
The Effect of the Application of Stimulants on the Photosynthetic Apparatus and the Yield of Winter Wheat

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
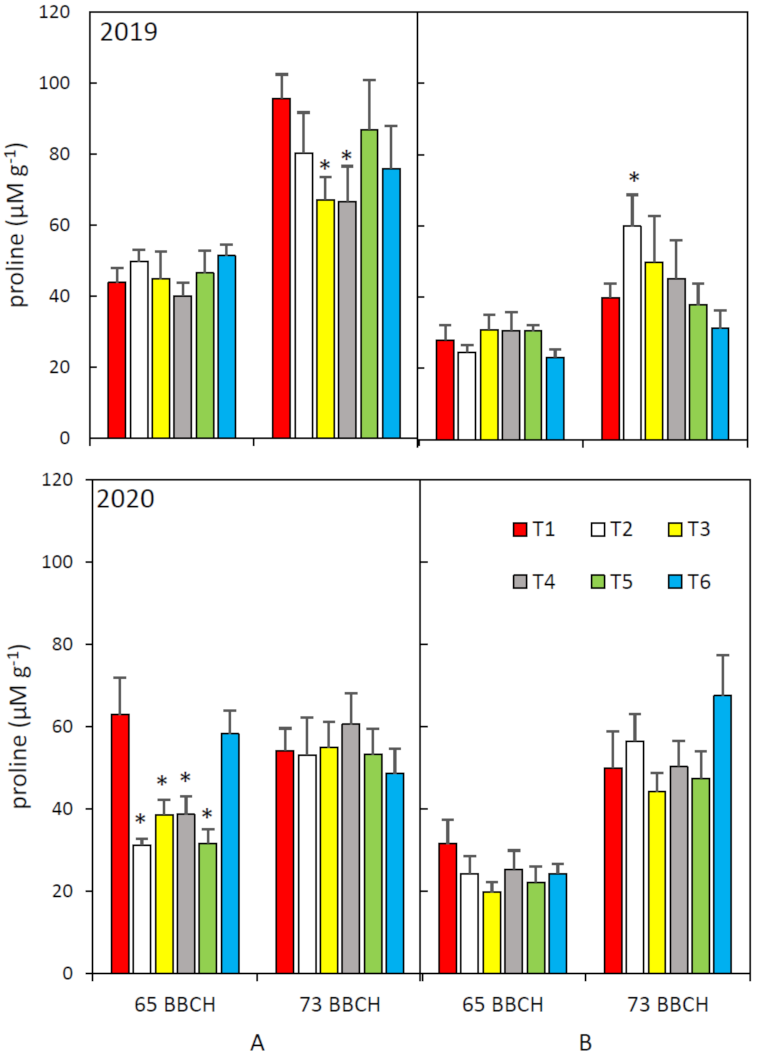
2.2. Photosynthetic Pigments and Free Proline Analyses
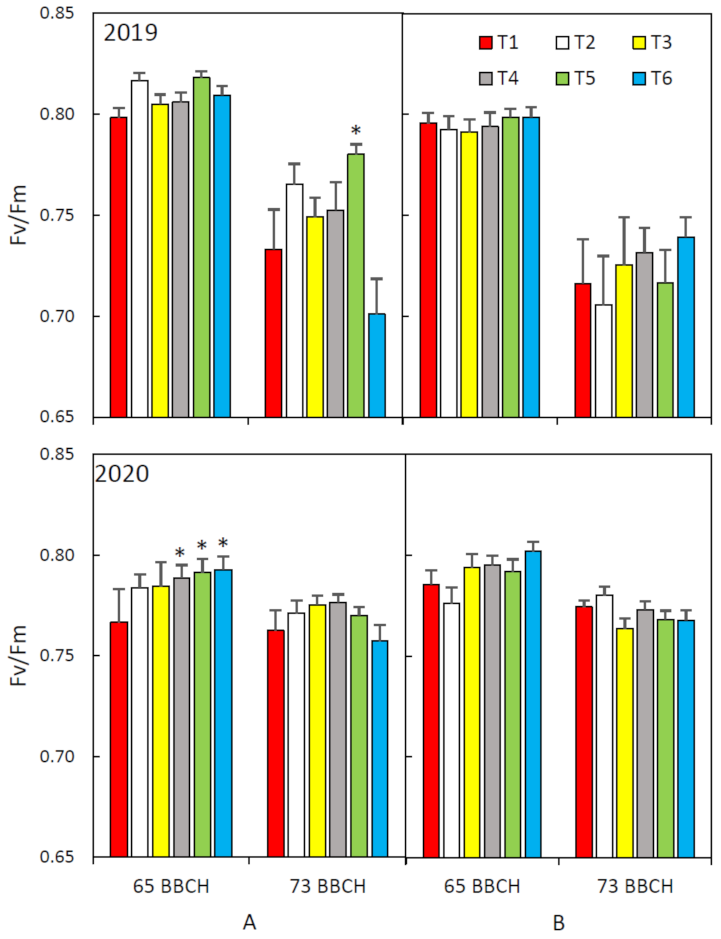
2.3. Chlorophyll Fluorescence
2.4. Analysis of the Yield Components
2.5. Statistical Analysis
3. Results and Discussion
3.1. Characteristics of the Experimental Period 2018–2020 and the State of the Stands
3.2. Photosynthetic Pigments
3.3. Proline Content
3.4. Yield Parameters
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Verma, A.K.; Deepti, S. Abiotic Stress and Crop Improvement: Current Scenario. Adv. Plants Agric. Res. 2016, 4, 345–346. [Google Scholar] [CrossRef] [Green Version]
- Mittler, R. Abiotic Stress, the Field Environment and Stress Combination. Trends Plant Sci. 2006, 11, 15–19. [Google Scholar] [CrossRef]
- Schroeder, E. FAO Forecasts Record Wheat Production in 2021. Available online: https://www.world-grain.com/articles/14970-fao-forecasts-record-wheat-production-in-2021 (accessed on 24 August 2021).
- Raza, A.; Razzaq, A.; Mehmood, S.S.; Zou, X.; Zhang, X.; Lv, Y.; Xu, J. Impact of Climate Change on Crops Adaptation and Strategies to Tackle Its Outcome: A Review. Plants 2019, 8, 34. [Google Scholar] [CrossRef] [Green Version]
- Bulgari, R.; Franzoni, G.; Ferrante, A. Biostimulants Application in Horticultural Crops under Abiotic Stress Conditions. Agronomy 2019, 9, 306. [Google Scholar] [CrossRef] [Green Version]
- Xiong, L.; Schumaker, K.S.; Zhu, J.-K. Cell Signaling during Cold, Drought, and Salt Stress. Plant Cell 2002, 14 (Suppl. 1), 165–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mariani, L.; Ferrante, A. Agronomic Management for Enhancing Plant Tolerance to Abiotic Stresses—Drought, Salinity, Hypoxia, and Lodging. Horticulturae 2017, 3, 52. [Google Scholar] [CrossRef] [Green Version]
- Du Jardin, P. Plant Biostimulants: Definition, Concept, Main Categories and Regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Rouphael, Y.; Colla, G. Synergistic Biostimulatory Action: Designing the Next Generation of Plant Biostimulants for Sustainable Agriculture. Front. Plant Sci. 2018, 9, 1655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahrajabian, M.H.; Chaski, C.; Polyzos, N.; Petropoulos, S.A. Biostimulants Application: A Low Input Cropping Management Tool for Sustainable Farming of Vegetables. Biomolecules 2021, 11, 698. [Google Scholar] [CrossRef]
- Paul, K.; Sorrentino, M.; Lucini, L.; Rouphael, Y.; Cardarelli, M.; Bonini, P.; Reynaud, H.; Canaguier, R.; Trtílek, M.; Panzarová, K.; et al. Understanding the Biostimulant Action of Vegetal-Derived Protein Hydrolysates by High-Throughput Plant Phenotyping and Metabolomics: A Case Study on Tomato. Front. Plant Sci. 2019, 10, 47. [Google Scholar] [CrossRef]
- Biostimulants Market Size Worth $4.14 Billion by 2025|CAGR: 10.2%. Available online: Https://www. Grandviewresearch.Com/Press-Release/Global-Biostimulants-Market (accessed on 8 March 2019).
- Wang, Z.; Yin, Z.; He, M.; Cao, H. Source-Sink Manipulation Effects on Postanthesis Photosynthesis and Grain Setting on Spike in Winter Wheat. Photosynthetica 1998, 35, 453–459. [Google Scholar] [CrossRef]
- Miralles, D.; Slafer, G. Sink Limitations to Yield in Wheat: How Could It Be Reduced? J. Agric. Sci. 2007, 145. [Google Scholar]
- Sreenivasulu, N.; Schnurbusch, T. A Genetic Playground for Enhancing Grain Number in Cereals. Trends Plant Sci. 2012, 17, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, M.; Pellegrineschi, A.; Skovmand, B. Sink-limitation to Yield and Biomass: A Summary of Some Investigations in Spring Wheat. Ann. Appl. Biol. 2005, 146, 39–49. [Google Scholar] [CrossRef]
- Dobrovolskaya, O.; Martinek, P.; Voylokov, A.V.; Korzun, V.; Röder, M.S.; Börner, A. Microsatellite Mapping of Genes That Determine Supernumerary Spikelets in Wheat (T. Aestivum) and Rye (S. Cereale). Theor. Appl. Genet. 2009, 119, 867–874. [Google Scholar] [CrossRef]
- Jiang, W.; Liu, T.; Nan, W.; Jeewani, D.C.; Niu, Y.; Li, C.; Wang, Y.; Shi, X.; Wang, C.; Wang, J.; et al. Two Transcription Factors TaPpm1 and TaPpb1 Co-Regulate Anthocyanin Biosynthesis in Purple Pericarps of Wheat. J. Exp. Bot. 2018, 69, 2555–2567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, N.M.; Carpenter, K.L.; Cannon, J.G. Estimating Contribution of Anthocyanin Pigments to Osmotic Adjustment during Winter Leaf Reddening. J. Plant Physiol. 2013, 170, 230–233. [Google Scholar] [CrossRef]
- Ma, D.; Sun, D.; Wang, C.; Li, Y.; Guo, T. Expression of Flavonoid Biosynthesis Genes and Accumulation of Flavonoid in Wheat Leaves in Response to Drought Stress. Plant Physiol. Biochem. 2014, 80, 60–66. [Google Scholar] [CrossRef]
- Shoeva (Tereshchenko), O.; Gordeeva, E.; Arbuzova, V.; Börner, A.; Khlestkina, E. The D Genome Carries a Gene Determining Purple Grain Colour in Wheat. Cereal Res. Commun. 2012, 40, 334–341. [Google Scholar] [CrossRef]
- Mbarki, S.; Sytar, O.; Zivcak, M.; Abdelly, C.; Cerda, A.; Brestic, M. Anthocyanins of Coloured Wheat Genotypes in Specific Response to SalStress. Molecules 2018, 23, 1518. [Google Scholar] [CrossRef] [Green Version]
- Burešová, V.; Kopecký, D.; Bartoš, J.; Martinek, P.; Watanabe, N.; Vyhnánek, T.; Doležel, J. Variation in Genome Composition of Blue-Aleurone Wheat. Theor. Appl. Genet. 2015, 128, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Dobrovolskaya, O.; Pont, C.; Sibout, R.; Martinek, P.; Badaeva, E.; Murat, F.; Chosson, A.; Watanabe, N.; Prat, E.; Gautier, N.; et al. Frizzy Panicle Drives Supernumerary Spikelets in Bread Wheat. Plant Physiol. 2015, 167, 189–199. [Google Scholar] [CrossRef] [Green Version]
- Rizwan, M.; Mahboob, W.; Faheem, M.; Shimelis, H.; Hameed, A.; Sial, M.A.; Shokat, S. Can We Exploit Supernumerary Spikelet and Spike Branching Traits to Boost Bread Wheat (Triticum Aestivum L.) Yield? Appl. Ecol. Environ. Res. 2020, 18, 6243–6258. [Google Scholar] [CrossRef]
- Meier, U. Growth Stages of Mono- and Dicotyledonous Plants: BBCH Monograph; Julius Kühn-Institut: Quendlinburg, Germany, 2018; ISBN 978-3-95547-071-5. [Google Scholar]
- Porra, R.J.; Thompson, W.A.; Kriedemann, P.E. Determination of Accurate Extinction Coefficients and Simultaneous Equations for Assaying Chlorophylls a and b Extracted with Four Different Solvents: Verification of the Concentration of Chlorophyll Standards by Atomic Absorption Spectroscopy. Biochim. Biophys. Acta 1989, 975, 384–394. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid Determination of Free Proline for Water-Stress Studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Zhao, C.; Liu, B.; Piao, S.; Wang, X.; Lobell, D.B.; Huang, Y.; Huang, M.; Yao, Y.; Bassu, S.; Ciais, P.; et al. Temperature Increase Reduces Global Yields of Major Crops in Four Independent Estimates. Proc. Natl. Acad. Sci. USA 2017, 114, 9326–9331. [Google Scholar] [CrossRef] [Green Version]
- Kyei-Mensah, C.; Kyerematen, R.; Adu-Acheampong, S. Impact of Rainfall Variability on Crop Production within the Worobong Ecological Area of Fanteakwa District, Ghana. Adv. Agric. 2019, 2019, e7930127. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Guan, K.; Schnitkey, G.D.; DeLucia, E.; Peng, B. Excessive Rainfall Leads to Maize Yield Loss of a Comparable Magnitude to Extreme Drought in the United States. Glob. Chang. Biol. 2019, 25, 2325–2337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baslam, M.; Mitsui, T.; Hodges, M.; Priesack, E.; Herritt, M.T.; Aranjuelo, I.; Sanz-Sáez, Á. Photosynthesis in a Changing Global Climate: Scaling Up and Scaling Down in Crops. Front. Plant Sci. 2020, 11, 882. [Google Scholar] [CrossRef]
- Dusenge, M.E.; Duarte, A.G.; Way, D.A. Plant Carbon Metabolism and Climate Change: Elevated CO2 and Temperature Impacts on Photosynthesis, Photorespiration and Respiration. New Phytol. 2019, 221, 32–49. [Google Scholar] [CrossRef] [Green Version]
- Moore, C.E.; Meacham-Hensold, K.; Lemonnier, P.; Slattery, R.A.; Benjamin, C.; Bernacchi, C.J.; Lawson, T.; Cavanagh, A.P. The Effect of Increasing Temperature on Crop Photosynthesis: From Enzymes to Ecosystems. J. Exp. Bot. 2021, 72, 2822–2844. [Google Scholar] [CrossRef]
- Pinheiro, C.; Chaves, M.M. Photosynthesis and Drought: Can We Make Metabolic Connections from Available Data? J. Exp. Bot. 2011, 62, 869–882. [Google Scholar] [CrossRef] [Green Version]
- Santos, E.F.; Zanchim, B.J.; Campos, A.G.; Garrone, R.F.; Lavres Junior, J. Photosynthesis Rate, Chlorophyll Content and Initial Development of Physic Nut without Micronutrient Fertilization. Rev. Bras. Ciênc. Solo 2013, 37, 1334–1342. [Google Scholar] [CrossRef] [Green Version]
- Glanz-Idan, N.; Wolf, S. Upregulation of Photosynthesis in Mineral Nutrition-Deficient Tomato Plants by Reduced Source-to-Sink Ratio. Plant Signal Behav. 2020, 15, 1712543. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.; Harris, P.J.C. Photosynthesis under Stressful Environments: An Overview. Photosynthetica 2013, 51, 163–190. [Google Scholar] [CrossRef]
- Li, Y.; He, N.; Hou, J.; Xu, L.; Liu, C.; Zhang, J.; Wang, Q.; Zhang, X.; Wu, X. Factors Influencing Leaf Chlorophyll Content in Natural Forests at the Biome Scale. Front. Ecol. Evol. 2018, 6, 64. [Google Scholar] [CrossRef] [Green Version]
- Croft, H.; Chen, J.M.; Luo, X.; Bartlett, P.; Chen, B.; Staebler, R.M. Leaf Chlorophyll Content as a Proxy for Leaf Photosynthetic Capacity. Glob. Chang. Biol. 2017, 23, 3513–3524. [Google Scholar] [CrossRef]
- Del Pozo, A.; Méndez-Espinoza, A.M.; Romero-Bravo, S.; Garriga, M.; Estrada, F.; Alcaíno, M.; Camargo-Rodriguez, A.V.; Corke, F.M.K.; Doonan, J.H.; Lobos, G.A. Genotypic Variations in Leaf and Whole-Plant Water Use Efficiencies Are Closely Related in Bread Wheat Genotypes under Well-Watered and Water-Limited Conditions during Grain Filling. Sci. Rep. 2020, 10, 460. [Google Scholar] [CrossRef] [Green Version]
- Gregersen, P.L.; Holm, P.B. Transcriptome Analysis of Senescence in the Flag Leaf of Wheat (Triticum Aestivum L.). Plant Biotechnol. J. 2007, 5, 192–206. [Google Scholar] [CrossRef]
- Liu, B.; Asseng, S.; Wang, A.; Wang, S.; Tang, L.; Cao, W.; Zhu, Y.; Liu, L. Modelling the Effects of Post-Heading Heat Stress on Biomass Growth of Winter Wheat. Agric. Forest Meteorol. 2017, 247, 476–490. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll Fluorescence—A Practical Guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Peripolli, M.; Dornelles, S.H.B.; Lopes, S.J.; Tabaldi, L.A.; Trivisiol, V.S.; Rubert, J. Application of Biostimulants in Tomato Subjected to Water Deficit: Physiological, Enzymatic and Production Responses1. Rev. Bras. Eng. Agríc. Ambient. 2021, 25, 90–95. [Google Scholar] [CrossRef]
- Desotgiu, R.; Pollastrini, M.; Cascio, C.; Gerosa, G.; Marzuoli, R.; Bussotti, F. Chlorophyll a Fluorescence Analysis along a Vertical Gradient of the Crown in a Poplar (Oxford Clone) Subjected to Ozone and Water Stress. Tree Physiol. 2012, 32, 976–986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotakis, C.; Kyzeridou, A.; Manetas, Y. Photosynthetic Electron Flow during Leaf Senescence: Evidence for a Preferential Maintenance of Photosystem I Activity and Increased Cyclic Electron Flow. Photosynthetica 2014, 52, 413–420. [Google Scholar] [CrossRef] [Green Version]
- Kottmann, L.; Wilde, P.; Schittenhelm, S. How Do Timing, Duration, and Intensity of Drought Stress Affect the Agronomic Performance of Winter Rye? Eur. J. Agron. 2016, 75, 25–32. [Google Scholar] [CrossRef]
- Zivcak, M.; Brestic, M.; Olsovska, K. Application of Photosynthetic Parameters in the Screening of Wheat (Triticum Aestivum L.) Genotypes for Improved Drought and High Temperature Tolerance. In Photosynthesis. Energy from the Sun; Allen, J.F., Gantt, E., Golbeck, J.H., Osmond, B., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 1247–1250. ISBN 978-1-4020-6707-5. [Google Scholar]
- Ertani, A.; Nardi, S.; Altissimo, A. Review: Long-Term Research Activity on the Biostimulant Properties of Natural Origin Compounds. Acta Hortic. 2013, 181–187. [Google Scholar] [CrossRef]
- Amanda, A.; Ferrante, A.; Valagussa, M.; Piaggesi, A. Effect of Biostimulants on Quality of Baby Leaf Lettuce Grown under Plastic Tunnel. Acta Hortic. 2009, 407–412. [Google Scholar] [CrossRef]
- Kaur, G.; Asthir, B. Proline: A Key Player in Plant Abiotic Stress Tolerance. Biol. Plant. 2015, 59, 609–619. [Google Scholar] [CrossRef]
- Liang, X.; Zhang, L.; Natarajan, S.K.; Becker, D.F. Proline Mechanisms of Stress Survival. Antioxid. Redox Signal. 2013, 19, 998–1011. [Google Scholar] [CrossRef] [Green Version]
- Nahar, K.; Hasanuzzaman, M.; Fujita, M. Roles of Osmolytes in Plant Adaptation to Drought and Salinity. In Osmolytes and Plants Acclimation to Changing Environment: Emerging Omics Technologies; Springer: New Delhi, India, 2016; pp. 37–68. ISBN 978-81-322-2615-4. [Google Scholar]
- Gould, K.S.; Jay-Allemand, C.; Logan, B.A.; Baissac, Y.; Bidel, L.P.R. When Are Foliar Anthocyanins Useful to Plants? Re-Evaluation of the Photoprotection Hypothesis Using Arabidopsis Thaliana Mutants That Differ in Anthocyanin Accumulation. Environ. Exp. Bot. 2018, 154, 11–22. [Google Scholar] [CrossRef]
- Pietrini, F.; Iannelli, M.A.; Massacci, A. Anthocyanin Accumulation in the Illuminated Surface of Maize Leaves Enhances Protection from Photo-Inhibitory Risks at Low Temperature, without Further Limitation to Photosynthesis. Plant Cell Environ. 2002, 25, 1251–1259. [Google Scholar] [CrossRef]
- Steyn, W.J.; Wand, S.J.E.; Holcroft, D.M.; Jacobs, G. Anthocyanins in Vegetative Tissues: A Proposed Unified Function in Photoprotection. New Phytol. 2002, 155, 349–361. [Google Scholar] [CrossRef]
- Li, X.; Lü, X.; Wang, X.; Peng, Q.; Zhang, M.; Ren, M. Biotic and Abiotic Stress-Responsive Genes Are Stimulated to Resist Drought Stress in Purple Wheat. J. Integr. Agric. 2020, 19, 33–50. [Google Scholar] [CrossRef]
- Kishor, P.B.K.; Sreenivasulu, N. Is Proline Accumulation per Se Correlated with Stress Tolerance or is Proline Homeostasis a More Critical Issue? Plant Cell Environ. 2014, 37, 300–311. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, S.; Funck, D.; Szabados, L.; Rentsch, D. Proline Metabolism and Transport in Plant Development. Amino Acids 2010, 39, 949–962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, X.; Nie, J.; Bai, R.; Sui, X. Amino Acid Transporters in Plants: Identification and Function. Plants 2020, 9, 972. [Google Scholar] [CrossRef]
- Pačuta, V.; Rašovský, M.; Michalska-Klimczak, B.; Wyszyňski, Z. Grain Yield and Quality Traits of Durum Wheat (Triticum Durum Desf.) Treated with Seaweed- and Humic Acid-Based Biostimulants. Agronomy 2021, 11, 1270. [Google Scholar] [CrossRef]
- Van Oosten, M.J.; Pepe, O.; De Pascale, S.; Silletti, S.; Maggio, A. The Role of Biostimulants and Bioeffectors as Alleviators of Abiotic Stress in Crop Plants. Chem. Biol. Technol. Agric. 2017, 4, 5. [Google Scholar] [CrossRef] [Green Version]
- Kocira, S.; Szparaga, A.; Hara, P.; Treder, K.; Findura, P.; Bartoš, P.; Filip, M. Biochemical and Economical Effect of Application Biostimulants Containing Seaweed Extracts and Amino Acids as an Element of Agroecological Management of Bean Cultivation. Sci. Rep. 2020, 10, 17759. [Google Scholar] [CrossRef]
- Balla, K.; Karsai, I.; Bónis, P.; Kiss, T.; Berki, Z.; Horváth, Á.; Mayer, M.; Bencze, S.; Veisz, O. Heat Stress Responses in a Large Set of Winter Wheat Cultivars (Triticum Aestivum L.) Depend on the Timing and Duration of Stress. PLoS ONE 2019, 14, e0222639. [Google Scholar] [CrossRef]
- Beltrano, J.; Ronco, M.G. Improved Tolerance of Wheat Plants (Triticum Aestivum L.) to Drought Stress and Rewatering by the Arbuscular Mycorrhizal Fungus Glomus Claroideum: Effect on Growth and Cell Membrane Stability. Braz. J. Plant Physiol. 2008, 20, 29–37. [Google Scholar] [CrossRef]
- Kukal, M.S.; Irmak, S. Climate-Driven Crop Yield and Yield Variability and Climate Change Impacts on the U.S. Great Plains Agricultural Production. Sci. Rep. 2018, 8, 3450. [Google Scholar] [CrossRef] [Green Version]
- Shew, A.M.; Tack, J.B.; Nalley, L.L.; Chaminuka, P. Yield Reduction under Climate Warming Varies among Wheat Cultivars in South Africa. Nat. Commun. 2020, 11, 4408. [Google Scholar] [CrossRef]
- Liliane, T.N.; Charles, M.S. Factors Affecting Yield of Crops; IntechOpen: London, UK, 2020; ISBN 978-1-83881-223-2. [Google Scholar]
- Ray, D.K.; West, P.C.; Clark, M.; Gerber, J.S.; Prishchepov, A.V.; Chatterjee, S. Climate Change Has Likely already Affected Global Food Production. PLoS ONE 2019, 14, e0217148. [Google Scholar] [CrossRef]
- He, W.; Yang, J.Y.; Qian, B.; Drury, C.F.; Hoogenboom, G.; He, P.; Lapen, D.; Zhou, W. Climate Change Impacts on Crop Yield, Soil Water Balance and Nitrate Leaching in the Semiarid and Humid Regions of Canada. PLoS ONE 2018, 13, e0207370. [Google Scholar] [CrossRef]
- Wing, I.S.; De Cian, E.; Mistry, M.N. Global Vulnerability of Crop Yields to Climate Change. J. Environ. Econ. Manag. 2021, 109, 102462. [Google Scholar] [CrossRef]
- Paleckiene, R.; Sviklas, A.; Šlinkšiene, R. Physicochemical Properties of a Microelement Fertilizer with Amino Acids. Russ. J. Appl. Chem. 2007, 80, 352–357. [Google Scholar] [CrossRef]
- Popko, M.; Michalak, I.; Wilk, R.; Gramza, M.; Chojnacka, K.; Górecki, H. Effect of the New Plant Growth Biostimulants Based on Amino Acids on Yield and Grain Quality of Winter Wheat. Molecules 2018, 23, 470. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, C.; Mrnka, L.; Frantík, T.; Bárnet, M.; Vosátka, M.; Švecová, E. Impact of Protein Hydrolysate Biostimulants on Growth of Barley and Wheat and Their Interaction with Symbionts and Pathogens. Agric. Food Sci. 2020, 29. [Google Scholar] [CrossRef]
- Colla, G.; Hoagland, L.; Ruzzi, M.; Cardarelli, M.; Bonini, P.; Canaguier, R.; Rouphael, Y. Biostimulant Action of Protein Hydrolysates: Unraveling Their Effects on Plant Physiology and Microbiome. Front. Plant Sci. 2017, 8, 2202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, P.; Irulappan, V.; Bagavathiannan, M.V.; Senthil-Kumar, M. Impact of Combined Abiotic and Biotic Stresses on Plant Growth and Avenues for Crop Improvement by Exploiting Physio-Morphological Traits. Front. Plant Sci. 2017, 8, 537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, H.S.S.; Fleming, C.; Selby, C.; Rao, J.R.; Martin, T. Plant Biostimulants: A Review on the Processing of Macroalgae and Use of Extracts for Crop Management to Reduce Abiotic and Biotic Stresses. J. Appl. Phycol. 2014, 26, 465–490. [Google Scholar] [CrossRef]
- Tarakhovskaya, E.R.; Maslov, Y.I.; Shishova, M.F. Phytohormones in Algae. Russ. J. Plant Physiol. 2007, 54, 163–170. [Google Scholar] [CrossRef]
- Kumar, R.; Trivedi, K.; Anand, K.G.V.; Ghosh, A. Science behind Biostimulant Action of Seaweed Extract on Growth and Crop Yield: Insights into Transcriptional Changes in Roots of Maize Treated with Kappaphycus Alvarezii Seaweed Extract under Soil Moisture Stressed Conditions. J. Appl. Phycol. 2020, 32, 599–613. [Google Scholar] [CrossRef]
- Gozzo, F.; Faoro, F. Systemic Acquired Resistance (50 Years after Discovery): Moving from the Lab to the Field. J. Agric. Food Chem. 2013, 61, 12473–12491. [Google Scholar] [CrossRef]
- Calvo, P.; Nelson, L.; Kloepper, J.W. Agricultural Uses of Plant Biostimulants. Plant Soil 2014, 383, 3–41. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Applied Dose of Monitored Stimulants (L ha−1) and the Phenological Phase of the Stand (BBCH) | Preparation Characteristics | |||
|---|---|---|---|---|---|
| BBCH 30 | BBCH 37 | BBCH 49–51 | BBCH 63 | ||
| T1 | - | - | - | - | - |
| T2 | Cleanstorm (0.1) | Cleanstorm (0.1) | Cleanstorm (0.1) | Cleanstorm (0.1) | Free amino acids 12%, combustible substances in dry matter 50% |
| T3 | Energen 3D Plus (0.1) | Energen 3D Plus (0.1) | Energen 3D Plus (0.2) | Energen 3D Plus (0.3) | Free amino acids 13%, combustible substances in dry matter 50% |
| T4 | Atlante-Cu-Prolina (0.6) | - | - | - | K2O 20%, P2O5 30%, Cu 0.5%, free amino acids (L-proline) 2% |
| T5 | Aminocat 30 (0.2) | - | Aminocat 30 (0.2) | - | Free amino acids 30%, total N 3%, P2O5 1%, K2O 1% |
| T6 | Fertileader 2M (2.0) | - | - | - | Seactive complex *, MgO 2%, Mn 11.7% |
| Chlt (mg m−2) | Car (mg m−2) | Fv/Fm | Proline (µM g−1) | Yield (t ha−1) | TKW (g) | |
|---|---|---|---|---|---|---|
| 2019 | 275.1 ± 4.7 a | 45.0 ± 0.7 a | 0.768 ± 0.003 b | 49.2 ± 2.0 a | 8.65 ± 0.14 b | 43.49 ± 0.86 b |
| 2020 | 254.2 ± 3.9 b | 42.7 ± 0.7 b | 0.778 ± 0.002 a | 43.7 ± 1.5 b | 9.37 ± 0.08 a | 46.49 ± 0.53 a |
| 65 BBCH | 326.7 ± 2.9 a | 53.5 ± 0.4 a | 0.793 ± 0.002 a | 35.5 ± 1.2 b | - | - |
| 73 BBCH | 182.9 ± 3.9 b | 31.0 ± 0.6 b | 0.751 ± 0.003 b | 57.6 ± 1.9 a | - | - |
| A | 272.6 ± 5.2 a | 43.8 ± 0.8 a | 0.776 ± 0.003 a | 55.6 ± 1.8 a | 9.42 ± 0.07 a | 48.94 ± 0.23 a |
| B | 262.1 ± 4.1 b | 44.5 ± 0.7 a | 0.769 ± 0.003 b | 37.3 ± 1.5 b | 8.60 ± 0.14 b | 41.05 ± 0.45 b |
| T1 | 267.9 ± 8.8 a | 45.0 ± 1.4 a | 0.764 ± 0.006 b | 50.7 ± 3.2 a | 8.71 ± 0.22 c | 44.23 ± 1.34 b |
| T2 | 263.4 ± 7.2 a | 43.1 ± 1.2 a | 0.774 ± 0.005 ab | 47.3 ± 3.1 a | 9.07 ± 0.23 ab | 45.11 ± 1.39 a |
| T3 | 268.2 ± 8.7 a | 44.0 ± 1.3 a | 0.773 ± 0.004 ab | 43.7 ± 2.8 a | 9.15 ± 0.25 ab | 45.52 ± 1.48 a |
| T4 | 275.3 ± 7.5 a | 45.7 ± 1.1 a | 0.775 ± 0.004 a | 44.7 ± 2.8 a | 9.14 ± 0.23 ab | 45.38 ± 1.26 a |
| T5 | 264.7 ± 8.0 a | 43.9 ± 1.3 a | 0.779 ± 0.004 a | 44.5 ± 3.2 a | 9.18 ± 0.23 a | 44.70 ± 1.28 ab |
| T6 | 263.5 ± 7.9 a | 43.2 ± 1.2 a | 0.771 ± 0.004 ab | 47.6 ± 3.1 a | 8.82 ± 0.16 bc | 45.03 ± 1.27 ab |
| 2019 | 2020 | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Chlt (mg m−2) | Car (mg m−2) | Chlt (mg m−2) | Car (mg m−2) | ||||||
| 65 BBCH | 73BBCH | 65 BBCH | 73BBCH | 65 BBCH | 73BBCH | 65 BBCH | 73BBCH | ||
| A | T1 | 333.1 ± 15.9 b | 156.7 ± 13.4 b | 51.4 ± 2.4 ab | 31.7 ± 2.9 a | 363.0 ± 15.7 a | 206.2 ± 5.9 a | 63.9 ± 2.4 a | 32.8 ± 0.7 a |
| T2 | 350.6 ± 10.5 ab | 181.4 ± 23.0 ab | 54.5 ± 1.5 a | 30.3 ± 3.9 a | 341.4 ± 13.6 a | 184.9 ± 7.6 a | 59.9 ± 2.1 ab | 28.9 ± 1.4 a | |
| T3 | 374.0 ± 13.2 a | 191.0 ± 28.2 a | 56.2 ± 1.8 a | 34.5 ± 4.5 a | 335.6 ± 15.3 ab | 193.3 ± 12.1 a | 59.4 ± 2.0 ab | 29.5 ± 2.1 a | |
| T4 | 353.7 ± 9.9 a | 128.2 ± 21.6 bc | 53.1 ± 1.4 ab | 25.1 ± 3.4 b | 309.2 ± 12.8 b | 208.5 ± 5.6 a | 54.9 ± 1.6 b | 32.9 ± 0.8 a | |
| T5 | 325.9 ± 14.2 b | 164.0 ± 22.6 ab | 49.2 ± 2.0 b | 28.4 ± 4.1 ab | 342.5 ± 15.6 a | 210.8 ± 6.8 a | 60.7 ± 2.4 ab | 33.2 ± 0.9 a | |
| T6 | 338.4 ± 13.8 ab | 111.7 ± 12.4 c | 51.1 ± 1.8 ab | 20.4 ± 2.4 b | 320.6 ± 12.5 b | 203.0 ± 8.1 a | 54.8 ± 3.3 b | 31.3 ± 1.4 a | |
| B | T1 | 332.7 ± 12.0 a | 137.6 ± 21.0 c | 55.4 ± 1.6 a | 26.8 ± 3.6 bc | 310.6 ± 8.2 a | 200.8 ± 6.3 a | 54.8 ± 1.7 a | 31.4 ± 1.0 a |
| T2 | 282.4 ± 8.8 b | 160.9 ± 19.6 b | 44.5 ± 1.7 c | 29.1 ± 3.5 bc | 305.4 ± 8.3 a | 204.8 ± 2.9 a | 53.7 ± 1.3 a | 31.7 ± 0.5 a | |
| T3 | 306.8 ± 12.4 ab | 140.5 ± 16.1 c | 51.8 ± 1.9 ab | 24.5 ± 2.7 c | 284.5 ± 6.1 b | 203.1 ± 3.9 a | 51.6 ± 1.2 ab | 31.0 ± 0.6 a | |
| T4 | 330.7 ± 8.6 a | 206.8 ± 19.4 a | 55.6 ± 1.2 a | 37.3 ± 2.8 ab | 326.5 ± 7.1 a | 198.7 ± 4.2 ab | 56.9 ± 1.3 a | 30.6 ± 0.6 a | |
| T5 | 325.2 ± 12.6 a | 184.0 ± 23.4 ab | 55.9 ± 2.1 a | 34.7 ± 3.1 ab | 285.8 ± 14.5 b | 190.2 ± 4.9 ab | 48.9 ± 1.8 b | 30.3 ± 0.8 a | |
| T6 | 307.8 ± 17.2 ab | 217.3 ± 15.8 a | 50.2 ± 2.5 b | 39.8 ± 2.6 a | 310.2 ± 10.8 a | 183.9 ± 4.4 b | 54.6 ± 1.9 a | 29.2 ± 0.7 a | |
| A | 346.0 ± 5.4 a | 155.5 ± 8.8 a | 52.6 ± 0.7 a | 28.4 ± 1.5 b | 335.4 ± 6.0 a | 201.1 ± 3.3 a | 58.9 ± 1.0 a | 31.4 ± 0.6 a | |
| B | 314.2 ± 5.1 b | 174.6 ± 8.4 a | 52.2 ± 0.8 a | 32.1 ± 1.3 a | 307.2 ± 4.1 b | 200.2 ± 4.3 a | 54.0 ± 0.7 b | 32.0 ± 0.7 a | |
| Treatments | Yield (t ha−1) | TKW (g) | |||
|---|---|---|---|---|---|
| 2019 | 2020 | 2019 | 2020 | ||
| A | T1 | 9.01 ± 0.19 b | 9.51 ± 0.15 a | 47.33 ± 0.38 c | 48.87 ± 0.41 ab |
| T2 | 9.31 ± 0.27 ab | 9.41 ± 0.43 a | 49.00 ± 0.15 ab | 49.27 ± 1.23 ab | |
| T3 | 9.41 ± 0.13 ab | 9.69 ± 0.24 a | 49.67 ± 0.24 a | 50.46 ± 1.09 a | |
| T4 | 9.37 ± 0.09 ab | 9.79 ± 0.05 a | 48.76 ± 0.43 ab | 49.63 ± 0.23 ab | |
| T5 | 9.64 ± 0.11 a | 9.67 ± 0.18 a | 48.53 ± 0.19 b | 48.00 ± 1.45 b | |
| T6 | 9.45 ± 0.09 ab | 8.83 ± 0.09 b | 47.67 ± 0.49 bc | 49.63 ± 0.78 ab | |
| B | T1 | 7.67 ± 0.25 a | 8.65 ± 0.27 b | 37.50 ± 0.40 c | 43.20 ± 0.42 a |
| T2 | 8.02 ± 0.34 a | 9.56 ± 0.15 a | 38.20 ± 0.06 bc | 43.97 ± 0.55 a | |
| T3 | 7.91 ± 0.42 a | 9.58 ± 0.12 a | 38.63 ± 0.30 ab | 43.30 ± 0.35 a | |
| T4 | 7.98 ± 0.26 a | 9.42 ± 0.20 a | 39.3 ± 0.35 a | 43.90 ± 0.31 a | |
| T5 | 7.95 ± 0.37 a | 9.42 ± 0.03 a | 38.57 ± 0.23 ab | 43.71 ± 0.44 a | |
| T6 | 8.09 ± 0.32 a | 8.89 ± 0.04 b | 38.73 ± 0.30 ab | 44.32 ± 0.33 a | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kraus, K.; Hnilickova, H.; Pecka, J.; Lhotska, M.; Bezdickova, A.; Martinek, P.; Kucirkova, L.; Hnilicka, F. The Effect of the Application of Stimulants on the Photosynthetic Apparatus and the Yield of Winter Wheat. Agronomy 2022, 12, 78. https://doi.org/10.3390/agronomy12010078
Kraus K, Hnilickova H, Pecka J, Lhotska M, Bezdickova A, Martinek P, Kucirkova L, Hnilicka F. The Effect of the Application of Stimulants on the Photosynthetic Apparatus and the Yield of Winter Wheat. Agronomy. 2022; 12(1):78. https://doi.org/10.3390/agronomy12010078
Chicago/Turabian StyleKraus, Kamil, Helena Hnilickova, Jan Pecka, Marie Lhotska, Alena Bezdickova, Petr Martinek, Lenka Kucirkova, and Frantisek Hnilicka. 2022. "The Effect of the Application of Stimulants on the Photosynthetic Apparatus and the Yield of Winter Wheat" Agronomy 12, no. 1: 78. https://doi.org/10.3390/agronomy12010078