
Drought Stress in Grain Legumes: Effects, Tolerance Mechanisms and Management

 ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Effects of Drought Stress in Grain Legumes
2.1. Morphological Effects
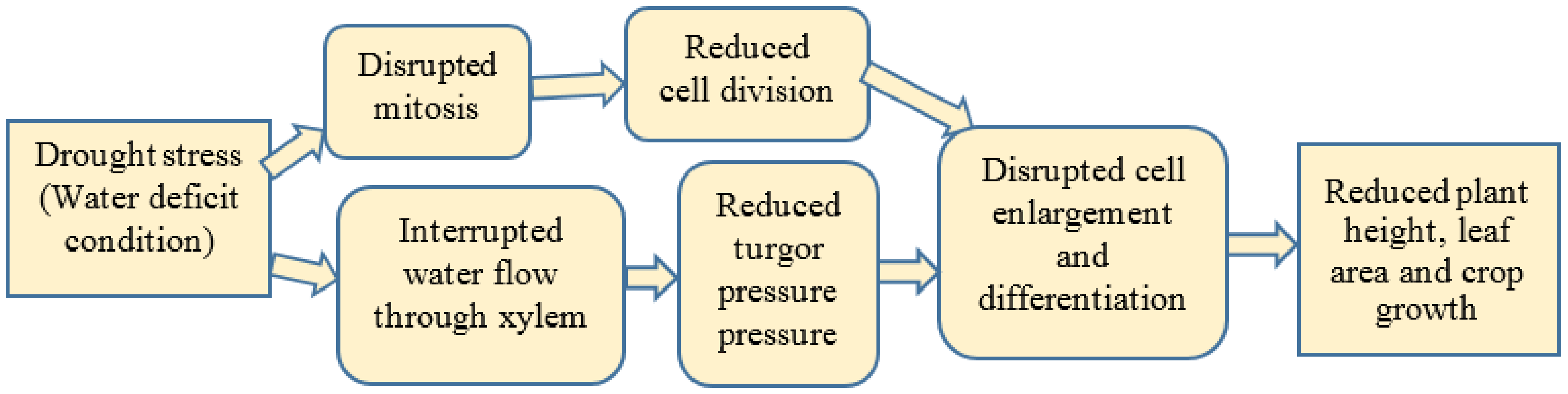
2.1.1. Plant Growth
2.1.2. Leaf Area
2.1.3. Pod Number
2.1.4. Nodulation of Grain Legumes
2.2. Physiological Effects
2.2.1. Leaf Temperature
2.2.2. Water-Use Efficiency
2.2.3. Chlorophyll Content
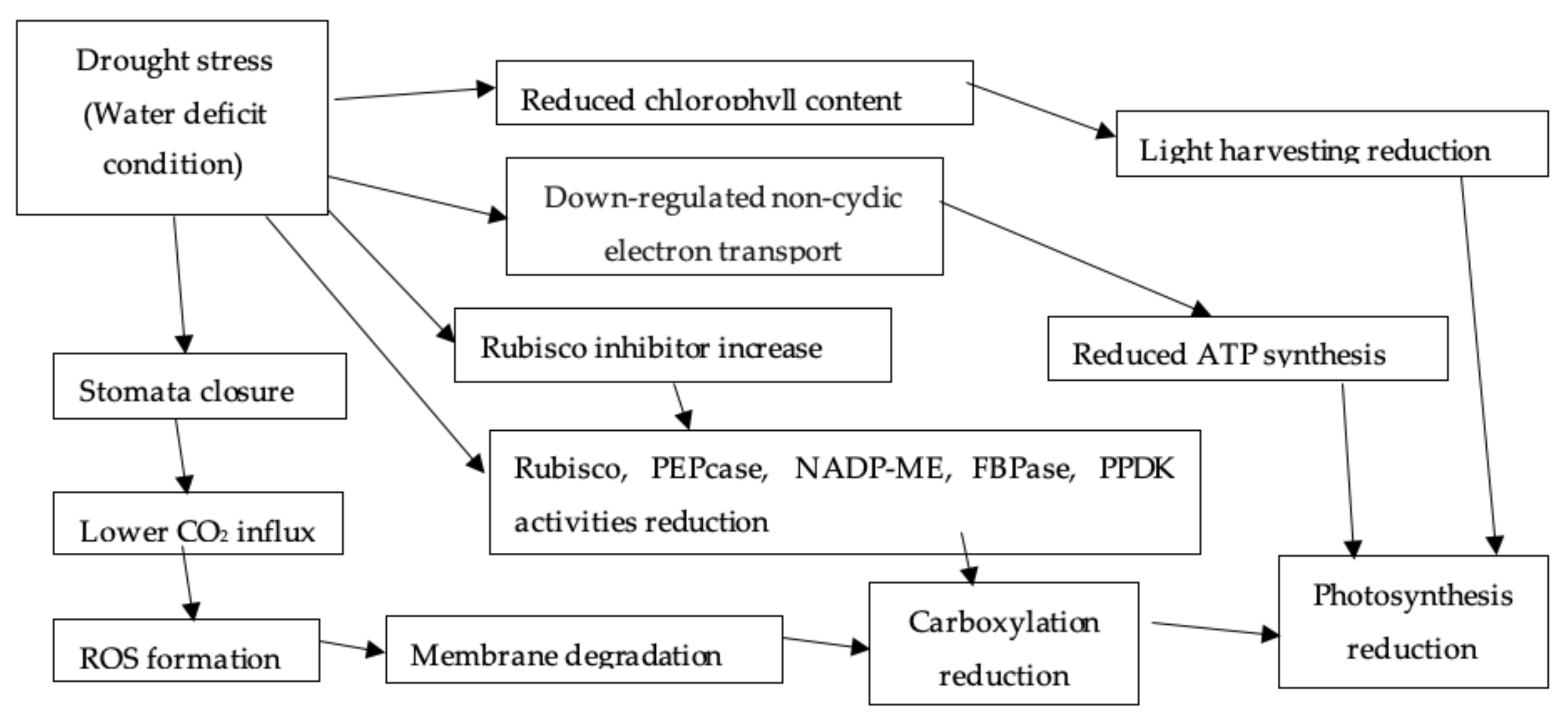
2.2.4. Photosynthesis
2.2.5. Transpiration and Stomatal Conductance
2.2.6. Plant–Water Relations
2.2.7. Plant Nutrient Relations
2.3. Morpho-Physiological Effects
2.3.1. Growth Stages
2.3.2. Grain Composition
2.3.3. Yield
2.3.4. Physio-Biochemical Level
2.3.5. Molecular Level
3. Tolerance Mechanisms of Grain Legumes against Drought Stress
3.1. Morphological Mechanisms
3.2. Phenotypic Plasticity
3.3. Leaf Abscission
4. Physio-Biochemical Mechanisms
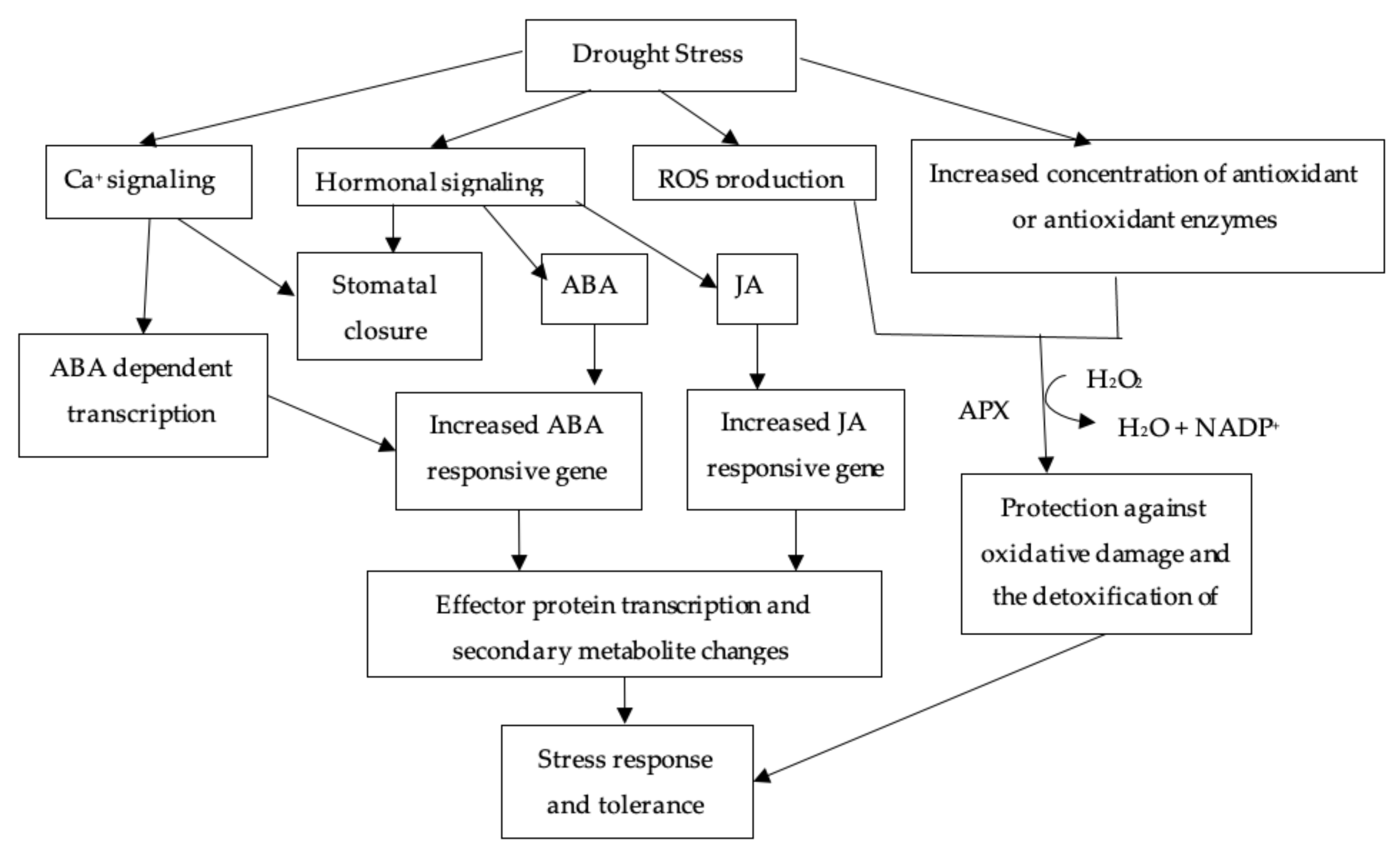
4.1. ABA Mediated Stomatal Closure
4.2. Antioxidant
4.3. Solute Accumulation
4.4. Plant Growth Regulators (PGRs)
4.5. Water-Use Efficiency (WUE) in Drought Tolerance
4.6. Molecular Mechanisms
5. Management of Drought Stress in Grain Legumes
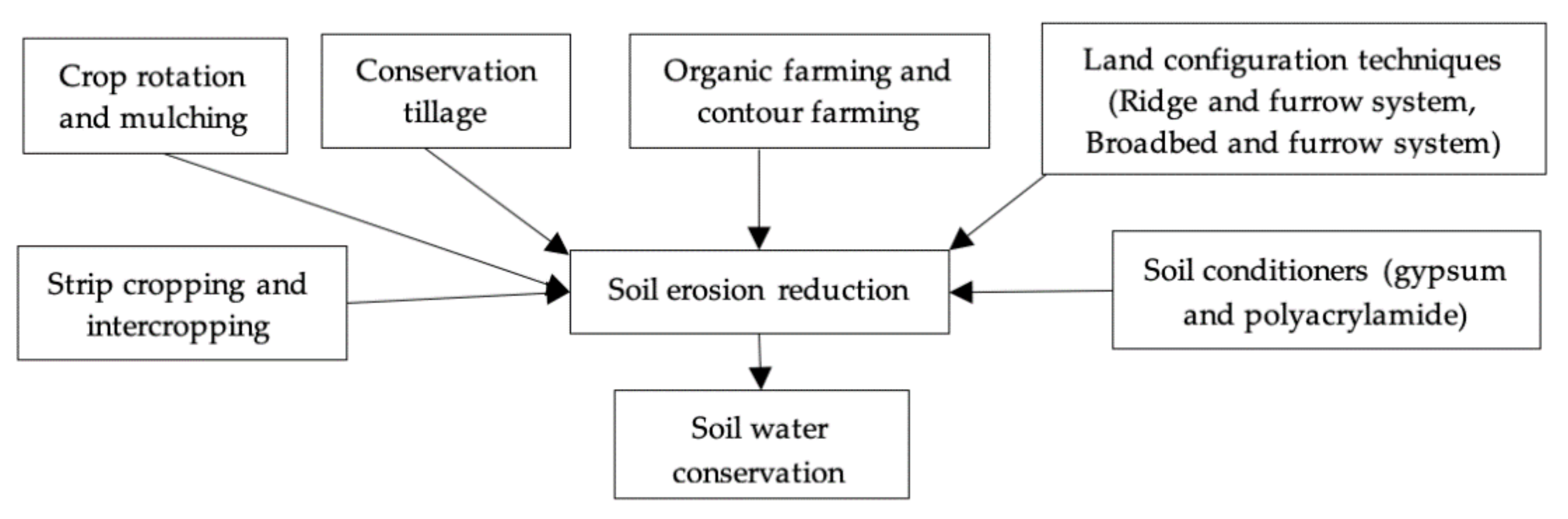
5.1. Traditional Agronomic Approaches
5.2. New Approaches in Agronomy
5.2.1. Biochar Application
5.2.2. Exogenous Application of Plant Growth Regulators (PGRs) and Osmoprotectants
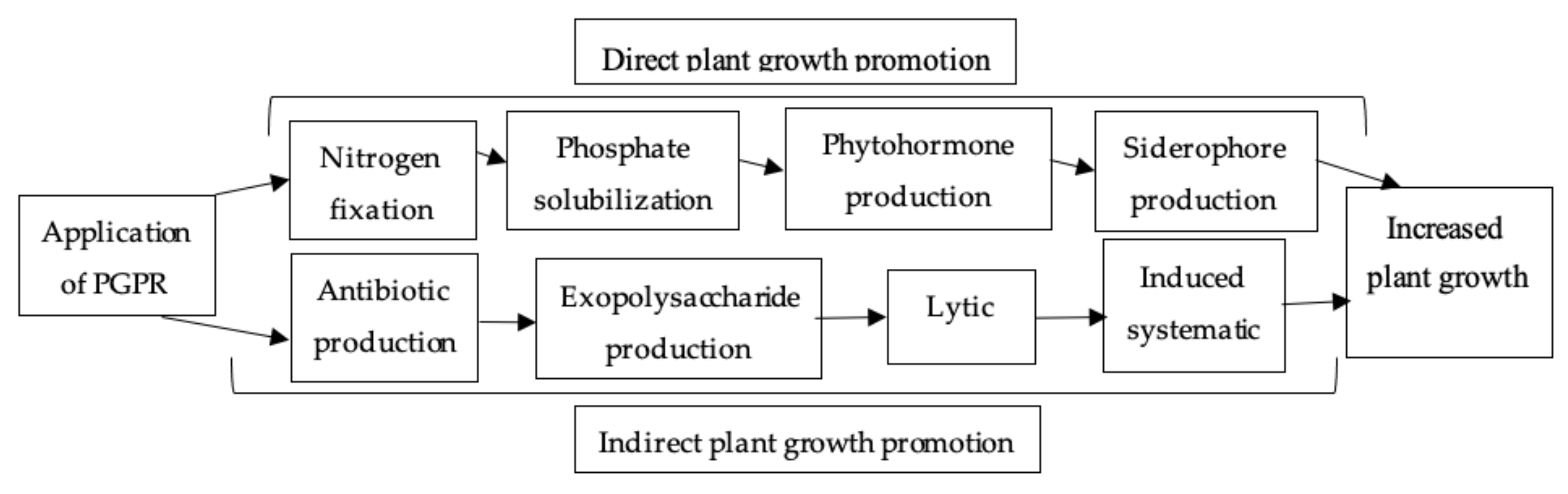
5.2.3. Plant-Growth-Promoting Rhizobacteria (PGPR)
5.2.4. Use of Arbuscular Mycorrhizal Fungi (AMF)
6. Breeding Approaches
6.1. Conventional Breeding
6.2. Genome-Wide Association Studies (GWASs)
6.3. Marker-Assisted Selection (MAS)
6.4. Genomic Selection (GS)
6.5. Biotechnological Approaches
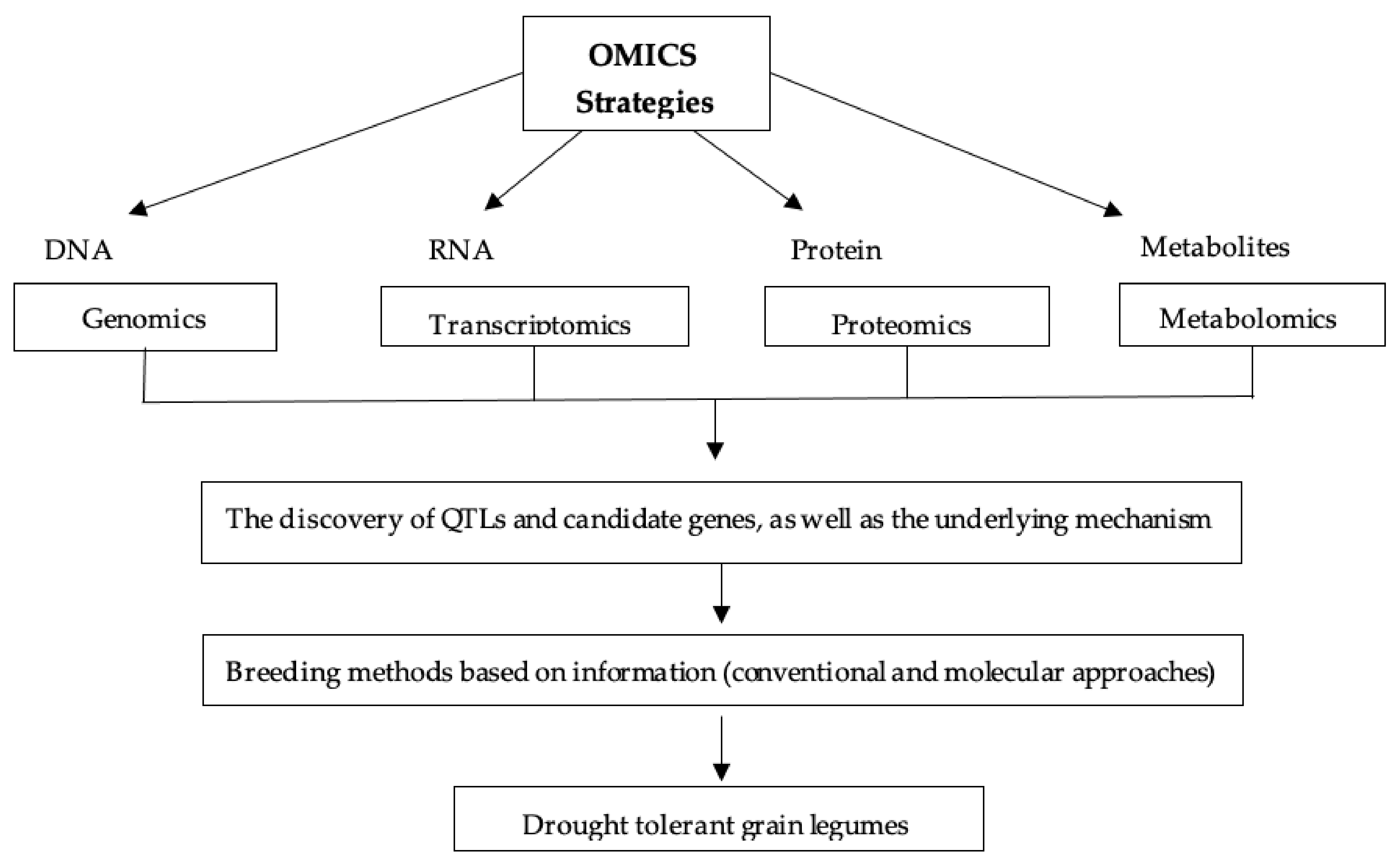
6.6. OMICS Strategy
6.7. CRISPR/Cas9: Sophisticated Technology for Genome Editing (GE)
7. Future Research Prospects
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Pradhan, J.; Katiyar, D.; Hemantaranjan, A. Drought mitigation strategies in pulses. Pharm. Innov. J. 2019, 8, 567–576. [Google Scholar]
- Abate, T.; Alene, A.D.; Bergvinson, D.; Shiferaw, B.; Silim, S.; Orr, A.; Asfaw, S. Tropical Grain Legumes in Africa and South Asia: Knowledge and Opportunities; International Crops Research Institute for the Semi-Arid Tropics: Patancheruvu, India, 2012. [Google Scholar]
- Daryanto, S.; Wang, L.; Jacinthe, P.A. Global synthesis of drought effects on food legume production. PLoS ONE 2015, 10, e0127401. [Google Scholar] [CrossRef] [Green Version]
- Araujo, S.S.; Beebe, S.; Crespi, M.; Delbreil, B.; Gonzalez, E.M.; Gruber, V.; Lejeune-Henaut, I.; Link, W.; Monteros, M.J.; Prats, E.; et al. Abiotic stress responses in legumes: Strategies used to cope with environmental challenges. Crit. Rev. Plant Sci. 2015, 34, 237–280. [Google Scholar] [CrossRef]
- Priyanka, J.P.; Goral, R.T.; Rupal, K.S.; Saraf, M. Rhizospheric microflora: A natural alleviator of drought stress in agricultural crops. In Plant Growth Promoting Rhizobacteria for Sustainable Stress Management; Springer: Singapore, 2019; pp. 103–115. [Google Scholar] [CrossRef]
- Kaushal, M.; Wani, S.P. Rhizobacterial-plant interactions: Strategies ensuring plant growth promotion under drought and salinity stress. Agric. Ecosyst. Environ. 2016, 231, 68–78. [Google Scholar] [CrossRef]
- Zhang, J.L.; Flowers, T.J.; Wang, S.M. Mechanisms of sodium uptake by roots of higher plants. Plant Soil 2010, 326, 45–60. [Google Scholar] [CrossRef]
- Ye, H.; Roorkiwal, M.; Valliyodan, B.; Zhou, L.; Chen, P.; Varshney, R.K.; Nguyen, H.T. Genetic diversity of root system architecture in response to drought stress in grain legumes. J. Exp. Bot. 2018, 69, 3267–3277. [Google Scholar] [CrossRef] [Green Version]
- Asma; Hussain, I.; Ashraf, M.Y.; Ashraf, M.A.; Rasheed, R.; Iqbal, M.; Anwar, S.; Shereen, A.; Khan, M.A. Assessment of rice (Oryza sativa L.) genotypes for drought stress tolerance using morpho-physiological indices as a screening technique. Pak. J. Bot. 2021, 53, 45–58. [Google Scholar] [CrossRef]
- Feng, S.; Sikdar, A.; Wang, J.; Memon, M.; Li, B.; Ma, H.; Lv, G. Response of Amorpha fruticosa seedlings to drought and rewatering in arid and semi-arid environment. Pak. J. Bot. 2021, 53, 419–424. [Google Scholar] [CrossRef]
- Lu, H.; Tabassum, A.; Zhou, G. Plant hydraulic conductivity determines photosynthesis in rice under PEG-induced drought stress. Pak. J. Bot. 2021, 53, 409–417. [Google Scholar]
- Foyer, C.H.; Lam, H.M.; Nguyen, H.T.; Siddique, K.H.; Varshney, R.K.; Colmer, T.D.; Cowling, W.; Bramley, H.; Mori, T.A.; Hodgson, J.M.; et al. Neglecting legumes has compromised human health and sustainable food production. Nat. Plants 2016, 2, 1–10. [Google Scholar] [CrossRef]
- Mourtzinis, S.; Specht, J.E.; Lindsey, L.E.; Wiebold, W.J.; Ross, J.; Nafziger, E.D.; Kandel, H.J.; Mueller, N.; Devillez, P.L.; Arriaga, F.J.; et al. Climate-induced reduction in US-wide soybean yields underpinned by region-and in-season-specific responses. Nat. Plants 2015, 1, 1–4. [Google Scholar] [CrossRef]
- Beebe, S.; Ramirez, J.; Jarvis, A.; Rao, I.M.; Mosquera, G.; Bueno, J.M.; Blair, M.W. Genetic improvement of common beans and the challenges of climate change. In Crop Adaptation to Climate Change; John Wiley & Sons: Hoboken, NJ, USA, 2011; pp. 356–369. [Google Scholar] [CrossRef]
- Tomer, A.; Singh, S.K. Drought Stress Tolerance in Legume Crops. In Agronomic Crops; Springer: Singapore, 2020; pp. 149–155. [Google Scholar] [CrossRef]
- Tiwari, S.; Lata, C.; Chauhan, P.S.; Nautiyal, C.S. Pseudomonas putida attunes morphophysiological, biochemical and molecular responses in Cicer arietinum L. during drought stress and recovery. Plant Physiol. Biochem. 2016, 99, 108–117. [Google Scholar] [CrossRef]
- Andrews, M.; Hodge, S. Climate change, a challenge for cool season grain legume crop production. In Climate Change and Management of Cool Season Grain Legume Crops; Springer: Dordrecht, The Netherlands, 2010; pp. 1–9. [Google Scholar] [CrossRef]
- Bhatia, V.S.; Singh, P.; Wani, S.P.; Rao, A.K.; Srinivas, K. Yield Gap Analysis of Soybean, Groundnut, Pigeonpea and Chickpea in India Using Simulation Modeling: Global Theme on Agroecosystems Report No. 31; International Crops Research Institute for the Semi-Arid Tropics: Patancheruvu, India, 2006. [Google Scholar]
- Anderson, W.; Johansen, C.; Siddique, K.H. Addressing the yield gap in rainfed crops: A review. Agron. Sustain. Dev. 2016, 36, 18. [Google Scholar] [CrossRef] [Green Version]
- Ngumbi, E.; Kloepper, J. Bacterial-mediated drought tolerance: Current and future prospects. Appl. Soil Ecol. 2016, 105, 109–125. [Google Scholar] [CrossRef]
- Mancosu, N.; Snyder, R.L.; Kyriakakis, G.; Spano, D. Water scarcity and future challenges for food production. Water 2015, 7, 975–992. [Google Scholar] [CrossRef]
- Mickelbart, M.V.; Hasegawa, P.M.; Bailey-Serres, J. Genetic mechanisms of abiotic stress tolerance that translate to crop yield stability. Nat. Rev. Genet. 2015, 16, 237–251. [Google Scholar] [CrossRef]
- Khoyerdi, F.F.; Shamshiri, M.H.; Estaji, A. Changes in some physiological and osmotic parameters of several pistachio genotypes under drought stress. Sci. Hortic. 2016, 198, 44–51. [Google Scholar] [CrossRef]
- Singh, S.; Gupta, A.K.; Kaur, N. Differential responses of antioxidative defence system to long-term field drought in wheat (Triticum aestivum L.) genotypes differing in drought tolerance. J. Agron. Crop Sci. 2012, 198, 185–195. [Google Scholar] [CrossRef]
- Deshmukh, R.; Sonah, H.; Patil, G.; Chen, W.; Prince, S.; Mutava, R.; Vuong, T.; Valliyodan, B.; Nguyen, H.T. Integrating omic approaches for abiotic stress tolerance in soybean. Front. Plant Sci. 2014, 5, 244. [Google Scholar] [CrossRef]
- Mir, R.R.; Zaman-Allah, M.; Sreenivasulu, N.; Trethowan, R.; Varshney, R.K. Integrated genomics, physiology and breeding approaches for improving drought tolerance in crops. Theor. Appl. Genet. 2012, 125, 625–645. [Google Scholar] [CrossRef] [Green Version]
- Fleury, D.; Jefferies, S.; Kuchel, H.; Langridge, P. Genetic and genomic tools to improve drought tolerance in wheat. J. Exp. Bot. 2010, 61, 3211–3222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ulemale, C.S.; Mate, S.N.; Deshmukh, D.V. Physiological indices for drought tolerance in chickpea (Cicer arietinum L.). World J. Agric. Sci. 2013, 9, 123–131. [Google Scholar]
- Miyauchi, Y.; Isoda, A.; Li, Z.; Wang, P. Soybean cultivation on desert sand using drip irrigation with mulch. Plant Prod. Sci. 2012, 15, 310–316. [Google Scholar] [CrossRef]
- Golldack, D.; Li, C.; Mohan, H.; Probst, N. Tolerance to drought and salt stress in plants: Unraveling the signaling networks. Front. Plant Sci. 2014, 5, 151. [Google Scholar] [CrossRef] [Green Version]
- Shavrukov, Y.; Kurishbayev, A.; Jatayev, S.; Shvidchenko, V.; Zotova, L.; Koekemoer, F.; Groot, S.D.; Kathleen Soole, K.; Langridge, P. Early flowering as a drought escape mechanism in plants: How can it aid wheat production? Front. Plant Sci. 2017, 8, 1950. [Google Scholar] [CrossRef]
- Kamanga, R.M.; Mbega, E.; Ndakidemi, P. Drought tolerance mechanisms in plants: Physiological responses associated with water deficit stress in Solanum lycopersicum. Adv. Crop Sci. Technol. 2018, 6, 1–8. [Google Scholar] [CrossRef]
- Bechtold, U. Plant life in extreme environments: How do you improve drought tolerance? Front. Plant Sci. 2018, 9, 543. [Google Scholar] [CrossRef] [Green Version]
- Taiz, L.; Zeiger, E. Plant Physiology, 4th ed.; Sinauer Associates Inc. Publishers: Sunderland, MA, USA, 2006. [Google Scholar]
- Farooq, M.; Aziz, T.; Basra, S.M.A.; Cheema, M.A.; Rehman, H. Chilling tolerance in hybrid maize induced by seed priming with salicylic acid. J. Agron. Crop Sci. 2008, 194, 161–168. [Google Scholar] [CrossRef]
- Vessal, S.; Arefian, M.; Siddique, K.H. Proteomic responses to progressive dehydration stress in leaves of chickpea seedlings. BMC Genom. 2020, 21, 523. [Google Scholar] [CrossRef]
- Garg, B.K. Nutrient uptake and management under drought: Nutrient-moisture interaction. Curr. Agric. 2003, 27, 1–8. [Google Scholar]
- Farooq, M.; Wahid, A.; Kobayashi, N.S.M.A.; Fujita, D.B.S.M.A.; Basra, S.M.A. Plant drought stress: Effects, mechanisms and management. Sustain. Agric. 2009, 153–188. [Google Scholar] [CrossRef] [Green Version]
- Fathi, A.; Tari, D.B. Effect of drought stress and its mechanism in plants. Int. J. Life Sci. 2016, 10, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Hudak, C.M.; Patterson, R.P. Root distribution and soil moisture depletion pattern of a drought-resistant soybean plant introduction. Agron. J. 1996, 88, 478–485. [Google Scholar] [CrossRef]
- Bagheri, A. Effects of drought stress on chlorophyll, proline and rates of photosynthesis and respiration and activity of superoxide dismutase and peroxidase in millet (Panicum milenaceum L.). In Proceedings of the National Conference on Water Scarcity and Drought Management in Agriculture, Arsanjan; 2009; p. 16. [Google Scholar]
- Kabiri, R.; Farahbakhsh, H.; Nasibi, F. Salicylic acid ameliorates the effects of oxidative stress induced by water deficit in hydroponic culture of Nigella sativa. J. Stress Physiol. Biochem. 2012, 8, 13–22. [Google Scholar]
- Sarkar, S.; Khatun, M.; Era, F.M.; Islam, A.K.M.M.; Anwar, M.P.; Danish, S.; Datta, R.; Islam, A.K.M.A. Abiotic stresses: Alteration of composition and grain quality in food legumes. Agronomy 2021, 11, 2238. [Google Scholar] [CrossRef]
- De Souza, P.I.; Egli, D.B.; Bruening, W.P. Water stress during seed filling and leaf senescence in soybean. Agron. J. 1997, 89, 807–812. [Google Scholar] [CrossRef]
- Mafakheri, A.; Siosemardeh, A.F.; Bahramnejad, B.; Struik, P.C.; Sohrabi, Y. Effect of drought stress on yield, proline and chlorophyll contents in three chickpea cultivars. Aust. J. Crop Sci. 2010, 4, 580–585. [Google Scholar]
- Busse, M.D.; Bottomley, P.J. Growth and nodulation responses of Rhizobium meliloti to water stress induced by permeating and nonpermeating solutes. Appl. Environ. Microbiol. 1989, 55, 2431–2436. [Google Scholar] [CrossRef] [Green Version]
- Chowdhury, J.A.; Karim, M.A.; Khaliq, Q.A.; Ahmed, A.U.; Mondol, A.M. Effect of drought stress on water relation traits of four soybean genotypes. SAARC J. Agric. 2017, 15, 163–175. [Google Scholar] [CrossRef] [Green Version]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; et al. Crop production under drought and heat stress: Plant responses and management options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef] [Green Version]
- Monclus, R.; Dreyer, E.; Villar, M.; Delmotte, F.M.; Delay, D.; Petit, J.M.; Barbaroux, C.; Thiec, D.L.; Bréchet, C.; Brignolas, F. Impact of drought on productivity and water use efficiency in 29 genotypes of Populus deltoides× Populus nigra. New Phytol. 2006, 169, 765–777. [Google Scholar] [CrossRef] [PubMed]
- Sohrawardy, H.; Hossain, M. Response of short duration tropical legumes and maize to water stress: A glasshouse study. Adv. Agric. 2014, 2014, 641319. [Google Scholar] [CrossRef] [Green Version]
- Anjum, S.A.; Xie, X.Y.; Wang, L.C.; Saleem, M.F.; Man, C.; Lei, W. Morphological, physiological and biochemical responses of plants to drought stress. Afr. J. Agric. Res. 2011, 6, 2026–2032. [Google Scholar]
- Ashraf, M.; Karim, F. Screening of some cultivars/lines of black gram (Vigna mungo L. Hepper) for resistance to water stress. Trop. Agric. 1991, 68, 57–62. [Google Scholar]
- Jain, M.; Tiwary, S.; Gadre, R. Sorbitol-induced changes in various growth and biochemici parameters in maize. Plant Soil Environ. 2010, 56, 263–267. [Google Scholar] [CrossRef] [Green Version]
- Awasthi, R.; Kaushal, N.; Vadez, V.; Turner, N.C.; Berger, J.; Siddique, K.H.; Nayyar, H. Individual and combined effects of transient drought and heat stress on carbon assimilation and seed filling in chickpea. Funct. Plant Biol. 2014, 41, 1148–1167. [Google Scholar] [CrossRef] [Green Version]
- Tang, A.C.; Kawamitsu, Y.; Kanechi, M.; Boyer, J.S. Photosynthetic oxygen evolution at low water potential in leaf discs lacking an epidermis. Ann. Bot. 2002, 89, 861–870. [Google Scholar] [CrossRef] [Green Version]
- Makbul, S.; Guler, N.S.; Durmus, N.; Guven, S. Changes in anatomical and physiological parameters of soybean under drought stress. Turk. J. Bot. 2011, 35, 369–377. [Google Scholar]
- Ohashi, Y.; Saneoka, H.; Fujita, K. Effect of water stress on growth, photosynthesis, and photoassimilate translocation in soybean and tropical pasture legume siratro. Soil Sci. Plant Nutr. 2000, 46, 417–425. [Google Scholar]
- Omae, H.; Kumar, A.; Kashiwaba, K.; Shono, M. Assessing drought tolerance of snap bean (Phaseolus vulgaris) from genotypic differences in leaf water relations, shoot growth and photosynthetic parameters. Plant Prod. Sci. 2007, 10, 28–35. [Google Scholar] [CrossRef]
- Omae, H.; Kumar, A.; Egawa, Y.; Kashiwaba, K.; Shono, M. Midday drop of leaf water content related to drought tolerance in snap bean (Phaseolus vulgaris L.). Plant Prod. Sci. 2005, 8, 465–467. [Google Scholar] [CrossRef]
- Barber, S.A. Soil Nutrient Bioavailability: A Mechanistic Approach; John Wiley and Sons: Hoboken, NJ, USA, 1995. [Google Scholar]
- Grossman, A.; Takahashi, H. Macronutrient utilization by photosynthetic eukaryotes and the fabric of interactions. Annu. Rev. Plant Biol. 2001, 52, 163–210. [Google Scholar] [CrossRef] [Green Version]
- Ullah, A.; Farooq, M. The challenge of drought stress for grain legumes and options for improvement. Arch. Agron. Soil Sci. 2021, 1–18. [Google Scholar] [CrossRef]
- Kaya, M.D.; Okçu, G.; Atak, M.; Cıkılı, Y.; Kolsarıcı, Ö. Seed treatments to overcome salt and drought stress during germination in sunflower (Helianthus annuus L.). Eur. J. Agron. 2006, 24, 291–295. [Google Scholar] [CrossRef]
- Harris, D.; Tripathi, R.S.; Joshi, A. On-farm seed priming to improve crop establishment and yield in dry direct-seeded rice. In Direct Seeding: Research Strategies and Opportunities; International Research Institute: Manila, Philippines, 2002; pp. 231–240. [Google Scholar]
- Okçu, G.; Kaya, M.D.; Atak, M. Effects of salt and drought stresses on germination and seedling growth of pea (Pisum sativum L.). Turk. J. Agric. For. 2005, 29, 237–242. [Google Scholar]
- Farooq, M.; Hussain, M.; Siddique, K.H. Drought stress in wheat during flowering and grain-filling periods. Crit. Rev. Plant Sci. 2014, 33, 331–349. [Google Scholar] [CrossRef]
- Fang, X.; Turner, N.C.; Yan, G.; Li, F.; Siddique, K.H. Flower numbers, pod production, pollen viability, and pistil function are reduced and flower and pod abortion increased in chickpea (Cicer arietinum L.) under terminal drought. J. Exp. Bot. 2010, 61, 335–345. [Google Scholar] [CrossRef] [Green Version]
- Al-Ghzawi, A.A.; Zaitoun, S.; Gosheh, H.Z.; Alqudah, A.M. The impacts of drought stress on bee attractively and flower pollination of Trigonella moabitica (fabaceae). Arch. Agron. Soil Sci. 2009, 55, 683–692. [Google Scholar] [CrossRef]
- Shrestha, R.; Turner, N.C.; Siddique, K.H.M.; Turner, D.W.; Speijers, J. A water deficit during pod development in lentils reduces flower and pod numbers but not seed size. Aust. J. Agric. Res. 2006, 57, 427–438. [Google Scholar] [CrossRef]
- Liu, F.; Jensen, C.R.; Andersen, M.N. Pod set related to photosynthetic rate and endogenous ABA in soybeans subjected to different water regimes and exogenous ABA and BA at early reproductive stages. Ann. Bot. 2004, 94, 405–411. [Google Scholar] [CrossRef] [Green Version]
- Davies, S.L.; Turner, N.C.; Siddique, K.H.M.; Leport, L.; Plummer, J.A. Seed growth of desi and kabuli chickpea (Cicer arietinum L.) in a short-season Mediterranean-type environment. Aust. J. Exp. Agric. 1999, 39, 181–188. [Google Scholar] [CrossRef]
- Singh, S.P. Drought resistance in the race Durango dry bean landraces and cultivars. Agron. J. 2007, 99, 1219–1225. [Google Scholar] [CrossRef]
- Ghanbari, A.A.; Shakiba, M.R.; Toorchi, M.; Choukan, R. Nitrogen changes in the leaves and accumulation of some minerals in the seeds of red, white and chitti beans (Phaseolus vulgaris) under water deficit conditions. Aust. J. Crop Sci. 2013, 7, 706–712. [Google Scholar]
- Ghanbari, A.A.; Mousavi, S.H.; Gorji, A.M.; Idupulapati, R.A.O. Effects of water stress on leaves and seeds of bean (Phaseolus vulgaris L.). Turk. J. Field Crop. 2013, 18, 73–77. [Google Scholar]
- Bellaloui, N.; Mengistu, A.; Kassem, M.A. Effects of genetics and environment on fatty acid stability in soybean seed. Food Nutr. Sci. 2013, 4, 165–175. [Google Scholar] [CrossRef] [Green Version]
- Dornbos, D.L.; Mullen, R.E. Soybean seed protein and oil contents and fatty acid composition adjustments by drought and temperature. J. Am. Oil Chem. Soc. 1992, 69, 228–231. [Google Scholar] [CrossRef]
- Labanauskas, C.K.; Shouse, P.; Stolzy, L.H.; Handy, M.F. Protein and free amino acids in field-grown cowpea seeds as affected by water stress at various growth stages. Plant Soil 1981, 63, 355–368. [Google Scholar] [CrossRef]
- Estrada-Campuzano, G.; Miralles, D.J.; Slafer, G.A. Genotypic variability and response to water stress of pre-and post-anthesis phases in triticale. Eur. J. Agron. 2008, 28, 171–177. [Google Scholar] [CrossRef]
- Samarah, N.H. Effects of drought stress on growth and yield of barley. Agron. Sustain. Dev. 2005, 25, 145–149. [Google Scholar] [CrossRef] [Green Version]
- Jukanti, A.K.; Bhatt, R.; Sharma, R.; Kalia, R.K. Morphological, agronomic, and yield characterization of cluster bean (Cyamopsis tetragonoloba L.) germplasm accessions. J. Crop Sci. Biotechnol. 2015, 18, 83–88. [Google Scholar] [CrossRef]
- Nayyar, H.; Kaur, S.; Singh, S.; Upadhyaya, H.D. Differential sensitivity of Desi (small-seeded) and Kabuli (large-seeded) chickpea genotypes to water stress during seed filling: Effects on accumulation of seed reserves and yield. J. Sci. Food Agric. 2006, 86, 2076–2082. [Google Scholar] [CrossRef]
- Samarah, N.H.; Haddad, N.; Alqudah, A.M. Yield potential evaluation in chickpea genotypes under late terminal drought in relation to the length of reproductive stage. Ital. J. Agron. 2009, 111–117. [Google Scholar] [CrossRef]
- Nam, N.H.; Chauhan, Y.S.; Johansen, C. Effect of timing of drought stress on growth and grain yield of extra-short-duration pigeonpea lines. J. Agric. Sci. 2001, 136, 179–189. [Google Scholar] [CrossRef] [Green Version]
- Kobraee, S.; Shamsi, K.; Rasekhi, B. Soybean production under water deficit conditions. Ann. Biol. Res. 2011, 2, 423–434. [Google Scholar]
- Maleki, A.; Naderi, A.; Naseri, R.; Fathi, A.; Bahamin, S.; Maleki, R. Physiological performance of soybean cultivars under drought stress. Bull. Environ. Pharmacol. Life Sci. 2013, 2, 38–44. [Google Scholar]
- Wei, Y.; Jin, J.; Jiang, S.; Ning, S.; Liu, L. Quantitative response of soybean development and yield to drought stress during different growth stages in the Huaibei Plain, China. Agronomy 2018, 8, 97. [Google Scholar] [CrossRef] [Green Version]
- Martínez, J.P.; Silva, H.F.L.J.; Ledent, J.F.; Pinto, M. Effect of drought stress on the osmotic adjustment, cell wall elasticity and cell volume of six cultivars of common beans (Phaseolus vulgaris L.). Eur. J. Agron. 2007, 26, 30–38. [Google Scholar] [CrossRef]
- Rosales-Serna, R.; Kohashi-Shibata, J.; Acosta-Gallegos, J.A.; Trejo-López, C.; Ortiz-Cereceres, J.; Kelly, J.D. Biomass distribution, maturity acceleration and yield in drought-stressed common bean cultivars. Field Crop. Res. 2004, 85, 203–211. [Google Scholar] [CrossRef]
- Ahmed, F.E.; Suliman, A.S.H. Effect of water stress applied at different stages of growth on seed yield and water-use efficiency of cowpea. Agric. Biol. J. North Am. 2010, 1, 534–540. [Google Scholar]
- Ogbonnaya, C.I.; Sarr, B.; Brou, C.; Diouf, O.; Diop, N.N.; Roy-Macauley, H. Selection of cowpea genotypes in hydroponics, pots, and field for drought tolerance. Crop Sci. 2003, 43, 1114–1120. [Google Scholar] [CrossRef]
- Ghassemi-Golezani, K.; Hosseinzadeh-Mahootchy, A. Changes in seed vigour of faba bean (Vicia faba L.) cultivars during development and maturity. Seed Sci. Technol. 2009, 37, 713–720. [Google Scholar] [CrossRef]
- Baroowa, B.; Gogoi, N. Biochemical changes in black gram and green gram genotypes after imposition of drought stress. J. Food Legumes 2014, 27, 350–353. [Google Scholar]
- Baroowa, B.; Gogoi, N. Biochemical changes in two Vigna spp. during drought and subsequent recovery. Indian J. Plant Physiol. 2013, 18, 319–325. [Google Scholar] [CrossRef]
- Allahmoradi, P.; Mansourifar, C.; Saiedi, M.; Jalali Honarmand, S. Effect of different water deficiency levels on some antioxidants at different growth stages of lentil (Lens culinaris L.). Adv. Environ. Biol. 2013, 7, 535–543. [Google Scholar]
- Frederick, J.R.; Camp, C.R.; Bauer, P.J. Drought-stress effects on branch and mainstem seed yield and yield components of determinate soybean. Crop Sci. 2001, 41, 759–763. [Google Scholar] [CrossRef]
- Thankamani, C.K.; Chempakam, B.; Ashokan, P.K. Water stress induced changes in enzyme activities and lipid peroxidation in black pepper (Piper nigrum). J. Med. Aromat. Plant Sci. 2003, 25, 646–650. [Google Scholar]
- Møller, I.M.; Jensen, P.E.; Hansson, A. Oxidative modifications to cellular components in plants. Annu. Rev. Plant Biol. 2007, 58, 459–481. [Google Scholar] [CrossRef] [Green Version]
- Moran, J.F.; Becana, M.; Iturbe-Ormaetxe, I.; Frechilla, S.; Klucas, R.V.; Aparicio-Tejo, P. Drought induces oxidative stress in pea plants. Planta 1994, 194, 346–352. [Google Scholar] [CrossRef] [Green Version]
- Ingram, J.; Bartels, D. The molecular basis of dehydration tolerance in plants. Annu. Rev. Plant Biol. 1996, 47, 377–403. [Google Scholar] [CrossRef] [Green Version]
- Gao, W.R.; Wang, X.S.; Liu, Q.Y.; Peng, H.; Chen, C.; Li, J.G.; Zhang, J.S.; Song-Nian Hub, S.N.; Ma, H. Comparative analysis of ESTs in response to drought stress in chickpea (Cicer arietinum L.). Biochem. Biophys. Res. Commun. 2008, 376, 578–583. [Google Scholar] [CrossRef]
- Varshney, R.K.; Tuberosa, R.; Tardieu, F. Progress in understanding drought tolerance: From alleles to cropping systems. J. Exp. Bot. 2018, 69, 3175–3179. [Google Scholar] [CrossRef]
- Kavar, T.; Maras, M.; Kidrič, M.; Šuštar-Vozlič, J.; Meglič, V. Identification of genes involved in the response of leaves of Phaseolus vulgaris to drought stress. Mol. Breed. 2008, 21, 159–172. [Google Scholar] [CrossRef]
- Kramer, P.J. Plant and soil water relationships: A modern synthesis. In Plant and Soil Water Relationships: A Modern Synthesis; McGraw-Hill Book Company: New York, NY, USA, 1969. [Google Scholar]
- Prince, S.J.; Murphy, M.; Mutava, R.N.; Durnell, L.A.; Valliyodan, B.; Shannon, J.G.; Nguyen, H.T. Root xylem plasticity to improve water use and yield in water-stressed soybean. J. Exp. Bot. 2017, 68, 2027–2036. [Google Scholar] [CrossRef] [Green Version]
- Jaganathan, D.; Thudi, M.; Kale, S.; Azam, S.; Roorkiwal, M.; Gaur, P.M.; Kishor, P.B.; Nguyen, H.; Sutton, T.; Varshney, R.K. Genotyping-by-sequencing based intra-specific genetic map refines a ‘‘QTL-hotspot” region for drought tolerance in chickpea. Mol. Genet. Genom. 2015, 290, 559–571. [Google Scholar] [CrossRef]
- Salazar, C.; Hernández, C.; Pino, M.T. Plant water stress: Associations between ethylene and abscisic acid response. Chil. J. Agric. Res. 2015, 75, 71–79. [Google Scholar] [CrossRef] [Green Version]
- Shekari, F. Effect of Drought Stress on Phenology, Water Relations, Growth, Yield and Quality Canola. Ph.D. Thesis, University of Tabriz, Tabriz, Iran, 2000. [Google Scholar]
- Kafi, M.; Damghany Mahdavi, A. Mechanism of Resistance of Plants to Environmental Stresses; University of Mashhad: Mashhad, Iran, 1999. [Google Scholar]
- Schachtman, D.P.; Goodger, J.Q. Chemical root to shoot signaling under drought. Trends Plant Sci. 2008, 13, 281–287. [Google Scholar] [CrossRef]
- Matysik, J.; Alia; Bhalu, B.; Mohanty, P. Molecular mechanisms of quenching of reactive oxygen species by proline under stress in plants. Curr. Sci. 2002, 82, 525–532. [Google Scholar]
- Nayyar, H.; Walia, D.P. Water stress induced proline accumulation in contrasting wheat genotypes as affected by calcium and abscisic acid. Biol. Plant. 2003, 46, 275–279. [Google Scholar] [CrossRef]
- Anjum, S.A.; Wang, L.; Farooq, M.; Khan, I.; Xue, L. Methyl jasmonate-induced alteration in lipid peroxidation, antioxidative defence system and yield in soybean under drought. J. Agron. Crop Sci. 2011, 197, 296–301. [Google Scholar] [CrossRef]
- Szabados, L.; Savouré, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef]
- Nadeem, M.; Li, J.; Yahya, M.; Sher, A.; Ma, C.; Wang, X.; Qiu, L. Research progress and perspective on drought stress in legumes: A review. Int. J. Mol. Sci. 2019, 20, 2541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farooq, M.; Irfan, M.; Aziz, T.; Ahmad, I.; Cheema, S.A. Seed priming with ascorbic acid improves drought resistance of wheat. J. Agron. Crop Sci. 2013, 199, 12–22. [Google Scholar] [CrossRef]
- Desoky, E.S.M.; Mansour, E.; El-Sobky, E.S.E.; Abdul-Hamid, M.I.; Taha, T.F.; Elakkad, H.A.; Arnaout, S.M.; Eid, R.S.; El-Tarabily, K.A.; Yasin, M.A. Physio-biochemical and agronomic responses of faba beans to exogenously applied nano-silicon under drought stress conditions. Front. Plant Sci. 2021, 12, 637783. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.H.; Murata, N. Enhancement of tolerance of abiotic stress by metabolic engineering of betaines and other compatible solutes. Curr. Opin. Plant Biol. 2002, 5, 250–257. [Google Scholar] [CrossRef]
- Verbruggen, N.; Hermans, C. Proline accumulation in plants: A review. Amino Acids 2008, 35, 753–759. [Google Scholar] [CrossRef] [PubMed]
- Amede, T.; Schubert, S.; Stahr, K. Mechanisms of drought resistance in grain legumes I: Osmotic adjustment. SINET Ethiopian J. Sci. 2003, 26, 37–46. [Google Scholar] [CrossRef]
- Fahad, S.; Hussain, S.; Matloob, A.; Khan, F.A.; Khaliq, A.; Saud, S.; Hassan, S.; Shan, D.; Khan, F.; Ullah, N.; et al. Phytohormones and plant responses to salinity stress: A review. Plant Growth Regul. 2015, 75, 391–404. [Google Scholar] [CrossRef]
- Abobatta, W.F. Drought adaptive mechanisms of plants—A review. Adv. Agric. Environ. Sci. 2019, 2, 62–65. [Google Scholar] [CrossRef]
- Saradadevi, R.; Palta, J.A.; Siddique, K.H. ABA-mediated stomatal response in regulating water use during the development of terminal drought in wheat. Front. Plant Sci. 2017, 8, 1251. [Google Scholar] [CrossRef]
- Rani, A.; Devi, P.; Jha, U.C.; Sharma, K.D.; Siddique, K.H.; Nayyar, H. Developing climate-resilient chickpea involving physiological and molecular approaches with a focus on temperature and drought stresses. Front. Plant Sci. 2020, 10, 1759. [Google Scholar] [CrossRef]
- Ali, F.; Bano, A.; Fazal, A. Recent methods of drought stress tolerance in plants. Plant Growth Regul. 2017, 82, 363–375. [Google Scholar] [CrossRef]
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M.R. Heat tolerance in plants: An overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Chinnusamy, V.; Schumaker, K.; Zhu, J.K. Molecular genetic perspectives on cross-talk and specificity in abiotic stress signalling in plants. J. Exp. Bot. 2004, 55, 225–236. [Google Scholar] [CrossRef]
- Bray, E.A. Classification of genes differentially expressed during water-deficit stress in Arabidopsis thaliana: An analysis using microarray and differential expression data. Ann. Bot. 2002, 89, 803–811. [Google Scholar] [CrossRef] [Green Version]
- Xiong, L.; Schumaker, K.S.; Zhu, J.K. Cell signaling during cold, drought, and salt stress. Plant Cell 2002, 14 Suppl. S1, S165–S183. [Google Scholar] [CrossRef] [Green Version]
- Wrzaczek, M.; Hirt, H. Plant MAP kinase pathways: How many and what for? Biol. Cell 2001, 93, 81–87. [Google Scholar] [CrossRef]
- Abe, H.; Urao, T.; Ito, T.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Arabidopsis AtMYC2 (bHLH) and AtMYB2 (MYB) function as transcriptional activators in abscisic acid signaling. Plant Cell 2003, 15, 63–78. [Google Scholar] [CrossRef] [Green Version]
- Bartels, D. Desiccation tolerance studied in the resurrection plant Craterostigma plantagineum. Integr. Comp. Biol. 2005, 45, 696–701. [Google Scholar] [CrossRef]
- Beebe, S.; Rao, I.; Blair, M.; Acosta, J. Phenotyping common beans for adaptation to drought. Front. Physiol. 2013, 4, 35. [Google Scholar] [CrossRef] [Green Version]
- Bajwa, A.A.; Farooq, M. Seed priming with sorghum water extract and benzyl amino purine along with surfactant improves germination metabolism and early seedling growth of wheat. Arch. Agron. Soil Sci. 2017, 63, 319–329. [Google Scholar] [CrossRef]
- Villar-Salvador, P.; Heredia, N.; Millard, P. Remobilization of acorn nitrogen for seedling growth in holm oak (Quercus ilex), cultivated with contrasting nutrient availability. Tree Physiol. 2010, 30, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Kumawat, A.; Yadav, D.; Samadharmam, K.; Rashmi, I. Soil and water conservation measures for agricultural sustainability. In Soil Moisture Importance; IntechOpen: London, UK, 2020. [Google Scholar]
- Fazal, A.; Bano, A. Role of plant growth-promoting rhizobacteria (pgpr), biochar, and chemical fertilizer under salinity stress. Commun. Soil Sci. Plant Anal. 2016, 47, 1985–1993. [Google Scholar] [CrossRef]
- Thomas, S.C.; Frye, S.; Gale, N.; Garmon, M.; Launchbury, R.; Machado, N.; Sarah Melamed, S.; Murray, J.; Petroff, A.; Winsborough, C. Biochar mitigates negative effects of salt additions on two herbaceous plant species. J. Environ. Manag. 2013, 129, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Pan, F.; Wang, G.; Zhang, G.; Wang, Y.; Chen, X.; Mao, Z. Effects of biochar on photosynthesis and antioxidative system of Malus hupehensis Rehd. seedlings under replant conditions. Sci. Hortic. 2014, 175, 9–15. [Google Scholar] [CrossRef]
- Atkinson, C.J.; Fitzgerald, J.D.; Hipps, N.A. Potential mechanisms for achieving agricultural benefits from biochar application to temperate soils: A review. Plant Soil 2010, 337, 1–18. [Google Scholar] [CrossRef]
- Zhang, M.; Duan, L.; Zhai, Z.; Li, J.; Tian, X.; Wang, B.; He, Z.; Li, Z. Effects of plant growth regulators on water deficit-induced yield loss in soybean. In Proceedings of the 4th International Crop Science Congress, Brisbane, QLD, Australia, 26 September–1 October 2004; pp. 252–256. [Google Scholar]
- Upreti, K.K.; Sharma, M. Role of plant growth regulators in abiotic stress tolerance. In Abiotic Stress Physiology of Horticultural Crops; Springer: New Delhi, India, 2016; pp. 19–46. [Google Scholar] [CrossRef]
- Miyashita, K.; Tanakamaru, S.; Maitani, T.; Kimura, K. Recovery responses of photosynthesis, transpiration, and stomatal conductance in kidney bean following drought stress. Environ. Exp. Bot. 2005, 53, 205–214. [Google Scholar] [CrossRef]
- Sharma, P.; Dubey, R.S. Drought induces oxidative stress and enhances the activities of antioxidant enzymes in growing rice seedlings. Plant Growth Regul. 2005, 46, 209–221. [Google Scholar] [CrossRef]
- Rathinasabapathi, B. Metabolic engineering for stress tolerance: Installing osmoprotectant synthesis pathways. Ann. Bot. 2000, 86, 709–716. [Google Scholar] [CrossRef]
- Ashraf, M.F.M.R.; Foolad, M.R. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Hussain, M.; Malik, M.A.; Farooq, M.; Ashraf, M.Y.; Cheema, M.A. Improving drought tolerance by exogenous application of glycinebetaine and salicylic acid in sunflower. J. Agron. Crop Sci. 2008, 194, 193–199. [Google Scholar] [CrossRef]
- Zhu, J.K. Cell signaling under salt, water and cold stresses. Curr. Opin. Plant Biol. 2001, 4, 401–406. [Google Scholar] [CrossRef]
- Wilkinson, S.; Davies, W.J. ABA-based chemical signalling: The co-ordination of responses to stress in plants. Plant Cell Environ. 2002, 25, 195–210. [Google Scholar] [CrossRef]
- Ober, E.S.; Sharp, R.E. Electrophysiological responses of maize roots to low water potentials: Relationship to growth and ABA accumulation. J. Exp. Bot. 2003, 54, 813–824. [Google Scholar] [CrossRef] [Green Version]
- Hayat, S.; Ali, B.; Ahmad, A. Salicylic acid: Biosynthesis, metabolism and physiological role in plants. In Salicylic Acid: A Plant Hormone; Springer: Dordrecht, The Netherlands, 2007; pp. 1–14. [Google Scholar] [CrossRef]
- Bao, A.K.; Wang, S.M.; Wu, G.Q.; Xi, J.J.; Zhang, J.L.; Wang, C.M. Overexpression of the Arabidopsis H+-PPase enhanced resistance to salt and drought stress in transgenic alfalfa (Medicago sativa L.). Plant Sci. 2009, 176, 232–240. [Google Scholar] [CrossRef]
- Peleg, Z.; Blumwald, E. Hormone balance and abiotic stress tolerance in crop plants. Curr. Opin. Plant Biol. 2011, 14, 290–295. [Google Scholar] [CrossRef]
- Coque, M.; Gallais, A. Genomic regions involved in response to grain yield selection at high and low nitrogen fertilization in maize. Theor. Appl. Genet. 2006, 112, 1205–1220. [Google Scholar] [CrossRef]
- Desikan, R.; Last, K.; Harrett-Williams, R.; Tagliavia, C.; Harter, K.; Hooley, R.; Hancock, J.T.; Neill, S.J. Ethylene-induced stomatal closure in Arabidopsis occurs via AtrbohF-mediated hydrogen peroxide synthesis. Plant J. 2006, 47, 907–916. [Google Scholar] [CrossRef]
- Vurukonda, S.S.K.P.; Vardharajula, S.; Shrivastava, M.; Skz, A. Enhancement of drought stress tolerance in crops by plant growth promoting rhizobacteria. Microbiol. Res. 2016, 184, 13–24. [Google Scholar] [CrossRef]
- Korver, R.A.; Koevoets, I.T.; Testerink, C. Out of shape during stress: A key role for auxin. Trends Plant Sci. 2018, 23, 783–793. [Google Scholar] [CrossRef]
- Colebrook, E.H.; Thomas, S.G.; Phillips, A.L.; Hedden, P. The role of gibberellin signalling in plant responses to abiotic stress. J. Exp. Biol. 2014, 217, 67–75. [Google Scholar] [CrossRef] [Green Version]
- Singh, J.S. Plant growth promoting rhizobacteria. Resonance 2013, 18, 275–281. [Google Scholar] [CrossRef]
- Marulanda, A.; Barea, J.M.; Azcón, R. Stimulation of plant growth and drought tolerance by native microorganisms (AM fungi and bacteria) from dry environments: Mechanisms related to bacterial effectiveness. J. Plant Growth Regul. 2009, 28, 115–124. [Google Scholar] [CrossRef]
- Glick, B.R. Plant growth-promoting bacteria: Mechanisms and applications. Scientifica 2012, 2012, 963401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saharan, B.S.; Nehra, V. Plant growth promoting rhizobacteria: A critical review. Life Sci. Med. Res. 2011, 21, 30. [Google Scholar]
- Sarma, R.K.; Saikia, R. Alleviation of drought stress in mung bean by strain Pseudomonas aeruginosa GGRJ21. Plant Soil 2014, 377, 111–126. [Google Scholar] [CrossRef]
- Augé, R.M. Water relations, drought and vesicular-arbuscular mycorrhizal symbiosis. Mycorrhiza 2001, 11, 3–42. [Google Scholar] [CrossRef]
- Smith, S.E.; Facelli, E.; Pope, S.; Smith, F.A. Plant performance in stressful environments: Interpreting new and established knowledge of the roles of arbuscular mycorrhizas. Plant Soil 2010, 326, 3–20. [Google Scholar] [CrossRef]
- Habibzadeh, Y.; Evazi, A.R.; Abedi, M. Alleviation drought stress of mungbean (Vigna radiata L.) plants by using arbuscular mycorrhizal fungi. J. Agric. Nat. Res. 2014, 1, 1–6. [Google Scholar] [CrossRef]
- Gaur, A.; Adholeya, A. Arbuscular-mycorrhizal inoculation of five tropical fodder crops and inoculum production in marginal soil amended with organic matter. Biol. Fertil. Soil. 2002, 35, 214–218. [Google Scholar] [CrossRef]
- Figueiredo, M.V.; Burity, H.A.; Martinez, C.R.; Chanway, C.P. Alleviation of drought stress in the common bean (Phaseolus vulgaris L.) by co-inoculation with Paenibacillus polymyxa and Rhizobium tropici. Appl. Soil Ecol. 2008, 40, 182–188. [Google Scholar] [CrossRef]
- Beebe, S.E.; Rao, I.M.; Cajiao, C.; Grajales, M. Selection for drought resistance in common bean also improves yield in phosphorus limited and favorable environments. Crop Sci. 2008, 48, 582–592. [Google Scholar] [CrossRef]
- Duc, G.; Agrama, H.; Bao, S.; Berger, J.; Bourion, V.; De Ron, A.M.; Gowda, C.L.L.; Mikic, A.; Millot, D.; Singh, K.B.; et al. Breeding annual grain legumes for sustainable agriculture: New methods to approach complex traits and target new cultivar ideotypes. Crit. Rev. Plant Sci. 2015, 34, 381–411. [Google Scholar] [CrossRef] [Green Version]
- Hall, A. Phenotyping cowpeas for adaptation to drought. Front. Physiol. 2012, 3, 155. [Google Scholar] [CrossRef] [Green Version]
- Khan, H.R.; Paull, J.G.; Siddique, K.H.M.; Stoddard, F.L. Faba bean breeding for drought-affected environments: A physiological and agronomic perspective. Field Crop Res. 2010, 115, 279–286. [Google Scholar] [CrossRef]
- Kashiwagi, J.; Krishnamurthy, L.; Crouch, J.H.; Serraj, R. Variability of root length density and its contributions to seed yield in chickpea (Cicer arietinum L.) under terminal drought stress. Field Crop Res. 2006, 95, 171–181. [Google Scholar] [CrossRef] [Green Version]
- Kumar, J.; Basu, P.S.; Srivastava, E.; Chaturvedi, S.K.; Nadarajan, N.; Kumar, S. Phenotyping of traits imparting drought tolerance in lentil. Crop Pasture Sci. 2012, 63, 547–554. [Google Scholar] [CrossRef]
- Greacen, E.L.; Oh, J.S. Physics of root growth. Nat. New Biol. 1972, 235, 24–25. [Google Scholar] [CrossRef]
- Chauhan, Y.S.; Wallace, D.H.; Johansen, C.; Singh, L. Genotype-by-environment interaction effect on yield and its physiological bases in short-duration pigeonpea. Field Crop Res. 1998, 59, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Saxena, N.P.; Krishnamurthy, L.; Johansen, C. Registration of a drought-resistant chickpea germplasm. Crop Sci. 1993, 33, 1424. [Google Scholar] [CrossRef] [Green Version]
- Nam, N.H.; Chauhan, Y.S.; Johansen, C. Comparison of extra-short-duration pigeonpea with short-season legumes under rainfed conditions on Alfisols. Exp. Agric. 1993, 29, 307–316. [Google Scholar] [CrossRef] [Green Version]
- Bushby, H.V.A.; Lawn, R.J. Accumulation and partitioning of nitrogen and dry matter by contrasting genotypes of mungbean (Vigna radiata (L.) Wilczek). Aust. J. Agric. Res. 1992, 43, 1609–1628. [Google Scholar] [CrossRef]
- Singh, K.B.; Bejiga, G.; Saxena, M.C.; Singh, M. Transferability of chickpea selection indices from normal to drought-prone growing conditions in a Mediterranean environment. J. Agron. Crop Sci. 1995, 175, 57–63. [Google Scholar] [CrossRef]
- Rao, I.; Beebe, S.; Polania, J.; Ricaurte, J.; Cajiao, C.; Garcia, R.; Rivera, M. Can tepary bean be a model for improvement of drought resistance in common bean? Afr. Crop Sci. J. 2013, 21, 265–281. [Google Scholar]
- Muchero, W.; Roberts, P.A.; Diop, N.N.; Drabo, I.; Cisse, N.; Close, T.J.; Muranaka, S.; Baukar, O.; Ehlers, J.D. Genetic architecture of delayed senescence, biomass, and grain yield under drought stress in cowpea. PLoS ONE 2013, 8, e70041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karamanos, A.J.; Papatheohari, A.Y. Assessment of drought resistance of crop genotypes by means of the water potential index. Crop Sci. 1999, 39, 1792–1797. [Google Scholar] [CrossRef]
- Kumar, N.; Kulwal, P.L.; Balyan, H.S.; Gupta, P.K. QTL mapping for yield and yield contributing traits in two mapping populations of bread wheat. Mol. Breed. 2007, 19, 163–177. [Google Scholar] [CrossRef]
- Baldocchi, D.D.; Verma, S.B.; Rosenberg, N.J. Water use efficiency in a soybean field: Influence of plant water stress. Agric. For. Meteorol. 1985, 34, 53–65. [Google Scholar] [CrossRef]
- Amede, T.; Kittlitz, E.V.; Schubert, S. Differential drought responses of faba bean (Vicia faba L.) inbred lines. J. Agron. Crop Sci. 1999, 183, 35–45. [Google Scholar] [CrossRef]
- Lopez, F.B.; Setter, T.L.; McDavid, C.R. Carbon dioxide and light responses of photosynthesis in cowpea and pigeonpea during water deficit and recovery. Plant Physiol. 1987, 85, 990–995. [Google Scholar] [CrossRef]
- Westgate, M.E.; Grant, D.T. Effect of Water Deficits on Seed Development in Soybean: I. Tissue Water Status. Plant Physiol. 1989, 91, 975–979. [Google Scholar] [CrossRef] [Green Version]
- Parsons, L.R.; Howe, T.K. Effects of water stress on the water relations of Phaseolus vulgaris and the drought resistant Phaseolus acutifolius. Physiol. Plant. 1984, 60, 197–202. [Google Scholar] [CrossRef]
- Saxena, N.P. Screening for adaptation to drought: Case studies with Chickpea and Pigeonpea. In Adaptation of Chickpea and Pigeonpea to Abiotic Stresses Proceedings of Consultants’ Workshop, Patancheruvu, India, 19–21 December 1984; International Crops Research Institute for the Semi-Arid Tropics: Patancheruvu, India, 1987; pp. 63–76. [Google Scholar]
- Berger, J.D.; Milroy, S.P.; Turner, N.C.; Siddique, K.H.M.; Imtiaz, M.; Malhotra, R. Chickpea evolution has selected for contrasting phenological mechanisms among different habitats. Euphytica 2011, 180, 1–15. [Google Scholar] [CrossRef]
- Vadez, V.; Soltani, A.; Sinclair, T.R. Modelling possible benefits of root related traits to enhance terminal drought adaptation of chickpea. Field Crop Res. 2012, 137, 108–115. [Google Scholar] [CrossRef] [Green Version]
- Zaman-Allah, M.; Jenkinson, D.M.; Vadez, V. Chickpea genotypes contrasting for seed yield under terminal drought stress in the field differ for traits related to the control of water use. Funct. Plant Biol. 2011, 38, 270–281. [Google Scholar] [CrossRef] [Green Version]
- Zaman-Allah, M.; Jenkinson, D.M.; Vadez, V. A conservative pattern of water use, rather than deep or profuse rooting, is critical for the terminal drought tolerance of chickpea. J. Exp. Bot. 2011, 62, 4239–4252. [Google Scholar] [CrossRef] [Green Version]
- Blair, M.W.; Iriarte, G.; Beebe, S. QTL analysis of yield traits in an advanced backcross population derived from a cultivated Andean× wild common bean (Phaseolus vulgaris L.) cross. Theor. Appl. Genet. 2006, 112, 1149–1163. [Google Scholar] [CrossRef]
- Miklas, P.N.; Kelly, J.D.; Beebe, S.E.; Blair, M.W. Common bean breeding for resistance against biotic and abiotic stresses: From classical to MAS breeding. Euphytica 2006, 147, 105–131. [Google Scholar] [CrossRef]
- Haghighi, K.R.; Ascher, P.D. Fertile, intermediate hybrids between Phaseolus vulgaris and P. acutifolius from congruity backcrossing. Sex. Plant Reprod. 1988, 1, 51–58. [Google Scholar] [CrossRef]
- White, J.W.; Singh, S.P. Sources and inheritance of earliness in tropically adapted indeterminate common bean. Euphytica 1991, 55, 15–19. [Google Scholar] [CrossRef]
- Devi, M.J.; Sinclair, T.R.; Beebe, S.E.; Rao, I.M. Comparison of common bean (Phaseolus vulgaris L.) genotypes for nitrogen fixation tolerance to soil drying. Plant Soil 2013, 364, 29–37. [Google Scholar] [CrossRef]
- Rao, I.M. Advances in Improving Adaptation of Common Bean and Brachiaria forage Grasses to Abiotic Stress in the Tropics; CRC Press: Boca Raton, FL, USA, 2014. [Google Scholar]
- Homrich, M.S.; Wiebke-Strohm, B.; Weber, R.L.M.; Bodanese-Zanettini, M.H. Soybean genetic transformation: A valuable tool for the functional study of genes and the production of agronomically improved plants. Genet. Mol. Biol. 2012, 35, 998–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khazaei, H.; Street, K.; Bari, A.; Mackay, M.; Stoddard, F.L. The FIGS (Focused Identification of Germplasm Strategy) approach identifies traits related to drought adaptation in Vicia faba genetic resources. PLoS ONE 2013, 8, e63107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khazaei, H.; Street, K.; Santanen, A.; Bari, A.; Stoddard, F.L. Do faba bean (Vicia faba L.) accessions from environments with contrasting seasonal moisture availabilities differ in stomatal characteristics and related traits? Genet. Resour. Crop Evol. 2013, 60, 2343–2357. [Google Scholar] [CrossRef]
- Odeny, D.A. The potential of pigeonpea (Cajanus cajan L.) Millsp.) in Africa. In Natural Resources Forum; Blackwell Publishing Ltd.: Oxford, UK, 2007; pp. 297–305. [Google Scholar] [CrossRef]
- Brachi, B.; Morris, G.P.; Borevitz, J.O. Genome-wide association studies in plants: The missing heritability is in the field. Genome Biol. 2011, 12, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Jha, U.C.; Bohra, A.; Nayyar, H. Advances in “omics” approaches to tackle drought stress in grain legumes. Plant Breed. 2020, 139, 1–27. [Google Scholar] [CrossRef]
- Dhanapal, A.P.; Ray, J.D.; Singh, S.K.; Hoyos-Villegas, V.; Smith, J.R.; Purcell, L.C.; King, C.A.; Cregan, P.C.; Song, Q.; Fritschi, F.B. Genome-wide association study (GWAS) of carbon isotope ratio (δ13C) in diverse soybean [Glycine max (L.) Merr.] genotypes. Theor. Appl. Genet. 2015, 128, 73–91. [Google Scholar] [CrossRef]
- Kaler, A.S.; Ray, J.D.; Schapaugh, W.T.; Asebedo, A.R.; King, C.A.; Gbur, E.E.; Purcell, L.C. Association mapping identifies loci for canopy temperature under drought in diverse soybean genotypes. Euphytica 2018, 214, 1–18. [Google Scholar] [CrossRef]
- Hoyos-Villegas, V.; Song, Q.; Kelly, J.D. Genome-wide association analysis for drought tolerance and associated traits in common bean. Plant Genome 2017, 10, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Cortés, A.J.; Blair, M.W. Genotyping by sequencing and genome–environment associations in wild common bean predict widespread divergent adaptation to drought. Front. Plant Sci. 2018, 9, 128. [Google Scholar] [CrossRef] [Green Version]
- Teran, J.C.B.M.Y.; Konzen, E.R.; Palkovic, A.; Tsai, S.M.; Rao, I.M.; Beebe, S.; Gepts, P. Effect of drought stress on the genetic architecture of photosynthate allocation and remobilization in pods of common bean (Phaseolus vulgaris L.), a key species for food security. BMC Plant Biol. 2019, 19, 171. [Google Scholar] [CrossRef]
- Radhika, P.; Gowda, S.J.M.; Kadoo, N.Y.; Mhase, L.B.; Jamadagni, B.M.; Sainani, M.N.; Chandra, S.; Gupta, V.S. Development of an integrated intraspecific map of chickpea (Cicer arietinum L.) using two recombinant inbred line populations. Theor. Appl. Genet. 2007, 115, 209–216. [Google Scholar] [CrossRef] [Green Version]
- Witcombe, J.R.; Hollington, P.A.; Howarth, C.J.; Reader, S.; Steele, K.A. Breeding for abiotic stresses for sustainable agriculture. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 703–716. [Google Scholar] [CrossRef] [Green Version]
- Eathington, S.R.; Crosbie, T.M.; Edwards, M.D.; Reiter, R.S.; Bull, J.K. Molecular markers in a commercial breeding program. Crop Sci. 2007, 47, 154–163. [Google Scholar] [CrossRef]
- Bohra, A. Emerging paradigms in genomics-based crop improvement. Sci. World J. 2013, 2013, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Varshney, R.K.; Gaur, P.M.; Chamarthi, S.K.; Krishnamurthy, L.; Tripathi, S.; Kashiwagi, J.; Samineni, S.; Singh, V.K.; Thudi, M.; Jaganathan, D. Fast-track introgression of “QTL-hotspot” for root traits and other drought tolerance traits in JG 11, an elite and leading variety of chickpea. Plant Genome 2013, 6, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Haleem, H.; Lee, G.J.; Boerma, R.H. Identification of QTL for increased fibrous roots in soybean. Theor. Appl. Genet. 2011, 122, 935–946. [Google Scholar] [CrossRef]
- Manavalan, L.P.; Prince, S.J.; Musket, T.A.; Chaky, J.; Deshmukh, R.; Vuong, T.D.; Song, L.; Cregan, P.B.; Nelson, J.C.; Shannon, J.G.; et al. Identification of novel QTL governing root architectural traits in an interspecific soybean population. PLoS ONE 2015, 10, e0120490. [Google Scholar] [CrossRef] [Green Version]
- Muchero, W.; Ehlers, J.D.; Close, T.J.; Roberts, P.A. Mapping QTL for drought stress-induced premature senescence and maturity in cowpea [Vigna unguiculata (L.) Walp.]. Theor. Appl. Genet. 2009, 118, 849–863. [Google Scholar] [CrossRef]
- Prince, S.J.; Song, L.; Qiu, D.; Dos Santos, J.V.M.; Chai, C.; Joshi, T.; Patil, G.; Valliyodan, B.; Vuong, T.D.; Murphy, M.; et al. Genetic variants in root architecture-related genes in a Glycine soja accession, a potential resource to improve cultivated soybean. BMC Genom. 2015, 16, 132. [Google Scholar] [CrossRef] [Green Version]
- Burridge, J.D.; Schneider, H.M.; Huynh, B.L.; Roberts, P.A.; Bucksch, A.; Lynch, J.P. Genome-wide association mapping and agronomic impact of cowpea root architecture. Theor. Appl. Genet. 2017, 130, 419–431. [Google Scholar] [CrossRef]
- Liao, H.; Yan, X.; Rubio, G.; Beebe, S.E.; Blair, M.W.; Lynch, J.P. Genetic mapping of basal root gravitropism and phosphorus acquisition efficiency in common bean. Funct. Plant Biol. 2004, 31, 959–970. [Google Scholar] [CrossRef] [PubMed]
- Hamwieh, A.; Imtiaz, M.; Malhotra, R.S. Multi-environment QTL analyses for drought-related traits in a recombinant inbred population of chickpea (Cicer arientinum L.). Theor. Appl. Genet. 2013, 126, 1025–1038. [Google Scholar] [CrossRef] [PubMed]
- Fondevilla, S.; Almeida, N.F.; Satovic, Z.; Rubiales, D.; Patto, M.C.V.; Cubero, J.I.; Torres, A.M. Identification of common genomic regions controlling resistance to Mycosphaerella pinodes, earliness and architectural traits in different pea genetic backgrounds. Euphytica 2011, 182, 43–52. [Google Scholar] [CrossRef]
- Saxena, K.B.; Singh, G.; Gupta, H.S.; Mahajan, V.; Kumar, R.V.; Singh, B.; Vales, M.I.; Sultana, R. Enhancing the livelihoods of Uttarakhand farmers by introducing pigeonpea cultivation in hilly areas. J. Food Legume. 2011, 24, 128–132. [Google Scholar]
- Varshney, R.K.; Pandey, M.K.; Bohra, A.; Singh, V.K.; Thudi, M.; Saxena, R.K. Toward the sequence-based breeding in legumes in the post-genome sequencing era. Theor. Appl. Genet. 2019, 132, 797–816. [Google Scholar] [CrossRef] [Green Version]
- Collins, N.C.; Tardieu, F.; Tuberosa, R. QTL approaches for improving crop performance under abiotic stress conditions: Where do we stand. Plant Physiol. 2008, 147, 469–486. [Google Scholar] [CrossRef] [Green Version]
- Singh, B.; Bohra, A.; Mishra, S.; Joshi, R.; Pandey, S. Embracing new-generation ‘omics’ tools to improve drought tolerance in cereal and food-legume crops. Biol. Plant. 2015, 59, 413–428. [Google Scholar] [CrossRef]
- Goddard, M.E.; Hayes, B.J. Genomic selection. J. Anim. Breed. Genet. 2007, 124, 323–330. [Google Scholar] [CrossRef]
- Meuwissen, T.H.; Hayes, B.J.; Goddard, M.E. Prediction of total genetic value using genome-wide dense marker maps. Genetics 2001, 157, 1819–1829. [Google Scholar] [CrossRef]
- Hickey, L.T.; Hafeez, A.N.; Robinson, H.; Jackson, S.A.; Leal-Bertioli, S.C.; Tester, M.; Gao, C.; Godwin, I.D.; Hayes, B.J.; Wulff, B.B.H. Breeding crops to feed 10 billion. Nat. Biotechnol. 2019, 37, 744–754. [Google Scholar] [CrossRef] [Green Version]
- Crossa, J.; Pérez-Rodríguez, P.; Cuevas, J.; Montesinos-López, O.; Jarquín, D.; de los Campos, G.; Burgueño, J.; Camacho-González, J.M.; Pérez-Elizalde, S.; Beyene, Y.; et al. Genomic selection in plant breeding: Methods, models, and perspectives. Trends Plant Sci. 2017, 22, 961–975. [Google Scholar] [CrossRef]
- Li, Y.; Ruperao, P.; Batley, J.; Edwards, D.; Khan, T.; Colmer, T.D.; Pang, J.; Kadambot, H.M.; Siddique, K.H.M.; Sutton, T. Investigating drought tolerance in chickpea using genome-wide association mapping and genomic selection based on whole-genome resequencing data. Front. Plant Sci. 2018, 9, 190. [Google Scholar] [CrossRef] [Green Version]
- Ashraf, M. Inducing drought tolerance in plants: Recent advances. Biotechnol. Adv. 2010, 28, 169–183. [Google Scholar] [CrossRef]
- Kishor, P.K.; Hong, Z.; Miao, G.H.; Hu, C.A.A.; Verma, D.P.S. Overexpression of [delta]-pyrroline-5-carboxylate synthetase increases proline production and confers osmotolerance in transgenic plants. Plant Physiol. 1995, 108, 1387–1394. [Google Scholar] [CrossRef] [Green Version]
- Anbazhagan, K.; Bhatnagar-Mathur, P.; Vadez, V.; Dumbala, S.R.; Kishor, P.K.; Sharma, K.K. DREB1A overexpression in transgenic chickpea alters key traits influencing plant water budget across water regimes. Plant Cell Rep. 2015, 34, 199–210. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zhang, J.; Zhang, J.; Hao, L.; Hua, J.; Duan, L.; Zhang, M.; Li, Z. Expression of an Arabidopsis molybdenum cofactor sulphurase gene in soybean enhances drought tolerance and increases yield under field conditions. Plant Biotechnol. J. 2013, 11, 747–758. [Google Scholar] [CrossRef]
- Ronde, J.A.D.; Cress, W.A.; Krüger, G.H.J.; Strasser, R.J.; Van Staden, J. Photosynthetic response of transgenic soybean plants, containing an Arabidopsis P5CR gene, during heat and drought stress. J. Plant Physiol. 2004, 161, 1211–1224. [Google Scholar] [CrossRef]
- Iuchi, S.; Kobayashi, M.; Yamaguchi-Shinozaki, K.; Shinozaki, K. A stress-inducible gene for 9-cis-epoxycarotenoid dioxygenase involved in abscisic acid biosynthesis under water stress in drought-tolerant cowpea. Plant Physiol. 2000, 123, 553–562. [Google Scholar] [CrossRef] [Green Version]
- Deokar, A.A.; Kondawar, V.; Jain, P.K.; Karuppayil, S.M.; Raju, N.L.; Vadez, V.; Varshney, R.K.; Srinivasan, R. Comparative analysis of expressed sequence tags (ESTs) between drought-tolerant and-susceptible genotypes of chickpea under terminal drought stress. BMC Plant Biol. 2011, 11, 70. [Google Scholar] [CrossRef] [Green Version]
- Nayak, S.N.; Balaji, J.; Upadhyaya, H.D.; Hash, C.T.; Kishor, P.K.; Chattopadhyay, D.; Rodriquez, L.M.; Blair, M.W.; Baum, M.; McNally, K.; et al. Isolation and sequence analysis of DREB2A homologues in three cereal and two legume species. Plant Sci. 2009, 177, 460–467. [Google Scholar] [CrossRef] [Green Version]
- Hiremath, P.J.; Farmer, A.; Cannon, S.B.; Woodward, J.; Kudapa, H.; Tuteja, R.; Kumar, A.; BhanuPrakash, A.; Mulaosmanovic, B.; Gujaria, N.; et al. Large-scale transcriptome analysis in chickpea (Cicer arietinum L.), an orphan legume crop of the semi-arid tropics of Asia and Africa. Plant Biotechnol. J. 2011, 9, 922–931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azeem, F.; Bilal, A.; Rana, M.A.; Muhammad, A.A.; Habibullah, N.; Sabir, H.; Sumaira, R.; Hamid, M.; Usama, A.; Muhammad, A. Drought affects aquaporins gene expression in important pulse legume chickpea (Cicer arietinum L.). Pak. J. Bot. 2019, 51, 81–88. [Google Scholar] [CrossRef]
- Baloda, A.; Madanpotra, S. Transformation of mungbean plants for salt and drought tolerance by introducing a gene for an osmoprotectant glycine betaine. J. Plant Stress Physiol. 2017, 3, 5–11. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, R.; Kumar, S.; Kobayashi, Y.; Kusunoki, K.; Tripathi, P.; Kobayashi, Y.; Koyama, H.; Sahoo, L. Comparative genome-wide analysis of WRKY transcription factors in two Asian legume crops: Adzuki bean and Mung bean. Sci. Rep. 2018, 8, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Zhu, J.; Li, X.; Wang, S.; Wu, J. Salt and drought stress and ABA responses related to bZIP genes from V. radiata and V. angularis. Gene 2018, 651, 152–160. [Google Scholar] [CrossRef]
- Cui, X.H.; Hao, F.S.; Chen, H.; Chen, J.; Wang, X.C. Expression of the Vicia faba VfPIP1 gene in Arabidopsis thaliana plants improves their drought resistance. J. Plant Res. 2008, 121, 207–214. [Google Scholar] [CrossRef]
- Cortés, A.J.; This, D.; Chavarro, C.; Madriñán, S.; Blair, M.W. Nucleotide diversity patterns at the drought-related DREB2 encoding genes in wild and cultivated common bean (Phaseolus vulgaris L.). Theor. Appl. Genet. 2012, 125, 1069–1085. [Google Scholar] [CrossRef]
- Barrera-Figueroa, B.E.; Peña-Castro, J.M.; Acosta-Gallegos, J.A.; Ruiz-Medrano, R.; Xoconostle-Cázares, B. Isolation of dehydration-responsive genes in a drought tolerant common bean cultivar and expression of a group 3 late embryogenesis abundant mRNA in tolerant and susceptible bean cultivars. Funct. Plant Biol. 2007, 34, 368–381. [Google Scholar] [CrossRef]
- Montalvo-Hernández, L.; Piedra-Ibarra, E.; Gómez-Silva, L.; Lira-Carmona, R.; Acosta-Gallegos, J.A.; Vazquez-Medrano, J.; Xoconostle-Cázares, B.; Ruíz-Medrano, R. Differential accumulation of mRNAs in drought-tolerant and susceptible common bean cultivars in response to water deficit. New Phytol. 2008, 177, 102–113. [Google Scholar] [CrossRef]
- Manavalan, L.P.; Guttikonda, S.K.; Phan Tran, L.S.; Nguyen, H.T. Physiological and molecular approaches to improve drought resistance in soybean. Plant Cell Physiol. 2009, 50, 1260–1276. [Google Scholar] [CrossRef] [Green Version]
- Bhatnagar-Mathur, P.; Vadez, V.; Devi, M.J.; Lavanya, M.; Vani, G.; Sharma, K.K. Genetic engineering of chickpea (Cicer arietinum L.) with the P5CSF129A gene for osmoregulation with implications on drought tolerance. Mol. Breed. 2009, 23, 591–606. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.S.; Chen, Q.S.; Xin, D.W.; Qi, Z.M.; Zhang, C.; Li, S.N.; Jin, Y.; Li, M.; Mei, H.Y.; Su, A.Y.; et al. Overexpression of GmBIN2, a soybean glycogen synthase kinase 3 gene, enhances tolerance to salt and drought in transgenic Arabidopsis and soybean hairy roots. J. Integr. Agric. 2018, 17, 1959–1971. [Google Scholar] [CrossRef]
- Yoo, J.H.; Park, C.Y.; Kim, J.C.; Do Heo, W.; Cheong, M.S.; Park, H.C.; Kim, M.C.; Moon, B.C.; Choi, M.S.; Kang, Y.H.; et al. Direct interaction of a divergent CaM isoform and the transcription factor, MYB2, enhances salt tolerance in Arabidopsis. J. Biol. Chem. 2005, 280, 3697–3706. [Google Scholar] [CrossRef] [Green Version]
- Guenther, J.F.; Chanmanivone, N.; Galetovic, M.P.; Wallace, I.S.; Cobb, J.A.; Roberts, D.M. Phosphorylation of soybean nodulin 26 on serine 262 enhances water permeability and is regulated developmentally and by osmotic signals. Plant Cell 2003, 15, 981–991. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, E.; Karakaya, H.C.; Knap, H.T. Molecular characterization of two soybean homologs of Arabidopsis thaliana CLAVATA1 from the wild type and fasciation mutant. Biochim. Biophys. Acta Gene Struct. Express. 2000, 1491, 333–340. [Google Scholar] [CrossRef]
- An, J.; Cheng, C.; Hu, Z.; Chen, H.; Cai, W.; Yu, B. The Panax ginseng PgTIP1 gene confers enhanced salt and drought tolerance to transgenic soybean plants by maintaining homeostasis of water, salt ions and ROS. Environ. Exp. Bot. 2018, 155, 45–55. [Google Scholar] [CrossRef]
- Savitri, E.S.; Fauziah, S.M. Characterization of drought tolerance of GmDREB2 soybean mutants (Glycine max (L.) Merr) by ethyl methane sulfonate induction. AIP Conf. Proc. 2018, 2019, 020017. [Google Scholar] [CrossRef]
- Li, D.H.; Chen, F.J.; Li, H.Y.; Li, W.; Guo, J.J. The soybean GmRACK1 gene plays a role in drought tolerance at vegetative stages. Russ. J. Plant Physiol. 2018, 65, 541–552. [Google Scholar] [CrossRef]
- Kim, H.J.; Cho, H.S.; Pak, J.H.; Kwon, T.; Lee, J.H.; Kim, D.H.; Lee, D.H.; Kim, C.; Chung, Y.S. Confirmation of drought tolerance of ectopically expressed AtABF3 gene in soybean. Mol. Cells 2018, 41, 413. [Google Scholar]
- Li, Y.; Chen, Q.; Nan, H.; Li, X.; Lu, S.; Zhao, X.; Liu, B.; Guo, C.; Kong, F.; Cao, D. Overexpression of GmFDL19 enhances tolerance to drought and salt stresses in soybean. PLoS ONE 2017, 12, e0179554. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Chi, Y.; Meng, Q.; Wang, X.; Yu, D. GmSK1, an SKP1 homologue in soybean, is involved in the tolerance to salt and drought. Plant Physiol. Biochem. 2018, 127, 25–31. [Google Scholar] [CrossRef]
- Yang, S.; Pang, W.; Ash, G.; Harper, J.; Carling, J.; Wenzl, P.; Huttner, E.; Zong, E.; Kilian, A. Low level of genetic diversity in cultivated pigeonpea compared to its wild relatives is revealed by diversity arrays technology. Theor. Appl. Genet. 2006, 113, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Muchero, W.; Ehlers, J.D.; Roberts, P.A. Restriction site polymorphism-based candidate gene mapping for seedling drought tolerance in cowpea [Vigna unguiculata (L.) Walp.]. Theor. Appl. Genet. 2010, 120, 509–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Q.X.; Liu, Y.F.; Hu, X.Y.; Zhang, W.K.; Ma, B.; Chen, S.Y.; Zhang, J.S. Identification of miRNAs and their target genes in developing soybean seeds by deep sequencing. BMC Plant Biol. 2011, 11, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joshi, T.; Yan, Z.; Libault, M.; Jeong, D.H.; Park, S.; Green, P.J.; Sherrier, D.J.; Farmer, A.; May, G.; Meyers, B.C.; et al. Prediction of novel miRNAs and associated target genes in Glycine max. BMC Bioinform. 2010, 11, S14. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Dong, S.; Liu, L.; Ma, Y.; Li, S.; Zu, W. Transcriptome profiling reveals PEG-simulated drought, heat and combined stress response mechanisms in soybean. Comput. Biol. Chem. 2018, 77, 413–419. [Google Scholar] [CrossRef]
- Prince, S.J.; Joshi, T.; Mutava, R.N.; Syed, N.; Vitor, M.D.S.J.; Patil, G.; Song, L.; Wang, J.J.; Lin, L.; Chen, W.; et al. Comparative analysis of the drought-responsive transcriptome in soybean lines contrasting for canopy wilting. Plant Sci. 2015, 240, 65–78. [Google Scholar] [CrossRef]
- Domenico, S.D.; Bonsegna, S.; Horres, R.; Pastor, V.; Taurino, M.; Poltronieri, P.; Imtiaz, M.; Kahl, G.; Flors, V.; Winter, P.; et al. Transcriptomic analysis of oxylipin biosynthesis genes and chemical profiling reveal an early induction of jasmonates in chickpea roots under drought stress. Plant Physiol. Biochem. 2012, 61, 115–122. [Google Scholar] [CrossRef]
- Strozycki, P.M.; Szczurek, A.; Lotocka, B.; Figlerowicz, M.; Legocki, A.B. Ferritins and nodulation in Lupinus luteus: Iron management in indeterminate type nodules. J. Exp. Bot. 2007, 58, 3145–3153. [Google Scholar] [CrossRef] [Green Version]
- Varshney, R.K.; Dubey, A. Novel genomic tools and modern genetic and breeding approaches for crop improvement. J. Plant Biochem. Biotechnol. 2009, 18, 127–138. [Google Scholar] [CrossRef] [Green Version]
- Pandey, A.; Chakraborty, S.; Datta, A.; Chakraborty, N. Proteomics approach to identify dehydration responsive nuclear proteins from chickpea (Cicer arietinum L.). Mol. Cell. Proteom. 2008, 7, 88–107. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Song, Q.; Li, D.; Yang, X.; Li, D. Multifunctional roles of plant dehydrins in response to environmental stresses. Front. Plant Sci. 2017, 8, 1018. [Google Scholar] [CrossRef] [Green Version]
- Das, A.; Rushton, P.J.; Rohila, J.S. Metabolomic profiling of soybeans (Glycine max L.) reveals the importance of sugar and nitrogen metabolism under drought and heat stress. Plants 2017, 6, 21. [Google Scholar] [CrossRef] [Green Version]
- Cai, Y.; Chen, L.; Liu, X.; Sun, S.; Wu, C.; Jiang, B.; Han, T.; Hou, W. CRISPR/Cas9-mediated genome editing in soybean hairy roots. PLoS ONE 2015, 10, e0136064. [Google Scholar] [CrossRef]
- Pandey, M.K.; Roorkiwal, M.; Singh, V.K.; Ramalingam, A.; Kudapa, H.; Thudi, M.; Chitikineni, A.; Rathore, A.; Varshney, R.K. Emerging genomic tools for legume breeding: Current status and future prospects. Front. Plant Sci. 2016, 7, 455. [Google Scholar] [CrossRef] [Green Version]
- Jain, M.; Misra, G.; Patel, R.K.; Priya, P.; Jhanwar, S.; Khan, A.W.; Shah, N.; Singh, V.K.; Garg, R.; Jeena, G.; et al. A draft genome sequence of the pulse crop chickpea (Cicer arietinum L.). Plant J. 2013, 74, 715–729. [Google Scholar] [CrossRef]
- Jacobs, T.B.; LaFayette, P.R.; Schmitz, R.J.; Parrott, W.A. Targeted genome modifications in soybean with CRISPR/Cas9. BMC Biotechnol. 2015, 15, 16. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Grain Legumes | Phenological Stages | Yield Losses (%) | References |
|---|---|---|---|
| Chickpea (Cicer arietinum L.) | Reproductive | 45–69 | Farooq et al. [35]; Nayyar et al. [81] |
| Late ripening | 49–54 | Samarah et al. [82] | |
| Anthesis | 27–40 | Mafakheri et al. [45] | |
| Pigeonpea (Cajanus cajan L.) | Reproductive | 40–55 | Farooq et al. [35] |
| Flowering | 40–55 | Nam et al. [83] | |
| Soybean (Lens culinaris L.) | Reproductive | 46–71 | Farooq et al. [35] |
| pod setting | 45–50 | Kobraee et al. [84] | |
| Grain filling | 42 | Maleki et al. [85] | |
| Pod setting | 73–82 | Wei et al. [86] | |
| Common bean (Phaseolus vulgaris L.) | Reproductive | 58–87 | Farooq et al. [35]; Martínez et al. [87] |
| Flowering | 49 | Rosales-Serna et al. [88] | |
| Pod filling | 40 | Ghanbari et al. [74] | |
| Cowpea (Vigna unguiculata L.) | Reproductive | 34–66 | Ahmed and Suliman [89] |
| Pod filling | 29 | Farooq et al. [11] | |
| Reproductive | 60 | Ogbonnaya et al. [90] | |
| Faba bean (Vicia faba L.) | Grain filling | 68 | Ghassemi-Golezani and Hosseinzadeh-Mahootchy [91] |
| Black gram (Vigna mungo L.) | Flowering | 31–57 | Baroowa and Gogoi [92] |
| Reproductive | 26 | Baroowa and Gogoi [93] | |
| Lentil (Lens culinaris L.) | Reproductive | 24 | Allahmoradi et al. [94] |
| Pod development | 70 | Shrestha et al. [69] |
| Parameter | Observed Yield Losses (%) | Observed Water Deficit (%) | |
|---|---|---|---|
| Grain legumes | Lentil | 21.7 | >65 |
| Groundnut | 28.6 | >65 | |
| Faba bean | 40 | >65 | |
| Pigeon pea | 21.8 | 60–65 | |
| Soybean | 28.0 | 60–65 | |
| Chickpea | 40.4 | 60–65 | |
| Cowpea | 44.3 | 60–65 | |
| Green gram | 45.3 | 60–65 | |
| Common bean | 60.8 | 60–65 | |
| Field pea | 20.2 | <60 | |
| Plant phenological stages | Vegetative phase | 15.5 | >65 |
| Reproductive stages (flowering and pod filling) | 43.4 | 60 or <60 | |
| Throughout the growing season | 42.1 | 60 or <60 | |
| Early reproductive (flowering) | 37.5 | >65 | |
| Late reproductive (pod filling) | 28.5 | >65 | |
| Soil texture | Medium-textured soil | 63.8 | >60 |
| Fine-textured soil | 30.9 | >60 | |
| Coarse-textured soil | 19.8 | >60 | |
| Agroclimatic-region | Tropical region | 35.4 | >60 |
| Non-tropical region | 36.6 | >60 | |
| Phytohormones | Functions | References |
|---|---|---|
| Abscisic acid | • Manages the water status of the plant by regulating the guard cell | Zhu [147] |
| • Transmits signals from the root to the shoot, leading in the closure of leaf stomata and a reduction in transpiration | Wilkinson and Davies [148] | |
| • Induces genes coding for protein and enzymes linked to drought tolerance | Ali et al. [124] | |
| • Limit excessive ethylene production and preserve root and shoot growth | Ober and Sharp [149] | |
| Salicylic acid | • Improved membrane stability index (MSI), photosynthetic parameters, leaf water potential, carbonic anhydrase, activity of nitrate reductase, relative water content and chlorophyll content | Hayat et al. [150] |
| Jasmonic acid | • Play a crucial part in antioxidant responses produced by drought, particularly ascorbate metabolism | Bao et al. [151] |
| Cytokinins | • Late leaf senescence | Peleg and Blumwald [152] |
| • Encouraging root development and more efficient nutrient uptake | Coque and Gallais [153] | |
| Ethylene | • Produces H2O2 in the guard cell, which causes stomatal closure | Desikan et al. [154] |
| • Abscission of the leaves | Salazar et al. [106] | |
| • Reduced root and shoot growth due to plant homoeostasis | Vurukonda et al. [155] | |
| Auxin | • Phenotypic plasticity with developmental changes to root system architecture and root growth | Korver et al. [156] |
| Gibberellin | • Signaling in either growth repression or promotion as a result of stress-induced growth regulation | Colebrook et al. [157] |
| Useful Traits | References |
|---|---|
| Short growth duration and erect plant type | Hall [170] |
| Root length, rooting depth, root density and fibrous root system | Khan et al. [171], Hall [170] and Duc et al. [169] |
| Prolific root system | Kashiwagi et al. [172] |
| Rooting depth | Farooq et al. [11] |
| Root length and dry root weight | Kumar et al. [173] |
| Osmotic adjustment of root | Greacen and Oh [174] |
| Root and shoot biomass | Chauhan et al. [175] |
| Smaller growth duration | Saxena et al. [176] |
| Extra short duration | Nam et al. [177] |
| Early flowering, podding and maturity | Duc et al. [169] |
| Dry matter partitioning | Bushby and Lawn [178] |
| Early maturity, grain size and short stature | Singh et al. [179] |
| Canopy biomass, stem biomass reduction, pod partitioning and harvest index | Rao et al. [180] |
| Delayed senescence | Muchero et al. [181] |
| Cooler canopies and higher stomatal conductance | Duc et al. [169] |
| Leaf water potential index | Karamanos and Papatheohari [182] |
| Leaf area maintenance | Lopez et al. [43] |
| Leaf relative water content (RWC) | Kumar et al. [183] |
| Presence of dense leaf pubescence | Baldochi et al. [184] |
| Leaf temperature and carbon isotope discrimination (Δ13C) | Khan et al. [171] |
| Earliness and vigorous growth | Khan et al. [171] |
| Water-use efficiency | Amede et al. [185] |
| Osmotic adjustment | Lopez et al. [186] |
| Carbohydrate store and remobilization | Westgate et al. [187] |
| Grain Legumes | Development Method and Institution Involved | Achievement in Breeding | Addressed Characteristics |
|---|---|---|---|
| Chickpea |
| ||
| Common bean |
| ||
| Faba bean | • Cytoplasmic genetic male sterility (CGMS) and Departamento de Genetica, Instituto de Biociencias, Universidade Federal do Rio Grande do Sul, Brazil [200] | • Drought-escaping and drought-tolerant germplasm selection [201,202] |
|
| Pigeonpea | • DSI and ICRISAT [175] | • Drought-tolerant cultivated germplasm selection [203] |
|
| Grain Legumes | QTL Name | Characteristics | References |
|---|---|---|---|
| Soybean (Glycine max) | FR_Gm01, FR_Gm03, FR_Gm04, FR_Gm08, FR_Gm20 | Fibrous rooting/surface area | Abdel-Haleem et al. [216] |
| SA_Gm06, TRL_Gm06, RDL3_Gm07, TRL_Gm08 | Root length | Manavalan et al. [217] | |
| LRN_Gm08 | Lateral root number | Manavalan et al. [217] | |
| Cowpea (Vigna unguiculata) | Dro-1, Dro-2 | Drought induced senescence | Muchero et al. [218] |
| Mat-1, Mat-2 | Maturity | Muchero et al. [218] | |
| RDT2_Gm08 | Root thickness | Prince et al. [219] | |
| Brg10 | Basal root angle | Burridge et al. [220] | |
| RD1–1 | Root diameter | Burridge et al. [220] | |
| MW6–6 | Median width | Burridge et al. [220] | |
| WA10–10 | Width accumulation | Burridge et al. [220] | |
| Common bean (Phaseolus vulgaris) | Brg1.1, Brg5.1, Brg5.2 | Basal root angle | Liao et al. [221] |
| Chickpea (Cicer arietinum) | Qncl.Sw1 NCPGR-50, | Grain weight | Radhika et al. [211] |
| TR-50, SCEA19, TAA-58 | Grain weight | Hamwieh et al. [222] | |
| RLD4 | Root length density | Jaganathan et al. [105] | |
| RSA6 | Root surface area | Jaganathan et al. [105] | |
| RDWR4 | Root dry weight ratio | Jaganathan et al. [105] | |
| QTL-hotspot | Rooting depth or root length density | Jaganathan et al. [105] | |
| H6C-07, H5G-01, H6C-07, H1B-04 | Grain yield | Hamwieh et al. [222] | |
| TA-113, TR-58, H6C-07, H1F-21 5 | Harvest index | Hamwieh et al. [222] | |
| H6C-07 | Drought-resistance score | Hamwieh et al. [222] | |
| Pea (Pisum sativum) | rl1, rl2, rl3 | Root length | Fondevilla et al. [223] |
| Pegion pea (Cajanus cajan) | QTL-RF-1, QTL-RF-2, QTL-RF-3, QTL-RF-4 | Fertility restoration | Saxena et al. [224] |
| Grain Legumes | Identified Gene | Functions | References |
|---|---|---|---|
| Chick pea (Cicer arietinum) | MyB, AP2/ERF, XPB1 | Transcription factors | Deokar et al. [239] |
| DREB2A | Transcription factors | Nayak et al. [240] | |
| MYB, WRKY, bZIP | Transcription factors | Hiremath et al. [241] | |
| Aquaporins | Drought-stress tolerance | Azeem et al. [242] | |
| Mungbean (Vigna radiata) | codA | Improve abiotic-stress tolerance | Baloda and Madanpotra [243] |
| VrWRKY | Enhance abiotic-stress tolerance | Srivastava et al. [244] | |
| VrbZIP | Drought-responsive gene | Wang et al. [245] | |
| Broad bean (Vicia faba) | VfPIP1 | Aquaporin/water transport | Cui et al. [246] |
| Common bean (Phaseolus vulgaris) | DREB2B | Non-ABA dependent responses | Cortés et al. [247] |
| Asr1, Asr2 | ABA signaling pathway | Cortés et al. [248] | |
| PvLEA3 | Protein stabilization | Barrera-Figueroa et al. [249] | |
| Aquaporin | Water allocation | Montalvo-Hernández et al. [250] | |
| Soybean (Glycine max) | GmNAC, GmDREB, GmZIP, ERF089 | Transcription factors | Manavalan et al. [250] |
| DREB1A, rd29A | Transcription factors | Bhatnagar-Mathur et al. [251] | |
| GmBIN2 | Enhance tolerance to drought | Wang et al. [252] | |
| GmCaM4 | Upregulate several drought-responsive genes | Yoo et al. [253] | |
| CDPK | Enhance water permeability across the membrane | Guenther et al. [254] | |
| GmHK, GmCLV1A, GmCLV1B, GmRLK1, GmRLK2, GmRLK3, GmRLK4 | Osmosensor | Yamamoto et al. [255] | |
| PgTIP1 | Confers drought tolerance | An et al. [256] | |
| GmDREB2 | Enhance drought tolerance | Savitri and Fauziah [257] | |
| GmRACK1 | Improve drought tolerance during vegetative growth | Li et al. [258] | |
| AtABF3 | Improve drought tolerance | Kim et al. [259] | |
| GmFDL19 | Enhance drought tolerance | Li et al. [260] | |
| GmSK1 | Enhance tolerance to drought | Chen et al. [261] | |
| Pigeon pea (Cajanus cajan) | CcM1522–CcM1821, CcM0047–CcM2332 | Improve drought tolerance | Saxena et al. [224] |
| MyB, WRKY, NF-Y | Transcription factors | Yang et al. [262] | |
| Cowpea (Vigna unguiculata) | CPRD8, CPRD12, CPRD14, CPRD22, CPRD46 phospholipase D, VuPLD1, 9-cisepoxycarotenoid dioxygenase, VuNCED1 | ABA biosynthesis | Muchero et al. [263] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khatun, M.; Sarkar, S.; Era, F.M.; Islam, A.K.M.M.; Anwar, M.P.; Fahad, S.; Datta, R.; Islam, A.K.M.A. Drought Stress in Grain Legumes: Effects, Tolerance Mechanisms and Management. Agronomy 2021, 11, 2374. https://doi.org/10.3390/agronomy11122374
Khatun M, Sarkar S, Era FM, Islam AKMM, Anwar MP, Fahad S, Datta R, Islam AKMA. Drought Stress in Grain Legumes: Effects, Tolerance Mechanisms and Management. Agronomy. 2021; 11(12):2374. https://doi.org/10.3390/agronomy11122374
Chicago/Turabian StyleKhatun, Marium, Sumi Sarkar, Farzana Mustafa Era, A. K. M. Mominul Islam, Md. Parvez Anwar, Shah Fahad, Rahul Datta, and A. K. M. Aminul Islam. 2021. "Drought Stress in Grain Legumes: Effects, Tolerance Mechanisms and Management" Agronomy 11, no. 12: 2374. https://doi.org/10.3390/agronomy11122374