 30.3 a | 29.6 a | 30.0 a | ||
| Mean | 29.6 | 29.7 | 28.7 | 29.3 | 29.9 | 29.8 | 29.0 | 29.6 | ||
| S2 | 0 | 28.8 bc | 28.8 d | 27.8 cd | 28.5 c | 28.9 b | 28.8 bc | 27.6 d | 28.4 c | |
| 150 | 29.1 b | 29.4 b | 28.1 c | 28.9 b | 29.5 b | 29.3 ab | 28.5 abc | 29.1 b | ||
| 300 | 29.2 ab | 29.6 ab | 28.2 bc | 29.0 b | 29.7 a | 29.5 ab | 28.9 ab | 29.4 a | ||
| Mean | 29.0 | 29.3 | 28.0 | 28.8 | 29.4 | 29.2 | 28.3 | 29.0 | ||
| S3 | 0 | 27.6 d | 27.7 f | 26.5 f | 27.3 e | 28.3 b | 28.4 c | 27.2 d | 28.0 d | |
| 150 | 28.2 c | 28.3 e | 27.4 e | 28.0 d | 28.7 b | 28.8 ab | 27.5 c | 28.3 b | ||
| 300 | 28.4 c | 28.9 d | 27.6 d | 28.3 cd | 28.9 b | 29.0 ab | 27.7 bc | 28.5 b | ||
| Mean | 28.1 | 28.3 | 27.2 | 27.8 | 28.6 | 28.7 | 27.5 | 28.3 | ||
| Alleviation coefficient | S2 | 150 | 100.0 | 150.0 | 75.0 | 133.3 | 100.0 | 125.0 | 128.6 | 117.6 |
| 300 | 133.3 | 200.0 | 100.0 | 166.7 | 133.3 | 175.0 | 185.7 | 164.7 | ||
| S3 | 150 | 40.0 | 40.0 | 41.2 | 40.0 | 33.3 | 50.0 | 27.3 | 35.5 | |
| 300 | 53.3 | 80.0 | 47.1 | 60.0 | 50.0 | 75.0 | 45.5 | 54.8 | ||
| Simian 3 | S1 | 0 | 28.9 bc | 28.4 bc | 28.3 cd | 28.5 b | 29.2 bc | 28.6 cde | 28.0 ab | 28.6 de |
| 150 | 29.6 ab | 29.7 a | 29.3 ab | 29.5 a | 29.5 abc | 29.2 abcd | 29.4 a | 29.4 bc | ||
| 300 | 29.7 a | 29.8 a | 29.6 a | 29.7 a | 30.4 a | 29.4 a | 29.5 a | 29.8 a | ||
| Mean | 29.4 | 29.3 | 29.1 | 29.3 | 29.7 | 29.1 | 29.0 | 29.2 | ||
| S2 | 0 | 28.0 d | 27.6 de | 27.7 de | 27.8 c | 28.6 c | 28.1 de | 27.4 bc | 28.0 e | |
| 150 | 28.7 c | 28.4 bc | 28.2 c | 28.4 b | 29.0 abc | 28.6 abc | 28.1 ab | 28.6 c | ||
| 300 | 28.9 bc | 28.8 b | 28.5 bc | 28.7 b | 29.3 ab | 28.9 ab | 28.4 a | 28.9 ab | ||
| Mean | 28.5 | 28.3 | 28.1 | 28.3 | 29.0 | 28.5 | 28.0 | 28.5 | ||
| S3 | 0 | 27.1 e | 27.2 e | 26.6 f | 27.0 d | 28.4 c | 27.6 e | 26.2 c | 27.4 f | |
| 150 | 27.6 d | 27.7 cd | 27.3 e | 27.5 c | 28.6 c | 28.0 bcde | 26.7 ab | 27.8 de | ||
| 300 | 27.8 d | 28.0 cd | 27.7 de | 27.8 c | 28.7 c | 28.1 ab | 27.0 ab | 27.9 cd | ||
| Mean | 27.5 | 27.6 | 27.2 | 27.4 | 28.6 | 27.9 | 26.6 | 27.7 | ||
| Alleviation coefficient | S2 | 150 | 77.8 | 100.0 | 83.3 | 87.0 | 66.7 | 100.0 | 116.7 | 94.1 |
| 300 | 100.0 | 150.0 | 133.3 | 126.1 | 166.7 | 160.0 | 166.7 | 147.1 | ||
| S3 | 150 | 27.8 | 41.7 | 41.2 | 36.2 | 25.0 | 40.0 | 27.8 | 30.6 | |
| 300 | 38.9 | 66.7 | 64.7 | 55.3 | 37.5 | 50.0 | 44.4 | 44.4 | ||
| Salinity(S) | ** | ** | ||||||||
| K rate(K) | ** | ** | ||||||||
| Cultivar(C) | ** | ** | ||||||||
| S × K | ** | ** | ||||||||
| S × C | ** | ** | ||||||||
| K × C | ** | ** | ||||||||
| Year | Parameter | LSCB | Fruiting Branch | Boll Shell | Fiber |
|---|---|---|---|---|---|
| 2019 | K+ | 0.8820 ** | 0.8943 ** | 0.9076 ** | 0.9726 ** |
| Na+ | −0.9138 ** | −0.9195 ** | −0.8896 ** | −0.8414 ** | |
| K+/Na+ | 0.9089 ** | 0.8895 ** | 0.9221 ** | 0.8599 ** | |
| 2020 | K+ | 0.9239 ** | 0.8666 ** | 0.9226 ** | 0.9436 ** |
| Na+ | −0.7864 ** | −0.9183 ** | −0.7408 ** | −0.7512 ** | |
| K+/Na+ | 0.7776 ** | 0.8666 ** | 0.7752 ** | 0.7718 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, J.; Sun, L.; Ju, F.; Wang, Z.; Xiong, C.; Yu, H.; Yu, K.; Huo, Y.; Khattak, W.A.; Hu, W.; et al. Potassium Application Increases Cotton (Gossypium hirsutum L.) Fiber Length by Improving K+/Na+ Homeostasis and Potassium Transport Capacity in the Boll-Leaf System under Moderate Salinity. Agronomy 2022, 12, 2962. https://doi.org/10.3390/agronomy12122962
Zhu J, Sun L, Ju F, Wang Z, Xiong C, Yu H, Yu K, Huo Y, Khattak WA, Hu W, et al. Potassium Application Increases Cotton (Gossypium hirsutum L.) Fiber Length by Improving K+/Na+ Homeostasis and Potassium Transport Capacity in the Boll-Leaf System under Moderate Salinity. Agronomy. 2022; 12(12):2962. https://doi.org/10.3390/agronomy12122962
Chicago/Turabian StyleZhu, Junjun, Liyuan Sun, Feiyan Ju, Zhuo Wang, Cai Xiong, Huilian Yu, Kai Yu, Yuyang Huo, Wajid Ali Khattak, Wei Hu, and et al. 2022. "Potassium Application Increases Cotton (Gossypium hirsutum L.) Fiber Length by Improving K+/Na+ Homeostasis and Potassium Transport Capacity in the Boll-Leaf System under Moderate Salinity" Agronomy 12, no. 12: 2962. https://doi.org/10.3390/agronomy12122962