
Regulation of Polyethylene Nano-Packaging on Postharvest Stipe Elongation of Flammulina velutipes

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Methods
2.2.1. Sample Processing
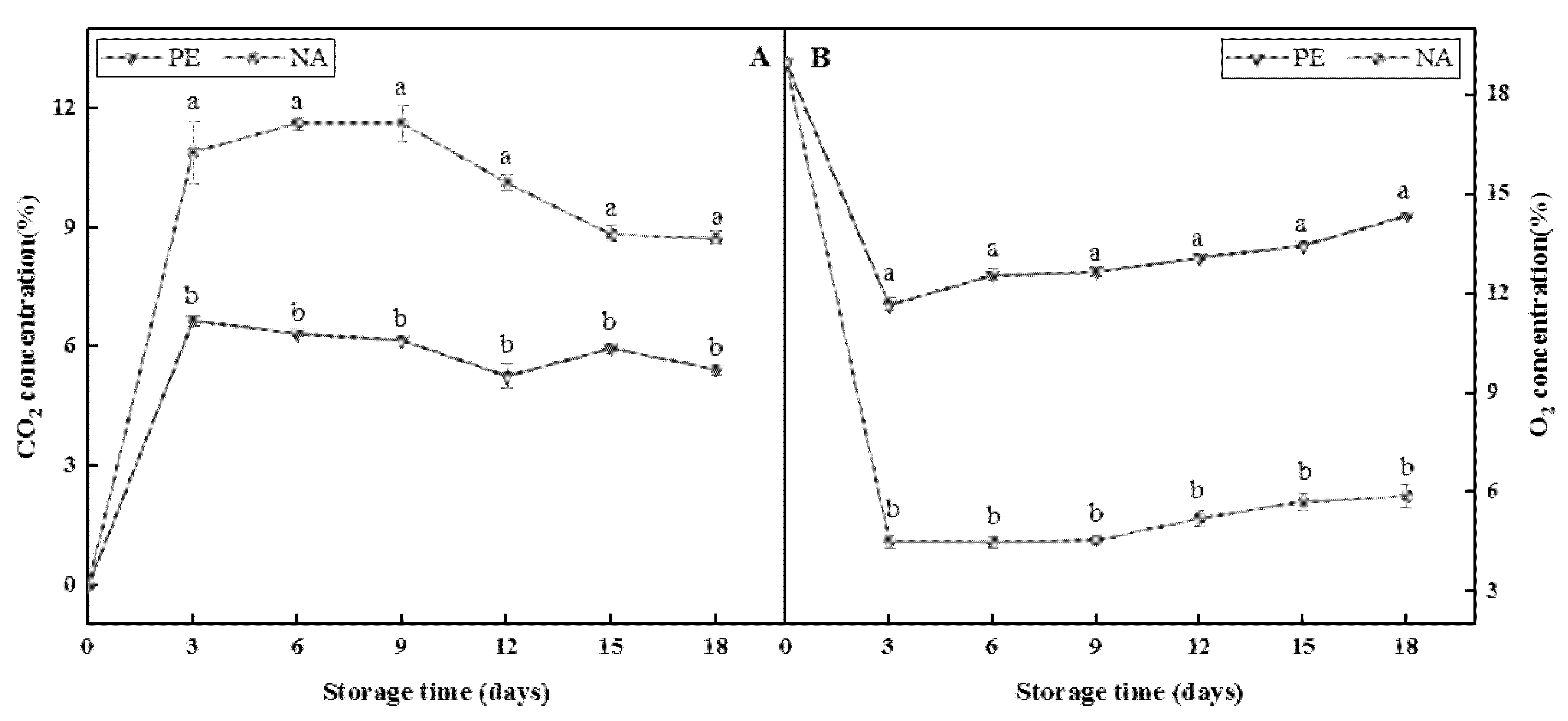
2.2.2. Determination of CO2 Concentrations
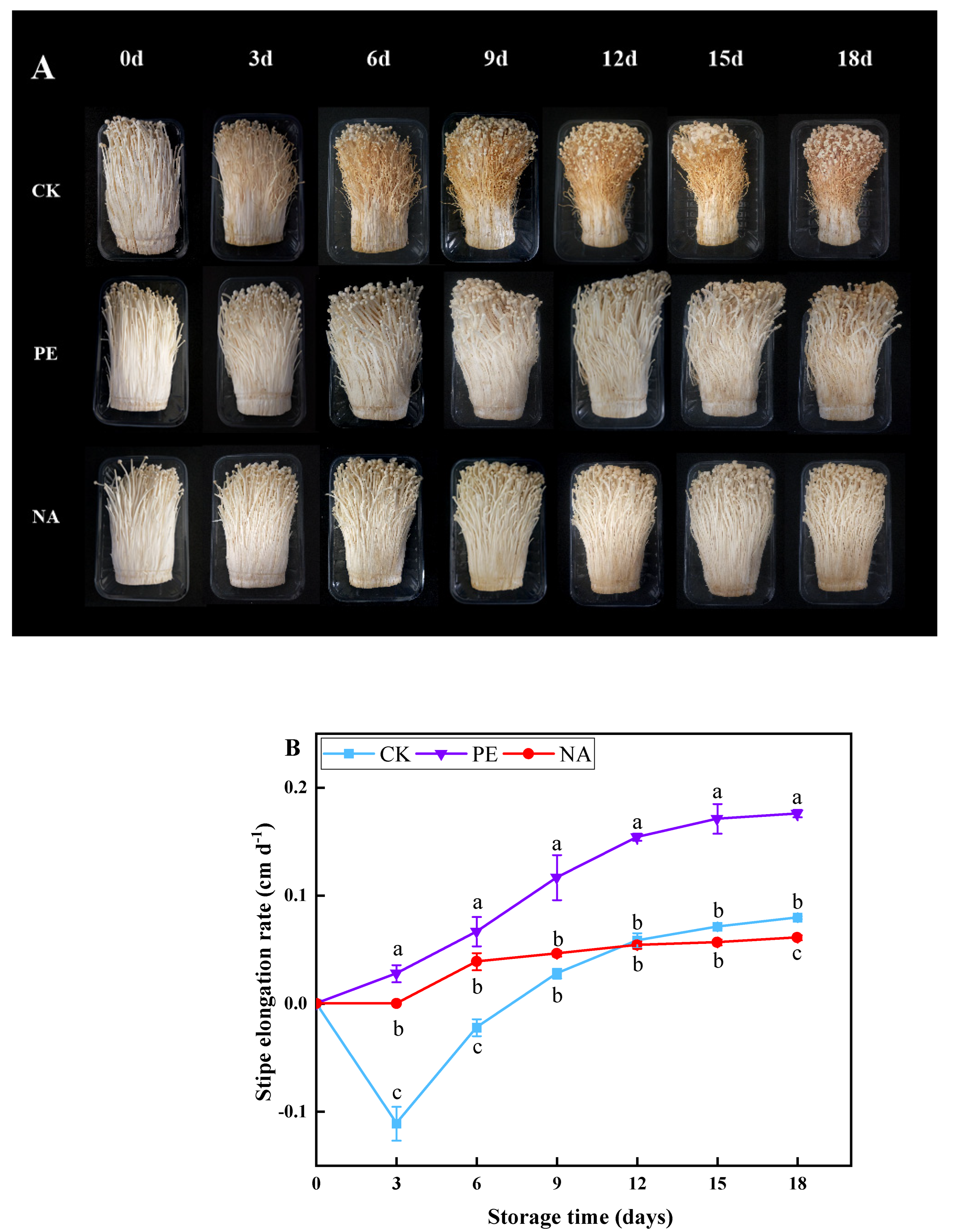
2.2.3. Determination of Stipe Elongation
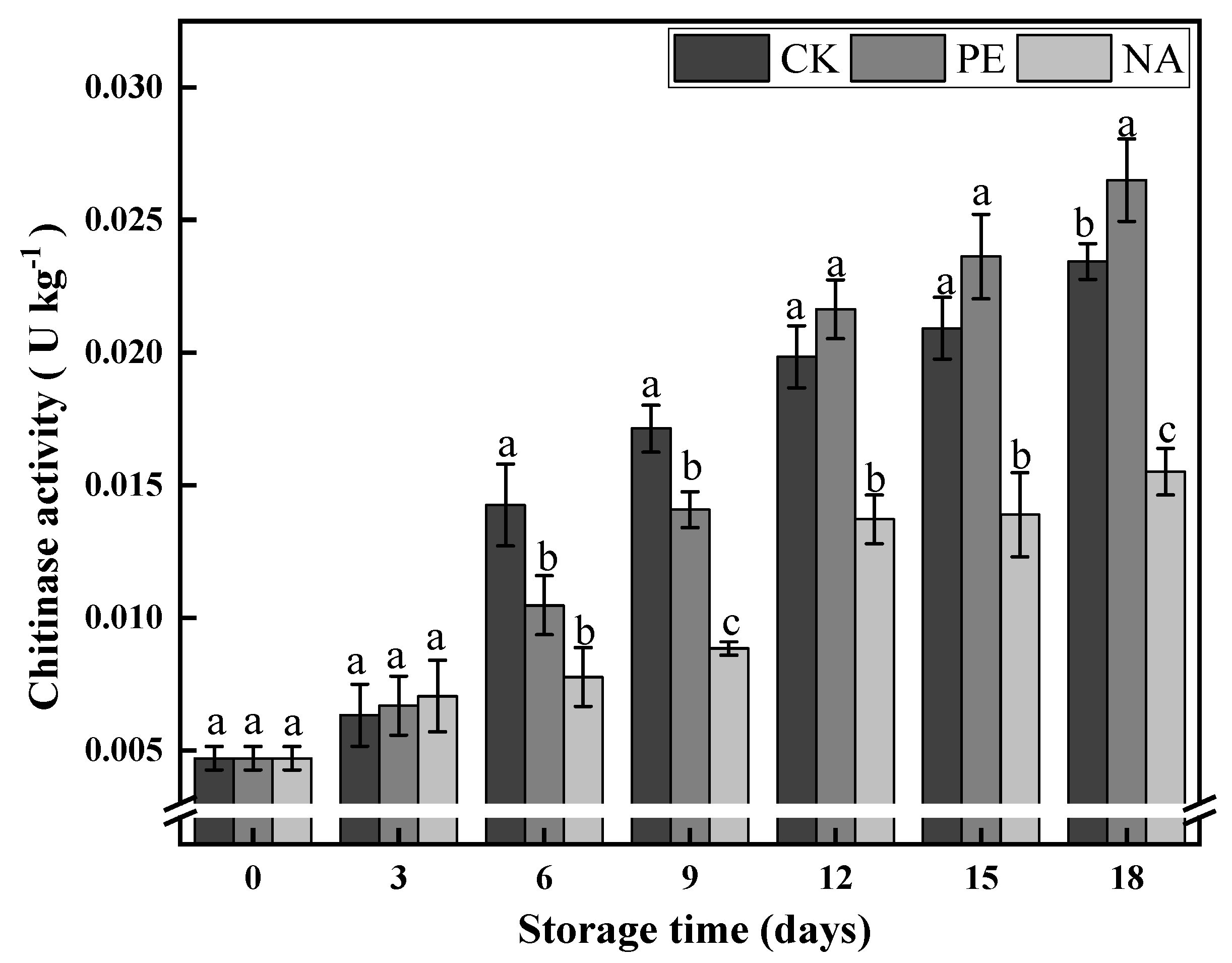
2.2.4. Chitinase Activity Determination
2.2.5. The β-1,3-Glucanase Activity Determination
2.2.6. Determination of Chitin Content
2.2.7. Determination of β-Glucan Content
2.2.8. Determination of Hormones
2.3. Data Analysis
3. Results
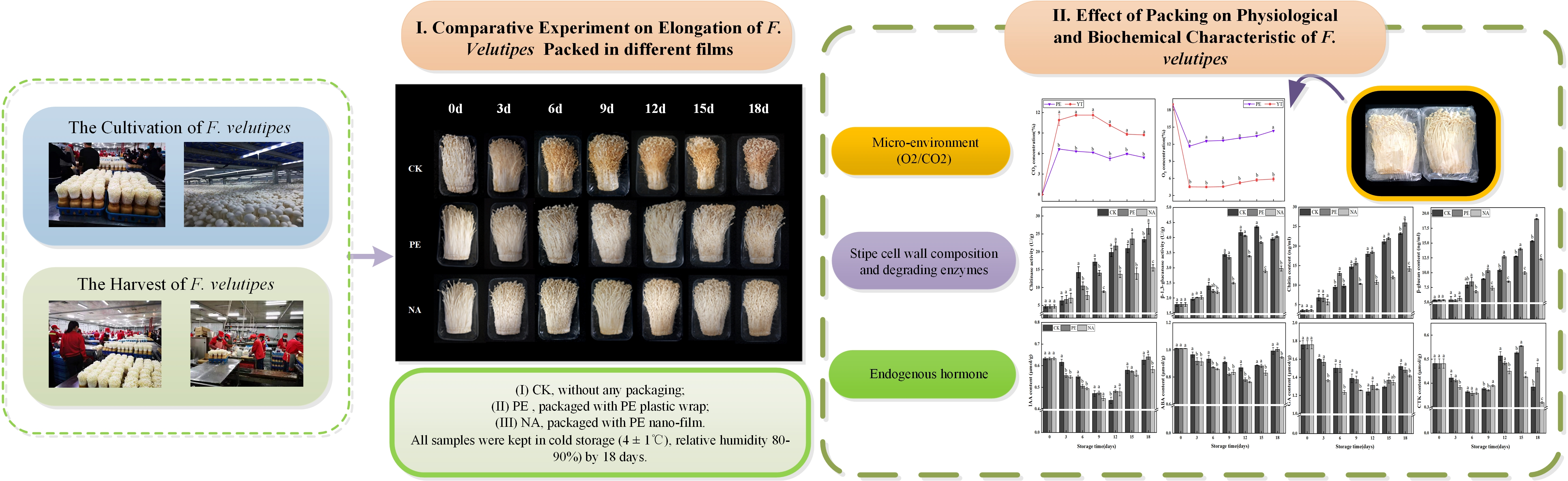
3.1. Changes in Microenvironment Gas in Different Packaging Boxes during Storage
3.2. Effects of Nano-Packaging on the Elongation of F. velutipes Stipes
3.3. Effects of PE Nano-Packaging on the Cell Wall of F. velutipes Stipes
3.3.1. Chitinase Activity
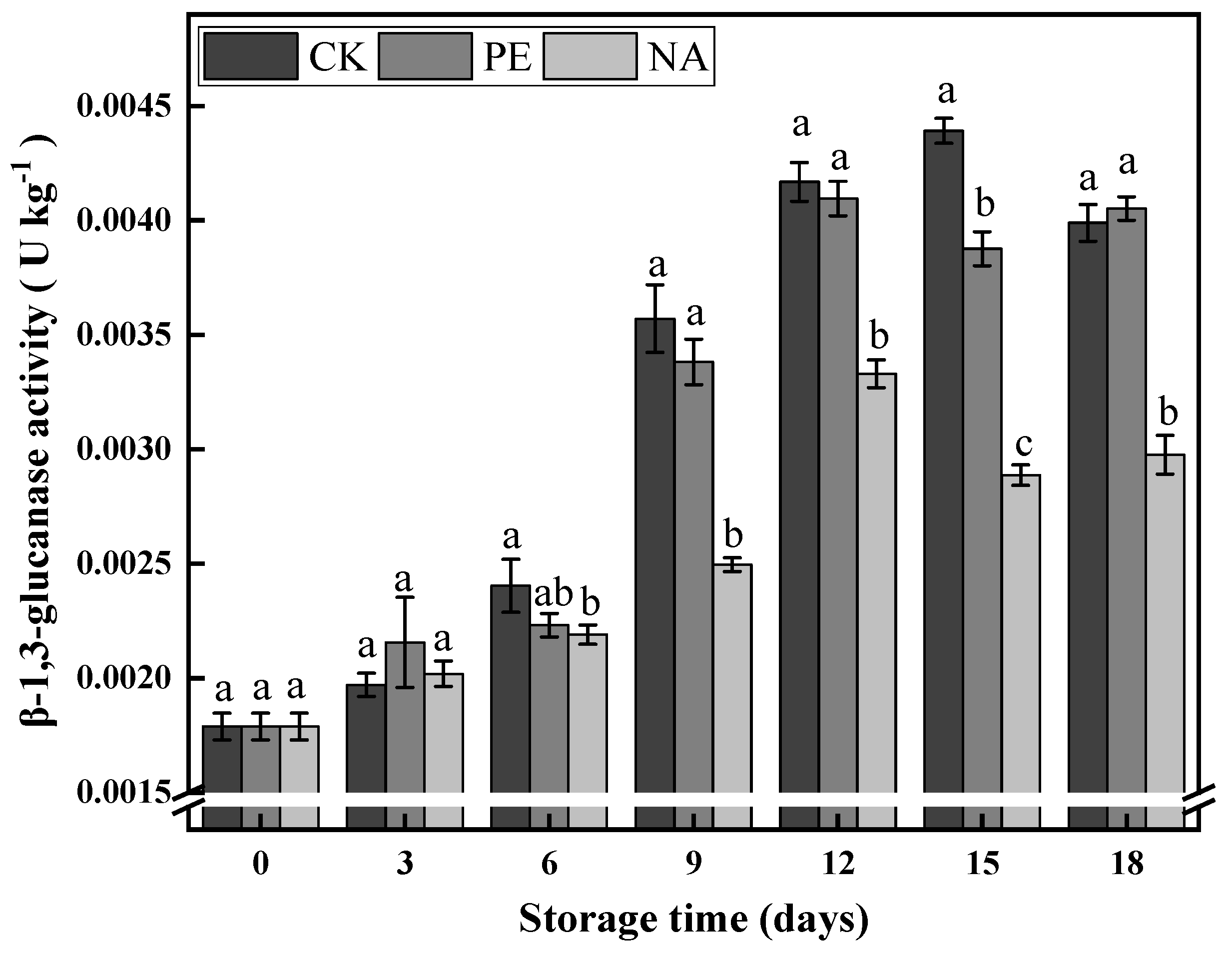
3.3.2. β-1,3-Glucanase Activity
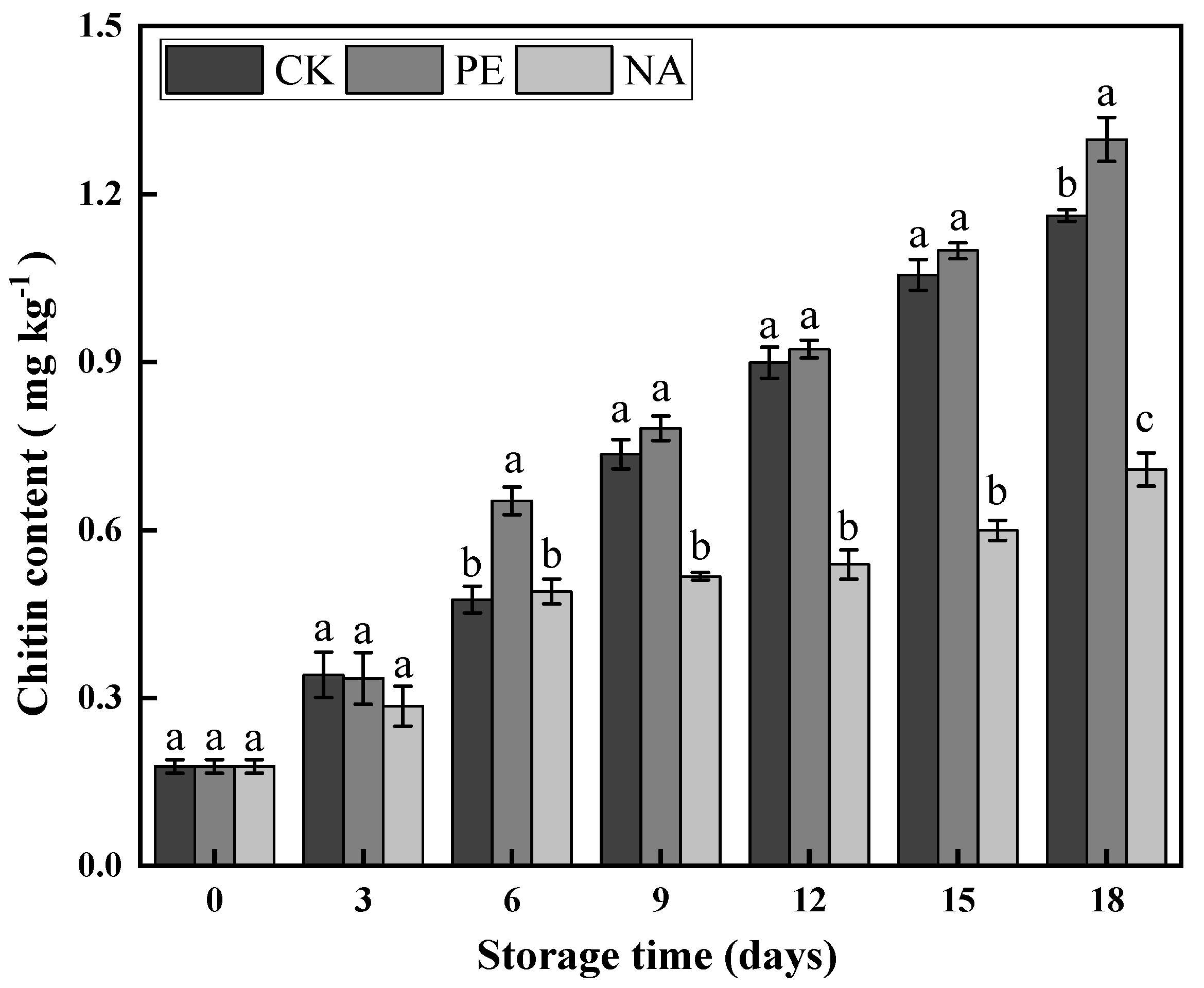
3.3.3. Chitin Content
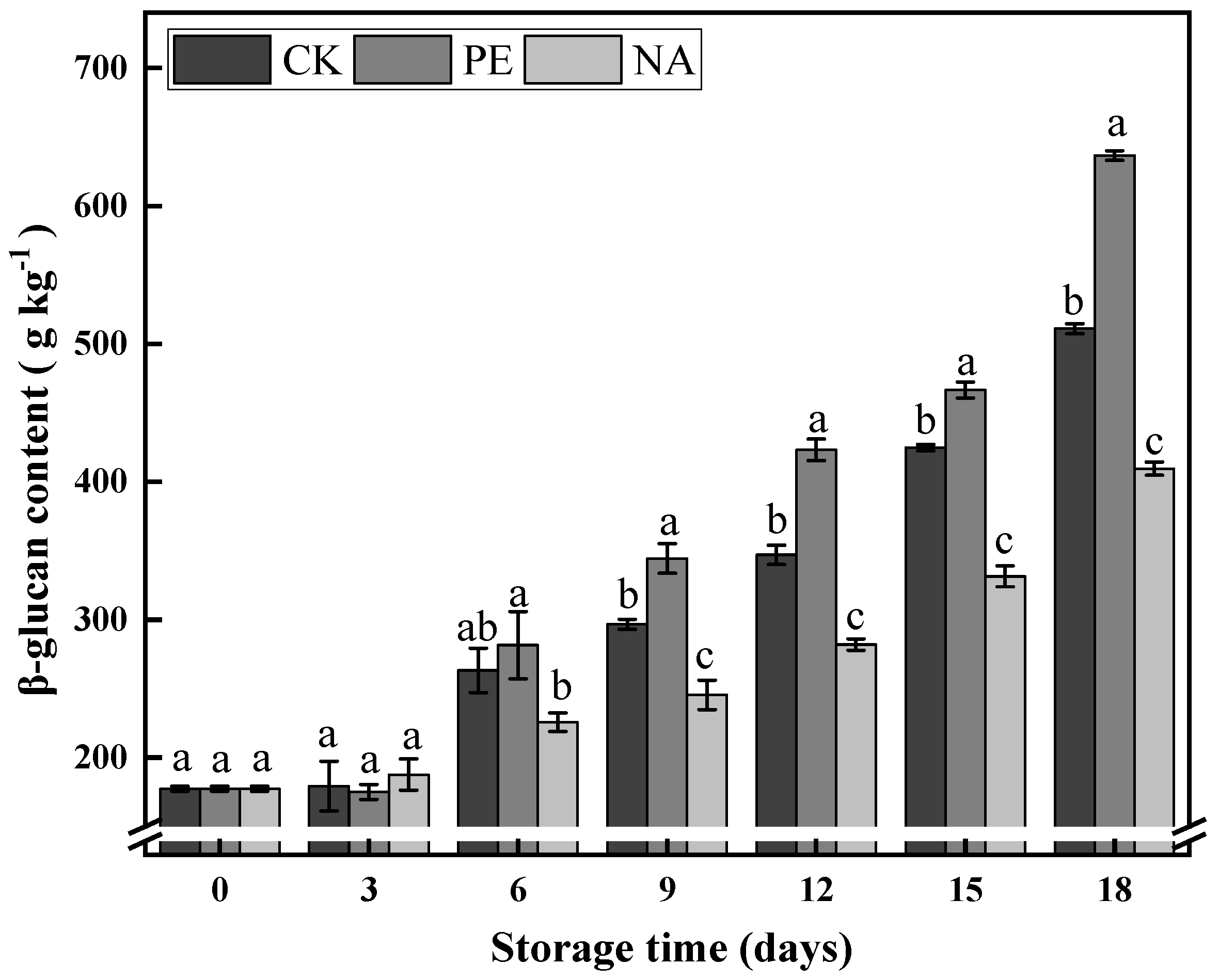
3.3.4. β-Glucan Content
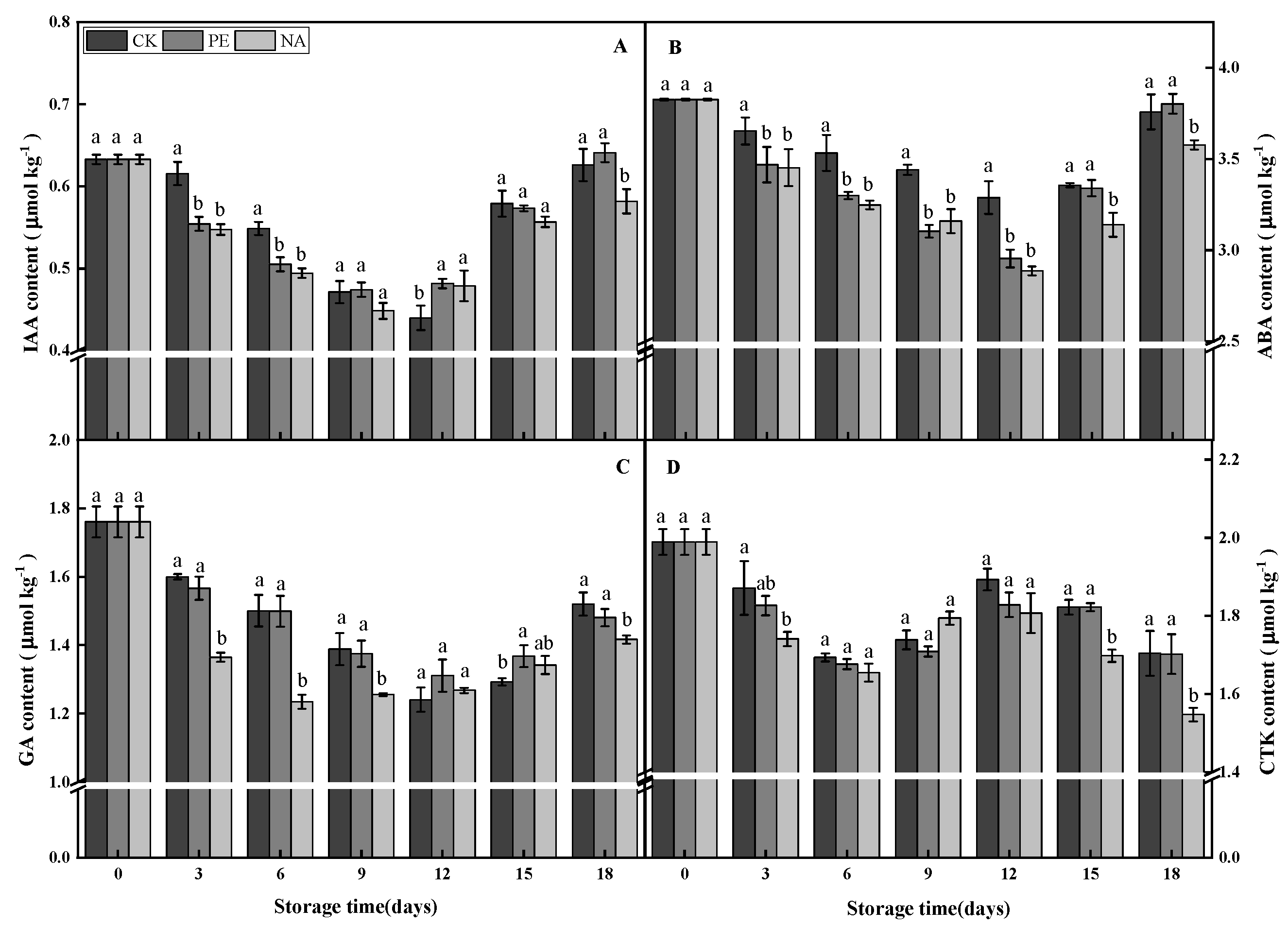
3.4. Effects of Polyethylene Nano-Packaging on Endogenous Hormones in F. velutipes
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jing, P.; Zhao, S.; Lu, M.; Cai, Z.; Pang, J.; Song, L. Multiple-fingerprint analysis for investigating quality control of Flammulina velutipes fruiting body polysaccharides. J. Agric. Food Chem. 2014, 62, 12128–12133. [Google Scholar] [CrossRef]
- Chen, F.; Zhang, Q.; Wu, P.; Zhao, Y.; Suo, X.; Xiao, A.; Ke, M.; He, X.; Tong, Z.; Chen, Y. Green fabrication of seedbed-like Flammulina velutipes polysaccharides–derived scaffolds accelerating full-thickness skin wound healing accompanied by hair follicle regeneration. Int. J. Biol. Macromol. 2021, 167, 117–129. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.; Wu, Y.; Fang, D.; Pei, F.; Mariga, A.M.; Yang, W.; Hu, Q. Effect of nanocomposite packaging on postharvest senescence of Flammulina velutipes. Food Chem. 2018, 246, 414–421. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Yun, J.; Bi, Y.; Wang, T.; Zhang, Y.; Liu, H.; Zhao, F. Predicting the shelf life of postharvest Flammulina velutipes at various temperatures based on mushroom quality and specific spoilage organisms. Postharvest Biol. Technol. 2020, 167, 111235. [Google Scholar] [CrossRef]
- Wang, C.; Wang, C.; Cao, Y.; Robert Nout, M.J.; Sun, B.; Liu, L. Effect of modified atmosphere packaging (MAP) with low and superatmospheric oxygen on the quality and antioxidant enzyme system of golden needle mushrooms (Flammulina velutipes) during postharvest storage. Eur. Food Res. Technol. 2011, 232, 851. [Google Scholar] [CrossRef]
- Xia, R.; Wang, L.; Xin, G.; Bao, X.; Sun, L.; Xu, H.; Hou, Z. Preharvest and postharvest applications of 1-MCP affect umami taste and aroma profiles of mushrooms (Flammulina velutipes). LWT 2021, 144, 111176. [Google Scholar] [CrossRef]
- Chetan, S.; Romika, D.; Namita, R.; Harsh, P. Nanotechnology: An untapped resource for food packaging. Front. Microbiol. 2017, 8, 1735. [Google Scholar] [CrossRef]
- Hu, Q.; Fang, Y.; Yang, Y.; Ma, N.; Zhao, L. Effect of nanocomposite based packaging on postharvest quality of ethylene-treated kiwifruit (Actinidia deliciosa) during cold storage. Food Res. Int. 2011, 44, 1589–1596. [Google Scholar] [CrossRef]
- Duarte, L.G.R.; Picone, C.S.F. Antimicrobial activity of lactoferrin-chitosan-gellan nanoparticles and their influence on strawberry preservation. Food Res. Int. 2022, 159, 111586. [Google Scholar] [CrossRef]
- Ezadi, F.; Rastegar, S. Effect of Nano Packaging on the Quality and Increasing Shelf Life of Pomegranate Arils of Rabab and Mallas Pomegranate (Punica granatum L.). J. Crop Prod. Process. 2019, 9, 41–51. [Google Scholar] [CrossRef] [Green Version]
- Ma, Q.; Cai, M.; Xie, C.; Yang, K.; Sun, P. Evaluation of Polyvinyl Alcohol-Based SiO2/TiO2 Nanocomposite Films and Their Preservation Effect on Agaricus bisporus. Food Sci. 2020, 41, 182–187. [Google Scholar] [CrossRef]
- Yang, W.; Shi, C.; Hu, Q.; Wu, Y.; Fang, D.; Pei, F.; Mariga, A.M. Nanocomposite packaging regulate respiration and energy metabolism in Flammulina velutipes. Postharvest Biol. Technol. 2019, 151, 119–126. [Google Scholar] [CrossRef]
- Zuo, C.; Hu, Q.; Su, A.; Pei, F.; Ma, G.; Xu, H.; Xie, M.; Liu, J.; Mariga, A.M.; Yang, W. Transcriptome analysis reveals the underlying mechanism of nanocomposite packaging in delaying quality deterioration of Flammulina velutipes. Postharvest Biol. Technol. 2021, 182, 111723. [Google Scholar] [CrossRef]
- Zhang, Y. Study on the Regulation of Stipe Elongation and Compound preservation Technology of Flammulina Filiformis after Harvest. Master’s Thesis, Gansu Agricultural University, Lanzhou, China, 2020. [Google Scholar] [CrossRef]
- Kadoo, M.R.; Badere, R.S. Modulation of the activity of chitinases and β-1,3 glucanase in seedlings of cucumber and chilli by the aqueous extract of Cleistanthus collinus. Physiol. Mol. Plant Pathol. 2017, 98, 46–53. [Google Scholar] [CrossRef]
- Kern, K.D.; Nunn, C.; Pichova, A.; Dickinson, J.R. Isoamyl alcohol-induced morphological change in Saccharomyces cerevisiae involves increases in mitochondria and cell wall chitin content. FEMS Yeast Res. 2004, 5, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Du, X.; Rao, Y. Measurement of 3-glucan form oats by congo red. J. Anhui Agric. Univ. 2007, 34, 23–26. [Google Scholar] [CrossRef]
- Meng, D.; Wang, H.; Zhang, Y.; Yang, R.; Fan, Z.; Sheng, J. The production law of phytohormones in different tissue parts of Agaricus bisporus after harvest. J. Chin. Inst. Food Sci. Technol. 2019, 19, 191–198. [Google Scholar] [CrossRef]
- Jin, W.; Gao, H.; Fan, X.; Cheng, W.; Shi, D. Research Progress of Modified Atmosphere Packaging in Edible Fungi Preservation. Storage Process 2017, 17, 126–131. [Google Scholar] [CrossRef]
- Liu, C.; Bi, J.; Kang, L.; Zhou, J.; Liu, X.; Liu, Z.; Yuan, S. The molecular mechanism of stipe cell wall extension for mushroom stipe elongation growth. Fungal Biol. Rev. 2020, 35, 14–26. [Google Scholar] [CrossRef]
- Leyser, O. Auxin signaling. Plant Physiol. 2018, 176, 465–479. [Google Scholar] [CrossRef]
- Zhao, Y. Essential roles of local auxin biosynthesis in plant development and in adaptation to environmental changes. Annu. Rev. Plant Biol. 2018, 29, 417–435. [Google Scholar] [CrossRef] [PubMed]
- Rupak, M.; Sandipan, C.; Bishnu, P.C.; Arun, K.G. Enhancement of biomass production of edible mushroom Pleurotus sajor-caju grown in whey by plant growth hormones. Process Biochem. 2004, 40, 1241–1244. [Google Scholar] [CrossRef]
- Wei, X.; Zhang, X.; Wei, F.; Hong, D.; Ma, J.; Ma, Y. Review of Hormone on Regulation of Cell Elongation. Mod. Agric. Sci. Technol. 2012, 21, 166–167, 170. [Google Scholar] [CrossRef]
- Serpil, Ü.; ŞFatih, T.; Suna, B. Abscisic acid production by pleurotus florida cultured in various conditions and its relation to growth. Isr. J. Plant Sci. 1997, 45, 19–22. [Google Scholar] [CrossRef]
- Xie, B.; Luo, L. Endogenesis Hormone Change in Different Developmental Stage Stipe of Volvariella volvacea. J. Huazhong Agric. Univ. 2004, 23, 64–66. [Google Scholar] [CrossRef]
- Davière, J.M.; Achard, P. Gibberellin signaling in plants. Development 2013, 140, 1147–1151. [Google Scholar] [CrossRef] [PubMed]
- Binenbaum, J.; Weinstain, R.; Shani, E. Gibberellin localization and transport in plants. Trends Plant Sci. 2018, 23, 410–421. [Google Scholar] [CrossRef]
- Ferrero, L.V.; Viola, I.L.; Ariel, F.D.; Gonzalez, D.H. Class I TCP transcription factors target the gibberellin biosynthesis gene GA20ox1 and the growth-promoting genes HBI1 and PRE6 during thermomorphogenic growth in Arabidopsis. Plant Cell Physiol. 2019, 60, 1633–1645. [Google Scholar] [CrossRef]
- Zhu, D.; Wang, C.; Liu, Y.; Winters, E.; Li, W.; Cheng, F. Gibberellic acid maintains postharvest quality of Agaricus bisporus mushroom by enhancing antioxidative system and hydrogen sulfide synthesis. J. Food Biochem. 2021, 45, e13939. [Google Scholar] [CrossRef]
- Vázquez-Chimalhua Ernesto Barrera-Ortiz, S.; Valencia-Cantero, E.; López-Bucio, J.; Ruiz-Herrera, L.F. The bacterial volatile N, N-dimethyl-hexadecylamine promotes Arabidopsis primary root elongation through cytokinin signaling and the AHK2 receptor. Plant Signal. Behav. 2021, 16, 1879542. [Google Scholar] [CrossRef]
- Fang, D. Effects of Nanacomposite Packaging Material on Flammulina velutipes preservation and its possible mechanisms. Ph.D. Thesis, Nanjing Agricultural University, Nanjing, China, 2017. [Google Scholar] [CrossRef]
- Niu, X.; Liu, Z.; Zhou, Y.; Wang, J.; Zhang, W.; Yuan, S. Stipe cell wall architecture varies with the stipe elongation of the mushroom Coprinopsis cinerea. Fungal Biol. 2015, 119, 946–956. [Google Scholar] [CrossRef] [PubMed]
- Fang, H.; Zhang, W.; Niu, X.; Liu, Z.; Lu, C.; Wei, H.; Yuan, S. Stipe wall extension of Flammulina velutipes could be induced by an expansin-like protein from Helix aspersa. Fungal Biol. 2014, 118, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Free, S.J. Fungal cell wall organization and biosynthesis. Adv. Genet. 2013, 81, 33–82. [Google Scholar] [CrossRef] [PubMed]
- Gow NA, R.; Latge, J.P.; Munro, C.A. The fungal cell wall: Structure, biosynthesis, and function. Microbiol. Spectr. 2017, 5, 3. [Google Scholar] [CrossRef]
- Kang, L.; Zhou, J.; Wang, R.; Zhang, X.; Liu, C.; Liu, Z.; Yuan, S. Glucanase-induced stipe wall extension shows distinct differences from chitinase-induced stipe wall extension of Coprinopsis cinerea. Appl. Environ. Microbiol. 2019, 85, e01345-19. [Google Scholar] [CrossRef]
- Zhou, J.; Kang, L.; Liu, C.; Niu, X.; Wang, X.; Liu, H.; Zhang, W.; Liu, Z.; Latge, J.P.; Yuan, S. Chitinases play a key role in stipe cell wall extension in the mushroom Coprinopsis cinerea. Appl. Environ. Microbiol. 2019, 85, e00532-19. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Yang, Q.; Yang, F.; Xin, Z.; Zhao, L.; Yu, Z.; Hu, Q. Effect of Nano-Packaging Material on Quality of Flammulina velutipes. Sci. Agric. Sin. 2009, 42, 3250–3258. [Google Scholar] [CrossRef]
- Wu, Y.; Hu, Q.; Li, Z.; Pei, F.; Mugambi Mariga, A.; Yang, W. Effect of nanocomposite-based packaging on microstructure and energy metabolism of Agaricus bisporus. Food Chem. 2018, 276, 790–796. [Google Scholar] [CrossRef]
- Wang, H. Changes of Nanocomposite Packaging Flammulina filiformis on the Postharvest Mitochondria Energy Metabolism. Master’s Thesis, Nanjing University of Finance and Economics, Nanjing, China, 2021. [Google Scholar] [CrossRef]
- Li, J. Study on the Elongation and Growth of the Stipe Cell Wall of the Fruiting Body of Trichomonas chinensis. Master’s Thesis, Nanjing Normal University, Nanjing, China, 2005. [Google Scholar] [CrossRef]
- Shioya, T.; Nakamura, H.; Ishii, N.; Takahashi, N.; Sakamoto, Y.; Ozaki, N.; Kobayashi, M.; Okano, K.; Kamada, T.; Muraguchi, H. The Coprinopsis cinerea septin Cc.Cdc3 is involved in stipe cell elongation. Fungal Genet. Biol. 2013, 58–59, 80–90. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; Yun, J.; Guo, G.; Li, W.; Wang, B.; Zhao, F.; Bi, Y. Regulation of Polyethylene Nano-Packaging on Postharvest Stipe Elongation of Flammulina velutipes. Agronomy 2022, 12, 2362. https://doi.org/10.3390/agronomy12102362
Zhao Y, Yun J, Guo G, Li W, Wang B, Zhao F, Bi Y. Regulation of Polyethylene Nano-Packaging on Postharvest Stipe Elongation of Flammulina velutipes. Agronomy. 2022; 12(10):2362. https://doi.org/10.3390/agronomy12102362
Chicago/Turabian StyleZhao, Yuxuan, Jianmin Yun, Gengxin Guo, Wenhui Li, Biao Wang, Fengyun Zhao, and Yang Bi. 2022. "Regulation of Polyethylene Nano-Packaging on Postharvest Stipe Elongation of Flammulina velutipes" Agronomy 12, no. 10: 2362. https://doi.org/10.3390/agronomy12102362