
Genome-Wide Transcriptome Analysis Reveals That Upregulated Expression of Aux/IAA Genes Is Associated with Defective Leaf Growth of the slf Mutant in Eggplant

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Leaf Size Measurement
2.3. Histological Analysis
2.4. Measuring the Main Hormone Content
2.5. RNA-seq Analysis
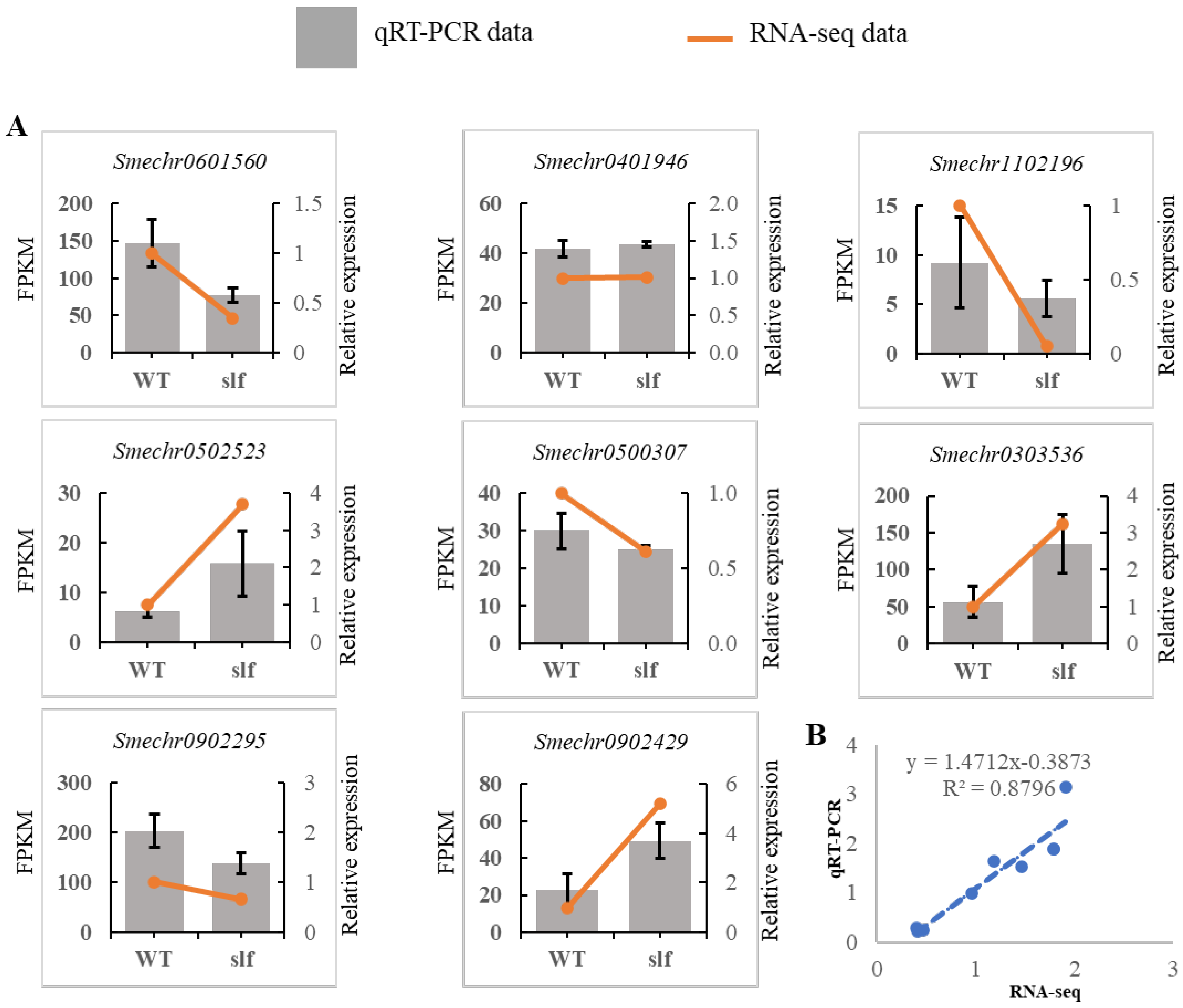
2.6. Validation of Selected DEGs Using Real-Time Quantitative Reverse Transcription PCR (qRT-PCR)
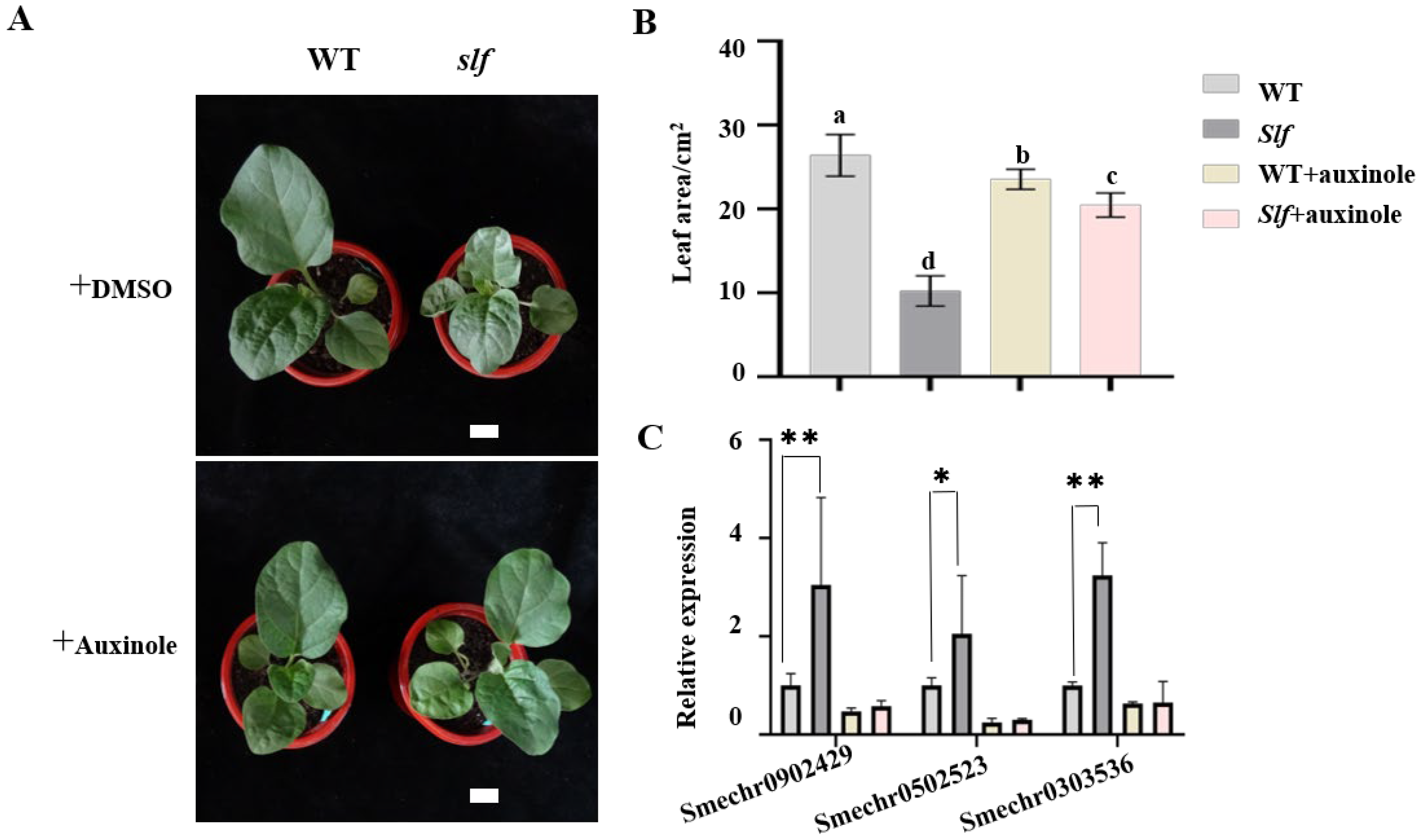
2.7. Exogenous Auxinole Treatments
2.8. Statistical Analysis
3. Results
3.1. Phenotypic Comparison of the WT and Slf Mutant
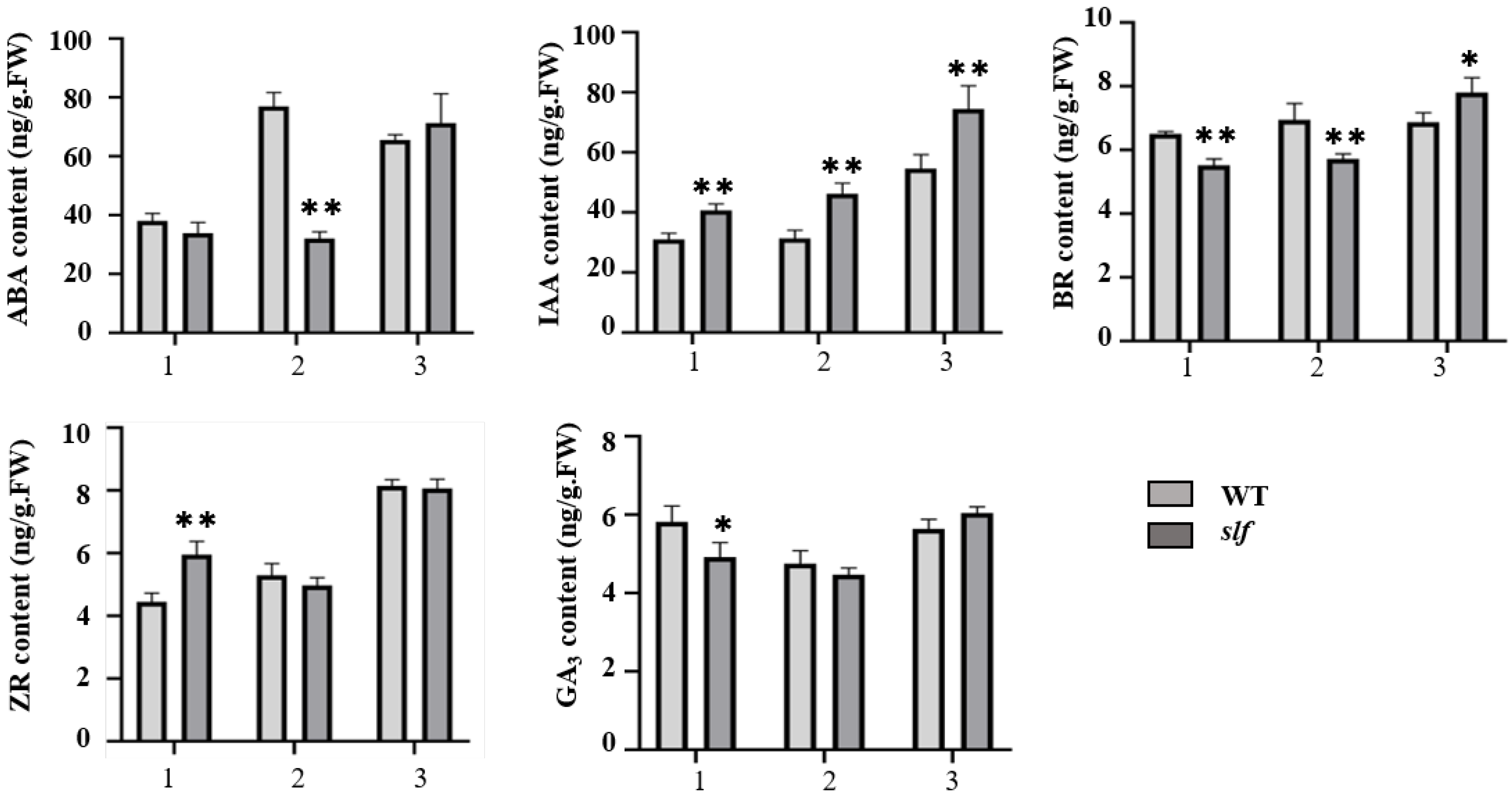
3.2. Analysis of the Contents of ABA, IAA, BR, GA3 and ZR
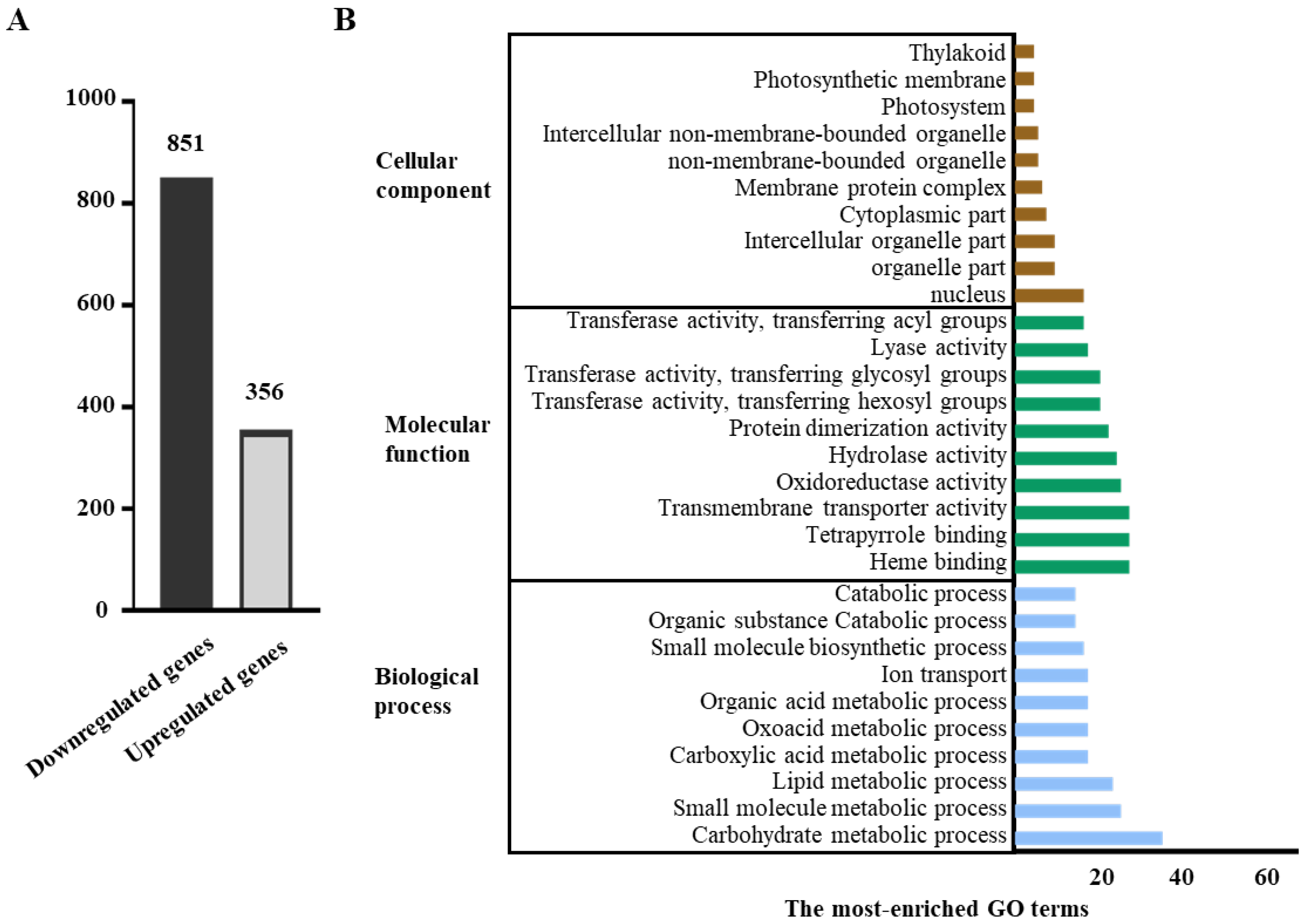
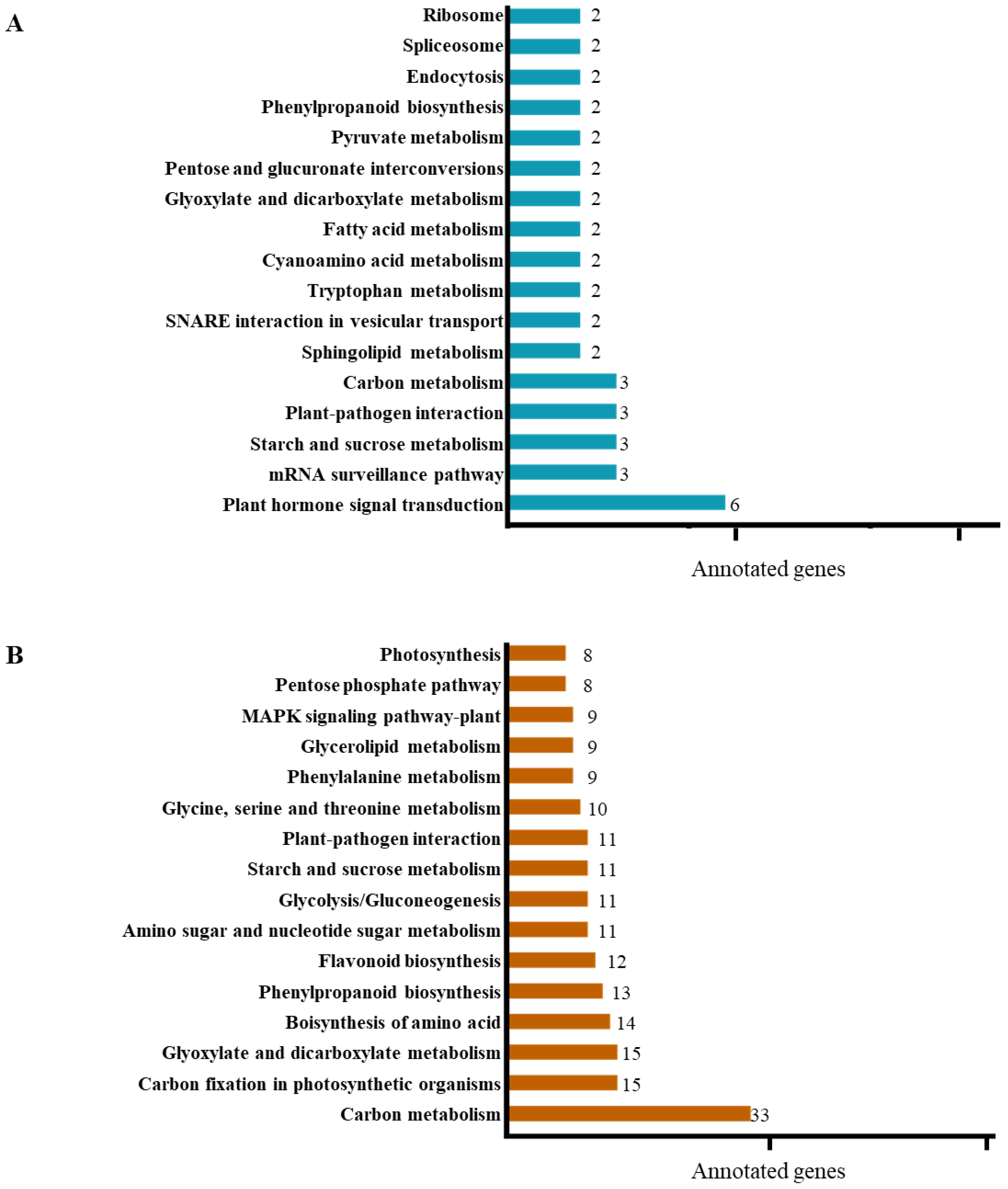
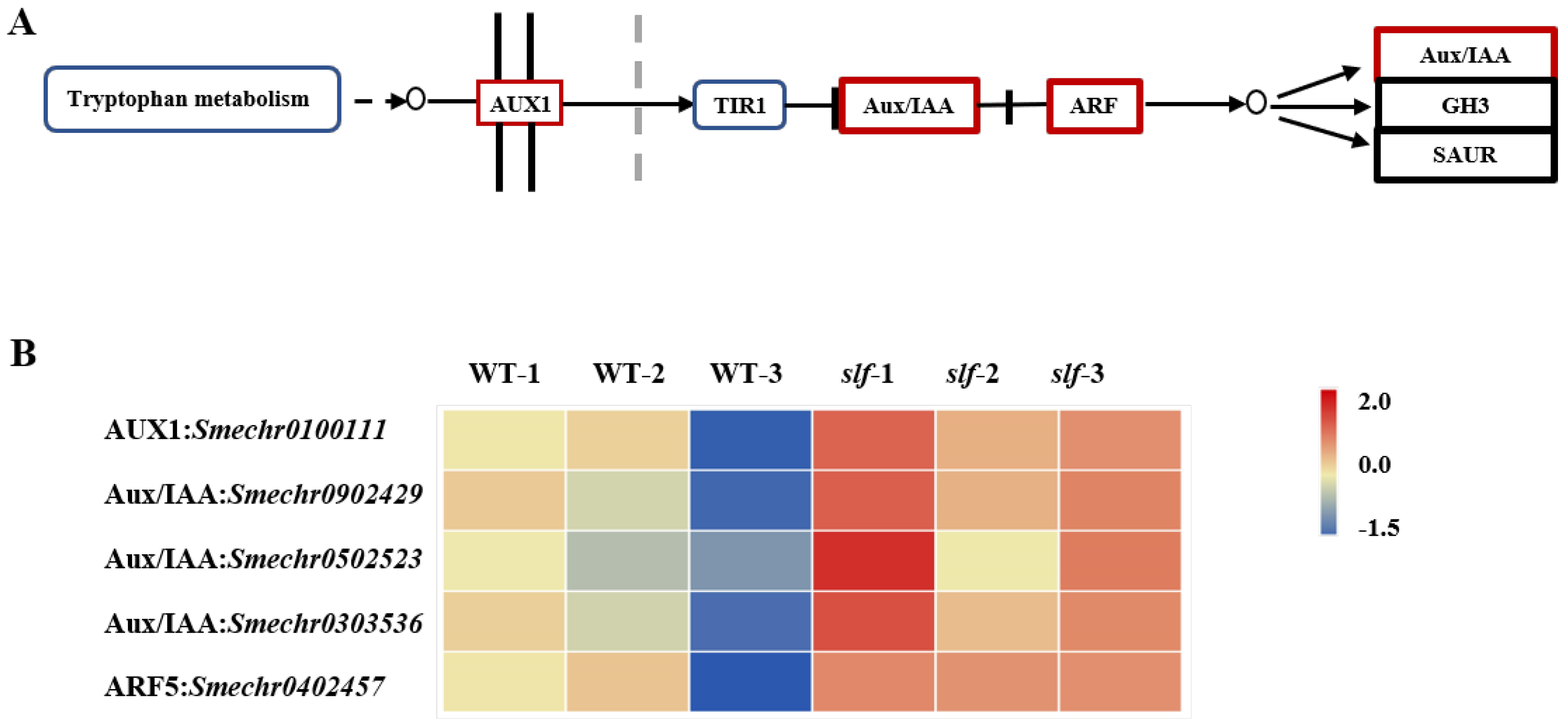
3.3. Transcriptome Profiles Showed That Genes Involved in Auxin Signalling Are Enhanced in Slf
3.4. The Effect of Exogenous Auxinole on the Leaf Size of WT and slf
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wei, Q.; Wang, J.; Wang, W.; Hu, T.; Hu, H.; Bao, C. A high-quality chromosome-level genome assembly reveals genetics for important traits in eggplant. Hortic. Res. 2020, 7, 153. [Google Scholar] [CrossRef] [PubMed]
- Kashyap, V.; Kumar, S.V.; Collonnier, C.; Fusari, F.; Haicour, R.; Rotino, G.L.; Sihachakr, D.; Rajam, M. Biotechnology of eggplant. Sci. Hortic. 2003, 97, 1–25. [Google Scholar] [CrossRef]
- Hanifah, A.; Maharijaya, A.; Putri, S.P.; Lavina, W.A.; Sobir. Untargeted metabolomics analysis of eggplant (Solanum melongena L.) fruit and its correlation to fruit morphologies. Metabolites 2018, 8, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mauceri, A.; Bassolino, L.; Lupini, A.; Badeck, F.; Rizza, F.; Schiavi, M.; Toppino, L.; Abenavoli, M.R.; Rotino, G.L.; Sunseri, F. Genetic variation in eggplant for nitrogen use efficiency under contrasting NO3− supply. J. Integr. Plant Biol. 2020, 62, 487–508. [Google Scholar] [CrossRef] [Green Version]
- Liu, N.; Wang, Q.; He, C.; An, B. CgMFS1, a major facilitator superfamily transporter, is required for sugar transport, oxidative stress resistance, and pathogenicity of colletotrichum gloeosporioides from Hevea brasiliensis. Curr. Issues Mol. Biol. 2021, 43, 1548–1557. [Google Scholar] [CrossRef] [PubMed]
- Mahalle, M.D.; Chetia, S.K.; Dey, P.C.; Sarma, R.N.; Baruah, A.R.; Kaldate, R.C.; Verma, R.K.; Modi, M.K. Assessing the leaf shape dynamic through marker-trait association under drought stress in a rice germplasm panel. Plant Genet. Resour. 2022, 19, 477–483. [Google Scholar] [CrossRef]
- Chitwood, D.H.; Ranjan, A.; Kumar, R.; Ichihashi, Y.; Zumstein, K.; Headland, L.R.; Ostria-Gallardo, E.; Aguilar-Martinez, J.A.; Bush, S.; Carriedo, L.; et al. Resolving distinct genetic regulators of tomato leaf shape within a heteroblastic and ontogenetic context. Plant Cell 2014, 26, 3616–3629. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.; Li, G.; Sun, S. The effects of leaf size, leaf habit, and leaf form on leaf/stem relationships in plant twigs of temperate woody species. J. Veg. Sci. 2009, 20, 359–366. [Google Scholar] [CrossRef]
- Fu, Z.; Martin, C.E.; Do, J.; Ho, C.-L.; Wagner, B. Functional relationship between leaf/stem pseudobulb size and photosynthetic pathway in the orchidaceae. Can. J. Plant Sci. 2022, 102, 419–426. [Google Scholar] [CrossRef]
- Valencia, E.; Quero, J.L.; Maestre, F.T. Functional leaf and size traits determine the photosynthetic response of 10 dryland species to warming. J. Plant Ecol. 2016, 9, 773–783. [Google Scholar] [CrossRef]
- Famuwagun, A.A.; Alashi, A.M.; Gbadamosi, S.O.; Taiwo, K.A.; Oyedele, D.; Adebooye, O.C.; Aluko, R.E. Effect of protease type and peptide size on the in vitro antioxidant, antihypertensive and anti-diabetic activities of eggplant leaf protein hydrolysates. Foods 2021, 10, 1112. [Google Scholar] [CrossRef]
- Hay, A.; Barkoulas, M.; Tsiantis, M. PINning down the connections: Transcription factors and hormones in leaf morphogenesis. Curr. Opin. Plant Biol. 2004, 7, 575–581. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, N.; Vanhaeren, H.; Inze, D. Leaf size control: Complex coordination of cell division and expansion. Trends Plant Sci. 2012, 17, 332–340. [Google Scholar] [CrossRef] [PubMed]
- Karamat, U.; Sun, X.; Li, N.; Zhao, J. Genetic regulators of leaf size in brassica crops. Hortic. Res. 2021, 8, 91. [Google Scholar] [CrossRef] [PubMed]
- Rojas, C.A.; Eloy, N.B.; Lima, M.d.F.; Rodrigues, R.L.; Franco, L.O.; Himanen, K.; Beemster, G.T.S.; Hemerly, A.S.; Gomes Ferreira, P.C. Overexpression of the Arabidopsis anaphase promoting complex subunit CDC27a increases growth rate and organ size. Plant Mol. Biol. 2009, 71, 307–318. [Google Scholar] [CrossRef]
- Kim, J.H.; Kende, H. A transcriptional coactivator, AtGIF1, is involved in regulating leaf growth and morphology in Arabidopsis. Proc. Natl. Acad. Sci. USA 2004, 101, 13374–13379. [Google Scholar] [CrossRef] [Green Version]
- Dewitte, W.; Scofield, S.; Alcasabas, A.A.; Maughan, S.C.; Menges, M.; Braun, N.; Collins, C.; Nieuwland, J.; Prinsen, E.; Sundaresan, V.; et al. Arabidopsis CYCD3 d-type cyclins link cell proliferation and endocycles and are rate-limiting for cytokinin responses. Proc. Natl. Acad. Sci. USA 2007, 104, 14537–14542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, H.T.; Cosgrove, D.J. Altered expression of expansin modulates leaf growth and pedicel abscission in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2000, 97, 9783–9788. [Google Scholar] [CrossRef] [Green Version]
- Horvath, B.M.; Magyar, Z.; Zhang, Y.; Hamburger, A.W.; Bako, L.; Visser, R.G.F.; Bachem, C.W.B.; Bogre, L. EBP1 regulates organ size through cell growth and proliferation in plants. Embo J. 2006, 25, 4909–4920. [Google Scholar] [CrossRef] [PubMed]
- Schruff, M.C.; Spielman, M.; Tiwari, S.; Adams, S.; Fenby, N.; Scott, R.J. The AUXIN RESPONSE FACTOR 2 gene of Arabidopsis links auxin signalling, cell division, and the size of seeds and other organs. Development 2006, 133, 251–261. [Google Scholar] [CrossRef]
- Ubeda-Tomas, S.; Federici, F.; Casimiro, I.; Beemster, G.T.S.; Bhalerao, R.; Swarup, R.; Doerner, P.; Haseloff, J.; Bennett, M.J. Gibberellin signaling in the endodermis controls Arabidopsis root meristem size. Curr. Biol. 2009, 19, 1194–1199. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, R.E.; Mecchia, M.A.; Debernardi, J.M.; Schommer, C.; Weigel, D.; Palatnik, J.F. Control of cell proliferation in Arabidopsis thaliana by microRNA mir396. Development 2010, 137, 103–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Zheng, L.; Corke, F.; Smith, C.; Bevan, M.W. Control of final seed and organ size by the DA1 gene family in Arabidopsis thaliana. Gene Dev. 2008, 22, 1331–1336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, B.; Yan, Z.; Tian, H.; Zhang, X.; Ding, Z. Local Auxin biosynthesis mediates plant growth and development. Trends Plant Sci. 2019, 24, 6–9. [Google Scholar] [CrossRef] [PubMed]
- Keller, C.P.; Stahlberg, R.; Barkawi, L.S.; Cohen, J.D. Long-term inhibition by auxin of leaf blade expansion in bean and arabidopsis. Plant Physiol. 2004, 134, 1217–1226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Vos, D.; Nelissen, H.; AbdElgawad, H.; Prinsen, E.; Broeckhove, J.; Inze, D.; Beemster, G.T.S. How grass keeps growing: An integrated analysis of hormonal crosstalk in the maize leaf growth zone. New Phytol. 2020, 225, 2513–2525. [Google Scholar] [CrossRef]
- Hagen, G. Auxin signal transduction. Plant Horm. Signal. 2015, 58, 1–12. [Google Scholar] [CrossRef]
- Salehin, M.; Bagchi, R.; Estelle, M. SCFTIR1/AFB-based Auxin perception: Mechanism and role in plant growth and development. Plant Cell 2015, 27, 9–19. [Google Scholar] [CrossRef] [Green Version]
- Todd, O.E.; Figueiredo, M.R.A.; Morran, S.; Soni, N.; Preston, C.; Kubes, M.F.; Napier, R.; Gaines, T.A. Synthetic auxin herbicides: Finding the lock and key to weed resistance. Plant Sci. 2020, 300, 110631. [Google Scholar] [CrossRef]
- Pan, J.; Fujioka, S.; Peng, J.; Chen, J.; Li, G.; Chen, R. The E3 Ubiquitin Ligase SCFTIR1/AFB and membrane sterols play key roles in Auxin regulation of endocytosis, recycling, and plasma membrane accumulation of the Auxin efflux transporter PIN2 in Arabidopsis thaliana. Plant Cell 2009, 21, 568–580. [Google Scholar] [CrossRef]
- Yu, Z.; Zhang, F.; Friml, J.; Ding, Z. Auxin signaling: Research advances over the past 30 years. J. Integr. Plant Biol. 2022, 64, 371–392. [Google Scholar] [CrossRef] [PubMed]
- Strader, L.C.; Zhao, Y. Auxin perception and downstream events. Curr. Opin. Plant Biol. 2016, 33, 8–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dezfulian, M.H.; Jalili, E.; Roberto, D.K.A.; Moss, B.L.; Khoo, K.; Nemhauser, J.L.; Crosby, W.L. Oligomerization of SCFTIR1 is essential for Aux/IAA degradation and Auxin signaling in Arabidopsis. PLoS Genet. 2016, 12, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quint, M.; Gray, W.M. Auxin signaling. Curr. Opin. Plant Biol. 2006, 9, 448–453. [Google Scholar] [CrossRef]
- Takato, S.; Kakei, Y.; Mitsui, M.; Ishida, Y.; Suzuki, M.; Yamazaki, C.; Hayashi, K.-i.; Ishii, T.; Nakamura, A.; Soeno, K.; et al. Auxin signaling through SCFTIR1/AFBs mediates feedback regulation of IAA biosynthesis. Biosci. Biotech Bioch. 2017, 81, 1320–1326. [Google Scholar] [CrossRef] [Green Version]
- Si Ammour, A.; Windels, D.; Arn Bouldoires, E.; Kutter, C.; Ailhas, J.; Meins, F., Jr.; Vazquez, F. miR393 and secondary siRNAs regulate expression of the TIR1/AFB2 Auxin receptor clade and Auxin-related development of Arabidopsis leaves. Plant Physiol. 2011, 157, 683–691. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Li, J.; Cui, L.; Zhang, T.; Wu, Z.; Zhu, P.; Meng, Y.; Zhang, K.; Yu, X.; Lou, Q.; et al. New insights into the roles of cucumber TIR1 homologs and miR393 in regulating fruit/seed set development and leaf morphogenesis. BMC Plant Biol. 2017, 17, 130. [Google Scholar] [CrossRef] [Green Version]
- Rouse, D.; Mackay, P.; Stirnberg, P.; Estelle, M.; Leyser, O. Changes in auxin response from mutations in an AUX/IAA gene. Science 1998, 279, 1371–1373. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Jones, B.; Li, Z.G.; Frasse, P.; Delalande, C.; Regad, F.; Chaabouni, S.; Latche, A.; Pech, J.C.; Bouzayen, M. The tomato Aux/IAA transcription factor IAA9 is involved in fruit development and leaf morphogenesis. Plant Cell 2005, 17, 2676–2692. [Google Scholar] [CrossRef] [Green Version]
- Deng, W.; Yan, F.; Liu, M.; Wang, X.; Li, Z. Down-regulation of SlIAA15 in tomato altered stem xylem development and production of volatile compounds in leaf exudates. Plant Signal Behav. 2012, 7, 911–913. [Google Scholar] [CrossRef] [PubMed]
- Windels, D.; Bielewicz, D.; Ebneter, M.; Jarmolowski, A.; Szweykowska Kulinska, Z.; Vazquez, F. miR393 is required for production of proper Auxin signalling outputs. PLoS ONE 2014, 9, e95972. [Google Scholar] [CrossRef] [Green Version]
- Trainotti, L.; Pavanello, A.; Casadoro, G. Differential expression of genes in apical and basal tissues of expanding tobacco leaves. Plant Sci. 2004, 167, 679–686. [Google Scholar] [CrossRef]
- Ku, S.; Park, J.; Ha, S.; Kim, J. Overexpression of iaa1 with domain II mutation impairs cell elongation and cell division in inflorescences and leaves of Arabidopsis. J. Plant Physiol. 2009, 166, 548–553. [Google Scholar] [CrossRef] [PubMed]
- Krogan, N.T.; Ckurshumova, W.; Marcos, D.; Caragea, A.E.; Berleth, T. Deletion of MP/ARF5 domains III and IV reveals a requirement for Aux/IAA regulation in Arabidopsis leaf vascular patterning. New Phytol. 2012, 194, 391–401. [Google Scholar] [CrossRef]
- Matthes, M.S.; Best, N.B.; Robil, J.M.; Malcomber, S.; Gallavotti, A.; McSteen, P. Auxin evodevo: Conservation and diversification of genes regulating Auxin biosynthesis, transport, and signaling. Mol. Plant 2019, 12, 298–320. [Google Scholar] [CrossRef] [Green Version]
- Okushima, Y.; Mitina, I.; Quach, H.L.; Theologis, A. AUXIN RESPONSE FACTOR 2 (ARF2): A pleiotropic developmental regulator. Plant J. 2005, 43, 29–46. [Google Scholar] [CrossRef]
- Bennett, M.J.; Marchant, A.; Green, H.G.; May, S.T.; Ward, S.P.; Millner, P.A.; Walker, A.R.; Schulz, B.; Feldmann, K.A. Arabidopsis AUX1 gene: A permease-like regulator of root gravitropism. Science 1996, 273, 948–950. [Google Scholar] [CrossRef]
- Schroeder, A.B.; Dobson, E.T.A.; Rueden, C.T.; Tomancak, P.; Jug, F.; Eliceiri, K.W. The imagej ecosystem: Open-source software for image visualization, processing, and analysis. Protein Sci. 2021, 30, 234–249. [Google Scholar] [CrossRef]
- Popova, L.P.; Tsonev, T.D.; Lazova, G.N.; Stoinova, Z.G. Drought- and ABA-induced changes in photosynthesis of barley plants. Physiol. Plant. 1996, 96, 623–629. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. Hisat: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. Tophat: Discovering splice junctions with RNA-seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Smyth, G.K.; Shi, W. Featurecounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with deseq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarazona, S.; Garcia-Alcalde, F.; Dopazo, J.; Ferrer, A.; Conesa, A. Differential expression in RNA-seq: A matter of depth. Genome Res. 2011, 21, 2213–2223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.; Zhang, H.; Tan, J.; Huang, S.; Chen, X.; Jiang, D.; Xiao, X. Transcriptome analysis of eggplant root in response to root-knot nematode infection. Pathogens 2021, 10, 470. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−∆∆Ct method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Hayashi, K.-i.; Neve, J.; Hirose, M.; Kuboki, A.; Shimada, Y.; Kepinski, S.; Nozaki, H. Rational design of an Auxin antagonist of the SCFTIR1 Auxin receptor complex. ACS Chem. Biol. 2012, 7, 590–598. [Google Scholar] [CrossRef]
- Mishra, P.; Singh, U.; Pandey, C.M.; Mishra, P.; Pandey, G. Application of student’s t-test, analysis of variance, and covariance. Ann. Card. Anaesth. 2019, 22, 407–411. [Google Scholar] [CrossRef]
- Schrader, J.; Shi, P.; Royer, D.L.; Peppe, D.J.; Gallagher, R.V.; Li, Y.; Wang, R.; Wright, I.J. Leaf size estimation based on leaf length, width and shape. Ann. Bot. 2021, 128, 395–406. [Google Scholar] [CrossRef]
- Wang, P.; Zhou, G.; Cui, K.; Li, Z.; Yu, S. Clustered QTL for source leaf size and yield traits in rice (Oryza sativa l.). Mol. Breed. 2012, 29, 99–113. [Google Scholar] [CrossRef]
- Zhao, C.; Bao, Y.; Wang, X.; Yu, H.; Ding, A.; Guan, C.; Cui, J.; Wu, Y.; Sun, H.; Li, X.; et al. QTL for flag leaf size and their influence on yield-related traits in wheat. Euphytica 2018, 214, 209. [Google Scholar] [CrossRef]
- Serce, S.; Navazio, J.P.; Gokce, A.F.; Staub, J.E. Nearly isogenic cucumber genotypes differing in leaf size and plant habit exhibit differential response to water stress. J. Am. Soc. Hortic. Sci. 1999, 124, 358–365. [Google Scholar] [CrossRef] [Green Version]
- Schurr, U.; Heckenberger, U.; Herdel, K.; Walter, A.; Feil, R. Leaf development in ricinus communis during drought stress: Dynamics of growth processes of cellular structure and of sink-source transition. J. Exp. Bot. 2000, 51, 1515–1529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shwartz, I.; Levy, M.; Ori, N.; Bar, M. Hormones in tomato leaf development. Dev. Biol. 2016, 419, 132–142. [Google Scholar] [CrossRef]
- Dindas, J.; Scherzer, S.; Roelfsema, M.R.G.; von Meyer, K.; Mueller, H.M.; Al Rasheid, K.A.S.; Palme, K.; Dietrich, P.; Becker, D.; Bennett, M.J.; et al. AUX1-mediated root hair auxin influx governs SCFTIR1/AFB-type Ca2+ signaling. Nat. Commun. 2018, 9, 1174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Street, I.H.; Mathews, D.E.; Yamburkenko, M.V.; Sorooshzadeh, A.; John, R.T.; Swarup, R.; Bennett, M.J.; Kieber, J.J.; Schaller, G.E. Cytokinin acts through the auxin influx carrier AUX1 to regulate cell elongation in the root. Development 2016, 143, 3982–3993. [Google Scholar] [CrossRef] [Green Version]
- Schuetz, M.; Fidanza, M.; Mattsson, J. Identification of Auxin response factor-encoding genes expressed in distinct phases of leaf vein development and with overlapping functions in leaf formation. Plants 2019, 8, 242. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.M.; Fei, T.; Zhang, H.X.; Xie, Z.S.; Li, B. Observation of the development of leaf vein and stomata and identification candidate transcription factors related to vein/stoma development in grapevine leaf (Vitis vinifera L.). Sci. Hortic. 2023, 307, 111518. [Google Scholar] [CrossRef]
- Kuang, J.; Wu, J.; Zhong, H.; Li, C.; Chen, J.; Lu, W.; Li, J. Carbohydrate stress affecting fruitlet abscission and expression of genes related to Auxin signal transduction pathway in Litchi. Int. J. Mol. Sci. 2012, 13, 16084–16103. [Google Scholar] [CrossRef]
- Eun, C.H.; Ko, S.M.; Matsubayashi, Y.; Sakagami, Y.; Kamada, H. Phytosulfokine-alpha requires auxin to stimulate carrot non-embryogenic cell proliferation. Plant Physiol. Bioch. 2003, 41, 447–452. [Google Scholar] [CrossRef]
- Niu, J.; Bi, Q.X.; Deng, S.Y.; Chen, H.P.; Yu, H.Y.; Wang, L.B.; Lin, S.Z. Identification of AUXIN RESPONSE FACTOR gene family from Prunus sibirica and its expression analysis during mesocarp and kernel development. BMC Plant Biol. 2018, 18, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geisler, M.; Bailly, A.; Ivanchenko, M. Master and servant: Regulation of auxin transporters by FKBPs and cyclophilins. Plant Sci. 2016, 245, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, T.; Feng, H.; Zhang, S.; Xiao, H.; Hu, Q.; Chen, G.; Xuan, W.; Moran, N.; Murphy, A.; Yu, L.; et al. The potassium transporter OsHAK5 alters rice architecture via ATP-dependent transmembrane Auxin fluxes. Plant Commun. 2020, 1, 100052. [Google Scholar] [CrossRef] [PubMed]
- Bishopp, A.; Benkova, E.; Helariutta, Y. Sending mixed messages: Auxin-cytokinin crosstalk in roots. Curr. Opin. Plant Biol. 2011, 14, 10–16. [Google Scholar] [CrossRef]
- Kurepa, J.; Shull, T.E.; Karunadasa, S.S.; Smalle, J.A. Modulation of auxin and cytokinin responses by early steps of the phenylpropanoid pathway. BMC Plant Biol. 2018, 18, 278. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.J.; Mei, L.H.; Wu, M.B.; Wei, W.; Shan, W.; Gong, Z.H.; Zhang, Q.; Yang, F.Q.; Yan, F.; Zhang, Q.; et al. SlARF10, an auxin response factor, is involved in chlorophyll and sugar accumulation during tomato fruit development. J. Exp. Bot. 2018, 69, 5507–5518. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, W.; Lu, Y.; Luo, S.; Yu, P.; Shen, J.; Wang, X.; Xuan, S.; Wang, Y.; Zhao, J.; Li, N.; et al. Genome-Wide Transcriptome Analysis Reveals That Upregulated Expression of Aux/IAA Genes Is Associated with Defective Leaf Growth of the slf Mutant in Eggplant. Agronomy 2022, 12, 2647. https://doi.org/10.3390/agronomy12112647
Du W, Lu Y, Luo S, Yu P, Shen J, Wang X, Xuan S, Wang Y, Zhao J, Li N, et al. Genome-Wide Transcriptome Analysis Reveals That Upregulated Expression of Aux/IAA Genes Is Associated with Defective Leaf Growth of the slf Mutant in Eggplant. Agronomy. 2022; 12(11):2647. https://doi.org/10.3390/agronomy12112647
Chicago/Turabian StyleDu, Wenchao, Yang Lu, Shuangxia Luo, Ping Yu, Jiajia Shen, Xing Wang, Shuxin Xuan, Yanhua Wang, Jianjun Zhao, Na Li, and et al. 2022. "Genome-Wide Transcriptome Analysis Reveals That Upregulated Expression of Aux/IAA Genes Is Associated with Defective Leaf Growth of the slf Mutant in Eggplant" Agronomy 12, no. 11: 2647. https://doi.org/10.3390/agronomy12112647