1. Introduction
Production of wheat (
Triticum aestivum L.), a major staple crop, is endangered by the increasing severity and incidence of yellow rust (YR) disease. YR is an epidemic disease caused by the fungus
Puccinia striiformis (
Pst.), infecting both spring as well as winter wheat [
1]. YR is one of the most diverse, destructive, and widespread diseases decimating wheat productivity [
2]. Around 88% of wheat productions are vulnerable to YR [
3]. According to an estimate, every year there is a loss of 5–6 million tonnes of wheat due to YR, at an estimated cost of USD 979 million [
3]. Grain-filling is directly impacted by YR, especially if the disease occurs after anthesis. This is because the affected photosynthetic cells are less effective in intercepting light and using radiation, which lowers yields [
4]. Efficient dispersal, i.e., the ability of
Pst. urediospores to travel long distances, even across continents, has resulted in its global spread [
5].
Severe yellow rust outbreaks have been reported, ranging in intensity from two percent to complete crop loss [
6]. The latest entry to the list of countries affected by the YR epidemic is Zimbabwe [
7]. However, the epidemic occurs more frequently (2 or 3 years of every 5 years) in Central and South Asia, China, West Africa, Australasia, the UK, the USA, and South America [
8]. Besides the higher dispersability of the YR pathogen, another reason for its widespread presence is the high mutation rate of
Pst. resulting in its higher adaptability as explained by its complicated life cycle [
9]. The sexual stage in the pathogen life cycle hosted by
Berberis vulgaris plays a key role in generating novel genetic combinations, which results in potential virulent
Pst. isolates that may lead to epidemics and swift alterations in wheat resistance [
10].
With five distinct sporulation stages,
Pst. has a highly complicated life cycle. The pathogen lifecycle is split into asexual stages, utilizing wheat (the primary host), and sexual cycle utilizing
B. vulgaris (alternate host;
Figure 1) [
11]. However, alternate host
Berberis spp. colonized by
Pst. are seldom seen in the wild [
12,
13]. Most of the wheat-growing regions around the world lack the opportunity for
Pst. to undergo complete genetic recombination (due to the absence of an alternate host) with the exception of the Himalayan region, i.e., China and Pakistan [
14,
15,
16,
17]. The seasons of the development of
B. vulgaris and wheat growing in the region are also simultaneous [
13].
In recent years,
Pst. populations have acclimated to hot climates, grown more aggressive, and have become genetically diverse. Control of the
Pst. and protection against YR primarily involves the use of fungicides and endogenous plant resistance. Fungicides can be used to reduce stripe rust, but their expense and potential environmental impacts raise serious questions [
18]. Growing resistant cultivars effectively and sustainably should control YR [
19,
20]. Utilization of YR-resistant genotypes to lessen YR damage is the most cost-effective and environmentally friendly strategy because it protects grain production and lowers the demand for fungicides [
21]. Widespread interest in the YR resistance discovery has led to the identification of a large number of YR resistance (Yr) and susceptibility genes [
22].
Sustainable protection against YR through the employment of resistant wheat cultivars has been complicated by the swift adaptability of the
Pst. [
23,
24]. However, using resistance genes individually to prevent yellow rust is no longer effective due to the appearance of
Pst. pathotypes adapted to a wide range of host resistance (Yr) genes. Genetic resistance conferred by a single gene may be prone to break down as a result of the pathogen’s virulence evolution [
25]. Since the 1950s, there have been five boom and bust cycles, where otherwise resistant wheat cultivars were devastated by newly adapted
Pst. Races [
26]. Breeding methods are now focusing on genomic and molecular approaches for developing durable YR-resistant wheat cultivars using multigenic approaches [
27].
Breeding programs use two types of stripe rust resistance genes, i.e., all-stage resistance (ASR) and adult plant resistance (APR) genes depending on the resistance conferred at different growth stages [
28]. The majority of the yellow rust ASR genes discovered are limited by specificity to particular races resistance genes [
29]. Race specificity makes germplasm carrying the ASR genes vulnerable and this kind of resistance may be broken down within a short span of time [
30]. Therefore, growing cultivars expressing race-specific ASR genes over large geographical areas exert selection pressure on the
Pst. resulting in the fast evolution of new virulent races [
31]. The ASR genes mostly encode for proteins containing nucleotide-binding sites with leucine-rich repeat (NBS-LRR). The NBS-LRR allows specific identification of the avirulence (Avr) proteins by respective sets of R gene products inside the host cells [
32]. Examples of Yr genes carrying NBS-LRR include YrSP, Yr5, and Yr10, etc. [
33].
In contrast to the ASR, the APR expresses in late plant growth stages, resulting in slow rusting phenotypes [
11]. The APR resistance may also be regulated by temperature, a phenomenon known as high-temperature adult plant (HTAP) resistance. APR is usually partial, and may or may not be sufficient to resist the
Pst. invasion [
20,
34]. However, the key positive dimension of the APR is that this resistance type is race non-specific and is considered more durable and effective against many
Pst. pathotypes. For example, Yr18 has remained durable and effective against YR for the longest duration [
35,
36]. To boost resistance level and durability, there is a need to combine APR with ASR [
37].
Screening for the Yr genes and associated quantitative traits loci (QTL) requires the use of molecular approaches such as genome-wide association studies (GWAS) and more frequently used associated DNA markers. DNA markers among others include microsatellite or simple sequence repeats (SSR), random amplified polymorphic DNA (RAPD), restriction fragments length polymorphism (RFLP), and single nucleotide polymorphism (SNPs), etc. [
38,
39]. DNA markers are needed to construct genetic maps, which may be utilized to find the location of Yr genes or Yr QTL [
40]. By means of marker-assisted selection (MAS), these Yr QTL may be introduced into the cultivated wheat genotypes to develop resistant varieties [
41]. Furthermore, markers that are frequently associated with important genes may serve as a starting point for map-based transgenic germplasm development [
42].
Allelic variations at different microsatellite markers were used to screen genetic differences among susceptible, moderately resistant, and resistant wheat genotypes as indicated by their low, moderate, and high relative resistance index (RRI), respectively. We used different analytical tools to identify novel microsatellite alleles associated with YR resistance. The associated microsatellite markers (of the total 19 markers) were further mined for their proximity with the known Yr genes/QTLs (Yr18, Yr26, Yr5, Ye15, Yr62) in the GrainGenes database [
43]. The recombinant inbred lines (RILs) developed using the diverse parents were further screened based on their RRI. Data from the study are envisioned to form the basis for sustainable YR-resistant breeding in wheat.
2. Materials and Methods
This study was conducted at two locations, i.e., The Center of Excellence, Cereal Crops Research Institute (CCRI), Pirsabak, Nowshera, and shuttle wheat breeding station (SWBS) Kaghan, Khyber Pakhtunkhwa Pakistan during the years 2018–2019, 2019–2020, and 2020–2021. CCRI is situated 288 m (945 ft) AMSL at the left bank of Kabul River, 3 km east of Nowshera, at 32° N latitude and 74° E longitude. Nowshera has a warm to hot, semi-arid, and sub-tropical climate with a mean annual rainfall of about 364 mm. It is a hot spot for yellow rust development due to its conducive environment. At 34°50′ N and 73°31′ E, Kaghan is a mountain valley with an elevation of 650 m (2134 ft) above sea level. Due to its favorable summer climate for wheat, this shuttle breeding site was built to promote wheat reproduction. With an average humidity of 59%, summertime temperatures typically range from 20 °C to 26 °C.
2.1. Germplasm Source, Matting Design and Sowing Layout
A diverse panel of wheat genotypes including fourteen parents (9 female lines and 5 male testers) and 45 recombinant inbred lines (RILs), resulted from lines × testers crossings, were evaluated under YR-stress conditions. Parental germplasm was procured from three wheat research groups, i.e., (i) two advance lines from State Key Laboratory, department of crop breeding and genetics, Anhui Agricultural University Hefei, China, (ii) five lines from International Maize and Wheat Improvement Center (CIMMYT) Mexico, and (iii) seven from Wheat Breeding Section of Cereal Crops Research Institute Pirsabak Pakistan [
44].
Hybridization (cross combinations of lines × testers) of breeding material (
Figure 2) was carried out at the wheat breeding section, CCRI Pirsabak, during the winter cropping season 2018–2019. Filial generation advancement was done with two generations per year at SWBS Kaghan (June to October), and CCRI Pirsabak (November to May) from 2019 to 2021. The initial selection of the RILs and the subsequent generation advancement were based on screening for YR-resistant phenotypes. The RRI data were ultimately collected on the F6 generation of the RILs. The selected 45 F6 RILs (selected resistant plants from the F5 population) along the respective parents (9 lines and 5 testers) were sown at CCRI Pirsabak in the 1st week of November 2021 using randomized complete block design, with three replications. Each plot had 4 rows of 5 m length with 0.25 m row-to-row distance and 0.6 m plot-to-plot distance. The experimental plots were fenced by a highly susceptible wheat genotype (Morocco) to encourage YR disease development. Standard agronomic practices were carried out as per the requirement of the wheat crop. The fertilizer was applied in split doses (half at sowing and the remaining half at the early boot stage).
2.2. Inoculation of Breeding Germplasm
Experimental material was inoculated at the vegetative growth (booting) stage with a fresh inoculum of the dominant Pakistani pathotype (
Pst. 574232) [
27]. The YR-inoculum was obtained from the Crop Diseases Research Institute, National Agricultural Research Centre (NARC), Islamabad. The uredospore’s suspension was prepared using 0.1 g of YR inoculum in 1 L of distilled water and added a few drops of Tween-20 for efficient germination of spores and fungal hyphae penetration. At sunset, each experimental plot was inoculated uniformly with the suspension by using a turbo-air sprayer.
2.3. YR-Disease Severity and Host Response
YR disease reaction and severity in the host plant were determined based on the percent flag leaf area affected. Peterson et al.’s [
45] method based on the visual determination of the proportion (1 to 100%) of leaf area infected by yellow rust was utilized to assess disease severity using a modified Cobb scale. The response of the host was detected by its reaction against the disease. The data on host responses and disease value were recorded and presented as, “I” denotes fully immune type by having no yellow rust infection, “R” denotes resistant by having observable with no YR pustules and necrosis on leaf, “MR” denotes moderately resistant by showing small and tiny pustules with little necrosis on leaf, “MS” denotes moderately susceptible by having an intermediate level of pustules and zero necrosis but having observable chlorosis on leaf, and “S” denotes fully susceptible with fresh and bulky pustules with necrosis and chlorotic area on leaf surface.
2.4. Disease Assessment and Host Phenotyping
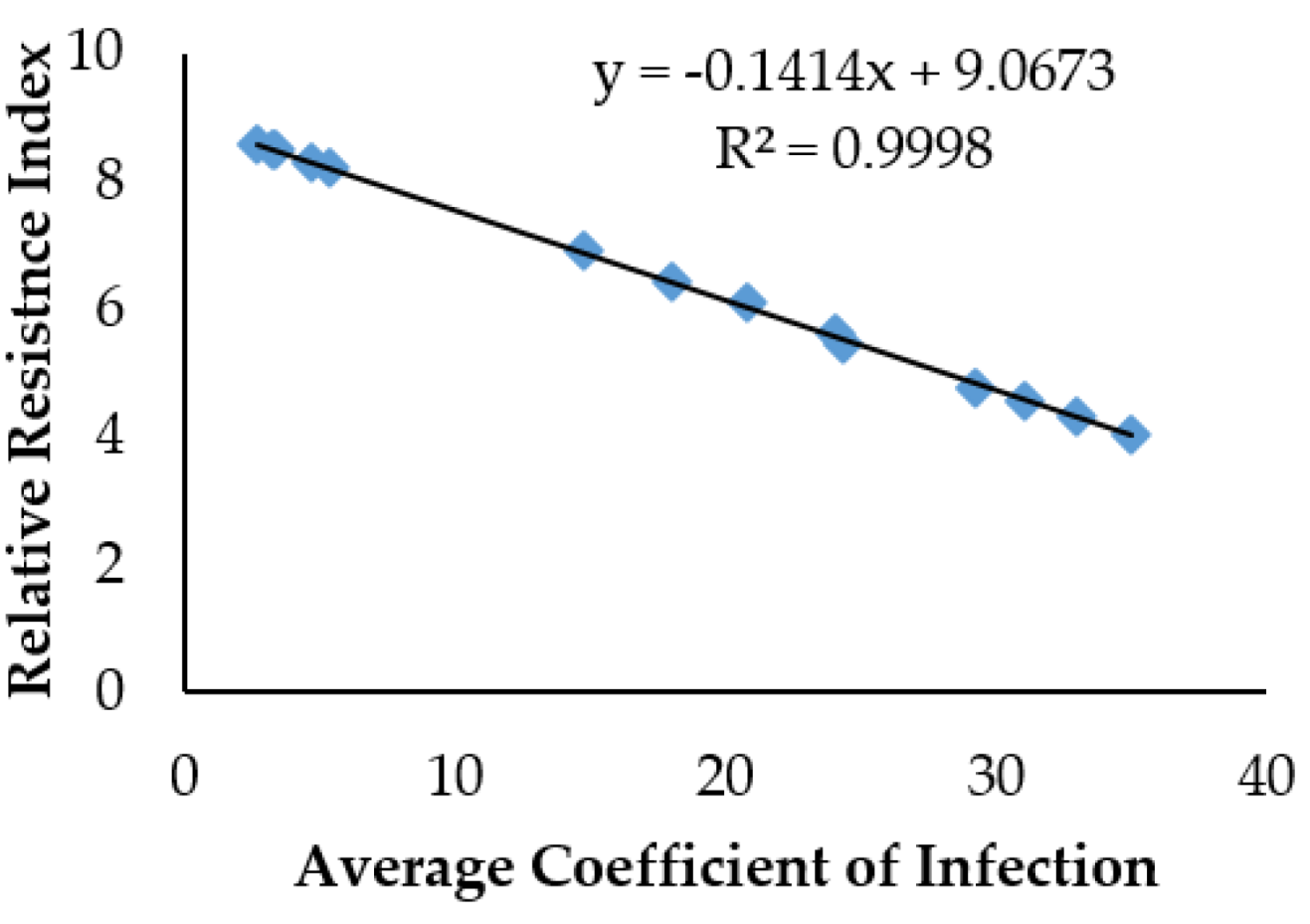
Yellow rust response of the tested wheat genotypes was characterized using two epidemiological parameters: average coefficient of infection (ACI) and RRI. The highest ACI of the tested candidate lines was set at 100 and all other lines were accustomed accordingly to calculate the country average relative percentage attack (CARPA). Using a 0 to 9 scale previously designated as resistance index (RI) was re-designated as relative resistance index (RRI). From CARPA the value of RRI was calculated on the 0 to 9 scale, where 0 denotes the most susceptible and 9 highly resistant [
46]. The RRI was calculated as “RRI = (100 − CARPA)100 × 9”. The susceptible (RRI < 5), moderate (RRI = 5–7), and resistant (RRI > 7) categorization was performed using the protocol of Aslam [
47].
2.5. YR-Coefficient of Infection
Yellow rust coefficient of infection (CI) was noted by multiplying prevailing disease severity with host hypersensitive response, using the protocol of Pathan and Park [
48]. CI was calculated by multiplying the disease infection severity value by its host reaction as follows.
The disease severity is the visual observation of the percent leaf area affected by YR as explained above. Infection type was 1.0 for susceptible (S) host response, 0.8 for moderately susceptible (MS), 0.6 for moderately resistant (MR), and 0.2 for resistant (R). The ACI was the mean CI of replicates for each experimental plot.
2.6. DNA Extraction
The parental genotypes were sampled for DNA and subjected to microsatellite analysis. Fresh young leaf samples were collected from the field in pre-labeled plastic bags and immediately placed on dry ice in a Styrofoam box. A total of 42 plants were sampled for leaves (3 replicates for each of the 14 parental genotypes). DNA was extracted from fresh leaf samples in the laboratory at the Institute of Biotechnology and Genetic Engineering, The University of Agriculture, Peshawar using the CTAB method [
49].
2.7. Polymerase Chain Reaction and Genotyping
Microsatellite markers were amplified in a thermal cycler (T100; Bio-Rad, Hercules, CA, USA). Each reaction was 25 µL with 12.5 µL of 2X DreamTaq Green PCR master mix (K1081, Thermo fisher scientific), 8.5 µL double distilled H
2O, 1 µL each of forward and reverse primers (
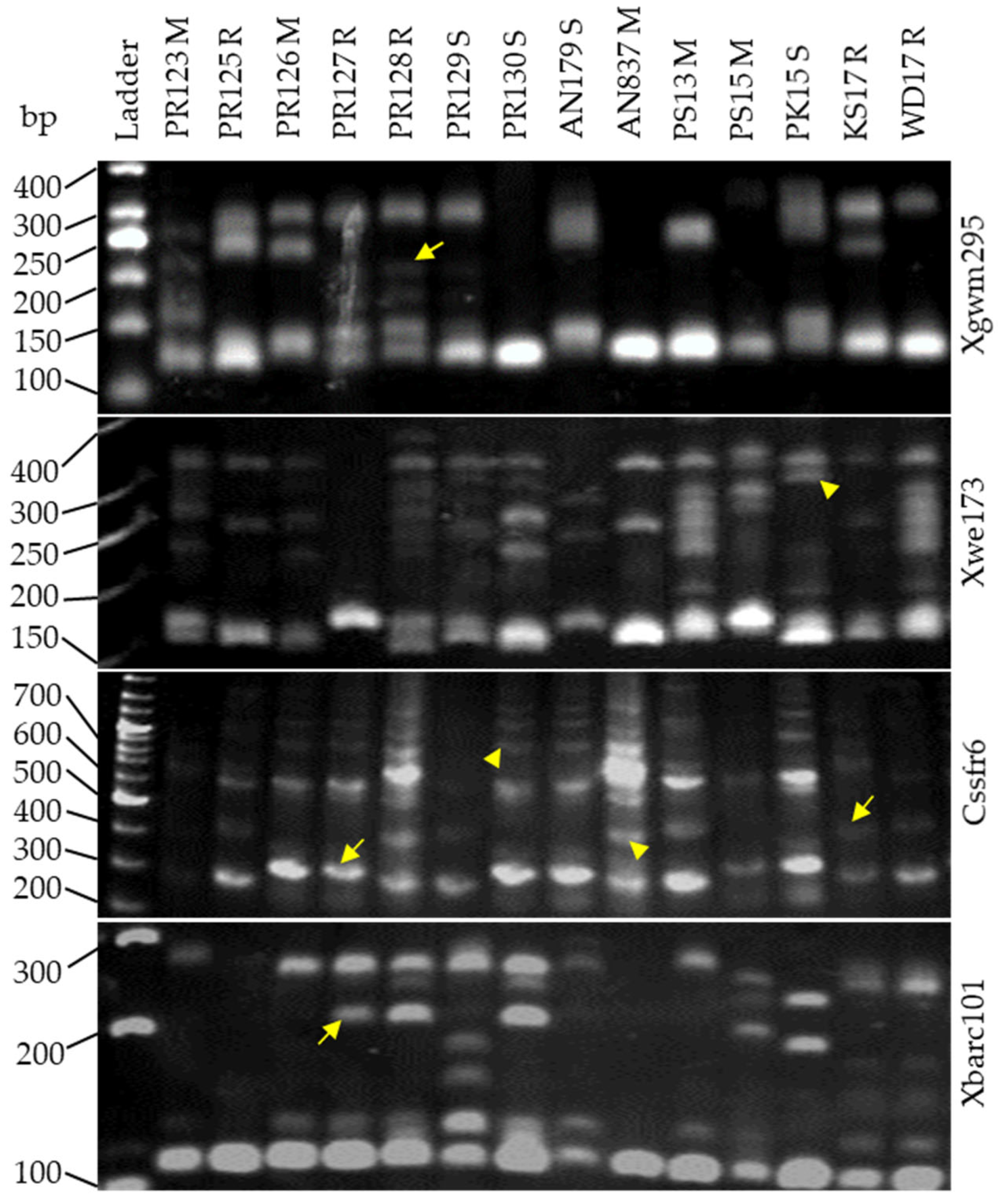
Table 1). A touchdown PCR method was developed to increase PCR specificity. The reaction was run with an initial denaturation at 95 °C for 5 min, followed by 10 repeat cycles (denature at 94 °C for 45 s, primer annealing at 64 °C for 45 s (with a gradual decrement of 1 °C at every cycle), and extension at 72 °C for 45 s. This was followed by another repeat of 25 cycles with a thermal profile of denaturation at 94 °C for 45 s, annealing at 54 °C for 45 s, and extension at 72 °C for 45 s; followed by one last extension cycle at 72 °C for 10 min. In control reactions, sterile distilled water was replaced with template DNA to check for contamination.
The PCR products were resolved on 2% agarose gel along with a 1 kb DNA ladder. The gels were manually scored for the presence and absence (0, 1) of alleles of different sizes for each microsatellite locus. The data were set in a matrix of excel sheets for further analysis.
2.8. Statistical Analysis
The ACI data were used in computer packages MS Excel and Statistix 10.0 for analysis of variance among genotypes. The overall comparison of individual means was calculated by least significant differences (LSD) at a 0.05% probability level. The data were then subjected to the calculation of RRI from the ACI values as explained above. Regression among RRI and ACI was computed by plotting the two variables on a scatter plot using MS Excel.
Means and standard errors for RRI among different parents and RILs, groups of parents (susceptible, moderate, and resistant), and among alleles of significantly associated genotypes were calculated using SPSS software version 23.0. One-way ANOVA along with post hoc Tukey’s test was performed for pair-wise comparisons.
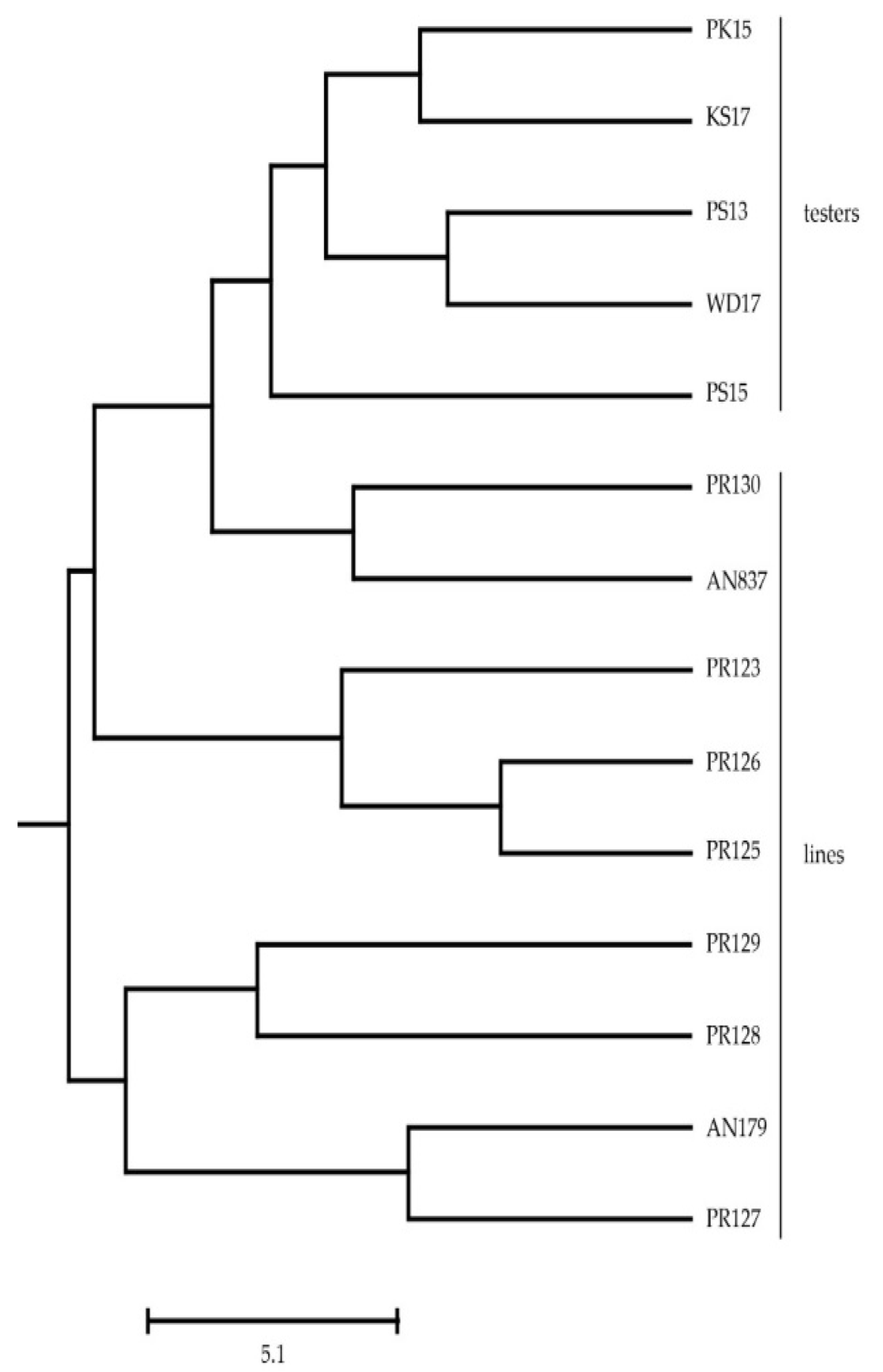
The allelic data of microsatellite markers were analyzed using POPGENE and STRUCTURE software for genetic diversity parameters. Allelic polymorphism and allele frequencies at the microsatellite markers were calculated for susceptible, moderate, and resistant parental groups using POPGENE software. The total number of alleles shared and unique alleles were calculated for each parental group. The microsatellite data were further evaluated for ancestral admixture among the three parental groups based on correlated allele frequencies using STRUCTURE software v2.3.4.
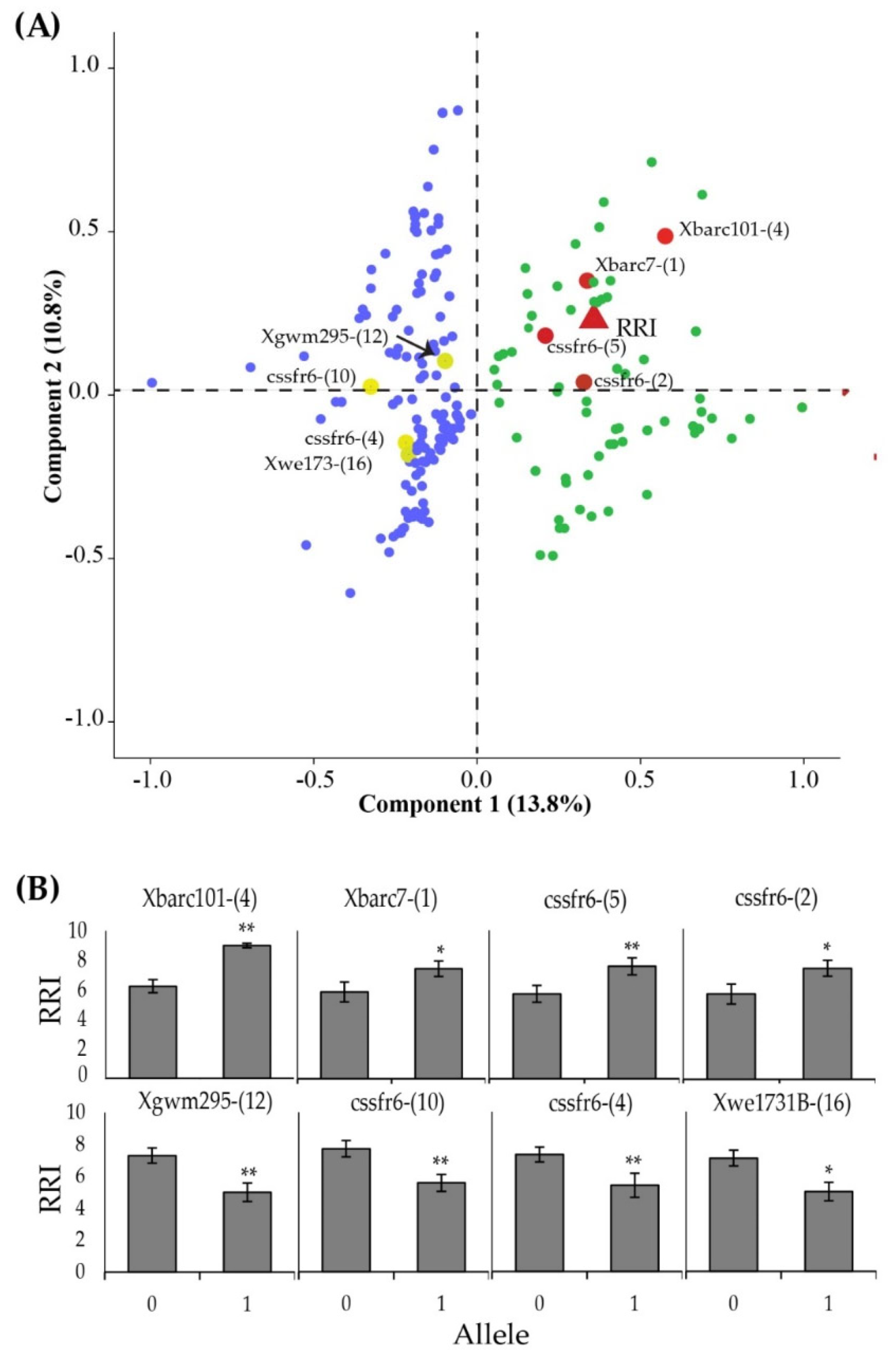
Association between microsatellite allelic polymorphism and RRI was performed using principal component analysis (PCA) in SPSS software. The analysis was based on correlations among RRI and different alleles at each microsatellite marker. Varimax rotation with a convergence of 25 iterations was used for constructing the PCA plot.
4. Discussion
Wheat resistance ratings have abruptly changed as a result of the introduction of genetically diverse exotic
Pst. races into local habitats and their displacement of the clonal
Pst. races. These
Pst. races are distinguished by a number of noteworthy characteristics, such as relatively large reductions in resistance in varieties that previously carried effective long-term APR and high production of sexual stage spores (teliospores) [
52]. Sexual recombination has led to the emergence of hypervirulent
Pst. races. Studies on
Pst. diversity have reported the emergence of new races and how this has led to evasion of resistance conferred by different Yr genes [
14,
17]. One of the current study locations, i.e., Kaghan, is vital as it is a natural habitat for YR alternate host
B. vulgarus, allowing the sexual reproductive cycle required for increasing diversity through recombination and thus the appearance of high virulence
Pst. Pathotypes.
A diverse gene pool from different geographical locations containing novel resistance genes is a prerequisite to developing YR-resistant wheat cultivars. A large and diverse wheat germplasm pool is being maintained by different working groups such as CIMMYT, the International Center for Agricultural Research in the Dry Areas (ICARDA), Pakistan Agricultural Research Council (PARC), the Center for Agricultural Resources Research (CARR), and Chinese Academy of Sciences (CAS) [
53,
54,
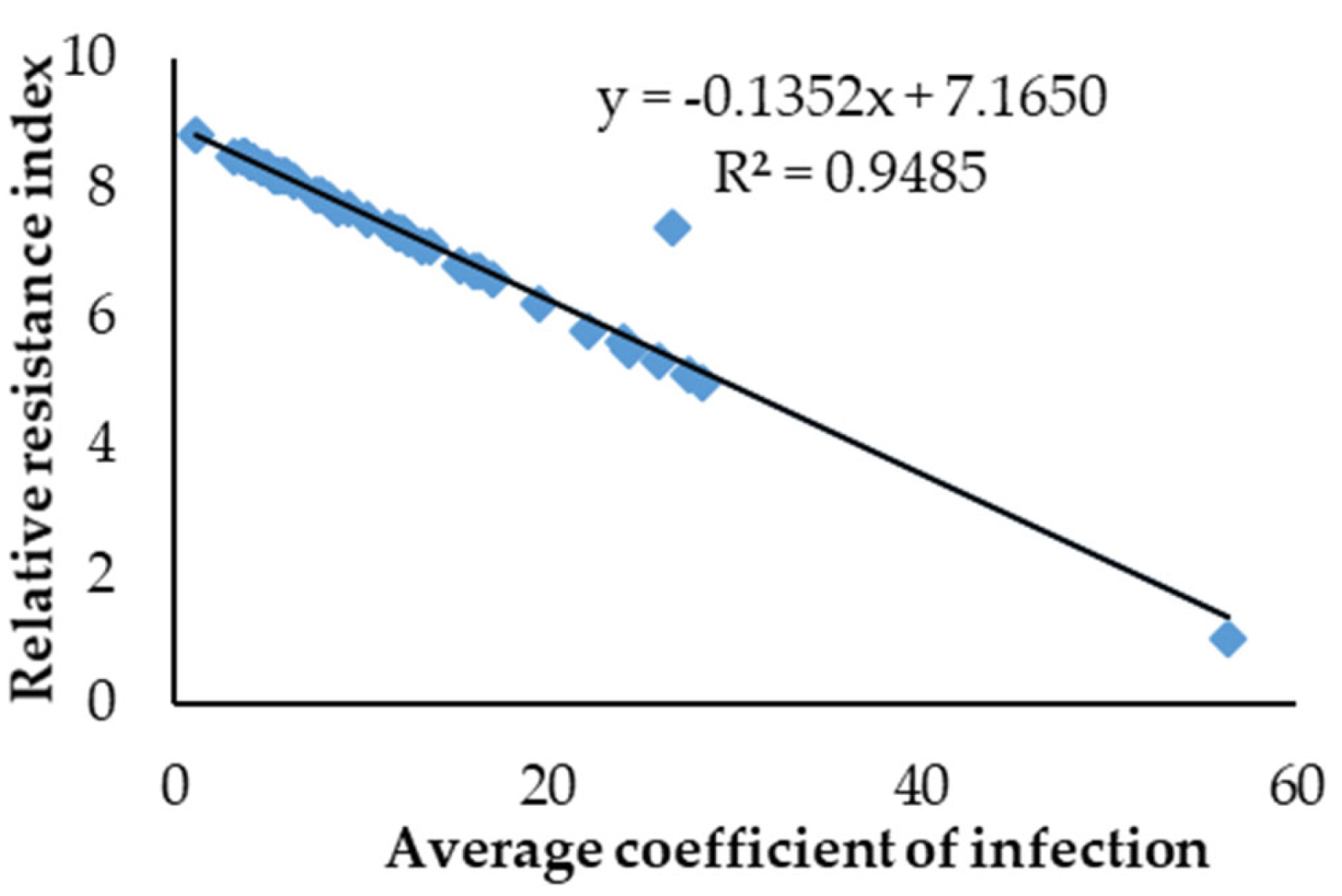
55]. These germplasms are potential gene pools for tackling different challenges, including the utilization of Yr genes. The currently studied wheat germplasms have been found to be highly diverse as indicated by the wide range of ACI expressions. Studies have shown that the RRI of the plants increases as the ACI decreases [
56], which is consistent with the findings from the current study.
This study in the YR hotspot aimed to profile different wheat genotypes using RRI based on ACI under artificial inoculation of a known
Pst. virulent strain
Pst. 574232. However, natural infection is also predominant in the region, which may also have infected the experimental plants. The
Pst. strain used in this study has been found to be avirulent against Yr5, 10, 15, 24, 32, SP, and Tye, whereas virulent against Yr1, 6, 7, 8, 9, 17, 27, 43, 44, and Exp2 [
57]. Desirable RRI values have been used as a successful tool for the incorporation of YR resistance in wheat-breeding programs [
58]. Three distinct resistance bulks were identified among the tested germplasm, exhibiting complete, moderate, and no resistance against YR. Findings from this study revealed that out of 14, five parental lines were fully resistant against YR, with an RRI value > 8; considered a highly desirable resistance index [
46]. In previous studies, nine out of fifty Egyptian wheat RILs have been identified as resistant based on the RRI values above [
59]. However, classical breeding approaches are too slow and outpaced by the fast-evolving pathogen [
60].
Faster alternates to classical approaches include the development of transgenic wheat lines expressing Yr genes [
61]. Therefore, identification of Yr genes/QTL is required for incorporation into speed breeding programs to improve YR resistance among the wheat cultivars and advance lines. Microsatellite markers have been described as markers of choice to identify alleles associated with Yr genes [
62]. In the current study, we have found novel alleles at different microsatellite markers unique to different resistance bulks. Moreover, we found some higher frequency alleles within the resistant genotypes bulk. These alleles may be of significant importance for the selection of YR-resistant RILs in successive generations.
Yr18 has been identified as an APR gene, which has conferred stable and durable resistance against YR over the last century [
36]. The marker Xgwm295 was reported to be in close proximity to Yr18 [
63]. Our results suggest a significant effect of polymorphism at marker Xgwm295 (due to the presence of unique alleles among resistance bulks) on resistance against YR due to its linkage with the Yr18 gene. In previous studies, Yr QTL in bread wheat has been identified at marker Xgwm295 [
64]. Furthermore, we have identified four alleles at the Yr18-linked marker “cssfr6” showing a significant correlation with RRI. This is suggestive of the presence of Yr18-based resistance among the studied genotypes. Previous studies have shown a close association of the cssfr6 marker with other resistance genes against leaf rust (Lr34) and powdery mildew (Pm38) as well [
65]. These results suggest the effectiveness of cssfr6 in differentiating resistant genotypes from susceptible. The absence of Yr18-linked alleles at cssfr6 and other markers has been shown to result in YR-susceptible Australian wheat genotypes [
66]. Two moderate and four resistant phenotype-linked alleles at the csLV34 marker identified in the current study are also known to be in close proximity to the Yr18 gene. Studies in China, Ukraine, and Russia involving Lr34/Yr18/Pm38-linked resistance breeding studies have found a strong linkage of these genes with the marker csLV34 [
67]. The highly polymorphic Yr18-linked markers Xgwm295, cssfr6, and csLV34 may be effective in MAS for resistance breeding in wheat.
The Yr5 and Yr15 associated markers analysis have resulted in the identification of unique alleles at markers Xbarc7 and Xgwm11. In previous studies, alleles Xbarc7 have been associated with the Lr13 gene [
68] but found weakly linked with the Yr5 gene on chromosome 2B. No explicit reports of its association with any of the known Yr genes have been found. However, this study has demonstrated that two alleles, i.e., Xbarc7–4 and Xbarc7–16, were frequently found in the moderately resistant bulk, whereas Xbarc7–1 was found among the resistant bulk. Findings from the current study indicate that Xbarc7–1 may be used as a novel marker for the selection of YR-resistant genotypes. The allele Xbrac1014 was significantly associated with RRI in the current study; is moderately linked to the Yr5 genes. In previous studies, marker Xbarc101 has been found in close proximity to the YrSP gene (seedling YR resistance) [
69]. Based on findings from this study, the Xbarc1014 allele may be co-located with a potential Yr resistance gene/QTL. Another important Yr gene (Yr15) was found in close proximity to marker Xgwm11 in the current study. Two unique alleles were found at marker Xgwm11 in moderately resistant and resistant genotypes, suggesting its positive association with RRI due to its linkage with the Yr15 gene. The Yr5 and Yr15 have been identified previously as ASR genes providing resistance against all prevailing races of YR in the USA, China, Pakistan, Egypt, and Africa [
70,
71,
72,
73,
74].
The Yr24/Yr26 has been widely used among Chinese wheat germplasm as a source of resistance against YR [
75]. Recently, reports suggest that highly virulent
Pst. strains have been able to break the Yr26-based resistance in some Chinese advanced lines, with a virulence percentage in the range of 73–98 percent [
26,
76,
77]. The current data from this study have identified the Yr26–linked marker Xwe173 (on chromosome 1B) as having a strong negative association with RRI. Higher infectivity in lines having Xwe173 negative alleles has been reported in RILs of synthetic-derived wheat line Soru#1 [
78]. Allele Xwe173–16 identified in the current study may be used as a marker to screen out highly susceptible YR phonotypes linked to the breakage of Yr26-based resistance.
The Yr62 gene was originally identified as a major effect QTL (QYrPI192252.wgp–4BL) on chromosome 4B with the ability to provide HTAP (explaining up to 60% phenotypic variance for YR resistance) [
79]. In this study, Yr62 gene-linked allele Xgwm25110 was found to be prevalent among moderate resistant bulk. The slow rusting phenotype in the APR-associated resistant genotypes expressing Yr62 genes is consistent with findings from the current study. Yr62 is another potential resistance gene, especially for areas where hot summers are prevalent.
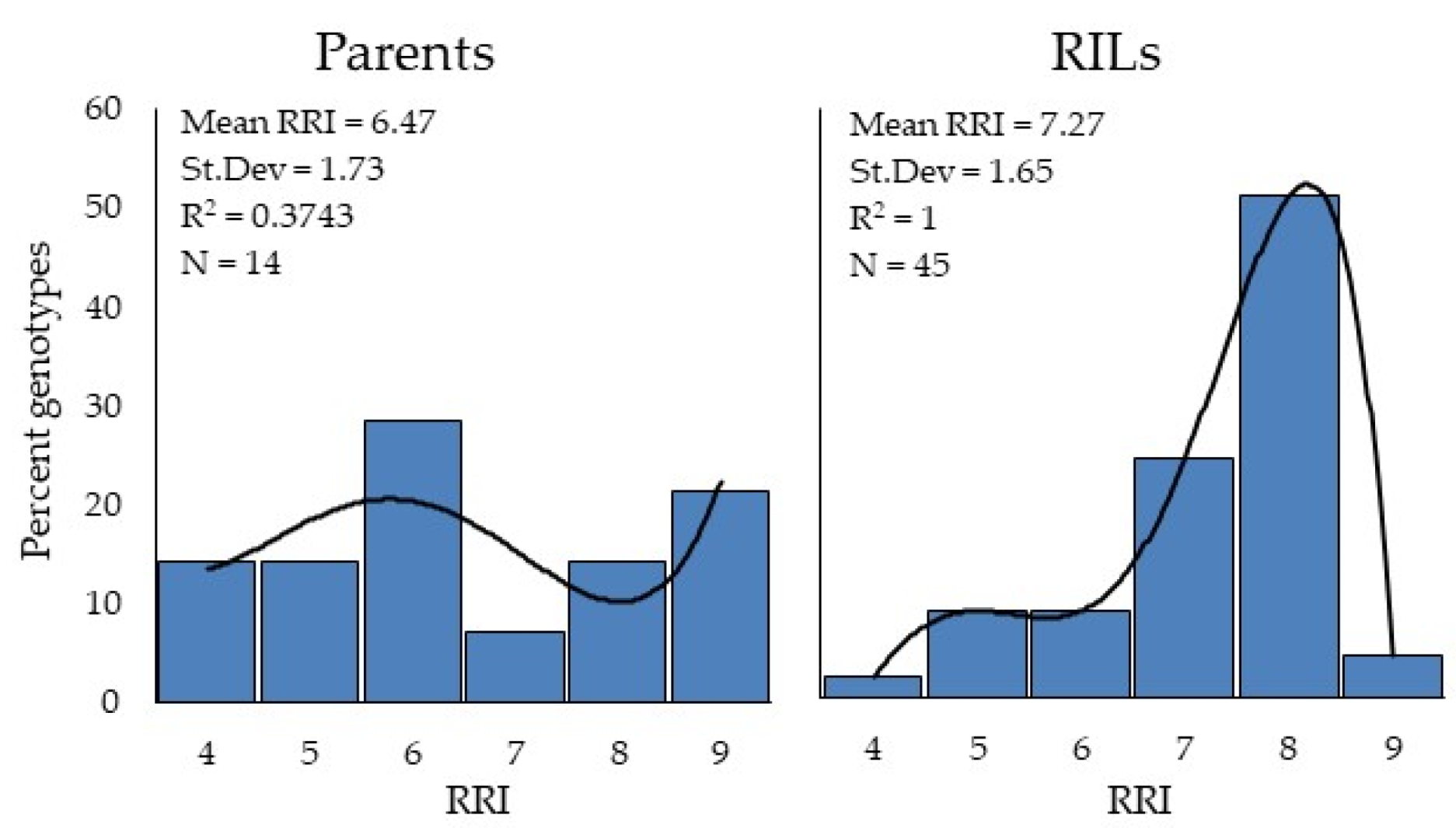
The data were recorded for the F6 generation of the RILs as by such time about 95% of the genotypes are expected to be homozygous for YR resistance. An upward shift in RRI (8.0) in half of the RILs as compared to 6.0 in one-third of the parental population may be explained by carryover and favorable recombination among potential Yr QTL and genes. Previous studies have found the carryover effect of Yr QTL, e.g., QYr.inra–2BL was found to count for 61% of the resistance phenotype variability among RIL [
69]. QYrco.wpg–1B.1, QYrco.wpg–1B.2, and other QTL together resulted in up to 48% disease reduction in the RIL populations [
80]. Based on the microsatellite marker analysis, diverse allelic distribution in the parent genotypes and their association with RRI could explain the significant increase in RRI among the resultant RIL population. This suggests that the identified genetic diversity among the parents, including novel alleles associated with variance in resistance phenotypes, is a rich genetic resource and could be used to improve YR resistance in wheat breeding programs.

 ,
, 











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}