
Ecological Intensification of Food Production by Integrating Forages

 ,
,  , , ,
, , ,  , and
, and 
Abstract
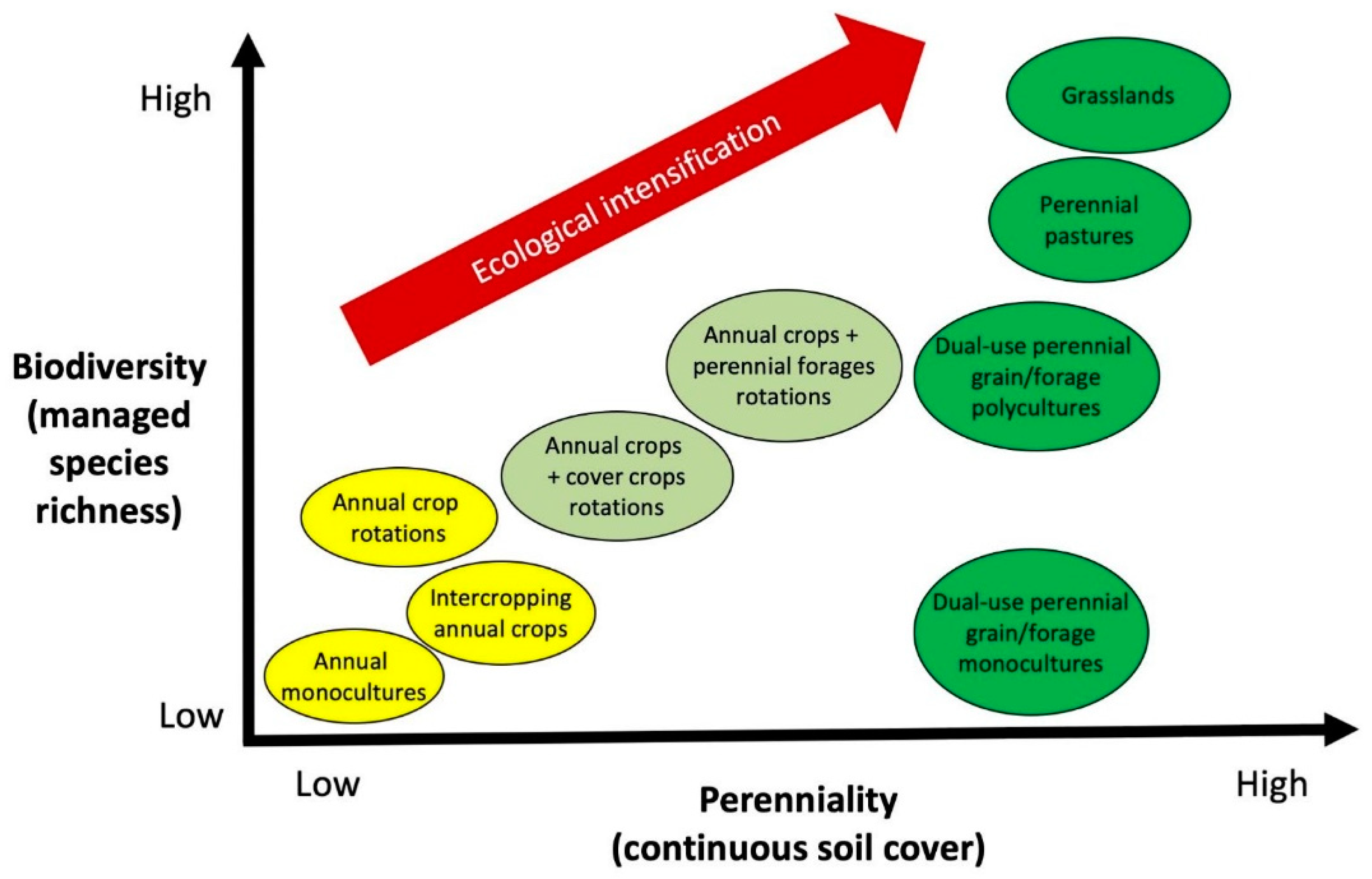
:1. Introduction
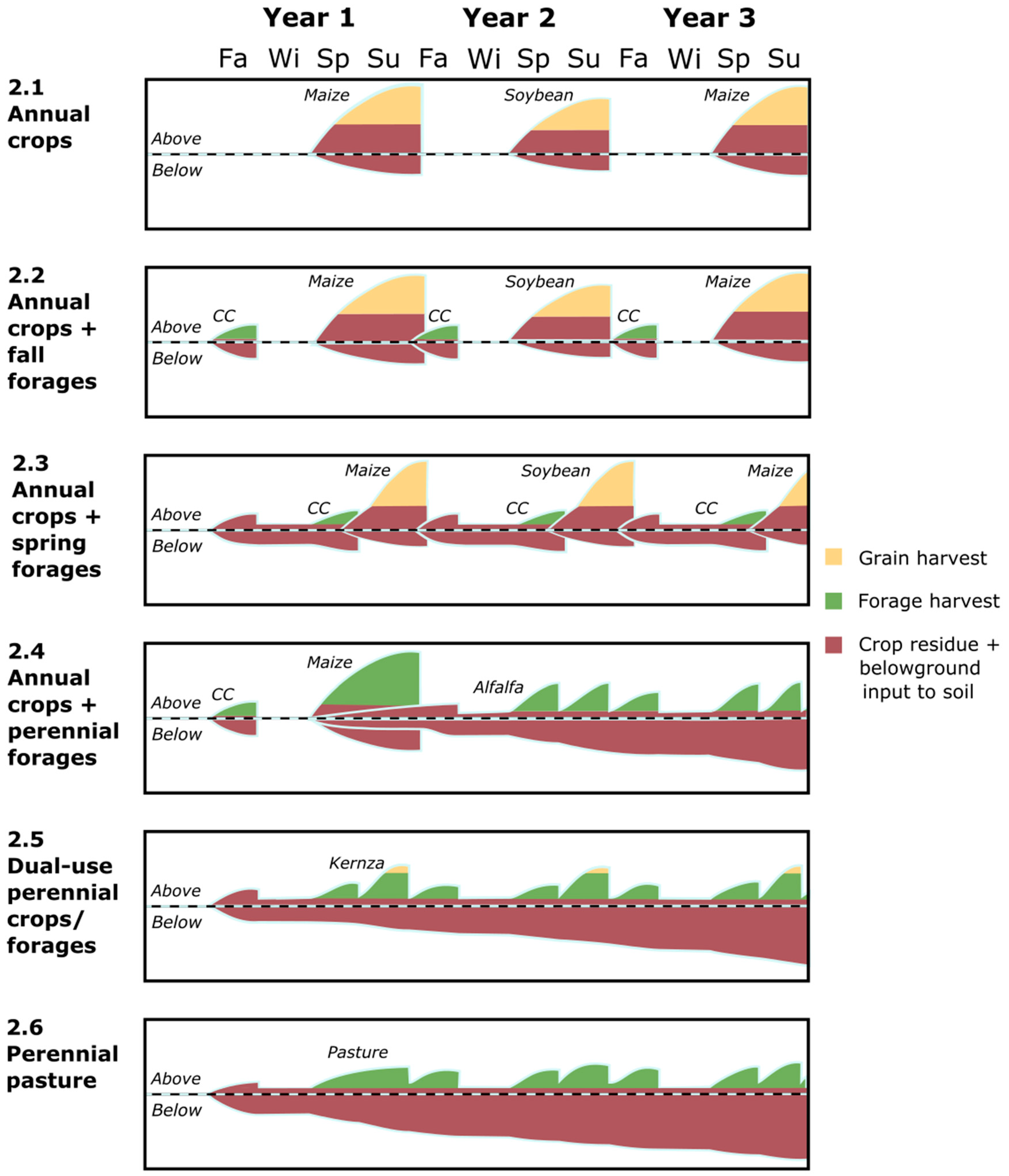
2. Annual Forages
2.1. Annual Forages after Cool-Season Cereals
2.2. Intercropped Annual Forages
2.3. Interseeded Annual Forages into Maize to Improve Grazing Value of Maize Stover
2.4. Winter Cover Crops for Spring Forage
2.5. Intercropping Annual Warm-Season Crops to Increase Value of Maize Silage
2.6. Annual Forages Seeded into Perennial Pasture
2.7. The Role of Annual Forages in Achieving Ecological Intensification
2.8. Annual Forages: Limitations and Knowledge Gaps
3. Perennial Forages
3.1. Perennial Forages in Annual Crop Rotations
3.2. Perennial Forages Intercropped or Interseeded with Annual Crops
3.3. The Role of Perennial Forages in Achieving Ecological Intensification
3.4. Perennial Forages: Limitations and Knowledge Gaps
4. Dual-Use Perennial Crops/Forages
4.1. Dual-Use Cereals
4.2. Dual-Use Oilseeds
4.3. The Role of Dual-Use Crops/Forages in Achieving Ecological Intensification
4.4. Dual-Use Crops/Forages: Limitations and Knowledge Gaps
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Disclaimer
References
- Van Dijk, M.; Morley, T.; Rau, M.L.; Saghai, Y. A meta-analysis of projected global food demand and population at risk of hunger for the period 2010–2050. Nat. Food 2021, 2, 494–501. [Google Scholar] [CrossRef]
- Alexandratos, N.; Bruinsma, J. World Agriculture toward 2030/2050: The 2012 Revision. Food and Agriculture Organization of the United Nations. ESA Working Paper no. 12-03. Available online: http://www.fao.org/3/ap106e/ap106e.pdf (accessed on 1 July 2021).
- United States Department of Agriculture. USDA Engages Public for Input on Agriculture Innovation Agenda (Press Release No. 0210.20). Available online: https://www.usda.gov/media/press-releases/2020/04/01/usda-engages-public-input-agriculture-innovation-agenda (accessed on 1 July 2021).
- Kremen, C.; Merenlender, A.M. Landscapes that work for biodiversity and people. Science 2018, 362, eaau6020. [Google Scholar] [CrossRef] [Green Version]
- Sanford, G.R.; Jackson, R.D.; Booth, E.G.; Hedtcke, J.L.; Picasso, V. Perenniality and diversity drive output stability and resilience in a 26-year cropping systems experiment. Field Crop. Res. 2021, 263, 108071. [Google Scholar] [CrossRef]
- Smith, P. Delivering food security without increasing pressure on land. Glob. Food Secur. 2013, 2, 18–23. [Google Scholar] [CrossRef]
- Pretty, J.; Bharucha, Z.P. Sustainable intensification in agricultural systems. Ann. Bot. 2014, 114, 1571–1596. [Google Scholar] [CrossRef] [PubMed]
- Royal Society. Reaping the Benefits: Science and the Sustainable Intensification of Global Agricultur; The Royal Society: London, UK, 2009. [Google Scholar]
- Godfray, H.C.J.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Pretty, J.; Robinson, S.; Thomas, S.M.; Toulmin, C. Food security: The challenge of feeding 9 billion people. Science 2010, 327, 812–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pretty, J. Intensification for redesigned and sustainable agricultural systems. Science 2018, 362, eaav0294. [Google Scholar] [CrossRef] [Green Version]
- Spiegal, S.; Bestelmeyer, B.T.; Archer, D.W.; Augustine, D.J.; Boughton, E.H.; Cavigelli, M.A.; Clark, P.E.; Derner, J.D.; Duncan, E.W.; Hapeman, C.J.; et al. Evaluating strategies for sustainable intensification of US agriculture through the Long-Term Agroecosystem Research Network. Environ. Res. Lett. 2018, 13, 034031. [Google Scholar] [CrossRef]
- Tittonell, P. Ecological intensification of agriculture-sustainable by nature. Curr. Opin. Environ. Sustain. 2014, 8, 53–61. [Google Scholar] [CrossRef]
- Barrios, E.; Herren, B.G.; Bicksler, A.; Siliprandi, E.; Brathwaite, R.; Moller, S.; Batello, C.; Tittonell, P. The 10 elements of Agroecology; enabling transitions towards sustainable agriculture and food systems through visual narratives. Ecosyst. People 2020, 16, 230–247. [Google Scholar] [CrossRef]
- Wezel, A.; Herren, B.G.; Kerr, R.B.; Barrios, E.; Goncalves, A.L.R.; Sinclair, F. Agroecological principles and elements and their implications for transitioning to sustainable food systems. A review. Agron. Sustain. Dev. 2020, 40, 40. [Google Scholar] [CrossRef]
- Crews, T.; Cox, T.; De Haan, L.; Damaraju, S.; Jackson, W.; Nabukalu, P.; Van Tassel, D.; Wang, S. New roots for ecological intensification. CSA News 2014, 59, 16–18. [Google Scholar] [CrossRef]
- Ausubel, J.H.; Wernick, I.K.; Waggoner, P.E. Peak farmland and the prospect for land sparing. Popul. Dev. Rev. 2013, 38, 221–242. [Google Scholar] [CrossRef] [Green Version]
- Ustaoglu, E.; Williams, B. Determinants of urban expansion and agricultural land conversion in 25 EU countries. Environ. Manage. 2017, 60, 717–746. [Google Scholar] [CrossRef]
- Entz, M.H.; Bullied, W.J.; Katepa-Mupondwa, F. Rotational benefits of forage crops in Canadian prairie cropping systems. J. Prod. Agric. 1995, 8, 521–529. [Google Scholar] [CrossRef]
- Cates, A.M.; Ruark, M.D.; Hedtcke, J.L.; Posner, J.L. Long-term tillage, rotation and perennialization effects on particulate and aggregate soil organic matter. Soil Tillage Res. 2016, 155, 371–380. [Google Scholar] [CrossRef]
- Franco, J.G.; Duke, S.E.; Hendrickson, J.R.; Liebig, M.A.; Archer, D.W.; Tanaka, D.L. Spring wheat yields following perennial forages in a semiarid no-till cropping system. Agron. J. 2018, 110, 2408–2416. [Google Scholar] [CrossRef]
- Sprunger, C.D.; Martin, T.; Mann, M. Systems with greater perenniality and crop diversity enhance soil biological health. Agric. Environ. Lett. 2020, 5, e20030. [Google Scholar] [CrossRef]
- Reynolds, J.; Bell, M.A.; Grace, J.; Gratton, C.; Jackson, R.D.; Keeley, K.O.; Mayerfeld, D. An agroecological vision of perennial agriculture. Agroecol. Sustain. Food Syst. 2021, 45, 1470–1479. [Google Scholar] [CrossRef]
- Snapp, S.S.; Swinton, S.M.; Labarta, R.; Mutch, D.; Black, J.R.; Leep, R.; Nyiraneza, J.; O’Neil, K. Evaluating cover crops for benefits, costs and performance within cropping systems niches. Agron. J. 2005, 97, 322–332. [Google Scholar] [CrossRef]
- Blanco-Canqui, H.; Holman, J.D.; Schlegel, A.J.; Tatarko, J.; Shaver, T.M. Replacing fallow with cover crops in a semiarid soil: Effects on soil properties. Soil Sci. Soc. Am. J. 2013, 77, 1026–1034. [Google Scholar] [CrossRef] [Green Version]
- Schipanski, M.E.; Barbercheck, M.; Douglas, M.R.; Finney, D.M.; Haider, K.; Kaye, J.P.; Kemanian, A.R.; Mortensen, D.A.; Ryan, M.R.; Tooker, J.; et al. A framework for evaluating ecosystem services provided by cover crops in agroecosystems. Agric. Syst. 2014, 125, 12–22. [Google Scholar] [CrossRef]
- Florence, A.M.; Higley, L.G.; Drijber, R.A.; Francis, C.A.; Lindquist, J.L. Cover crop mixture diversity, biomass productivity, weed suppression, and stability. PLoS ONE 2019, 14, e0206195. [Google Scholar] [CrossRef] [Green Version]
- Mallinger, R.E.; Franco, J.G.; Prischmann-Voldseth, D.A.; Prasifka, J.R. Annual cover crops for managed and wild bees: Optimal plant mixtures depend on pollinator enhancement goals. Agric. Ecosyst. Environ. 2019, 273, 107–116. [Google Scholar] [CrossRef]
- Sanderson, M.; Johnson, H.; Hendrickson, J. Cover crop mixtures grown for annual forage in a semi-arid environment. Agron. J. 2018, 110, 525–534. [Google Scholar] [CrossRef]
- Planisich, A.; Utsumi, S.A.; Larripa, M.; Galli, J.R. Grazing of cover crops in integrated crop-livestock systems. Animal 2021, 15, 100054. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Abagandura, G.O.; Kumar, S. Short-term grazing of cover crops and maize residue impacts on soil greenhouse gas fluxes in two Mollisols. J. Environ. Qual. 2020, 49, 628–639. [Google Scholar] [CrossRef] [Green Version]
- Kelly, C.; Schipanski, M.E.; Tucker, A.; Trujillo, W.; Holman, J.D.; Obour, A.K.; Johnson, S.K.; Brummer, J.E.; Haag, L.; Fonte, S.J. Dryland cover crop soil health benefits are maintained with grazing in the U.S. High and Central Plains. Agric. Ecosyst. Environ. 2021, 313, 107358. [Google Scholar] [CrossRef]
- Hilimire, K. Integrated crop/livestock agriculture in the United States: A review. J. Sustain. Agric. 2011, 35, 376–393. [Google Scholar] [CrossRef]
- Ryschawy, J.; Choisis, N.; Choisis, J.P.; Joannon, A.; Gibon, A. Mixed crop-livestock systems: An economic and environmental-friendly way of farming? Animal 2012, 6, 1722–1730. [Google Scholar] [CrossRef] [Green Version]
- Lemaire, G.; Franzluebbers, A.; de Faccio Carvalho, P.C.; Dedieu, B. Integrated crop-livestock systems: Strategies to achieve synergy between agricultural production and environmental quality. Agric. Ecosyst. Environ. 2014, 190, 4–8. [Google Scholar] [CrossRef]
- Sulc, R.M.; Franzluebbers, A.J. Exploring integrated crop-livestock systems in different ecoregions of the United States. Eur. J. Agron. 2014, 57, 21–30. [Google Scholar] [CrossRef]
- Hendrickson, J.R. Crop-livestock integrated systems for more sustainable agricultural production: A review. CABI Rev. 2020, 15, 1–11. [Google Scholar] [CrossRef]
- Schut, A.G.T.; Cooledge, E.C.; Moraine, M.; van de Ven, G.W.J.; Jones, D.L.; Chadwick, D.R. Reintegration of crop-livestock systems in Europe: An overview. Front. Agric. Sci. Eng. 2020, 8, 111–129. [Google Scholar] [CrossRef]
- Smart, A.J.; Redfearn, D.; Mitchell, R.; Wang, T.; Zilverberg, C.; Bauman, P.J.; Derner, J.D.; Walker, J.; Wright, C. Forum: Integration of crop-livestock systems: An opportunity to protect grasslands from conversion to cropland in the US Great Plains. Rangel. Ecol. Manag. 2020, in press. [Google Scholar] [CrossRef]
- Henchion, M.; McCarthy, M.; Resconi, V.C.; Troy, D. Meat consumption: Trends and quality matters. Meat Sci. 2014, 98, 561–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoll-Kleemann, S.; O’Riordan, T. The sustainability challenges of our meat and dairy diets. Environ. Sci. Policy Sustain. Dev. 2015, 57, 34–48. [Google Scholar] [CrossRef]
- Rockström, J.; Steffen, W.; Noone, K.; Persson, A.; Chapin, F.S., III; Lambin, E.F.; Lenton, T.M.; Scheffer, M.; Folke, C.; Schellnhuber, H.J.; et al. A safe operating space for humanity. Nature 2009, 461, 472–475. [Google Scholar] [CrossRef]
- Thorup-Kristensen, K.; Halberg, N.; Nicolaisen, M.; Olesen, J.E.; Crews, T.E.; Hinsinger, P.; Kirkegaard, J.; Pierret, A.; Dresboll, D.B. Digging deeper for agricultural resources, the value of deep rooting. Trends Plant Sci. 2020, 25, 406–417. [Google Scholar] [CrossRef] [Green Version]
- Heaton, E.A.; Schulte, L.A.; Berti, M.; Langeveld, H.; Zegada-Lizarazu, W.; Parrish, D.; Monti, A. Managing a second-generation crop portfolio through sustainable intensification: Examples from the USA and the EU. Biofuels Bioprod. Biorefin. 2013, 7, 701–714. [Google Scholar] [CrossRef]
- Drewnoski, M.; Parsons, J.; Blanco, H.; Redfearn, D.; Hales, K.; MacDonald, J. Forages and pastures symposium: Cover crops in livestock production: Whole-system approach. Can cover crops pull double duty: Conservation and profitable forage production in the Midwestern United States? J. Anim. Sci. 2018, 96, 3503–3512. [Google Scholar] [CrossRef] [PubMed]
- Andersen, B.J.; Samarappuli, D.P.; Wick, A.; Berti, M.T. Faba bean and pea can provide late-fall forage grazing without affecting maize yield the following season. Agronomy 2020, 10, 80. [Google Scholar] [CrossRef] [Green Version]
- Tobin, C.; Singh, S.; Kumar, S.; Wang, T.; Sexton, P. Demonstrating Short-term impacts of grazing and cover crops on soil health and economic benefits in an integrated crop-livestock system in South Dakota. Open J. Soil Sci. 2020, 10, 109–136. [Google Scholar] [CrossRef] [Green Version]
- Samarappuli, D.P.; Johnson, B.L.; Kandel, H.; Berti, M.T. Biomass yield and nitrogen content of annual energy/forage crops preceded by cover crops. Field Crop. Res. 2014, 167, 31–39. [Google Scholar] [CrossRef]
- Li, L.; Sun, J.; Zhang, F.; Guo, T.; Smith, F.A.; Smith, S.E. Root distribution and interactions between intercropped species. Oecologia 2006, 147, 280–290. [Google Scholar] [CrossRef]
- Hauggaard-Nielsen, H.; Jornsgaard, B.; Kinane, J.; Jensen, E.S. Grain legume-cereal intercropping: The practical application of diversity, competition and facilitation in arable and organic cropping systems. Renew. Agric. Food Syst. 2007, 23, 3–12. [Google Scholar] [CrossRef] [Green Version]
- Strydhorst, S.; King, J.R.; Lopetinsky, K.J.; Harker, K.N. Forage potential of intercropping barley with faba bean, lupin, or field pea. Agron. J. 2008, 100, 182–190. [Google Scholar] [CrossRef]
- Anil, L.; Park, J.; Phipps, R.H.; Miller, F.A. Temperate intercropping of cereal for forage: A review of the potential for growth and utilization with particular reference to the UK. Grass Forage Sci. 1998, 53, 301–317. [Google Scholar] [CrossRef]
- Sarunaite, L.; Deveikyte, I.; Arlauskiene, A.; Kadziuliene, Z.; Maiksteniene, S. Pea and spring cereal intercropping systems: Advantages and suppression of broad-leaved weeds. Pol. J. Environ. Stud. 2013, 22, 541–551. [Google Scholar]
- Zhang, G.; Zhang, C.; Yang, Z.; Dong, S. Root distribution and N acquisition in an alfalfa and corn intercropping system. J. Agric. Sci. 2013, 5, 128. [Google Scholar] [CrossRef]
- Hauggaard-Nielsen, H.; Ambus, P.; Jensen, E.S. Interspecific competition, N use and interference with weeds in pea-barley intercropping. Field Crop. Res. 2001, 70, 101–109. [Google Scholar] [CrossRef]
- Lauriault, L.M.; Guldan, S.J.; Popiel-Powers, F.G.; Steiner, R.L.; Martin, C.A.; Heyduck, R.F.; Falk, C.L.; Petersen, M.K.; May, T. Relay intercropping with cover crops improved autumn forage potential of sweet maize stover. Agriculture 2018, 8, 103. [Google Scholar] [CrossRef] [Green Version]
- Villalobos, L.; Brummer, J.E. Interseeding cool-season forages into corn to increase yield and quality of residue grazed in the fall. Proc. West. Sect. Am. Soc. Anim. Sci. 2015, 66, 268–271. [Google Scholar]
- Dabney, S.M.; Delgado, J.A.; Reeves, D.W. Using winter cover crops to improve soil and water quality. Commun. Soil Sci. Plant Anal. 2001, 32, 1221–1250. [Google Scholar] [CrossRef]
- Shah, S.; Hookway, S.; Pullen, H.; Clarke, T.; Wilkinson, S.; Reeve, V.; Fletcher, J.M. The role of cover crops in reducing nitrate leaching and increasing soil organic matter. Asp. Appl. Biol. 2017, 134, 243–251. [Google Scholar]
- Everett, L.A.; Wilson, M.L.; Pepin, R.J.; Coulter, J.A. Winter rye cover crop with liquid manure injection reduces spring soil nitrate but not maize yield. Agronomy 2019, 9, 852. [Google Scholar] [CrossRef] [Green Version]
- Coblentz, W.K.; Akins, M.S.; Kalscheur, K.F.; Brink, G.E.; Cavadini, J.S. Effects of growth stage and growing degree day accumulations on triticale forages: 1. Dry matter yield, nutritive value, and in vitro dry matter disappearance. J. Dairy Sci. 2018, 101, 8965–8985. [Google Scholar] [CrossRef]
- Coblentz, W.K.; Akins, M.S.; Kalscheur, K.F.; Brink, G.E.; Cavadini, J.S. Effects of growth stage and growing degree day accumulations on triticale forages: 2. In vitro disappearance on neutral detergent fiber. J. Dairy Sci. 2018, 101, 8986–9003. [Google Scholar] [CrossRef]
- Landry, E.; Janovicek, K.; Lee, E.A.; Deen, W. Winter cereal cover crops for spring forage in temperate climates. Agron. J. 2019, 111, 217–223. [Google Scholar] [CrossRef] [Green Version]
- Adhikari, L.; Mohseni-Moghadam, M.; Missaoui, A. Allelopathic effects of cereal rye on weed suppression and forage yield in alfalfa. Am. J. Plant Sci. 2018, 9, 685–700. [Google Scholar] [CrossRef] [Green Version]
- Samarappuli, D.; Berti, M.T. Intercropping forage sorghum with maize is a promising alternative to maize silage for biogas production. J. Clean. Prod. 2018, 194, 515–524. [Google Scholar] [CrossRef]
- La Guardia Nave, R.; Corbin, M.D. Forage warm-season legumes and grasses intercropped with corn as an alternative for corn silage production. Agronomy 2018, 8, 199. [Google Scholar] [CrossRef] [Green Version]
- Abdin, O.; Bruce, E.; Coulman, D.; Cloutier, M.A.; Faris, J.; Zhou, X.; Smith, D.L. Yield and yield components of maize interseeded with cover crops. Agron. J. 1998, 90, 63–68. [Google Scholar] [CrossRef]
- Parr, M.; Grossman, J.M.; Reberg-Horton, S.C.; Brinton, C.; Crozier, C. Nitrogen delivery from legume cover crops in no-till organic maize production. Agron. J. 2011, 103, 1578–1590. [Google Scholar] [CrossRef]
- Kallenbach, R.L.; Crawford, R.J., Jr.; Massie, M.D.; Kerley, M.S.; Bailey, N.J. Integrating bermudagrass into tall fescue-based pasture systems for stocker cattle. J. Anim. Sci. 2012, 90, 387–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stuedemann, J.A.; Hoveland, C.S. Fescue endophyte: History and impact on animal agriculture. J. Prod. Agric. 1988, 1, 39–44. [Google Scholar] [CrossRef]
- Utley, P.R.; Marchant, W.H.; McCormick, W.C. Evaluation of annual grass forages in prepared seedbeds and overseeded into perennial sods. J. Anim. Sci. 1976, 42, 16–20. [Google Scholar] [CrossRef]
- Moyer, J.L.; Coffey, K.P. Forage quality and production of small grains interseeded into bermudagrass sod or grown in monoculture. Agron. J. 2000, 92, 748–753. [Google Scholar] [CrossRef]
- Coffey, K.P.; Coblentz, W.K.; Montgomery, T.G.; Shockey, J.D.; Bryant, K.J.; Francis, P.B.; Rosenkrans, C.F., Jr.; Gunter, S.A. Growth performance of stocker calves backgrounded on sod-seeded winter annuals or hay and grain. J. Animal Sci. 2002, 80, 926–932. [Google Scholar] [CrossRef] [Green Version]
- Beck, P.A.; Gunter, S.A.; Lusby, K.S.; West, C.P.; Watkins, K.B.; Hubbell, D.S., III. Animal performance and economic comparison of novel and toxic endophyte tall fescues to cool-season annuals. J. Anim. Sci. 2008, 86, 2043–2055. [Google Scholar] [CrossRef]
- Gunter, S.A.; Lusby, K.S.; Hubbell, D.S., III. Tall fescue for backgrounding in preparation for small-grain or Bermudagrass pasture. Prof. Anim. Sci. 2005, 21, 93–96. [Google Scholar] [CrossRef]
- Moyer, J.L.; Coffey, K.P.; Brazle, F.K.; Schneider, J.E.; Coppock, C.E.; DelCurto, T. Forage intake, diet quality, and performance of stocker cattle grazing interseeded or monocultured rye and bermudagrass. Prof. Anim. Sci. 1995, 11, 67–73. [Google Scholar] [CrossRef]
- Ball, D.M.; Hoveland, C.S.; Lacefield, G.D. Southern Forages, 3rd ed.; Potash and Phosphate Institute and the Foundation for Agronomic Research: Norcross, GA, USA, 2002. [Google Scholar]
- Hoveland, C.S.; Anthony, W.B.; McGuire, J.A.; Starling, J.G. Beef cow-calf performance on coastal bermudagrass overseeded with winter annual clovers and grasses. Agron. J. 1978, 70, 418–420. [Google Scholar] [CrossRef]
- Gunter, S.A.; Whitworth, W.A.; Montgomery, T.G.; Beck, P.A. Cool-season annual pastures with clovers to supplement wintering beef cows nursing calves. J. Anim. Sci. Biotechnol. 2012, 3, 25. [Google Scholar] [CrossRef] [Green Version]
- Hoveland, C.S.; Evers, G.W. Arrowleaf, crimson and other annual clovers. Forages 1995, 1, 249–260. [Google Scholar]
- Beck, P.A.; Hubbell, D.S.; Watkins, K.B.; Gunter, S.A.; Daniels, L.B. Performance of stocker cattle grazing cool-season annual grass mixtures in northern Arkansas. Prof. Anim. Sci. 2005, 21, 465–473. [Google Scholar] [CrossRef]
- Hill, G.M.; Utley, P.R.; McCormick, W.C. Evaluation of cow-calf systems using ryegrass sod-seeded in perennial pastures. J. Anim. Sci. 1985, 61, 1088–1094. [Google Scholar] [CrossRef]
- Roberts, C.; Andrae, J. Public Education on Tall Fescue Toxicosis. Neotyphodium in Cool-Season Grasses. 2005. Available online: https://onlinelibrary.wiley.com/doi/10.1002/9780470384916.ch18 (accessed on 1 July 2021).
- Nieman, C.C.; Albrecht, K.A.; Schaefer, D.M. Temporal composition of alfalfa–grass pastures and productivity response of Holstein steers. Agron. J. 2019, 111, 686–693. [Google Scholar] [CrossRef]
- Minson, D.J. Forage in Ruminant Nutrition; Academic Press: San Diego, CA, USA, 1990. [Google Scholar]
- Lusby, K.S.; McMurphy, W.E.; Strasia, C.A.; Smith, S.C.; Muntz, S.H. Effects of fescue endophyte and interseeded clovers on subsequent finishing performance of steers. J. Prod. Agric. 1990, 3, 103–105. [Google Scholar] [CrossRef]
- McMurphy, W.E.; Lusby, K.S.; Smith, S.C.; Muntz, S.H.; Strasia, C.A. Steer performance on tall fescue pasture. J. Prod. Agric. 1990, 3, 100–102. [Google Scholar] [CrossRef]
- Beck, P.A.; Haque, M.; Biermacher, J.T.; Hopkins, A.A.; Hubbell, D.S.; Hess, T. Impact of clover additions to toxic or nontoxic endophyte-infected tall fescue on animal performance and economics of stocker programs. Prof. Anim. Sci. 2012, 28, 433–442. [Google Scholar] [CrossRef] [Green Version]
- Hoveland, C.S.; McCann, M.A.; Bouton, J.H. Influence of endophyte, alfalfa, and grazing pressure on steer performance and plant persistence of Jesup tall fescue. J. Prod. Agric. 1997, 10, 546–550. [Google Scholar] [CrossRef]
- Melchior, E.A.; Myer, P.R. Fescue toxicosis and its influence on the rumen microbiome: Mitigation of production losses through clover isoflavones. J. Appl. Anim. Res. 2018, 46, 1280–1288. [Google Scholar] [CrossRef] [Green Version]
- Rhodes, M.; Paterson, J.; Kerley, M.; Garner, H.; Laughlin, M. Reduced blood flow to peripheral and core body tissues in sheep and cattle induced by endophyte-infected tall fescue. J. Anim. Sci. 1991, 69, 2033–2043. [Google Scholar] [CrossRef]
- Aiken, G.; Piper, E.; Miesner, C. Influence of protein supplementation and implant status on alleviating fescue toxicosis. J. Anim. Sci. 2001, 79, 827–832. [Google Scholar] [CrossRef] [Green Version]
- Nevala, R.; Korpela, R.; Vapaatalo, H. Plant derived estrogens relax rat mesenteric artery in vitro. Life Sci. 1998, 63, 95–100. [Google Scholar] [CrossRef]
- Simoncini, T.; Fornari, L.; Mannella, P.; Caruso, A.; Garibaldi, S.; Baldacci, C.; Genazzani, A.R. Activation of nitric oxide synthesis in human endothelial cells by red clover extracts. Menopause 2005, 12, 69–77. [Google Scholar] [CrossRef]
- Jia, Y.; Harmon, D.L.; Flythe, M.D.; Klotz, J.L. Interaction of isoflavones and endophyte-infected tall fescue seed extract on vasoactivity of bovine mesenteric vasculature. Front. Nutr. 2015, 2, 1–10. [Google Scholar] [CrossRef]
- Aiken, G.E.; Flythe, M.D.; Kagan, I.A.; Ji, H.; Bush, L.P. Mitigation of ergot vasoconstriction by clover isoflavones in goats (Capra hircus). Front. Vet. Sci. 2016, 3, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.; Yang, Z.; Dong, S. Interspecific competitiveness affects the total biomass yield in an alfalfa and corn intercropping system. Field Crop. Res. 2011, 124, 66–73. [Google Scholar] [CrossRef]
- Franco, J.G.; King, S.R.; Volder, A. Component crop physiology and water use efficiency in response to intercropping. Eur. J. Agron. 2018, 93, 27–39. [Google Scholar] [CrossRef]
- Blanco-Canqui, H.; Ruis, S.J.; Proctor, C.A.; Creech, C.F.; Drewnoski, M.E.; Redfearn, D.D. Harvesting cover crops for biofuel and livestock production: Another ecosystem service? Agron. J. 2020, 112, 2373–2400. [Google Scholar] [CrossRef]
- Kaspar, T.C.; Jaynes, D.B.; Parkin, T.B.; Moorman, T.B.; Singer, J.W. Effectiveness of oat and rye cover crops in reducing nitrate losses in drainage water. Agric. Water Manage. 2012, 110, 25–33. [Google Scholar] [CrossRef]
- Kladivko, E.J.; Kaspar, T.C.; Jaynes, D.B.; Malone, R.W.; Singer, J.; Morin, X.K.; Searchinger, T. Cover crops in the upper midwestern United States: Potential adoption and reduction of nitrate leaching in the Mississippi River Basin. J. Soil Water Conserv. 2014, 69, 279–291. [Google Scholar] [CrossRef] [Green Version]
- Blanco-Canqui, H. Cover crops and water quality. Agron. J. 2018, 110, 1633–1647. [Google Scholar] [CrossRef]
- Peterson, A.T.; Berti, M.T.; Samarappuli, D. Intersowing cover crops into standing soybean in the US upper Midwest. Agronomy 2019, 9, 264. [Google Scholar] [CrossRef] [Green Version]
- Morris, A.H.; Isbell, S.A.; Saha, D.; Kaye, J.P. Mitigating nitrogen pollution with under-sown legume-grass cover crop mixtures in winter cereals. J. Environ. Qual. 2020, 50, 324–335. [Google Scholar] [CrossRef] [PubMed]
- Wall, G.J.; Pringle, E.A.; Sheard, R.W. Intercropping red clover with silage maize for soil erosion control. Can. J. Soil Sci. 1991, 71, 137–145. [Google Scholar] [CrossRef]
- Mohammed, Y.A.; Patel, S.; Matthees, H.L.; Lenssen, A.W.; Johnson, B.L.; Wells, M.S.; Forcella, F.; Berti, M.T.; Gesch, R.W. Soil nitrogen in response to interseeded cover crops in maize-soybean production systems. Agronomy 2020, 10, 1439. [Google Scholar] [CrossRef]
- Cecchin, A.; Pourhashem, G.; Gesch, R.W.; Lenssen, A.W.; Mohammed, Y.A.; Patel, S.; Berti, M.T. Environmental trade-offs of relay-cropping winter cover crops with soybean in a maize-soybean cropping system. Agric. Syst. 2021, 189, 103062. [Google Scholar] [CrossRef]
- Schulz, V.S.; Schumann, C.; Weisenburger, S.; Müller-Lindenlauf, M.; Stolzenburg, K.; Möller, K. Row-intercropping maize (Zea mays L.) with biodiversity-enhancing flowering-partners-effect on plant growth, silage yield, and composition of harvest material. Agriculture 2020, 10, 524. [Google Scholar] [CrossRef]
- Gadberry, M.S.; Hawley, J.; Beck, P.A.; Jennings, J.A.; Kegley, E.B.; Coffey, K.P.; Bill, E. Kunkle interdisciplinary beef symposium: A meta-analysis of research efforts aimed at reducing the impact of fescue toxicosis on cattle weight gain and feed intake. J. Anim. Sci. 2015, 93, 5496–5505. [Google Scholar] [CrossRef]
- Gentile, R.M.; Martino, D.L.; Entz, M.H. Influence of perennial forages on subsoil organic carbon in a long-term rotation study in Uruguay. Agric. Ecosyst. Environ. 2005, 105, 419–423. [Google Scholar] [CrossRef]
- Gamble, A.V.; Howe, J.A.; Balkcom, K.B.; Wood, C.W.; DiLorenzo, N.; Watts, D.B.; van Santen, E. Soil organic carbon storage and greenhouse gas emissions in a grazed perennial forage–crop rotation system. Agrosyst. Geosci. Environ. 2019, 2, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Liebig, M.A.; Hendrickson, J.R.; Franco, J.G.; Archer, D.W.; Nichols, K.; Tanaka, D.L. Near-surface soil property responses to forage production in a semiarid region. Soil Sci. Soc. Am. J. 2018, 82, 223–230. [Google Scholar] [CrossRef]
- Ward, P.R.; Dunin, F.X.; Micin, S.F. Water use and root growth by annual and perennial pastures and subsequent crops in a phase rotation. Agric. Water Manag. 2002, 53, 83–97. [Google Scholar] [CrossRef]
- Tan, I.Y.S.; Van Es, H.M.; Duxbury, J.M.; Melkonian, J.J.; Schindelbeck, R.R.; Geohring, L.D.; Hively, W.D.; Moebius, B.N. Single-event nitrous oxide losses under maize production as affected by soil type, tillage, rotation, and fertilization. Soil Tillage Res. 2009, 102, 19–26. [Google Scholar] [CrossRef]
- Taylor, A.M.; Amiro, B.D.; Fraser, T.J. Net CO2 exchange and carbon budgets of a three-year crop rotation following conversion of perennial lands to annual cropping in Manitoba, Canada. Agric. For. Meteorol. 2013, 182, 67–75. [Google Scholar] [CrossRef]
- Tenuta, M.; Amiro, B.D.; Gao, X.; Wagner-Riddle, C.; Gervais, M. Agricultural management practices and environmental drivers of nitrous oxide emissions over a decade for an annual and an annual-perennial crop rotation. Agric. For. Meteorol. 2019, 276, 107636. [Google Scholar] [CrossRef]
- Wortman, S.E.; Francis, C.A.; Lindquist, J.L. Cover crop mixtures for the western Corn Belt: Opportunities for increased productivity and stability. Agron. J. 2012, 104, 699–705. [Google Scholar] [CrossRef] [Green Version]
- Ominski, P.D.; Entz, M.H.; Kenkel, N. Weed suppression by Medicago sativa in subsequent cereal crops: A comparative survey. Weed Sci. 1999, 47, 282–290. [Google Scholar] [CrossRef] [Green Version]
- Meiss, H.; Médiène, S.; Waldhardt, R.; Caneill, J.; Bretagnolle, V.; Reboud, X.; Munier-Jolain, N. Perennial lucerne affects weed community trajectories in grain crop rotations. Weed Res. 2010, 50, 331–340. [Google Scholar] [CrossRef]
- Dear, B.S.; Sandral, G.A.; Virgona, J.M.; Swan, A.D. Yield and grain protein of wheat following phased perennial grass, lucerne, and annual pastures. Aust. J. Agric. Res. 2004, 55, 775–785. [Google Scholar] [CrossRef]
- Clemensen, A.K.; Grusak, M.A.; Duke, S.E.; Hendrickson, J.R.; Franco, J.G.; Archer, D.W.; Roemmich, J.N.; Liebig, M.A. Perennial forages influence mineral and protein concentrations in annual wheat cropping systems. Crop. Sci. 2021, 61, 2080–2089. [Google Scholar] [CrossRef]
- Crews, T.E.; Rumsey, B.E. What agriculture can learn from native ecosystems in building soil organic matter: A review. Sustainability 2017, 9, 578. [Google Scholar] [CrossRef] [Green Version]
- Kantola, I.B.; Masters, M.D.; DeLucia, E.H. Soil particulate organic matter increases under perennial bioenergy crop agriculture. Soil Biol. Biochem. 2017, 113, 184–191. [Google Scholar] [CrossRef]
- Wood, S.A.; Tirfessa, D.; Baudron, F. Soil organic matter underlies crop nutritional quality and productivity in smallholder agriculture. Agric. Ecosyst. Environ. 2018, 266, 100–108. [Google Scholar] [CrossRef]
- Grover, K.K.; Karsten, H.D.; Roth, G.W. Corn grain yields and yield stability in four long-term cropping systems. Agron. J. 2009, 101, 940–946. [Google Scholar] [CrossRef]
- Nielsen, D.C.; Vigil, M.F. Wheat yield and yield stability of eight dryland crop rotations. Agron. J. 2018, 110, 594–601. [Google Scholar] [CrossRef] [Green Version]
- Fuentes, R.; Taliaferro, C. Biomass Yield Stability of Switchgrass Cultivars. Trends in New Crops and New Uses; Janick, J., Whipkey, A., Eds.; ASHS Press: Alexandria, VA, USA, 2002; pp. 276–282. Available online: https://hort.purdue.edu/newcrop/ncnu02/v5-276.html (accessed on 15 September 2021).
- Halling, M.A. Yield stability of Festulolium and perennial ryegrass in southern and central Sweden. Grassl. Sci. Eur. 2012, 17, 118–120. [Google Scholar]
- Olmstead, J.; Brummer, E.C. Benefits and barriers to perennial forage crops in Iowa corn and soybean rotations. Renew. Agric. Food Syst. 2008, 23, 97–107. [Google Scholar] [CrossRef]
- Martin, N.P.; Russelle, M.P.; Powell, J.M.; Sniffen, C.J.; Smith, S.I.; Tricarico, J.M.; Grant, R.J. Invited review: Sustainable forage and grain crop production for the US dairy industry. J. Dairy Sci. 2017, 100, 9479–9494. [Google Scholar] [CrossRef] [Green Version]
- Hall, M.H.; Hebrock, N.S.; Pierson, P.E.; Caddel, J.L.; Owens, V.N.; Sulc, R.M.; Undersander, D.J.; Whitesides, R.E. The effects of glyphosate-tolerant technology on reduced alfalfa seeding rates. Agron. J. 2010, 102, 911–916. [Google Scholar] [CrossRef]
- Stranger, T.F.; Lauer, J.G.; Chavas, J.-P. The profitability and risk of long-term cropping systems featuring different rotations and nitrogen rates. Agron. J. 2008, 100, 105–113. [Google Scholar]
- Grabber, J.H. Prohexadione-calcium improves stand density and yield of alfalfa interseeded into silage corn. Agron. J. 2016, 108, 726–735. [Google Scholar] [CrossRef]
- Osterholz, W.R.; Renz, M.J.; Lauer, J.G.; Grabber, J.H. Prohexadione-calcium rate and timing effects on alfalfa interseeded into silage corn. Agron. J. 2018, 110, 85–94. [Google Scholar] [CrossRef]
- Osterholz, W.R.; Dias, J.L.C.S.; Grabber, J.H.; Renz, M.J. PRE- and POST-applied herbicide options for alfalfa interseeded with corn silage. Weed Technol. 2021, 35, 263–270. [Google Scholar] [CrossRef]
- Grabber, J.H.; Smith, D.L.; Osterholz, W.R.; Renz, M. Establishment and first year yield of interseeded alfalfa as influenced by corn plant density and treatment with prohexadione, fungicide and insecticide. Agronomy 2021, 11, 2343. [Google Scholar] [CrossRef]
- Grabber, J.H.; Osterholz, W.R.; Riday, H.; Cassida, K.A.; Williamson, J.A.; Renz, M. Differential survival of alfalfa varieties interseeded into corn silage. Crop. Sci. 2021, 61, 1797–1808. [Google Scholar] [CrossRef]
- Berti, M.T.; Lukaschewsky, J.; Samarappuli, D.P. Intercropping alfalfa into silage maize can be more profitable than maize silage followed by spring-seeded alfalfa. Agronomy 2021, 11, 1196. [Google Scholar] [CrossRef]
- Berti, M.T.; Cecchin, A.; Samarappuli, D.P.; Patel, S.; Lenssen, A.W.; Moore, K.J.; Wells, S.S.; Kazula, M.J. Alfalfa established successfully in intercropping with corn in the US Midwest. Agronomy 2021, 11, 1676. [Google Scholar] [CrossRef]
- Osterholz, W.R.; Ruark, M.D.; Renz, M.; Grabber, J.H. Benefits of alfalfa interseeding include reduced residual soil nitrate pools following corn production. Agric. Environ. Lett. 2021, 6, e20053. [Google Scholar] [CrossRef]
- Grabber, J.H.; Jokela, W.E.; Lauer, J.G. Soil nitrogen and forage yields of corn grown with clover or grass companion crops and manure. Agron. J. 2014, 106, 952–961. [Google Scholar] [CrossRef] [Green Version]
- Osterholz, W.R.; Ruark, M.D.; Renz, M.J.; Grabber, J.H. Interseeding alfalfa into corn silage increases corn N fertilizer demand and increases system yield. Agron. Sustain. Dev. 2021, 41, 1–3. [Google Scholar] [CrossRef]
- Duiker, S.W.; Hartwig, N.L. Living mulches of legumes in imidazolinone-resistant corn. Agron. J. 2004, 96, 1021–1028. [Google Scholar] [CrossRef]
- Zemenchik, R.A.; Albrecht, K.A.; Boerboom, C.M.; Lauer, J.G. Maize production with kura clover as a living mulch. Agron. J. 2000, 92, 698–705. [Google Scholar] [CrossRef] [Green Version]
- Martin, R.C.; Greyson, P.R.; Gordon, R. Competition between corn and a living mulch. Can. J. Plant Sci. 1999, 79, 579–586. [Google Scholar] [CrossRef] [Green Version]
- Sanders, Z.P.; Andrews, J.S.; Saha, U.K.; Vencill, W.; Lee, R.D.; Hill, N.S. Optimizing Agronomic Practices for Clover Persistence and Corn Yield in a White Clover-Corn Living Mulch System. Agron. J. 2017, 109, 2025–2032. [Google Scholar] [CrossRef] [Green Version]
- Flynn, E.S.; Moore, K.J.; Singer, J.W.; Lamkey, K.R. Evaluation of grass and legume species as perennial ground covers in corn production. Crop. Sci. 2013, 53, 611–620. [Google Scholar] [CrossRef]
- Bartel, C.A.; Banik, C.; Lenssen, A.W.; Moore, K.J.; Laird, D.A.; Archontoulis, S.V.; Lamkey, K.R. Establishment of perennial groundcovers for maize-based bioenergy production systems. Agron. J. 2017, 109, 822–835. [Google Scholar] [CrossRef] [Green Version]
- Schlautman, B.; Bartel, C.; Diaz-Garcia, L.; Fei, S.; Flynn, S.; Haramoto, E.; Moore, K.J.; Raman, D.R. Perennial groundcovers: An emerging technology for soil conservation and the sustainable intensification of agriculture. Emerg. Top. Life Sci. 2021, 5, 337–347. [Google Scholar] [CrossRef]
- Dobbratz, M.; Baker, J.M.; Grossman, J.; Wells, M.S.; Ginakes, P. Rotary zone tillage improves corn establishment in a kura clover living mulch. Soil Tillage Res. 2019, 189, 229–235. [Google Scholar] [CrossRef]
- Alexander, J.R.; Baker, J.M.; Venterea, R.T.; Coulter, J.A. Kura clover living mulch reduces fertilizer N requirements and increases profitability of maize. Agronomy 2019, 9, 432. [Google Scholar] [CrossRef] [Green Version]
- Andrews, J.S.; Sanders, Z.P.; Cabrera, M.L.; Saha, U.K.; Hill, N.S. Nitrogen dynamics in living mulch and annual cover crop corn production systems. Agron. J. 2018, 110, 1309–1317. [Google Scholar] [CrossRef] [Green Version]
- Ziyomo, C.; Albrecht, K.A.; Baker, J.M.; Bernardo, R. Corn performance under managed drought stress and in a kura clover living mulch intercropping system. Agron J. 2013, 105, 579–586. [Google Scholar] [CrossRef] [Green Version]
- Carter, P.R.; Sheaffer, C.C. Alfalfa response to soil water deficits. Growth, forage quality, yield, water use, and water-use efficiency. Crop. Sci. 1983, 23, 669–675. [Google Scholar] [CrossRef]
- Sulc, R.M.; Tracy, B.F. Integrated crop-livestock systems in the US Maize Belt. Agron. J. 2007, 99, 335–345. [Google Scholar] [CrossRef] [Green Version]
- Power, A.G. Ecosystem Services and Agriculture: Tradeoffs and Synergies. 2010. Available online: http://rstb.royalsocietypublishing.org/content/365/1554/2959.short (accessed on 14 September 2021).
- Dell, C.J.; Gollany, H.T.; Adler, P.R.; Skinner, R.H.; Polumsky, R.W. Implications of observed and simulated soil carbon sequestration for management options in maize-based rotations. J. Environ. Qual. 2018, 47, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Osterholz, W.R.; Renz, M.J.; Jokela, W.E.; Grabber, J.H. Interseeded alfalfa reduces soil and nutrient runoff losses during and after corn silage production. J. Soil Water Conser. 2019, 74, 85–90. [Google Scholar] [CrossRef]
- Niu, Y.; Luo, Z.; Cai, L.; Coulter, J.A.; Zhang, Y.; Berti, M. Continuous monoculture of alfalfa and annual crops influence soil organic matter and microbial communities in the rainfed loess plateau of China. Agronomy 2020, 10, 1054. [Google Scholar] [CrossRef]
- Wu, S.; Wu, P.; Feng, H.; Merkley, G.P. Effects of alfalfa coverage on runoff, erosion and hydraulic characteristics of overland flow on loess slope plots. Front. Environ. Sci. Eng. China 2011, 5, 76–83. [Google Scholar] [CrossRef]
- Siller, A.R.; Albrecht, K.A.; Jokela, W.E. Soil erosion and nutrient runoff in corn silage production with kura clover living mulch and winter rye. Agron. J. 2016, 108, 989–999. [Google Scholar] [CrossRef]
- Grabber, J.H.; Jokela, W.E. Off-season groundcover and runoff characteristics of perennial clover and annual grass companion crops for no-till corn fertilized with manure. J. Soil Water Conserv. 2013, 68, 411–418. [Google Scholar] [CrossRef]
- Kleinman, P.J.A.; Salon, P.; Sharpley, A.N.; Saporito, L.S. Effect of cover crops established at time of corn planting on phosphorus runoff from soils before and after dairy manure application. J. Soil Water Conserv. 2005, 60, 311–322. [Google Scholar]
- Ochsner, T.E.; Albrecht, K.A.; Schumacher, T.W.; Baker, J.M.; Berkevich, R.J. Water balance and nitrate leaching under corn in kura clover living mulch. Agron. J. 2010, 102, 1169–1178. [Google Scholar] [CrossRef]
- Osterholz, W.R.; Renz, M.J.; Grabber, J.H. Alfalfa establishment by interseeding with silage corn projected to increase profitability of corn silage-alfalfa rotations. Agron. J. 2020, 112, 4120–4132. [Google Scholar] [CrossRef]
- Entz, M.; Baron, V.; Carr, P.M.; Meyer, D.W.; Smith, S.R.; McCaughey, W.P. Potential of forages to diversify cropping systems in the Northern Great Plains. Agron. J. 2002, 94, 240–250. [Google Scholar] [CrossRef]
- Hendrickson, J.R.; Tanaka, D.L.; Liebig, M.A. Tillage and grazing impact on annual crop yields following conversion from perennial grass to annual crops. Crop. Manag. 2014, 13, 1–7. [Google Scholar] [CrossRef]
- Schmer, M.R.; Hendrickson, J.R.; Liebig, M.A.; Johnson, H.A. Perennial plant establishment and productivity can be influenced by previous annual crops. Agron. J. 2017, 109, 1423–1432. [Google Scholar] [CrossRef]
- Zimbric, J.W.; Stoltenberg, D.E.; Picasso, V.D. Effective weed suppression in dual-use intermediate wheatgrass systems. Agron. J. 2020, 112, 2164–2175. [Google Scholar] [CrossRef] [Green Version]
- Glover, J.D.; Culman, S.W.; DuPont, S.T.; Broussard, W.; Young, L.; Mangan, M.E.; Mia, J.G.; Crews, T.E.; DeHaan, L.R.; Buckley, D.H.; et al. Harvested perennial grasslands provide ecological benchmarks for agricultural sustainability. Agric. Ecosyst. Environ. 2010, 137, 3–12. [Google Scholar] [CrossRef]
- Culman, S.W.; Snapp, S.; Ollenburger, M.; Basso, B. Soil and water quality rapidly responds to the perennial grain Kernza wheatgrass. Agron J. 2013, 105, 735–744. [Google Scholar] [CrossRef] [Green Version]
- Jungers, J.M.; DeHaan, L.H.; Mulla, D.J.; Sheaffer, C.C.; Wyse, D.L. Reduced nitrate leaching in a perennial grain crop compared to maize in the Upper Midwest, USA. Agric. Ecosyst. Environ. 2019, 272, 63–73. [Google Scholar] [CrossRef]
- Culman, S.W.; DuPont, S.T.; Glover, J.D.; Buckley, D.H.; Fick, G.W.; Ferris, H.; Crews, T.E. Long-term impacts of high-input annual cropping and unfertilized perennial grass production on soil properties and belowground food webs in Kansas, USA. Agric. Ecosyst. Environ. 2010, 137, 13–24. [Google Scholar] [CrossRef]
- Crews, T.E.; Carton, W.; Olsson, L. Is the future of agriculture perennial? Imperatives and opportunities to reinvent agriculture by shifting from annual monocultures to perennial polycultures. Glob. Sustain. 2018, 1, e9. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira, G.; Brunsell, N.A.; Crews, T.E.; DeHaan, L.R.; Vico, G. Carbon and water relations in perennial Kernza (Thinopyrum intermedium): An overview. Plant Sci. 2020, 295, 110279. [Google Scholar] [CrossRef] [PubMed]
- Lanker, M.; Bell, M.; Picasso, V.D. Farmer perspectives and experiences introducing the novel perennial grain Kernza intermediate wheatgrass in the US Midwest. Renew. Agric. Food Syst. 2019, 35, 653–662. [Google Scholar] [CrossRef] [Green Version]
- Wayman, S.; Debray, V.; Parry, S.; David, C.; Ryan, M.R. Perspectives on perennial grain crop production among organic and conventional farmers in France and the United States. Agriculture 2019, 9, 244. [Google Scholar] [CrossRef] [Green Version]
- Marti, A.; Bock, J.E.; Pagani, M.A.; Ismail, B.; Seetharaman, K. Structural characterization of proteins in wheat flour doughs enriched with intermediate wheatgrass (Thinopyrum intermedium) flour. Food Chem. 2016, 194, 994–1002. [Google Scholar] [CrossRef] [PubMed]
- DeHaan, L.R.; Ismail, B.P. Perennial cereals provide ecosystem benefits. Cereal Foods World 2017, 62, 278–281. [Google Scholar] [CrossRef]
- Lazaroiu, G.; Andronie, M.; Uta, C.; Hurloiu, I. Trust management in organic agriculture: Sustainable consumption behavior, environmentally conscious purchase intention, and healthy food choices. Front. Public Health 2019, 7, 340. [Google Scholar] [CrossRef]
- Fernandez, C.W.; Ehlke, N.; Sheaffer, C.C.; Jungers, J.M. Effects of nitrogen fertilization and planting density on intermediate wheatgrass yield. Agron. J. 2020, 112, 4159–4170. [Google Scholar] [CrossRef]
- Sakiroglu, M.; Picasso, V.; Dong, C.; Hall, M.B.; Jungers, J.M. How does nitrogen and forage harvest affect belowground biomass and nonstructural carbohydrates in dual-use Kernza intermediate wheatgrass? Crop Sci. 2020, 60, 2562–2573. [Google Scholar] [CrossRef]
- Hunter, M.C.; Sheaffer, C.C.; Culman, S.W.; Jungers, J.M. Effects of defoliation and row spacing on intermediate wheatgrass I: Grain production. Agron. J. 2020, 112, 1748–1763. [Google Scholar] [CrossRef]
- Pinto, P.; De Haan, L.; Picasso, V. Post-harvest management practices impact on light penetration and kernza intermediate wheatgrass yield components. Agronomy 2021, 11, 442. [Google Scholar] [CrossRef]
- Law, E.P.; Pelzer, C.J.; Wayman, S.; DiTommaso, A.; Ryan, M.R. Strip-tillage renovation of intermediate wheatgrass (Thinopyrum intermedium) for maintaining grain yield in mature stands. Renew Agric. Food Syst. 2020, 36, 321–327. [Google Scholar] [CrossRef]
- Favre, J.R.; Munoz, T.; Combs, D.K.; Wattiaux, M.A.; Picasso, V.D. Forage nutritive value and predicted fiber digestibility of Kernza intermediate wheatgrass in monoculture and in mixture with red clover during the first production year. Anim. Feed. Sci. Technol. 2019, 258, 114298. [Google Scholar] [CrossRef]
- Mazzenga, A.; Gianesella, M.; Brscic, M.; Cozzi, G. Feeding behavior, diet digestibility, rumen fluid and metabolic parameters of beef cattle fed total mixed rations with a stepped substitution of wheat straw with maize silage. Livest. Sci. 2009, 122, 16–23. [Google Scholar] [CrossRef]
- Hunter, M.C.; Sheaffer, C.C.; Culman, S.W.; Lazarus, W.F.; Jungers, J.M. Effects of defoliation and row spacing on intermediate wheatgrass II: Forage yield and economics. Agron. J. 2020, 112, 1862–1880. [Google Scholar] [CrossRef]
- Pugliese, J.Y. Above- and Belowground Response to Managing Kernza (Thinopyrum intermedium) as a Dual-Use Crop for Forage and Grain. 2017. Available online: https://etd.ohiolink.edu/apexprod/rws_olink/r/1501/10?clear=10&p10_accession_num=osu1494000853982965 (accessed on 1 July 2021).
- Bell, L.W.; Byrne, F.; Ewing, M.A.; Wade, L.J. A preliminary whole-farm economic analysis of perennial wheat in an Australian dryland farming system. Agric. Syst. 2008, 96, 166–174. [Google Scholar] [CrossRef]
- Tautges, N.E.; Jungers, J.M.; Dehaan, L.R.; Wyse, D.L.; Sheaffer, C.C. Maintaining grain yields of the perennial cereal intermediate wheatgrass in monoculture v. bi-culture with alfalfa in the Upper Midwestern USA. J. Agric. Sci. 2018, 156, 758–773. [Google Scholar] [CrossRef]
- Zimbric, J.W.; Stoltenberg, D.E.; Picasso, V.D. Strategies to reduce plant height in dual-use intermediate wheatgrass cropping systems. Agron. J. 2020, 113, 1563–1573. [Google Scholar] [CrossRef]
- Weaver, J.E.; Stoddart, L.A. Response of the prairie to the great drought of 1934. Ecology 1935, 16, 612–629. [Google Scholar] [CrossRef]
- Leopold, A. A Sand County Almanac; Oxford University Press: Oxford, UK, 1968; ISBN 01950077785-62-311-7. [Google Scholar]
- Peni, D.; Stolarski, M.J.; Bordiean, A.; Krzyżaniak, M.; Devowski, M. Silphium perfoliatum—A herbaceous crop with increased interest in recent years for multi-purpose use. Agriculture 2020, 10, 640. [Google Scholar] [CrossRef]
- Pichard, G. Manejo, producción, y características nutricionales del silfo (Silphium perfoliatum) en climas templados del sur de Chile. Cien. Inv. Agric. 2012, 39, 61–77. [Google Scholar] [CrossRef] [Green Version]
- Vilela, A.E.; González-Paleo, L.; Ravetta, D.A.; Murrell, E.G.; van Tassel, D.L. Balancing forage production, seed yield, and pest management in the perennial sunflower Silphium integrifolium (Asteraceae). Agronomy 2020, 10, 1471. [Google Scholar] [CrossRef]
- Cumplido-Marin, L.; Graves, A.R.; Burgess, P.J.; Morhart, C.; Paris, P.; Jablonowsk, N.D.; Facciotto, G.; Bury, M.; Martens, R.; Nahm, M. Two novel energy crops: Sida hermaphrodita (L.) Rusby and Silphium perfoliatum L.-state of knowledge. Agronomy 2020, 10, 928. [Google Scholar] [CrossRef]
- Von Cossel, M.; Amarysti, C.; Wilhelm, H.; Priya, N.; Winkler, B.; Hoerner, L. The replacement of maize (Zea mays L.) by cup plant (Silphium perfoliatum L.) as biogas substrate and its implications for the energy and material flows of a large biogas plant. Biofuels Bioprod. Biorefin. 2020, 14, 152–179. [Google Scholar] [CrossRef] [Green Version]
- Wever, C.; Höller, M.; Becker, L.; Biertümpfel, A.; Köhler, J.; van Inghelandt, D.; Westhoff, P.; Pude, R.; Pestsova, E. Towards high-biomass yielding bioenergy crop Silphium perfoliatum L.: Phenotypic and genotypic evaluation of five cultivated populations. Biomass Bioenergy 2019, 124, 102–113. [Google Scholar] [CrossRef]
- Grunwald, D.; Panten, K.; Schwarz, A.; Bischoff, W.A.; Schittenhelm, S. Comparison of maize, permanent cup plant and a perennial grass mixture with regard to soil and water protection. GCB Bioenergy 2020, 12, 694–705. [Google Scholar] [CrossRef]
- Emmerling, C.; Ruf, T.; Audu, V.; Werner, W.; Udelhoven, T. Earthworm communities are supported by perennial bioenergy cropping systems. Eur. J. Soil Biol. 2021, 105, 103331. [Google Scholar] [CrossRef]
- Schorpp, Q.; Schrader, S. Earthworm functional groups respond to the perennial energy cropping system of the cup plant (Silphium perfoliatum L.). Biomass Bioenergy 2016, 87, 61–68. [Google Scholar] [CrossRef]
- Mueller, A.L.; Biertümpfel, A.; Friedritz, L.; Power, E.F.; Wright, G.A.; Dauber, J. Floral resources provided by the new energy crop, Silphium perfoliatum L. (Asteraceae). J. Apic. Res. 2019, 59, 232–245. [Google Scholar] [CrossRef]
- Teleuţă, A.; Coşman, S.; Ababii, V. Introduction of Silphium perfoliatum L. and its utilization possibilities. In Conservation of Plant Diversity; Gradina Botanica (Institut): Chișinău, Moldova, 2012; Volume 2, pp. 262–268. ISBN 978-997. [Google Scholar]
- Vilela, A.; González-Paleo, L.; Turner, K.; Peterson, K.; Ravetta, D.; Crews, T.E.; Van Tassel, D. Progress and bottlenecks in the early domestication of the perennial oilseed Silphium integrifolium, a sunflower substitute. Sustainability 2018, 10, 638. [Google Scholar] [CrossRef] [Green Version]
- Van Tassel, D.L.; Albrecht, K.A.; Bever, J.D.; Boe, A.A.; Brandvain, Y.; Crews, T.E.; Gansberger, M.; Gerstberger, P.; Gonzalez-Paleo, L.; Hulke, B.S.; et al. Accelerating Silphium domestication: An opportunity to develop new crop ideotypes and breeding strategies informed by multiple disciplines. Crop Sci. 2017, 57, 1274–1284. [Google Scholar] [CrossRef] [Green Version]
- Raduski, A.R.; Herman, A.; Pogoda, C.; Dorn, K.M.; Van Tassel, D.L.; Kane, N.; Brandvain, Y. Patterns of genetic variation in a prairie wildflower, Silphium integrifolium, suggest a non-prairie origin and locally adaptive variation. Am. J. Bot. 2021, 108, 145–158. [Google Scholar] [CrossRef]
- Reinert, S.; Price, J.H.; Smart, B.C.; Pogoda, C.S.; Kane, N.C.; Van Tassel, D.L.; Hulke, B.S. Mating compatibility and fertility studies in an herbaceous perennial Aster undergoing de novo domestication to enhance agroecosystems. Agron. Sustain. Dev. 2020, 40, 27. [Google Scholar] [CrossRef]
- Reinert, S.; Money, K.L.; Rockstad, G.B.G.; Kane, N.C.; Van Tassel, D.L.; Hulke, B.S. Two contrasting laboratory methods improve Silphium integrifolum Michx. Germination rate to agronomically acceptable levels. Euphytica 2018, 214, 156. [Google Scholar] [CrossRef]
- Von Gehren, P.; Gansberger, M.; Mayr, M.; Liebhard, P. The effect of sowing date and seed pretreatments on establishment of the energy plant Silphium perfolatium by sowing. Seed Sci. Technol. 2016, 44, 310–319. [Google Scholar] [CrossRef]
- Schiffner, S.; Jungers, J.M.; Van Tassel, D.; Smith, K.P.; Sheaffer, C.C. Seeding date affects seed and biomass yield of Silphium integrifolium Michx. (silflower). Nativ. Plants J. 2021, 22, 30–44. [Google Scholar] [CrossRef]
- Barbosa, C.H.; Andrade, M.A.; Vilarinho, F.; Castanheira, I.; Fernando, A.L.; Loizzo, M.R.; Silva, A.S. A new insight on cardoon: Exploring new uses besides cheese making with a view to zero waste. Foods 2020, 9, 564. [Google Scholar] [CrossRef]
- Savard, M.; Nesbitt, M.; Gale, R. Archaeobotanical evidence for early Neolithic diet and subsistence at M’lefaat (Iraq). Paléorient 2003, 29, 93–106. [Google Scholar] [CrossRef]
- Hind, N. 763. Gundelia tournefortii. Curtis’s Bot. Mag. 2013, 30, 114–138. [Google Scholar] [CrossRef]
- Yazdanshen, H.; Tavili, A.; Arzani, H.; Azarnivand, H. Traditional Gundelia tournefortii usage and its habitat destruction in Tiran va Karvan district in Iran’s Isfahan province. Ecologia 2016, 6, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Khojasted, F.; Chahouki, M.A.Z. Spatial patterns and coexistence of the native forb, Psathyrostachys fragilis (Asteracea) and the native invader, Gundelia tournefortii (Poaceae) in a semi-arid rangeland of Iran. Pol. J. Ecol. 2013, 61, 373–377. [Google Scholar]
- Karabulut, A.; Ozgur Ozcan, C.; Kamalak, A.; Canbolat, O. Comparison of the nutritive value of a turkey native forage, tumbleweed hay (Gundelia tournefortii L.), wheat straw and alfalfa hay using in situ and in vitro measurements with sheep. Arch. Latinoam. Prod. Anim. 2006, 14, 78–83. [Google Scholar]
- Dehghani-Samani, A.; Madreseh-Ghahfarokhi, S.; Dehghani-Samani, A. Evaluation of performance rate, some hematological and biochemical parameters in Iranian Afshari breed fattened sheep fed diet containing gundelia (Gundelia tournefortii L.). Iraqi J. Vet. Sci. 2019, 33, 33–38. [Google Scholar] [CrossRef] [Green Version]
- Van Tassel, D.L.; Tesdell, O.; Schlautman, B.; Rubin, M.J.; DeHaan, L.R.; Crews, T.E.; Streit Krug, A. New food crop domestication in the age of gene editing: Genetic, agronomic and cultural change remain co-evolutionarily entangled. Front. Plant Sci. 2020, 11, 789. [Google Scholar] [CrossRef]
- Khanzadeh, F.; Khodaparast, M.H.H.; Elhami Rad, A.H.; Rahmani, F.R. Physiochemical properties of Gundelia tournefortii L. seed oil. J. Agric. Sci. Technol. 2012, 14, 1535–1542. [Google Scholar]
- Sanderson, M.A.; Adler, P.R. Perennial forages as second generation bioenergy crops. Int. J. Mol. Sci. 2008, 9, 768–788. [Google Scholar] [CrossRef] [Green Version]
- Mortenson, J.S.; Waldron, B.L.; Larson, S.R.; Jensen, K.B.; DeHaan, L.R.; Peel, M.D.; Peel, M.D.; Creech, J.E. Quantitative trait loci (QTL) for forage traits in intermediate wheatgrass when grown as spaced-plants versus monoculture and polyculture swards. Agronomy 2019, 9, 580. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Grain Yield (kg ha−1) | Vegetative Biomass (kg ha−1) | |||||
|---|---|---|---|---|---|---|
| Location | First Year | Older Stands | First Year | Older Stands | Ref. | |
| Colorado | Fort Collins | 724 | 3 | 11,753 | 1808 | [188] |
| Kansas | Salina | 526 | 71 | 4098 | 4696 | [188] |
| Michigan | Hickory Corners | 112–157 | 1390–1662 | 3881–4984 | 12,202–17,131 | [170] |
| Minnesota | Crookston/Roseau | 452–1150 | 32–986 | 4037–8421 | 738–11,638 | [180,190] |
| Lamberton/Waseca | 33–1110 | 4684–13,161 | [190] | |||
| Morris | 107–809 | 4373–10,379 | [190] | |||
| Saint Paul | 535–876 | 183–664 | 9697–10,200 | 6604–8200 | [181,182,188] | |
| New York | Aurora/Ithaca | 1043 | 134–219 | 4723 | 4059–7290 | [184,188] |
| Wisconsin | Arlington | 446–902 | 105–479 | 6607 | 4666–5690 | [181,188,191] |
| Lancaster/Montfort | 143–203 | 59–326 | 6141 | 3183–19,495 | [183,185] | |
| Ohio | S. Charleston/Wooster | 651–758 | 36–655 | 3243–6069 | 2337–6040 | [188] |
| Dimension of Ecological Intensification * | Annual Forages | Perennial Forages | Dual-Use Crops/Forages |
|---|---|---|---|
| Increase in forage production | ++ | +++ | ++ |
| Increase in grain production | + | ++ | +++ |
| Increase in soil quality | ++ | +++ | +++ |
| Increase in soil carbon | ++ | +++ | +++ |
| Reduction of soil erosion | ++ | +++ | +++ |
| Reduction of nutrient leaching/ improvement in water quality | ++ | +++ | +++ |
| Suppression of weeds & other agronomic pests | +++ | ++ | ++ |
| Increase in managed biodiversity † | +–+++ | +–+++ | +–+++ |
| Increase in associated biodiversity | + | ++ | ++ |
| Economic benefits | ++ | ++ | +++ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Franco, J.G.; Berti, M.T.; Grabber, J.H.; Hendrickson, J.R.; Nieman, C.C.; Pinto, P.; Van Tassel, D.; Picasso, V.D. Ecological Intensification of Food Production by Integrating Forages. Agronomy 2021, 11, 2580. https://doi.org/10.3390/agronomy11122580
Franco JG, Berti MT, Grabber JH, Hendrickson JR, Nieman CC, Pinto P, Van Tassel D, Picasso VD. Ecological Intensification of Food Production by Integrating Forages. Agronomy. 2021; 11(12):2580. https://doi.org/10.3390/agronomy11122580
Chicago/Turabian StyleFranco, José G., Marisol T. Berti, John H. Grabber, John R. Hendrickson, Christine C. Nieman, Priscila Pinto, David Van Tassel, and Valentín D. Picasso. 2021. "Ecological Intensification of Food Production by Integrating Forages" Agronomy 11, no. 12: 2580. https://doi.org/10.3390/agronomy11122580