
Heat Stress in Cotton: A Review on Predicted and Unpredicted Growth-Yield Anomalies and Mitigating Breeding Strategies


 ,
,  ,
,  , ,
, ,
Abstract
:1. Introduction
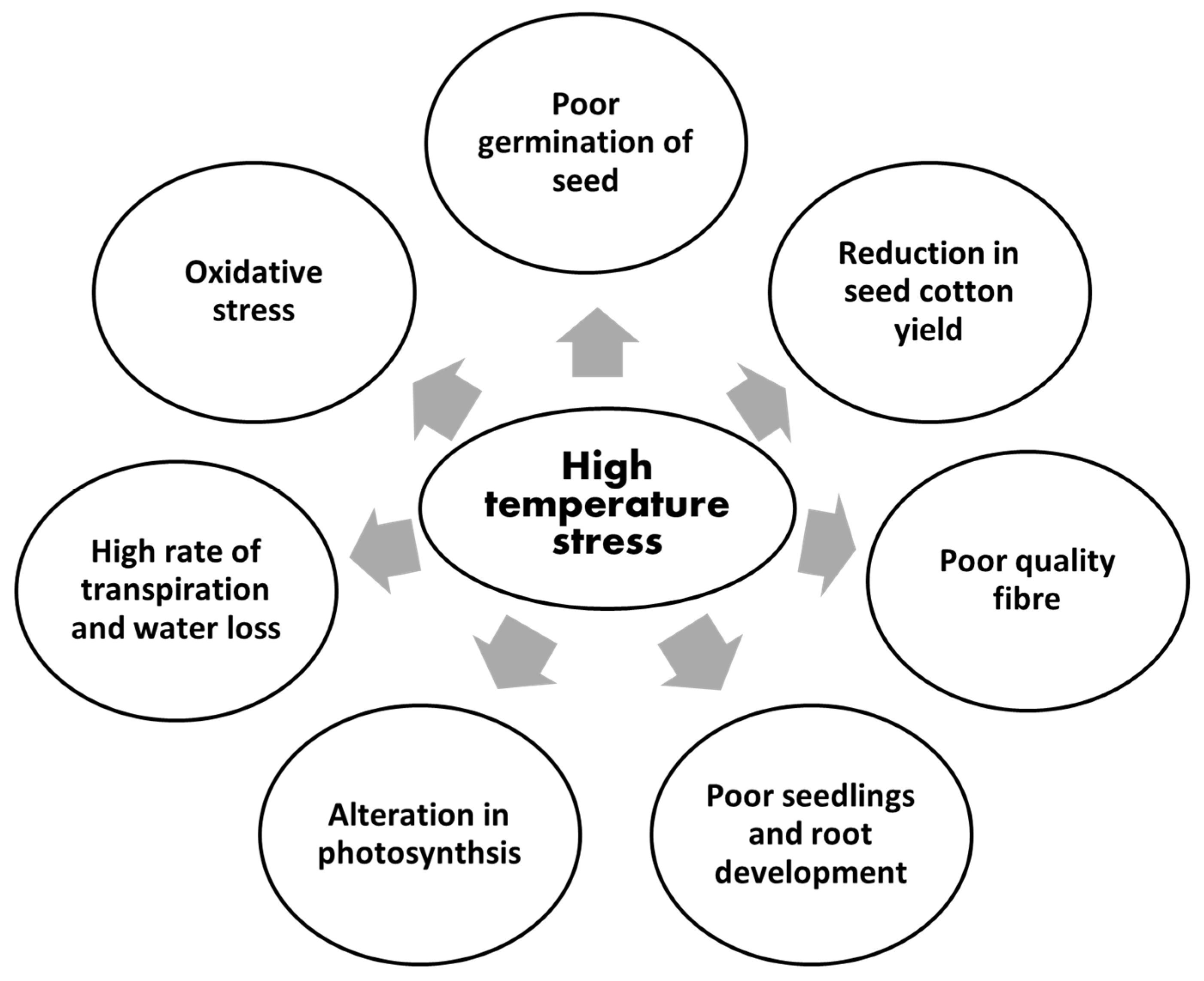
2. Effects of High Temperature Stress on Cotton
2.1. Effects on Germination of Cotton Seed
2.2. Effects on Early Vegetative Growth Stages
2.3. Effects on Lateral Vegetative Growth Stages
2.4. Effects on Yield and Quality of Fibre
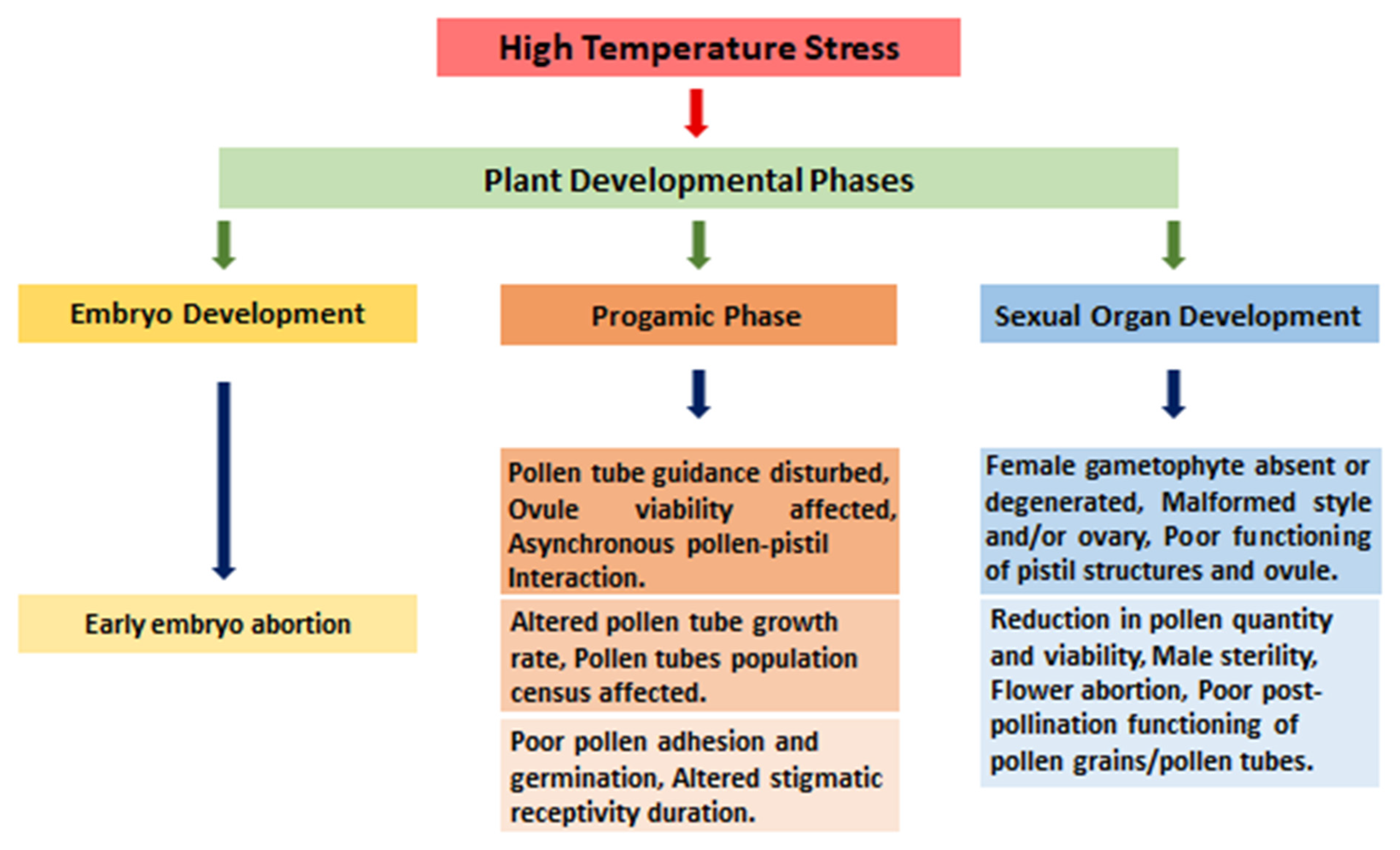
2.5. Effect on Floral Parts
2.6. Effects on Physiological Attributes
3. Mechanisms of Heat Tolerance
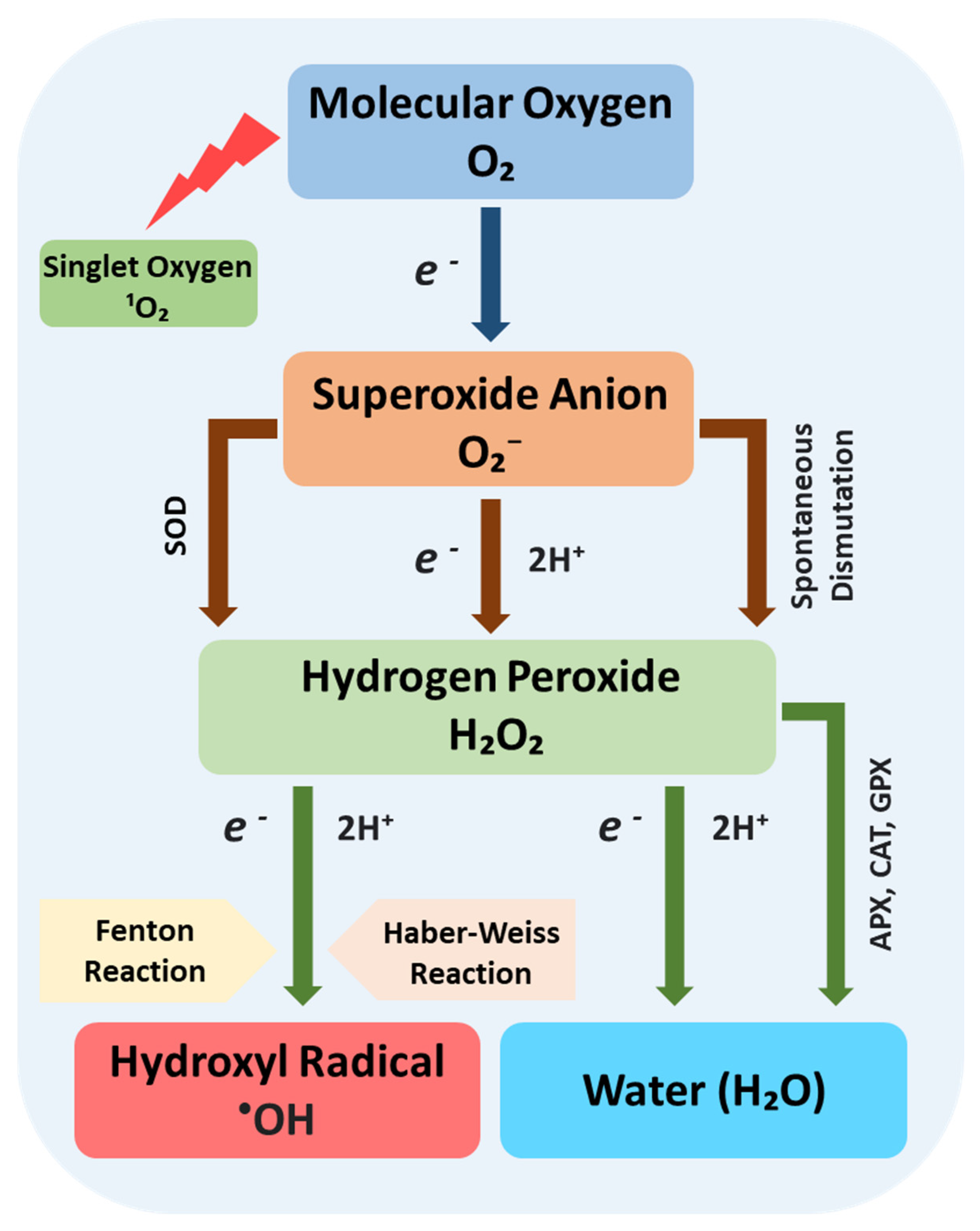
3.1. Antioxidant Activity in Response to Oxidative Stress
3.2. Role of Heat Shock Proteins
3.3. Small RNAs Activity in Regulating Heat Stress
4. Breeding Strategies for High Temperature Stress Tolerance
4.1. Conventional Breeding Approaches
4.2. Molecular and Biotechnological Approaches
4.3. Transgenic Approaches
4.4. CRISPR-Cas Mediated Genome Editing
5. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Disclaimer
References
- Meyer, L.A. The World and US Cotton Outlook for 2019/20. 2019. Available online: https://www.usda.gov/sites/default/files/documents/Leslie_Meyer.pdf (accessed on 12 May 2021).
- Khan, M.A.; Wahid, A.; Ahmad, M.; Tahir, M.T.; Ahmed, M.; Ahmad, S.; Hasanuzzaman, M. World cotton production and consumption: An overview. Cotton Prod. Uses 2020, 1–7. [Google Scholar] [CrossRef]
- Zahra, N.; Shaukat, K.; Hafeez, M.B.; Raza, A.; Hussain, S.; Chaudhary, M.T.; Wahid, A. Physiological and molecular responses to high chilling and freezing temperature in plant growth and production: Consequences and mitigation possibilities. In Harsh Environment and Plant Resilience: Molecular and Functional Aspects; Springer: New York, NY, USA, 2021; p. 235. [Google Scholar]
- Zhu, Y.N.; Shi, D.Q.; Ruan, M.B.; Zhang, L.L.; Meng, Z.H.; Liu, J.; Yang, W.C. Transcriptome analysis reveals crosstalk of responsive genes to multiple abiotic stresses in cotton (Gossypium hirsutum L.). PLoS ONE 2013, 8, e80218. [Google Scholar]
- Downes, S.; Walsh, T.; Tay, W.T. Bt resistance in Australian insect pest species. Curr. Opin. Insect Sci. 2016, 15, 78–83. [Google Scholar] [CrossRef]
- Van der Sluijs, M.J.; Hunter, L. A review on the formation, causes, measurement, implications and reduction of neps during cotton processing. Text. Prog. 2016, 48, 221–323. [Google Scholar] [CrossRef]
- Hall, A.E.; Botany and Plant Sciences Department University of California, Riverside. Heat Stress and Its Impact. 2001. Available online: https://plantstress.com/heat/ (accessed on 13 June 2021).
- Dhyani, K.; Ansari, M.W.; Rao, Y.R.; Verma, R.S.; Shukla, A.; Tuteja, N. Comparative physiological response of wheat genotypes under terminal heat stress. Plant Signal. Behav. 2013, 8, e24564. [Google Scholar] [CrossRef] [PubMed]
- Phillips, J.B. Cotton Response to High Temperature Stress During Reproductive Development. 2012. Available online: https://scholarworks.uark.edu/etd/394 (accessed on 22 July 2021).
- Raza, A.; Ahmad, M. Analysing the Impact of Climate Change on Cotton Productivity in Punjab and Sindh, Pakistan. 2015. Available online: https://mpra.ub.uni-muenchen.de/72867/ (accessed on 7 July 2021).
- Zahid, K.R.; Ali, F.; Shah, F.; Younas, M.; Shah, T.; Shahwar, D.; Hassan, W.; Ahmad, Z.; Qi, C.; Lu, Y. Response and tolerance mechanism of cotton Gossypium hirsutum L. to elevated temperature stress: A review. Front. Plant Sci. 2016, 7, 937. [Google Scholar] [CrossRef] [Green Version]
- Shamsuzzaman, K.; Hamid, M.; Azad, M.; Hussain, M.; Majid, M. Varietal improvement of cotton (Gossypium hirsutum) through mutation breeding. In Improvement of New and Traditional Industrial Crops by Induced Mutations and Related Biotechnology; International Atomic Energy Agency: Vienna, Austria, 2003; pp. 81–94. [Google Scholar]
- Ahloowalia, B.; Maluszynski, M.; Nichterlein, K. Global impact of mutation-derived varieties. Euphytica 2004, 135, 187–204. [Google Scholar] [CrossRef]
- Challinor, A.; Wheeler, T.; Craufurd, P.; Slingo, J. Simulation of the impact of high temperature stress on annual crop yields. Agric. For. Meteorol. 2005, 135, 180–189. [Google Scholar] [CrossRef] [Green Version]
- Snider, J.L.; Oosterhuis, D.M.; Skulman, B.W.; Kawakami, E.M. Heat stress-induced limitations to reproductive success in Gossypium hirsutum. Physiol. Plant. 2009, 137, 125–138. [Google Scholar] [CrossRef]
- Khan, A.H.; Min, L.; Ma, Y.; Wu, Y.; Ding, Y.; Li, Y.; Xie, S.; Ullah, A.; Shaban, M.; Manghwar, H. High day and night temperatures distinctively disrupt fatty acid and jasmonic acid metabolism, inducing male sterility in cotton. J. Exp. Bot. 2020, 71, 6128–6141. [Google Scholar] [CrossRef]
- Nabi, G.; Mullins, C. Soil temperature dependent growth of cotton seedlings before emergence. Pedosphere 2008, 18, 54–59. [Google Scholar] [CrossRef]
- Zia, S.; Khan, M.A. Effect of light, salinity, and temperature on seed germination of Limonium stocksii. Can. J. Bot. 2004, 82, 151–157. [Google Scholar] [CrossRef] [Green Version]
- Riley, G.J. Effects of high temperature on the germination of maize (Zea mays L.). Planta 1981, 151, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Burke, J.J.; Upchurch, D.R. Leaf temperature and transpirational control in cotton. Environ. Exp. Bot. 1989, 29, 487–492. [Google Scholar] [CrossRef]
- Krzyzanowski, F.C.; Delouche, J.C. Germination of cotton seed in relation to temperature. Rev. Bras. Sementes 2011, 33, 543–548. [Google Scholar] [CrossRef] [Green Version]
- Parera, C.A.; Cantliffe, D.J. Presowing seed priming. Hortic. Rev. 1994, 16, 109–141. [Google Scholar]
- Raphael, J.; Gazola, B.; Nunes, J.G.; Macedo, G.C.; Rosolem, C.A. Cotton germination and emergence under high diurnal temperatures. Crop Sci. 2017, 57, 2761–2769. [Google Scholar] [CrossRef]
- Carmo-Silva, A.E.; Gore, M.A.; Andrade-Sanchez, P.; French, A.N.; Hunsaker, D.J.; Salvucci, M.E. Decreased CO2 availability and inactivation of Rubisco limit photosynthesis in cotton plants under heat and drought stress in the field. Environ. Exp. Bot. 2012, 83, 1–11. [Google Scholar] [CrossRef]
- Sethar, M.A.; Laidman, D.; Pahoja, V.M.; Chachar, Q.; Mirjat, M.A. Thermotolerance in Cotton Roots at Early Stage of Emergence. Pak. J. Biol. Sci. 2001, 4, 361–364. [Google Scholar] [CrossRef]
- Reddy, K.R.; Hodges, H.F.; McKinion, J.M. A comparison of scenarios for the effect of global climate change on cotton growth and yield. Funct. Plant Biol. 1997, 24, 707–713. [Google Scholar] [CrossRef]
- Kamal, M.; Saleem, M.; Shahid, M.; Awais, M.; Khan, H.; Ahmed, K. Ascorbic acid triggered physiochemical transformations at different phenological stages of heat-stressed Bt cotton. J. Agron. Crop Sci. 2017, 203, 323–331. [Google Scholar] [CrossRef]
- Saleem, M.F.; Kamal, M.A.; Anjum, S.A.; Shahid, M.; Raza, M.A.S.; Awais, M. Improving the performance of Bt-cotton under heat stress by foliar application of selenium. J. Plant Nutr. 2018, 41, 1711–1723. [Google Scholar] [CrossRef]
- Brown, P. Cotton Heat Stress. 2008. Available online: https://cals.arizona.edu/azmet/az1448.pdf (accessed on 14 July 2021).
- Tariq, M.; Yasmeen, A.; Ahmad, S.; Hussain, N.; Afzal, M.N.; Hasanuzzaman, M. Shedding of fruiting structures in cotton: Factors, compensation and prevention. Trop. Subtrop. Agroecosystems 2017, 20, 251–262. [Google Scholar]
- Karademir, E.; Karademir, Ç.; Ekinci, R.; Başbağ, S.; Başal, H. Screening cotton varieties (Gossypium hirsutum L.) for heat tolerance under field conditions. Afr. J. Agric. Res. 2012, 7, 6335–6342. [Google Scholar]
- Rauf, S.; Khan, T.M.; Naveed, A.; Munir, H. Modified path to high lint yield in upland cotton (Gossypium hirsutum L.) under two temperature regimes. Turk. J. Biol. 2007, 31, 119–126. [Google Scholar]
- Kamal, M.; Saleem, M.; Wahid, M.; Shakeel, A. Effects of ascorbic acid on membrane stability and yield of heatstressed BT cotton. J. Anim. Plant Sci. 2017, 27, 192–199. [Google Scholar]
- Singh, K.; Wijewardana, C.; Gajanayake, B.; Lokhande, S.; Wallace, T.; Jones, D.; Reddy, K.R. Genotypic variability among cotton cultivars for heat and drought tolerance using reproductive and physiological traits. Euphytica 2018, 214, 57. [Google Scholar] [CrossRef]
- Zeiher, C.A.; Brown, P.W.; Silvertooth, J.C.; Matumba, N.; Mitton, N. The Effect of Night Temperature on Cotton Reproductive Development. 1994. Available online: https://repository.arizona.edu/bitstream/handle/10150/209598/370096-089-096.pdf?sequence=1 (accessed on 27 June 2021).
- Zeiher, C.; Matumba, N.; Brown, P.; Silvertooth, J. Response of upland cotton to elevated night temperatures. II. Results of controlled environmental studies. In Proceedings of the Beltwide Cotton Conferences, San Antonio, TX, USA, 4–7 January 1995. [Google Scholar]
- Brown, P.; Zeiher, C. A model to estimate cotton canopy temperature in the desert southwest. In Proceedings of the Beltwide Cotton Conferences, San Diego, CA, USA, 5–9 January 1998; National Cotton Council of America: Memphis, TN, USA, 1998. [Google Scholar]
- Sabagh, A.E.; Hossain, A.; Islam, M.S.; Barutcular, C.; Ratnasekera, D.; Gormus, O.; Amanet, K.; Mubeen, M.; Nasim, W.; Fahad, S. Drought and heat stress in cotton (Gossypium hirsutum L.): Consequences and their possible mitigation strategies. In Agroomic Crops; Springer: New York, NY, USA, 2020. [Google Scholar] [CrossRef]
- Yaqoob, M.; Fiaz, S.; Ijaz, B. Correlation analysis for yield and fiber quality traits in upland cotton. Commun. Plant Sc. 2016, 6, 55–60. [Google Scholar]
- Peng, S.; Krieg, D.; Hicks, S. Cotton lint yield response to accumulated heat units and soil water supply. Field Crops Res. 1989, 19, 253–262. [Google Scholar] [CrossRef]
- Reddy, K.R.; Davidonis, G.H.; Johnson, A.S.; Vinyard, B.T. Temperature regime and carbon dioxide enrichment alter cotton boll development and fiber properties. Agron. J. 1999, 91, 851–858. [Google Scholar] [CrossRef]
- Pettigrew, W. The effect of higher temperatures on cotton lint yield production and fiber quality. Crop Sci. 2008, 48, 278–285. [Google Scholar] [CrossRef] [Green Version]
- Abro, S.; Rajput, M.T.; Khan, M.A.; Sial, M.A.; Tahir, S.S. Screening of cotton (Gossypium hirsutum L.) genotypes for heat tolerance. Pak. J. Bot 2015, 47, 2085–2091. [Google Scholar]
- Liakatas, A.; Roussopoulos, D.; Whittington, W. Controlled-temperature effects on cotton yield and fibre properties. J. Agric. Sci. 1998, 130, 463–471. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Shi, W.; Broughton, K.; Smith, R.; Sharwood, R.; Payton, P.; Bange, M.; Tissue, D.T. Impacts of growth temperature, water deficit and heatwaves on carbon assimilation and growth of cotton plants (Gossypium hirsutum L.). Environ. Exp. Bot. 2020, 179, 104204. [Google Scholar] [CrossRef]
- He, X.-Y.; Zhou, Z.-G.; Dai, Y.-J.; Qiang, Z.-Y.; Chen, B.-L.; Wang, Y.-H. Effect of increased temperature in boll period on fiber yield and quality of cotton and its physiological mechanism. Yingyong Shengtai Xuebao 2013, 24, 12. [Google Scholar]
- Bo, X.; Zhou, Z.-G.; Guo, L.-T.; Xu, W.-Z.; Zhao, W.-Q.; Chen, B.-L.; Meng, Y.-L.; Wang, Y.-H. Susceptible time window and endurable duration of cotton fiber development to high temperature stress. J. Integr. Agric. 2017, 16, 1936–1945. [Google Scholar]
- Abdelgadir, H.; Johnson, S.; Van Staden, J. Pollen viability, pollen germination and pollen tube growth in the biofuel seed crop Jatropha curcas (Euphorbiaceae). S. Afr. J. Bot. 2012, 79, 132–139. [Google Scholar] [CrossRef] [Green Version]
- Kakani, V.; Reddy, K.; Koti, S.; Wallace, T.; Prasad, P.; Reddy, V.; Zhao, D. Differences in in vitro pollen germination and pollen tube growth of cotton cultivars in response to high temperature. Ann. Bot. 2005, 96, 59–67. [Google Scholar] [CrossRef] [Green Version]
- Burke, J.J.; Velten, J.; Oliver, M.J. In vitro analysis of cotton pollen germination. Agron. J. 2004, 96, 359–368. [Google Scholar] [CrossRef]
- Hedhly, A.; Hormaza, J.; Herrero, M. The effect of temperature on stigmatic receptivity in sweet cherry (Prunus avium L.). Plant Cell Environ. 2003, 26, 1673–1680. [Google Scholar] [CrossRef] [Green Version]
- Hedhly, A.; Hormaza, J.; Herrero, M. The effect of temperature on pollen germination, pollen tube growth, and stigmatic receptivity in peach. Plant Biol. 2005, 7, 476–483. [Google Scholar] [CrossRef] [Green Version]
- Snider, J.L.; Oosterhuis, D.M. Heat stress and pollen-pistil interactions. In Flowering and Fruiting in Cotton; Publ. Cotton Foundation: Memphis, TN, USA, 2012; pp. 59–78. [Google Scholar]
- Gago, J.; de Menezes Daloso, D.; Figueroa, C.M.; Flexas, J.; Fernie, A.R.; Nikoloski, Z. Relationships of leaf net photosynthesis, stomatal conductance, and mesophyll conductance to primary metabolism: A multispecies meta-analysis approach. Plant Physiol. 2016, 171, 265–279. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.; Roychowdhury, R.; Fujita, M. Physiological, biochemical, and molecular mechanisms of heat stress tolerance in plants. Int. J. Mol. Sci. 2013, 14, 9643–9684. [Google Scholar] [CrossRef] [PubMed]
- Crafts-Brandner, S.; Law, R. Effect of heat stress on the inhibition and recovery of the ribulose-1, 5-bisphosphate carboxylase/oxygenase activation state. Planta 2000, 212, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Schrader, S.; Wise, R.; Wacholtz, W.; Ort, D.R.; Sharkey, T. Thylakoid membrane responses to moderately high leaf temperature in Pima cotton. Plant Cell Environ. 2004, 27, 725–735. [Google Scholar] [CrossRef]
- Karademir, E.; Karademir, C.; Sevilmis, U.; Basal, H. Correlations between canopy temperature, chlorophyll content and yield in heat tolerant cotton (Gossypium hirsutum L.) genotypes. Fresenius Environ. Bull. 2018, 27, 5230–5237. [Google Scholar]
- Bita, C.; Gerats, T. Plant tolerance to high temperature in a changing environment: Scientific fundamentals and production of heat stress-tolerant crops. Front. Plant Sci. 2013, 4, 273. [Google Scholar] [CrossRef] [Green Version]
- Bibi, A.; Oosterhuis, D.; Brown, R.; Gonias, E.; Bourland, F. The physiological response of cotton to high temperatures for germplasm screening. Summ. Ark. Cotton Res. 2003, 521, 87–93. [Google Scholar]
- Rahman, H.; Malik, S.A.; Saleem, M. Heat tolerance of upland cotton during the fruiting stage evaluated using cellular membrane thermostability. Field Crops Res. 2004, 85, 149–158. [Google Scholar] [CrossRef]
- Khan, N.; Ahmad, I.; Azhar, M.T. Genetic variation in relative cell injury for breeding upland cotton under high temperature stress. Turk. J. Field Crops 2017, 22, 152–159. [Google Scholar] [CrossRef]
- Urban, J.; Ingwers, M.; McGuire, M.A.; Teskey, R.O. Stomatal conductance increases with rising temperature. Plant Signal. Behav. 2017, 12, e1356534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Z.; Percy, R.G.; Qualset, C.O.; Zeiger, E. Stomatal conductance predicts yields in irrigated Pima cotton and bread wheat grown at high temperatures. J. Exp. Bot. 1998, 453–460. [Google Scholar] [CrossRef] [Green Version]
- Lu, Z.; Chen, J.; Percy, R.G.; Zeiger, E. Photosynthetic rate, stomatal conductance and leaf area in two cotton species (Gossypium barbadense and Gossypium hirsutum) and their relation with heat resistance and yield. Funct. Plant Biol. 1997, 24, 693–700. [Google Scholar] [CrossRef]
- Lu, Z.; Quiñones, M.A.; Zeiger, E. Temperature dependence of guard cell respiration and stomatal conductance co-segregate in an F2 population of Pima cotton. Funct. Plant Biol. 2000, 27, 457–462. [Google Scholar] [CrossRef]
- Khan, A.I.; Khan, I.A.; Sadaqat, H.A. Heat tolerance is variable in cotton (Gossypium hirsutum L.) and can be exploited for breeding of better yielding cultivars under high temperature regimes. Pak. J. Bot. 2008, 40, 2053–2058. [Google Scholar]
- Rahman, H.; Murtaza, N.; Shah, K.; Qayyum, A.; Ullah, I.; Malik, W. Genetic variation for stomatal conductance in upland cotton as influenced by heat-stressed and non-stressed growing regimes. Acta Agron. Hung. 2008, 56, 11–19. [Google Scholar] [CrossRef]
- Suzuki, N.; Mittler, R. Reactive oxygen species and temperature stresses: A delicate balance between signaling and destruction. Physiol. Plant. 2006, 126, 45–51. [Google Scholar] [CrossRef]
- Suzuki, N.; Koussevitzky, S.; Mittler, R.; Miller, G. ROS and redox signalling in the response of plants to abiotic stress. Plant Cell Environ. 2012, 35, 259–270. [Google Scholar] [CrossRef]
- Considine, M.J.; María Sandalio, L.; Helen Foyer, C. Unravelling how plants benefit from ROS and NO reactions, while resisting oxidative stress. Ann. Bot. 2015, 116, 469–473. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.; Singh, S.; Parihar, P.; Mishra, R.K.; Tripathi, D.K.; Singh, V.P.; Chauhan, D.K.; Prasad, S.M. Reactive oxygen species (ROS): Beneficial companions of plants’ developmental processes. Front. Plant Sci. 2016, 7, 1299. [Google Scholar] [CrossRef] [Green Version]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Majeed, S.; Malik, T.A.; Rana, I.A.; Azhar, M.T. Antioxidant and physiological responses of upland cotton accessions grown under high-temperature regimes. Iran. J. Sci. Technol. Trans. A Sci. 2019, 43, 2759–2768. [Google Scholar] [CrossRef]
- Sekmen, A.H.; Ozgur, R.; Uzilday, B.; Turkan, I. Reactive oxygen species scavenging capacities of cotton (Gossypium hirsutum) cultivars under combined drought and heat induced oxidative stress. Environ. Exp. Bot. 2014, 99, 141–149. [Google Scholar] [CrossRef]
- Gür, A.; Demirel, U.; Özden, M.; Kahraman, A.; Çopur, O. Diurnal gradual heat stress affects antioxidant enzymes, proline accumulation and some physiological components in cotton (Gossypium hirsutum L.). Afr. J. Biotechnol. 2010, 9, 1008–1015. [Google Scholar]
- Sarwar, M.; Saleem, M.F.; Ullah, N.; Rizwan, M.; Ali, S.; Shahid, M.R.; Alamri, S.A.; Alyemeni, M.N.; Ahmad, P. Exogenously applied growth regulators protect the cotton crop from heat-induced injury by modulating plant defense mechanism. Sci. Rep. 2018, 8, 17086. [Google Scholar] [CrossRef] [Green Version]
- Loka, D.A.; Oosterhuis, D.M. Effect of high night temperatures during anthesis on cotton (Gossypium hirsutum L.) pistil and leaf physiology and biochemistry. Aust. J. Crop Sci. 2016, 10, 741. [Google Scholar] [CrossRef]
- Wang, W.; Vinocur, B.; Shoseyov, O.; Altman, A. Role of plant heat-shock proteins and molecular chaperones in the abiotic stress response. Trends Plant Sci. 2004, 9, 244–252. [Google Scholar] [CrossRef]
- Kotak, S.; Larkindale, J.; Lee, U.; von Koskull-Döring, P.; Vierling, E.; Scharf, K.-D. Complexity of the heat stress response in plants. Curr. Opin. Plant Biol. 2007, 10, 310–316. [Google Scholar] [CrossRef]
- Waters, E.R.; Lee, G.J.; Vierling, E. Evolution, structure and function of the small heat shock proteins in plants. J. Exp. Bot. 1996, 47, 325–338. [Google Scholar] [CrossRef]
- Mogk, A.; Schlieker, C.; Friedrich, K.L.; Schönfeld, H.-J.; Vierling, E.; Bukau, B. Refolding of substrates bound to small Hsps relies on a disaggregation reaction mediated most efficiently by ClpB/DnaK. J. Biol. Chem. 2003, 278, 31033–31042. [Google Scholar] [CrossRef] [Green Version]
- Van Montfort, R.L.; Basha, E.; Friedrich, K.L.; Slingsby, C.; Vierling, E. Crystal structure and assembly of a eukaryotic small heat shock protein. Nat. Struct. Mol. Biol. 2001, 8, 1025. [Google Scholar] [CrossRef]
- Malik, M.K.; Slovin, J.P.; Hwang, C.H.; Zimmerman, J.L. Modified expression of a carrot small heat shock protein gene, Hsp17. 7, results in increased or decreased thermotolerance. Plant J. 1999, 20, 89–99. [Google Scholar] [CrossRef]
- Maqbool, A.; Zahur, M.; Irfan, M.; Qaiser, U.; Rashid, B.; Husnain, T. Identification, characterization and expression of drought related alpha-crystalline heat shock protein gene (GHSP26) from Desi cotton. Crop Sci. 2007, 47, 2437–2444. [Google Scholar] [CrossRef]
- Lee, G.J.; Vierling, E. A small heat shock protein cooperates with heat shock protein 70 systems to reactivate a heat-denatured protein. Plant Physiol. 2000, 122, 189–198. [Google Scholar] [CrossRef] [Green Version]
- Haslbeck, M.; Vierling, E. A first line of stress defense: Small heat shock proteins and their function in protein homeostasis. J. Mol. Biol. 2015, 427, 1537–1548. [Google Scholar] [CrossRef] [Green Version]
- Boston, R.S.; Viitanen, P.V.; Vierling, E. Molecular chaperones and protein folding in plants. In Post-Transcriptional Control of Gene Expression in Plants; Springer: Berlin/Heidelberg, Germany, 1996; pp. 191–222. [Google Scholar]
- Hartl, F.U.; Bracher, A.; Hayer-Hartl, M. Molecular chaperones in protein folding and proteostasis. Nature 2011, 475, 324. [Google Scholar] [CrossRef]
- Sung, D.Y.; Kaplan, F.; Guy, C.L. Plant Hsp70 molecular chaperones: Protein structure, gene family, expression and function. Physiol. Plant. 2001, 113, 443–451. [Google Scholar] [CrossRef]
- Scarpeci, T.E.; Zanor, M.I.; Valle, E.M. Investigating the role of plant heat shock proteins during oxidative stress. Plant Signal. Behav. 2008, 3, 856–857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Liu, S.S.; Yi, C.Y.; Wang, F.; Zhou, J.; Xia, X.J.; Shi, K.; Zhou, Y.H.; Yu, J.Q. Hydrogen peroxide mediates abscisic acid-induced HSP 70 accumulation and heat tolerance in grafted cucumber plants. Plant Cell Environ. 2014, 37, 2768–2780. [Google Scholar] [CrossRef] [PubMed]
- Rehman, A.; Atif, R.M.; Qyyum, A.; Du, X.; Hinze, L.; Azhar, M.T. Genome-wide identification and characterization of HSP70 gene family in four species of cotton. Genomics 2020, 112, 4442–4453. [Google Scholar] [CrossRef] [PubMed]
- Krishna, P.; Gloor, G. The Hsp90 family of proteins in Arabidopsis thaliana. Cell Stress Chaperones 2001, 6, 238. [Google Scholar] [CrossRef]
- Kadota, Y.; Shirasu, K. The HSP90 complex of plants. Biochim. Et Biophys. Acta (BBA)-Mol. Cell Res. 2012, 1823, 689–697. [Google Scholar] [CrossRef] [Green Version]
- Sable, A.; Rai, K.M.; Choudhary, A.; Yadav, V.K.; Agarwal, S.K.; Sawant, S.V. Inhibition of heat shock proteins HSP90 and HSP70 induce oxidative stress, suppressing cotton fiber development. Sci. Rep. 2018, 8, 3620. [Google Scholar] [CrossRef] [PubMed]
- Zietkiewicz, S.; Krzewska, J.; Liberek, K. Successive and synergistic action of the Hsp70 and Hsp100 chaperones in protein disaggregation. J. Biol. Chem. 2004, 279, 44376–44383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winkler, J.; Tyedmers, J.; Bukau, B.; Mogk, A. Hsp70 targets Hsp100 chaperones to substrates for protein disaggregation and prion fragmentation. J. Cell Biol 2012, 198, 387–404. [Google Scholar] [CrossRef] [Green Version]
- Hemmingsen, S.M.; Woolford, C.; van der Vies, S.M.; Tilly, K.; Dennis, D.T.; Georgopoulos, C.P.; Hendrix, R.W.; Ellis, R.J. Homologous plant and bacterial proteins chaperone oligomeric protein assembly. Nature 1988, 333, 330. [Google Scholar] [CrossRef]
- Apuya, N.R.; Yadegari, R.; Fischer, R.L.; Harada, J.J.; Zimmerman, J.L.; Goldberg, R.B. The Arabidopsis embryo mutant schlepperless has a defect in the chaperonin-60α gene. Plant Physiol. 2001, 126, 717–730. [Google Scholar] [CrossRef] [Green Version]
- Ishikawa, A.; Tanaka, H.; Nakai, M.; Asahi, T. Deletion of a chaperonin 60β gene leads to cell death in the Arabidopsis lesion initiation 1 mutant. Plant Cell Physiol. 2003, 44, 255–261. [Google Scholar] [CrossRef]
- Zabaleta, E.; Oropeza, A.; Assad, N.; Mandel, A.; Salerno, G.; Herrera-Estrella, L. Antisense expression of chaperonin 60β in transgenic tobacco plants leads to abnormal phenotypes and altered distribution of photoassimilates. Plant J. 1994, 6, 425–432. [Google Scholar] [CrossRef]
- Miemyk, J. The 70 kDa stress-related proteins as molecular chaperones. Trends Plant Sci. 1997, 2, 180–187. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, M.; Chen, J.; Rong, J.; Ding, M. Genome-wide analysis of HSP70 superfamily in Gossypium raimondii and the expression of orthologs in Gossypium hirsutum. Yi Chuan Hered. 2014, 36, 921–933. [Google Scholar]
- Young, J.C.; Moarefi, I.; Hartl, F.U. Hsp90: A specialized but essential protein-folding tool. J. Cell Biol. 2001, 154, 267. [Google Scholar] [CrossRef] [PubMed]
- Buchner, J. Hsp90 & Co.—A holding for folding. Trends Biochem. Sci. 1999, 24, 136–141. [Google Scholar]
- Zhang, Z.; Quick, M.K.; Kanelakis, K.C.; Gijzen, M.; Krishna, P. Characterization of a plant homolog of hop, a cochaperone of hsp90. Plant Physiol. 2003, 131, 525–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Latterich, M.; Patel, S. The AAA team: Related ATPases with diverse functions. Trends Cell Biol. 1998, 8, 65–71. [Google Scholar] [CrossRef]
- Lee, U.; Rioflorido, I.; Hong, S.W.; Larkindale, J.; Waters, E.R.; Vierling, E. The Arabidopsis ClpB/Hsp100 family of proteins: Chaperones for stress and chloroplast development. Plant J. 2007, 49, 115–127. [Google Scholar] [CrossRef] [Green Version]
- Mishra, R.C.; Grover, A. ClpB/Hsp100 proteins and heat stress tolerance in plants. Crit. Rev. Biotechnol. 2016, 36, 862–874. [Google Scholar] [CrossRef]
- Barrera-Figueroa, B.E.; Gao, L.; Wu, Z.; Zhou, X.; Zhu, J.; Jin, H.; Liu, R.; Zhu, J.-K. High throughput sequencing reveals novel and abiotic stress-regulated microRNAs in the inflorescences of rice. BMC Plant Biol. 2012, 12, 132. [Google Scholar] [CrossRef] [Green Version]
- Barakat, A.; Sriram, A.; Park, J.; Zhebentyayeva, T.; Main, D.; Abbott, A. Genome wide identification of chilling responsive microRNAs in Prunus persica. BMC Genom. 2012, 13, 481. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Yang, M.; Zhang, X. The function of small RNAs in plant biotic stress response. J. Integr. Plant Biol. 2016, 58, 312–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.-W.; Czech, B.; Weigel, D. miR156-regulated SPL transcription factors define an endogenous flowering pathway in Arabidopsis thaliana. Cell 2009, 138, 738–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.; Li, J.; Liu, Y.; Zhang, Q.; Gao, Y.; Fang, K.; Cao, Q.; Qin, L.; Xing, Y. Roles of the GA-mediated SPL Gene Family and miR156 in the Floral Development of Chinese Chestnut (Castanea mollissima). Int. J. Mol. Sci. 2019, 20, 1577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, N.; Tu, L.; Wang, L.; Hu, H.; Xu, J.; Zhang, X. MicroRNA 157-targeted SPL genes regulate floral organ size and ovule production in cotton. BMC Plant Biol. 2017, 17, 7. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Wang, H.; Lu, Y.; de Ruiter, M.; Cariaso, M.; Prins, M.; van Tunen, A.; He, Y. Identification of conserved and novel microRNAs that are responsive to heat stress in Brassica rapa. J. Exp. Bot. 2011, 63, 1025–1038. [Google Scholar] [CrossRef]
- Song, C.; Fang, J.; Li, X.; Liu, H.; Chao, C.T. Identification and characterization of 27 conserved microRNAs in citrus. Planta 2009, 230, 671–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stief, A.; Altmann, S.; Hoffmann, K.; Pant, B.D.; Scheible, W.-R.; Bäurle, I. Arabidopsis miR156 regulates tolerance to recurring environmental stress through SPL transcription factors. Plant Cell 2014, 26, 1792–1807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Sun, F.; Cao, H.; Peng, H.; Ni, Z.; Sun, Q.; Yao, Y. TamiR159 directed wheat TaGAMYB cleavage and its involvement in anther development and heat response. PLoS ONE 2012, 7, e48445. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.; Ma, Y.; Liu, N.; Xu, J.; Hu, Q.; Li, Y.; Wu, Y.; Xie, S.; Zhu, L.; Min, L. micro RNA s involved in auxin signalling modulate male sterility under high-temperature stress in cotton (Gossypium hirsutum). Plant J. 2017, 91, 977–994. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Wang, Q.; Zhang, B. Response of miRNAs and their targets to salt and drought stresses in cotton (Gossypium hirsutum L.). Gene 2013, 530, 26–32. [Google Scholar] [CrossRef]
- Sailaja, B.; Anjum, N.; Prasanth, V.V.; Sarla, N.; Subrahmanyam, D.; Voleti, S.; Viraktamath, B.; Mangrauthia, S.K. Comparative study of susceptible and tolerant genotype reveals efficient recovery and root system contributes to heat stress tolerance in rice. Plant Mol. Biol. Rep. 2014, 32, 1228–1240. [Google Scholar] [CrossRef]
- Ruan, M.-B.; Zhao, Y.-T.; Meng, Z.-H.; Wang, X.-J.; Yang, W.-C. Conserved miRNA analysis in Gossypium hirsutum through small RNA sequencing. Genomics 2009, 94, 263–268. [Google Scholar] [CrossRef] [Green Version]
- Boopathi, M.; Sathish, S.; Kavitha, P.; Dachinamoorthy, P.; Ravikesavan, R. Comparative miRNAome analysis revealed numerous conserved and novel drought responsive miRNAs in cotton (Gossypium spp.). Cotton Genom. Genet. 2016. [Google Scholar] [CrossRef] [Green Version]
- Fang, Y.; Xie, K.; Xiong, L. Conserved miR164-targeted NAC genes negatively regulate drought resistance in rice. J. Exp. Bot. 2014, 65, 2119–2135. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.R.; Pathak, H.; Sharma, S.K.; Kala, Y.K.; Nirjal, M.K.; Singh, G.P.; Goswami, S.; Rai, R.D. Novel and conserved heat-responsive microRNAs in wheat (Triticum aestivum L.). Funct. Integr. Genom. 2015, 15, 323–348. [Google Scholar] [CrossRef] [PubMed]
- Matthews, C.; Arshad, M.; Hannoufa, A. Alfalfa response to heat stress is modulated by microRNA156. Physiol. Plant. 2019, 165, 830–842. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; He, Q.; Chen, G.; Wang, L.; Jin, B. Regulation of non-coding RNAs in heat stress responses of plants. Front. Plant Sci. 2016, 7, 1213. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Liu, J.; Yang, Z.E.; Chen, E.Y.; Zhang, C.J.; Zhang, X.Y.; Li, F.G. Genome-wide analysis of GRAS transcription factor gene family in Gossypium hirsutum L. BMC Genom. 2018, 19, 348. [Google Scholar] [CrossRef]
- Aukerman, M.J.; Sakai, H. Regulation of flowering time and floral organ identity by a microRNA and its APETALA2-like target genes. Plant Cell 2003, 15, 2730–2741. [Google Scholar] [CrossRef] [Green Version]
- François, L.; Verdenaud, M.; Fu, X.; Ruleman, D.; Dubois, A.; Vandenbussche, M.; Bendahmane, A.; Raymond, O.; Just, J.; Bendahmane, M. A miR172 target-deficient AP2-like gene correlates with the double flower phenotype in roses. Sci. Rep. 2018, 8, 12912. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.; Park, M.Y.; Conway, S.R.; Wang, J.-W.; Weigel, D.; Poethig, R.S. The sequential action of miR156 and miR172 regulates developmental timing in Arabidopsis. Cell 2009, 138, 750–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Q.-H.; Upadhyaya, N.M.; Gubler, F.; Helliwell, C.A. Over-expression of miR172 causes loss of spikelet determinacy and floral organ abnormalities in rice (Oryza sativa). BMC Plant Biol. 2009, 9, 149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, Z.; Han, X.; Li, Y.; Wang, J.; Wang, D.; Wang, S.; Fu, X.; Ye, W. Comparative analysis of cotton small RNAs and their target genes in response to salt stress. Genes 2017, 8, 369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wójcik, A.M.; Gaj, M.D. miR393 contributes to the embryogenic transition induced in vitro in Arabidopsis via the modification of the tissue sensitivity to auxin treatment. Planta 2016, 244, 231–243. [Google Scholar] [CrossRef] [Green Version]
- Jeong, D.-H.; Green, P.J. The role of rice microRNAs in abiotic stress responses. J. Plant Biol. 2013, 56, 187–197. [Google Scholar] [CrossRef]
- Xie, F.; Wang, Q.; Sun, R.; Zhang, B. Deep sequencing reveals important roles of microRNAs in response to drought and salinity stress in cotton. J. Exp. Bot. 2014, 66, 789–804. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Huang, J.; Lai, Z.; Xue, Y. F-box proteins in flowering plants. Chin. Sci. Bull. 2002, 47, 1497–1501. [Google Scholar] [CrossRef] [Green Version]
- Xu, G.; Ma, H.; Nei, M.; Kong, H. Evolution of F-box genes in plants: Different modes of sequence divergence and their relationships with functional diversification. Proc. Natl. Acad. Sci. USA 2009, 106, 835–840. [Google Scholar] [CrossRef] [Green Version]
- Song, J.B.; Gao, S.; Sun, D.; Li, H.; Shu, X.X.; Yang, Z.M. miR394 and LCR are involved in Arabidopsis salt and drought stress responses in an abscisic acid-dependent manner. BMC Plant Biol. 2013, 13, 210. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Yang, T.; Yu, T.; Zhang, S.; Mao, X.; Zhao, J.; Wang, X.; Dong, J.; Liu, B. Integrating small RNA sequencing with QTL mapping for identification of miRNAs and their target genes associated with heat tolerance at the flowering stage in rice. Front. Plant Sci. 2017, 8, 43. [Google Scholar] [CrossRef]
- Zhang, B. MicroRNA: A new target for improving plant tolerance to abiotic stress. J. Exp. Bot. 2015, 66, 1749–1761. [Google Scholar] [CrossRef]
- Majeed, S.; Chaudhary, M.T.; Hulse-Kemp, A.M.; Azhar, M.T. Introduction: Crop Wild Relatives in Plant Breeding. In Wild Germplasm for Genetic Improvement in Crop Plants; Elsevier: Amsterdam, The Netherlands, 2021; pp. 1–18. [Google Scholar]
- Raza, A.; Tabassum, J.; Kudapa, H.; Varshney, R.K. Can omics deliver temperature resilient ready-to-grow crops? Crit. Rev. Biotechnol. 2021, 1–24. [Google Scholar] [CrossRef]
- Percy, R.G. Comparison of bulk F2 performance testing and pedigree selection in thirty Pima cotton populations. J. Cotton Sci. 2003, 7, 123. [Google Scholar]
- Tokatlidis, I.; Tsikrikoni, C.; Lithourgidis, A.; Tsialtas, J.; Tzantarmas, C. Intra-cultivar variation in cotton: Response to single-plant yield selection at low density. J. Agric. Sci. 2011, 149, 197–204. [Google Scholar] [CrossRef]
- Mubarik, M.S.; Ma, C.; Majeed, S.; Du, X.; Azhar, M.T. Revamping of Cotton Breeding Programs for Efficient Use of Genetic Resources under Changing Climate. Agronomy 2020, 10, 1190. [Google Scholar] [CrossRef]
- Badigannavar, A.; Myers, G.O.; Jones, D.C. Molecular diversity revealed by AFLP markers in upland cotton genotypes. J. Crop Improv. 2012, 26, 627–640. [Google Scholar] [CrossRef]
- Mohamed, H.; Abdel-Hamid, A. Molecular and biochemical studies for heat tolerance on four cotton genotypes. Rom. Biotechnol. Lett. 2013, 18, 8823–8831. [Google Scholar]
- Du, L.; Cai, C.; Wu, S.; Zhang, F.; Hou, S.; Guo, W. Evaluation and exploration of favorable QTL alleles for salt stress related traits in cotton cultivars (G. hirsutum L.). PLoS ONE 2016, 11, e0151076. [Google Scholar] [CrossRef] [Green Version]
- Majeed, S.; Rana, I.A.; Atif, R.M.; Zulfiqar, A.; Hinze, L.; Azhar, M.T. Role of SNPs in determining QTLs for major traits in cotton. J. Cotton Res. 2019, 2, 5. [Google Scholar] [CrossRef] [Green Version]
- Demirel, U.; Gür, A.; Can, N.; Memon, A. Identification of heat responsive genes in cotton. Biol. Plant. 2014, 58, 515–523. [Google Scholar] [CrossRef]
- Zhang, J.; Srivastava, V.; Stewart, J.M.; Underwood, J. Heat-tolerance in Cotton Is Correlated with Induced Overexpression of Heat-Shock Factors, Heat-Shock Proteins, and General Stress Response Genes. J. Cotton Sci. 2016, 20, 253–262. [Google Scholar]
- Shaheen, T.; Asif, M.; Zafar, Y. Single nucleotide polymorphism analysis of MT-SHSP gene of Gossypium arboreum and its relationship with other diploid cotton genomes, G. hirsutum and Arabidopsis thaliana. Pak. J. Bot. 2009, 41, 117–183. [Google Scholar]
- Ma, W.; Zhao, T.; Li, J.; Liu, B.; Fang, L.; Hu, Y.; Zhang, T. Identification and characterization of the GhHsp20 gene family in Gossypium hirsutum. Sci. Rep. 2016, 6, 32517. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.S.; Fang, D.D.; Jenkins, J.N.; Guo, J.; McCarty, J.C.; Jones, D.C. Evaluation of genomic selection methods for predicting fiber quality traits in Upland cotton. Mol. Genet. Genom. 2020, 295, 67–79. [Google Scholar] [CrossRef] [PubMed]
- Baytar, A.A.; Peynircioğlu, C.; Sezener, V.; Basal, H.; Frary, A.; Frary, A.; Doğanlar, S. Genome-wide association mapping of yield components and drought tolerance-related traits in cotton. Mol. Breed. 2018, 38, 74. [Google Scholar] [CrossRef]
- Su, J.; Pang, C.; Wei, H.; Li, L.; Liang, B.; Wang, C.; Song, M.; Wang, H.; Zhao, S.; Jia, X. Identification of favorable SNP alleles and candidate genes for traits related to early maturity via GWAS in upland cotton. BMC Genom. 2016, 17, 687. [Google Scholar] [CrossRef] [Green Version]
- Batcho, A.A.; Sarwar, M.B.; Rashid, B.; Hassan, S.; Husnain, T. Heat shock protein gene identified from Agave sisalana (As HSP70) confers heat stress tolerance in transgenic cotton (Gossypium hirsutum). Theor. Exp. Plant Physiol. 2021, 33, 141–156. [Google Scholar] [CrossRef]
- Esmaeili, N.; Cai, Y.; Tang, F.; Zhu, X.; Smith, J.; Mishra, N.; Hequet, E.; Ritchie, G.; Jones, D.; Shen, G. Towards doubling fibre yield for cotton in the semiarid agricultural area by increasing tolerance to drought, heat and salinity simultaneously. Plant Biotechnol. J. 2021, 19, 462. [Google Scholar] [CrossRef]
- Burke, J.J.; Chen, J. Enhancement of reproductive heat tolerance in plants. PLoS ONE 2015, 10, e0122933. [Google Scholar] [CrossRef] [Green Version]
- Mishra, N.; Sun, L.; Zhu, X.; Smith, J.; Prakash Srivastava, A.; Yang, X.; Pehlivan, N.; Esmaeili, N.; Luo, H.; Shen, G. Overexpression of the rice SUMO E3 ligase gene OsSIZ1 in cotton enhances drought and heat tolerance, and substantially improves fiber yields in the field under reduced irrigation and rainfed conditions. Plant Cell Physiol. 2017, 58, 735–746. [Google Scholar] [CrossRef]
- Hozain, M.d.; Abdelmageed, H.; Lee, J.; Kang, M.; Fokar, M.; Allen, R.D.; Holaday, A.S. Expression of AtSAP5 in cotton up-regulates putative stress-responsive genes and improves the tolerance to rapidly developing water deficit and moderate heat stress. J. Plant Physiol. 2012, 169, 1261–1270. [Google Scholar] [CrossRef]
- Ali, S.; Rizwan, M.; Arif, M.S.; Ahmad, R.; Hasanuzzaman, M.; Ali, B.; Hussain, A. Approaches in enhancing thermotolerance in plants: An updated review. J. Plant Growth Regul. 2020, 39, 456–480. [Google Scholar] [CrossRef]
- Meriç, S.; Ayan, A.; Atak, Ç. Molecular Abiotic Stress Tolerans Strategies: From Genetic Engineering to Genome Editing Era. In Abiotic Stress in Plants; IntechOpen: London, UK, 2020. [Google Scholar]
- Mubarik, M.S.; Khan, S.H.; Ahmad, A.; Khan, Z.; Sajjad, M.; Khan, I.A. Disruption of Phytoene Desaturase Gene using Transient Expression of Cas9: gRNA Complex. Int. J. Agric. Biol. 2016, 18. [Google Scholar] [CrossRef]
- Bhatnagar-Mathur, P.; Vadez, V.; Sharma, K.K. Transgenic approaches for abiotic stress tolerance in plants: Retrospect and prospects. Plant Cell Rep. 2008, 27, 411–424. [Google Scholar] [CrossRef]
- Ruiz-Vera, U.M.; Siebers, M.H.; Drag, D.W.; Ort, D.R.; Bernacchi, C.J. Canopy warming caused photosynthetic acclimation and reduced seed yield in maize grown at ambient and elevated [CO2]. Glob. Chang. Biol. 2015, 21, 4237–4249. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Z.; Kang, S.; He, L.; Zhao, J.; Zhang, S.; Hu, J.; Zeng, D.; Zhang, G.; Dong, G.; Gao, Z. The newly identified heat-stress sensitive albino 1 gene affects chloroplast development in rice. Plant Sci. 2018, 267, 168–179. [Google Scholar] [CrossRef] [PubMed]
- Klap, C.; Yeshayahou, E.; Bolger, A.M.; Arazi, T.; Gupta, S.K.; Shabtai, S.; Usadel, B.; Salts, Y.; Barg, R. Tomato facultative parthenocarpy results from Sl AGAMOUS-LIKE 6 loss of function. Plant Biotechnol. J. 2017, 15, 634–647. [Google Scholar] [CrossRef]
- Li, J.; Zhang, H.; Si, X.; Tian, Y.; Chen, K.; Liu, J.; Chen, H.; Gao, C. Generation of thermosensitive male-sterile maize by targeted knockout of the ZmTMS5 gene. J. Genet. Genom. Yi Chuan Xue Bao 2017, 44, 465–468. [Google Scholar] [CrossRef]
- Mubarik, M.S.; Khan, S.H.; Sajjad, M.; Raza, A.; Hafeez, M.B.; Yasmeen, T.; Rizwan, M.; Ali, S.; Arif, M.S. A manipulative interplay between positive and negative regulators of phytohormones: A way forward for improving drought tolerance in plants. Physiol. Plant. 2021. [Google Scholar] [CrossRef] [PubMed]
- Yin, W.; Xiao, Y.; Niu, M.; Meng, W.; Li, L.; Zhang, X.; Liu, D.; Zhang, G.; Qian, Y.; Sun, Z. ARGONAUTE2 enhances grain length and salt tolerance by activating BIG GRAIN3 to modulate cytokinin distribution in rice. Plant Cell 2020, 32, 2292–2306. [Google Scholar] [CrossRef]
- Zhai, N.; Jia, H.; Liu, D.; Liu, S.; Ma, M.; Guo, X.; Li, H. GhMAP3K65, a cotton Raf-like MAP3K gene, enhances susceptibility to pathogen infection and heat stress by negatively modulating growth and development in transgenic Nicotiana benthamiana. Int. J. Mol. Sci. 2017, 18, 2462. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Group (Sub-Families) | Representative Members | Intracellular Localization | Major Role | Reference |
|---|---|---|---|---|
| sHSP | Stabilization of non-native proteins and prevent aggregation | [86,87] | ||
| I | Hsp17.6 | Cytosol | ||
| II | Hsp17.9 | Cytosol | ||
| III | Hsp21, Hsp26.2 5 | Chloroplast | ||
| IV | Hsp22 | Endoplasmic reticulum | ||
| V | Hsp23 5 | Mitochondria | ||
| VI | Hsp22.3 | Membrane | ||
| HSP60 | Folding of proteins | [88,89] | ||
| Group I | Cpn60 2 | Chloroplast, mitochondria | ||
| Group II | CCT3 | Cytosol | ||
| HSP70 | Protein import, signal transduction, transcriptional activation, assist refolding and prevent aggregation of proteins | [90,91,92,93] | ||
| DnaK | Hsp/Hsc70 | Cytosol | ||
| Hsp70 | Chloroplast, mitochondria | |||
| Bip 1 | Endoplasmic reticulum | |||
| Hsp110/SSE | Hsp91 | Cytosol | ||
| HSP90 | AtHsp90-1 | Cytosol | Facilitate in genetic buffering and maturation of signaling molecules | [94,95,96] |
| AtHsp90-5 | Chloroplast | |||
| AtHsp90-6 | Mitochondria | |||
| AtHsp90-7 | Endoplasmic reticulum | |||
| HSP100 | Unfolding and disaggregation of proteins | [97,98] | ||
| Class I | ClpB, ClpA/C, ClpD | Cytosol, mitochondria | ||
| Class II | ClpM, ClpN, ClpX, ClpY | Chloroplast |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Majeed, S.; Rana, I.A.; Mubarik, M.S.; Atif, R.M.; Yang, S.-H.; Chung, G.; Jia, Y.; Du, X.; Hinze, L.; Azhar, M.T. Heat Stress in Cotton: A Review on Predicted and Unpredicted Growth-Yield Anomalies and Mitigating Breeding Strategies. Agronomy 2021, 11, 1825. https://doi.org/10.3390/agronomy11091825
Majeed S, Rana IA, Mubarik MS, Atif RM, Yang S-H, Chung G, Jia Y, Du X, Hinze L, Azhar MT. Heat Stress in Cotton: A Review on Predicted and Unpredicted Growth-Yield Anomalies and Mitigating Breeding Strategies. Agronomy. 2021; 11(9):1825. https://doi.org/10.3390/agronomy11091825
Chicago/Turabian StyleMajeed, Sajid, Iqrar Ahmad Rana, Muhammad Salman Mubarik, Rana Muhammad Atif, Seung-Hwan Yang, Gyuhwa Chung, Yinhua Jia, Xiongming Du, Lori Hinze, and Muhammad Tehseen Azhar. 2021. "Heat Stress in Cotton: A Review on Predicted and Unpredicted Growth-Yield Anomalies and Mitigating Breeding Strategies" Agronomy 11, no. 9: 1825. https://doi.org/10.3390/agronomy11091825