
Biochemical and Rapid Molecular Analyses to Identify Glyphosate Resistance in Lolium spp.


 ,
,  , ,
, ,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Dose-Response Experiment
2.3. Shikimate Measurement
2.4. RNA Isolation and EPSPS Target-Site Resistance Analyses
2.5. Real-Time Polymerase Chain Reaction (PCR) Experiments
3. Results
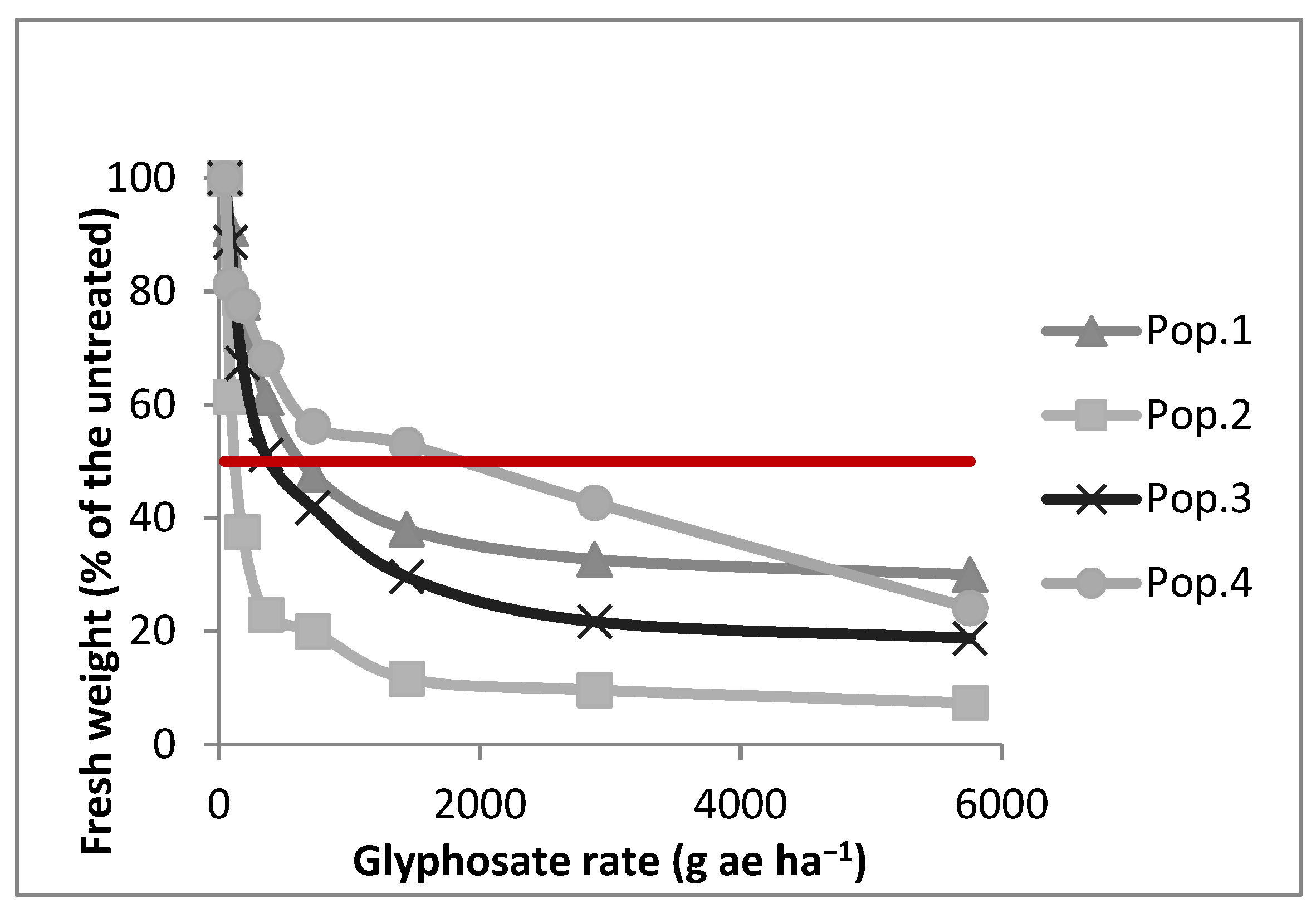
3.1. Dose-Response Experiment
3.2. Shikimate Measurements
3.3. EPSPS cDNA Sequencing
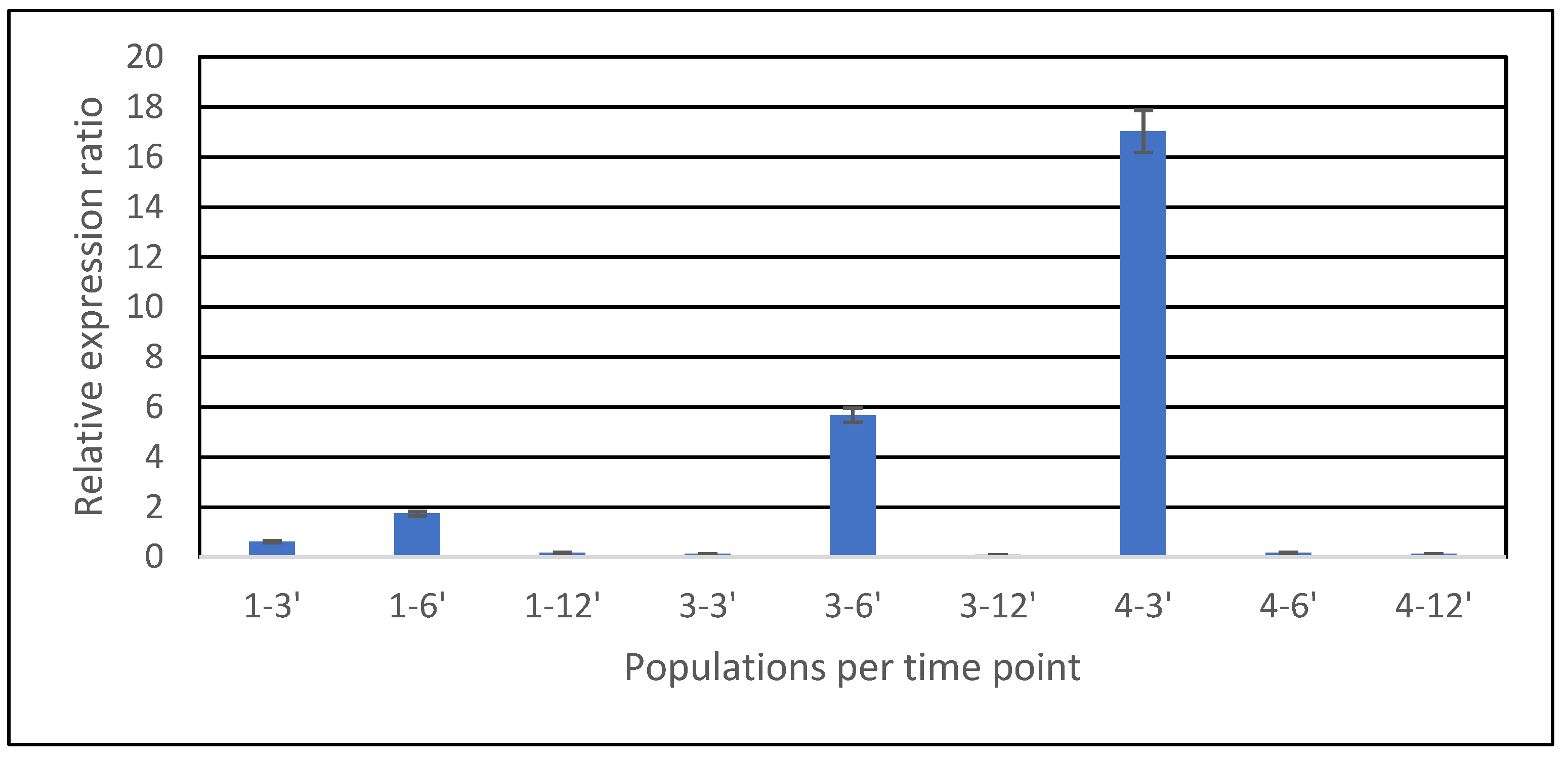
3.4. Analysis of the Expression of ABC Transporter Genes
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Broster, J.; Pratley, J. A decade of monitoring herbicide resistance in Lolium rigidum in Australia. Aust. J. Exp. Agric. 2006, 46, 1151–1160. [Google Scholar] [CrossRef]
- Burnet, M.W.M.; Hart, Q.; Holtum, J.A.M.; Powles, S.B. Resistance to Nine Herbicide Classes in a Population of Rigid Ryegrass (Lolium rigidum). Weed Sci. 1994, 42, 369–377. [Google Scholar] [CrossRef]
- Scarabel, L.; Panozzo, S.; Loddo, D.; Mathiassen, S.K.; Kristensen, M.; Kudsk, P.; Gitsopoulos, T.; Travlos, I.; Tani, E.; Chachalis, D.; et al. Diversified Resistance Mechanisms in Multi-Resistant Lolium spp. in Three European Countries. Front. Plant Sci. 2020, 11, 608845. [Google Scholar] [CrossRef]
- Travlos, I.S.; Cheimona, N.; De Prado, R.; Jhala, A.J.; Chachalis, D.; Tani, E. First Case of Glufosinate-Resistant Rigid Ryegrass (Lolium rigidum Gaud.) in Greece. Agronomy 2018, 8, 35. [Google Scholar] [CrossRef] [Green Version]
- Schönbrunn, E.; Eschenburg, S.; Shuttleworth, W.A.; Schloss, J.V.; Amrhein, N.; Evans, J.N.; Kabsch, W. Interaction of the herbicide glyphosate with its target enzyme 5-enolpyruvylshikimate 3-phosphate synthase in atomic detail. Proc. Natl. Acad. Sci. USA 2001, 98, 1376–1380. [Google Scholar] [CrossRef] [Green Version]
- Powles, S.B.; Lorraine-Colwill, D.F.; Dellow, J.J.; Preston, C. Evolved resistance to glyphosate in rigid ryegrass (Lolium rigidum) in Australia. Weed Sci. 1998, 46, 604–607. [Google Scholar] [CrossRef]
- Powles, S.B. Evolved glyphosate-resistant weeds around the world: Lessons to be learnt. Pest Manag. Sci 2008, 64, 360–365. [Google Scholar] [CrossRef]
- Heap, I. The International Survey of Herbicide Resistant Weeds. Available online: http://www.weedscience.org/Home.aspx (accessed on 1 November 2021).
- Koger, C.H.; Reddy, K.N. Role of absorption and translocation in the mechanism of glyphosate resistance in horseweed Conyza canadensis). Weed Sci. 2005, 53, 84–89. [Google Scholar] [CrossRef]
- Dayan, F.E.; Owens, D.K.; Corniani, N.; Silva, F.M.L.; Watson, S.B.; Howell, J.L.; Shaner, D.L. Biochemical Markers and Enzyme Assays for Herbicide Mode of Action and Resistance Studies. Weed Sci. 2015, 63, 23–63. [Google Scholar] [CrossRef] [Green Version]
- Sammons, R.D.; Gaines, T.A. Glyphosate resistance: State of knowledge. Pest Manag. Sci. 2014, 70, 1367–1377. [Google Scholar] [CrossRef] [Green Version]
- Jasieniuk, M.; Ahmad, R.; Sherwood, A.M.; Firestone, J.L.; Perez-Jones, A.; Lanini, W.T.; Mallory-Smith, C.; Stednick, Z. Glyphosate-Resistant Italian Ryegrass (Lolium multiflorum) in California: Distribution, Response to Glyphosate, and Molecular Evidence for an Altered Target Enzyme. Weed Sci. 2008, 56, 496–502. [Google Scholar] [CrossRef]
- Gherekhloo, J.; Fernández-Moreno, P.T.; Alcántara-de la Cruz, R.; Sánchez-González, E.; Cruz-Hipolito, H.E.; Domínguez-Valenzuela, J.A.; De Prado, R. Pro-106-Ser mutation and EPSPS overexpression acting together simultaneously in glyphosate-resistant goosegrass (Eleusine indica). Sci. Rep. 2017, 7, 6702. [Google Scholar] [CrossRef] [PubMed]
- Perez-Jones, A.; Park, K.W.; Polge, N.; Colquhoun, J.; Mallory-Smith, C.A. Investigating the mechanisms of glyphosate resistance in Lolium multiflorum. Planta 2007, 226, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Powles, S.B.; Preston, C. Evolved Glyphosate Resistance in Plants: Biochemical and Genetic Basis of Resistance. Weed Technol. 2006, 20, 282–289. [Google Scholar] [CrossRef]
- Yu, Q.; Cairns, A.; Powles, S. Glyphosate, paraquat and ACCase multiple herbicide resistance evolved in a Lolium rigidum biotype. Planta 2007, 225, 499–513. [Google Scholar] [CrossRef]
- De Prado, J.L.; Osuna, M.D.; Heredia, A.; De Prado, R. Lolium rigidum, a pool of resistance mechanisms to ACCase inhibitor herbicides. J. Agric. Food Chem. 2005, 53, 2185–2191. [Google Scholar] [CrossRef]
- Vila-Aiub, M.M.; Gundel, P.E.; Yu, Q.; Powles, S.B. Glyphosate resistance in Sorghum halepense and Lolium rigidum is reduced at suboptimal growing temperatures. Pest Manag. Sci. 2013, 69, 228–232. [Google Scholar] [CrossRef]
- Ge, X.; d’Avignon, D.A.; Ackerman, J.J.H.; Sammons, R.D. Rapid vacuolar sequestration: The horseweed glyphosate resistance mechanism. Pest Manag. Sci. 2010, 66, 345–348. [Google Scholar] [CrossRef] [Green Version]
- de Carvalho, L.B.; Alves, P.L.d.C.A.; González-Torralva, F.; Cruz-Hipolito, H.E.; Rojano-Delgado, A.M.; De Prado, R.; Gil-Humanes, J.; Barro, F.; Luque de Castro, M.D. Pool of Resistance Mechanisms to Glyphosate in Digitaria insularis. J. Agric. Food Chem. 2012, 60, 615–622. [Google Scholar] [CrossRef] [PubMed]
- Bai, S.; Liu, W.; Wang, H.; Zhao, N.; Jia, S.; Zou, N.; Guo, W.; Wang, J. Enhanced Herbicide Metabolism and Metabolic Resistance Genes Identified in Tribenuron-Methyl Resistant Myosoton aquaticum L. J. Agric. Food Chem. 2018, 66, 9850–9857. [Google Scholar] [CrossRef]
- Giacomini, D.A.; Patterson, E.L.; Küpper, A.; Beffa, R.; Gaines, T.A.; Tranel, P.J. Coexpression Clusters and Allele-Specific Expression in Metabolism-Based Herbicide Resistance. Genome Biol. Evol. 2020, 12, 2267–2278. [Google Scholar] [CrossRef] [PubMed]
- Gil, M.; Vega, T.; Felitti, S.; Picardi, L.; Balzergue, S.; Nestares, G. Characterization of Non-Target-Site Mechanisms in Imidazolinone-Resistant Sunflower by RNA-seq. Helia 2018, 41, 267–278. [Google Scholar] [CrossRef]
- Ghanizadeh, H.; Harrington, K.C. Non-target Site Mechanisms of Resistance to Herbicides. Crit. Rev. Plant Sci. 2017, 36, 24–34. [Google Scholar] [CrossRef]
- Peng, Y.; Abercrombie, L.; Yuan, J.; Riggins, C.; Sammons, R.; Tranel, P.; Stewart, C. Characterization of the horseweed (Conyza canadensis) transcriptome using GS-FLX 454 pyrosequencing and its application for expression analysis of candidate non-target herbicide resistance genes. Pest Manag. Sci. 2010, 66, 1053–1062. [Google Scholar] [CrossRef]
- Yuan, J.S.; Abercrombie, L.L.G.; Cao, Y.; Halfhill, M.D.; Zhou, X.; Peng, Y.; Hu, J.; Rao, M.R.; Heck, G.R.; Larosa, T.J.; et al. Functional Genomics Analysis of Horseweed (Conyza canadensis) with Special Reference to the Evolution of Non–Target-Site Glyphosate Resistance. Weed Sci. 2010, 58, 109–117. [Google Scholar] [CrossRef]
- Tani, E.; Chachalis, D.; Travlos, I.S. A Glyphosate Resistance Mechanism in Conyza canadensis Involves Synchronization of EPSPS and ABC-transporter Genes. Plant Mol. Biol. Report. 2015, 33, 1721–1730. [Google Scholar] [CrossRef]
- Pan, L.; Yu, Q.; Wang, J.; Han, H.; Mao, L.; Nyporko, A.; Maguza, A.; Fan, L.; Bai, L.; Powles, S. An ABCC-type transporter endowing glyphosate resistance in plants. Proc. Natl. Acad. Sci. USA 2021, 118, e2100136118. [Google Scholar] [CrossRef]
- Ghanizadeh, H.; Harrington, K.C. Perspective: Root exudation of herbicides as a novel mode of herbicide resistance in weeds. Pest Manag. Sci. 2020, 76, 2543–2547. [Google Scholar] [CrossRef]
- Cechin, J.; Piasecki, C.; Benemann, D.P.; Kremer, F.S.; Galli, V.; Maia, L.C.; Agostinetto, D.; Vargas, L. Transcriptome Analysis Identifies Candidate Target Genes Involved in Glyphosate-Resistance Mechanism in Lolium multiflorum. Plants 2020, 9, 685. [Google Scholar] [CrossRef]
- Seefeldt, S.S.; Jensen, J.E.; Fuerst, E.P. Log-Logistic Analysis of Herbicide Dose-Response Relationships. Weed Technol. 1995, 9, 218–227. [Google Scholar] [CrossRef]
- Travlos, I.; Tabaxi, E.; Papadimitriou, D.; Bilalis, D.; Chachalis, D. Lolium rigidum Gaud. biotypes from Greece with Resistance to Glyphosate and other Herbicides. Bull. Univ. Agric. Sci. Vet. Med. Cluj-Napoca. Hortic. 2016, 73, 1–2. [Google Scholar] [CrossRef] [Green Version]
- Kaundun, S.S.; Dale, R.P.; Zelaya, I.A.; Dinelli, G.; Marotti, I.; McIndoe, E.; Cairns, A. A Novel P106L Mutation in EPSPS and an Unknown Mechanism(s) Act Additively To Confer Resistance to Glyphosate in a South African Lolium rigidum Population. J. Agric. Food Chem. 2011, 59, 3227–3233. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Hu, T.; Fu, J. Identification of genes associated with adaptation to NaCl toxicity in perennial ryegrass (Lolium perenne L.). Ecotoxicol. Environ. Saf. 2012, 79, 153–162. [Google Scholar] [CrossRef]
- Salas, R.A.; Dayan, F.E.; Pan, Z.; Watson, S.B.; Dickson, J.W.; Scott, R.C.; Burgos, N.R. EPSPS gene amplification in glyphosate-resistant Italian ryegrass (Lolium perenne ssp. multiflorum) from Arkansas. Pest Manag. Sci. 2012, 68, 1223–1230. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Suzukawa, A.K.; Bobadilla, L.K.; Mallory-Smith, C.; Brunharo, C.A.C.G. Non-target-Site Resistance in Lolium spp. Globally: A Review. Front. Plant Sci. 2021, 11, 2137. [Google Scholar] [CrossRef]
- Panozzo, S.; Collavo, A.; Sattin, M. Sensitivity Analysis of Italian Lolium spp. to Glyphosate in Agricultural Environments. Plants 2020, 9, 165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tehranchian, P.; Nandula, V.; Jugulam, M.; Putta, K.; Jasieniuk, M. Multiple resistance to glyphosate, paraquat and ACCase-inhibiting herbicides in Italian ryegrass populations from California: Confirmation and mechanisms of resistance. Pest Manag. Sci. 2018, 74, 868–877. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, D.; Gil, D.; Cruz-Hipolito, H.; Ruiz-Santaella, J.P.; Christoffoleti, P.; Vidal, R.A.; Prado, R. Rapid assays for detection of glyphosate-resistant Lolium spp. J. Plant Dis. Proctection Suppl. 2008, 2, 95–99. [Google Scholar]
- Yanniccari, M.; Istilart, C.; Giménez, D.O.; Castro, A.M. Glyphosate resistance in perennial ryegrass (Lolium perenne L.) from Argentina. Crop Prot. 2012, 32, 12–16. [Google Scholar] [CrossRef]
- Yu, Q.; Abdallah, I.; Han, H.; Owen, M.; Powles, S. Distinct non-target site mechanisms endow resistance to glyphosate, ACCase and ALS-inhibiting herbicides in multiple herbicide-resistant Lolium rigidum. Planta 2009, 230, 713–723. [Google Scholar] [CrossRef] [PubMed]
- González-Torralva, F.; Gil-Humanes, J.; Barro, F.; Brants, I.; De Prado, R. Target site mutation and reduced translocation are present in a glyphosate-resistant Lolium multiflorum Lam. biotype from Spain. Plant Physiol. Biochem. PPB 2012, 58, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Karn, E.; Jasieniuk, M. Nucleotide Diversity at Site 106 of EPSPS in Lolium perenne L. ssp. multiflorum from California Indicates Multiple Evolutionary Origins of Herbicide Resistance. Front. Plant Sci. 2017, 8, 777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández-Moreno, P.T.; Travlos, I.; Brants, I.; De Prado, R. Different levels of glyphosate-resistant Lolium rigidum L. among major crops in southern Spain and France. Sci. Rep. 2017, 7, 13116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moretti, M.L.; Alárcon-Reverte, R.; Pearce, S.; Morran, S.; Hanson, B.D. Transcription of putative tonoplast transporters in response to glyphosate and paraquat stress in Conyza bonariensis and Conyza canadensis and selection of reference genes for qRT-PCR. PLoS ONE 2017, 12, e0180794. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population Code | Crop | Area |
|---|---|---|
| pop.1 | Olives | western Greece |
| pop.2 | barley | central Greece |
| pop.3 | Vineyard | central Greece |
| pop.4 | cereal stubble in no-till fields | Italy |
| Primer Name | Primer Sequence |
|---|---|
| LpABCIII-F | 5′-AGAGCTGCAAAGGCTGGTAG-3′ |
| LpABCIII-R | 5′-TCTAAGCGGAAGCAAAGCCA-3′ |
| Lpmultidrugprotein-F | 5′-GGTCATGGACTGCGACAGAG-3′ |
| Lpmultidrugprotein-R | 5′-CACGTCAGATGACCGGTTTG-3′ |
| LpM10-F | 5′-TATGTTGTGGCTGACACGCT-3′ |
| LpM10-R | 5′-ATCGGCGTTGTGCAAGAAAT-3′ |
| LpironinhibitedABC-F | 5′-TAAACTCCCACCACCAGTGC-3′ |
| LpironinhibitedABC-R | 5′-TCACCGGTCATGAGCTTCAG-3′ |
| Reference gene LpCCR-F | 5′-GATGTCGAACCAGAAGCTCCA-3′ |
| Reference gene LpCCR-R | 5′-GCAGCTAGGGTTTCCTTGTCC-3′ |
| Populations | ||||
|---|---|---|---|---|
| Pop.1 | Pop.2 | Pop.3 | Pop.4 | |
| LD50 | >5760 (60.18) | 270 (10.04) | >5760 (49.94) | 2850 (33.82) |
| LD90 | >5760 (40.32) | 1580 (31.53) | >5760 (55.20) | >5760 (39.21) |
| GR50 | 490 (19.05) | 126 (5.87) | 360 (13.08) | 740 (13.45) |
| GR90 | >5760 (46.51) | 1240 (12.29) | >5760 (46.67) | >5760 (47.60) |
| GR90 (pop.1)/GR90 (pop.2) | GR90 (pop.3)/GR90 (pop.2) | GR90 (pop.4)/GR90 (pop.2) | |
|---|---|---|---|
| RI | 5.2 | 4.9 | 5.1 |
| Population | Pro 106 CCA | Ala 109 GCT |
|---|---|---|
| 1 s1 (R) | CCA | GCA |
| 1 s2 (R) | CCA | GCA |
| 1 s3 (R) | CCA | GCG |
| 1 s4 (R) | CCA | GCG |
| 2 s1(R) | CCA | GCA |
| 2 s2 (R) | CCG | GCT |
| 2 s3 (R) | CCA | GCA |
| 3 s1 (S) | CCA | - |
| 3 s2 (S) | CCA | - |
| 4 s1 (R) | CCG | GCG |
| 4 s2 (R) | CCG | GCG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gerakari, M.; Cheimona, N.; Tani, E.; Travlos, I.; Chachalis, D.; Loddo, D.; Mathiassen, S.K.; Gitsopoulos, T.K.; Scarabel, L.; Panozzo, S.; et al. Biochemical and Rapid Molecular Analyses to Identify Glyphosate Resistance in Lolium spp. Agronomy 2022, 12, 40. https://doi.org/10.3390/agronomy12010040
Gerakari M, Cheimona N, Tani E, Travlos I, Chachalis D, Loddo D, Mathiassen SK, Gitsopoulos TK, Scarabel L, Panozzo S, et al. Biochemical and Rapid Molecular Analyses to Identify Glyphosate Resistance in Lolium spp. Agronomy. 2022; 12(1):40. https://doi.org/10.3390/agronomy12010040
Chicago/Turabian StyleGerakari, Maria, Nikolina Cheimona, Eleni Tani, Ilias Travlos, Demosthenis Chachalis, Donato Loddo, Solvejg Kopp Mathiassen, Thomas K. Gitsopoulos, Laura Scarabel, Silvia Panozzo, and et al. 2022. "Biochemical and Rapid Molecular Analyses to Identify Glyphosate Resistance in Lolium spp." Agronomy 12, no. 1: 40. https://doi.org/10.3390/agronomy12010040