
Effect of Nitrogen Supply on Growth and Nitrogen Utilization in Hemp (Cannabis sativa L.)


Abstract
:1. Introduction
2. Materials and Methods
- Total plant DW (DWP, g/plant) = root DW + stem DW + leaf DW;
- Ratio of root to shoot = root DW/(stem DW + leaf DW);
- Stem mass density (SMD, g/cm3) = stem DW/stem volume;
- NA of plant part (g/plant) = plant part DW × TN;
- NA per plant (NAP, g/plant) = root NA + stem NA + leaf NA;
- Nitrogen use efficiency (NUE, g/g) = DWP/NAP.
3. Results
3.1. Effect of N Supply on Plant Growth
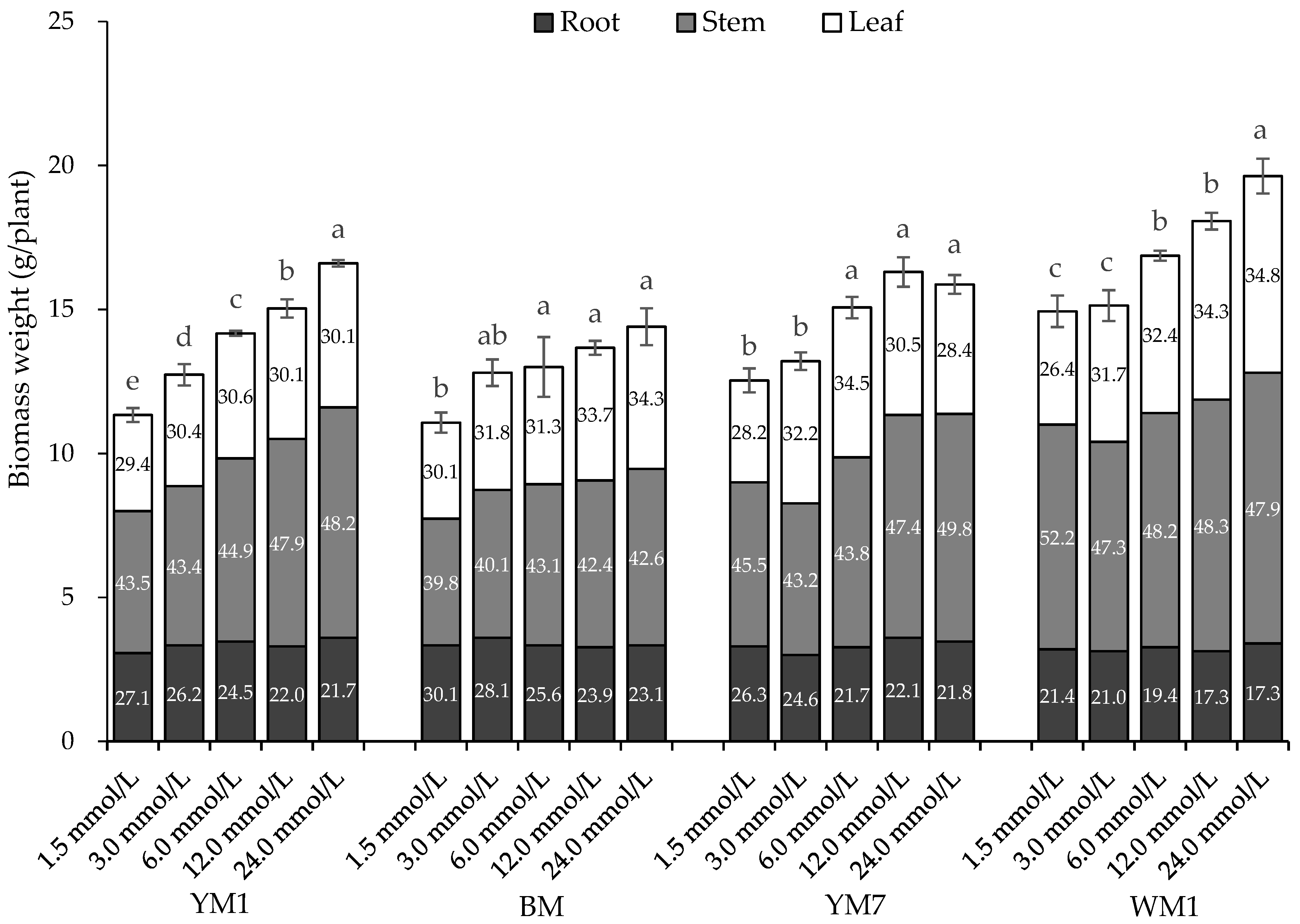
3.2. Effect of N Supply on Hemp Biomass
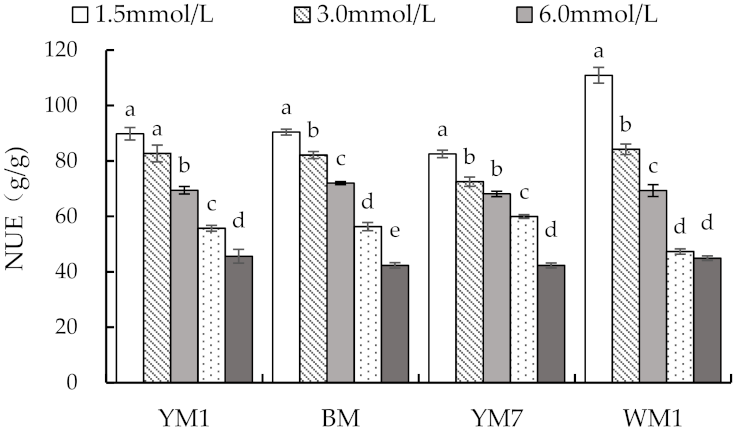
3.3. Effect of N Supply on NA and NUE in Hemp Plant
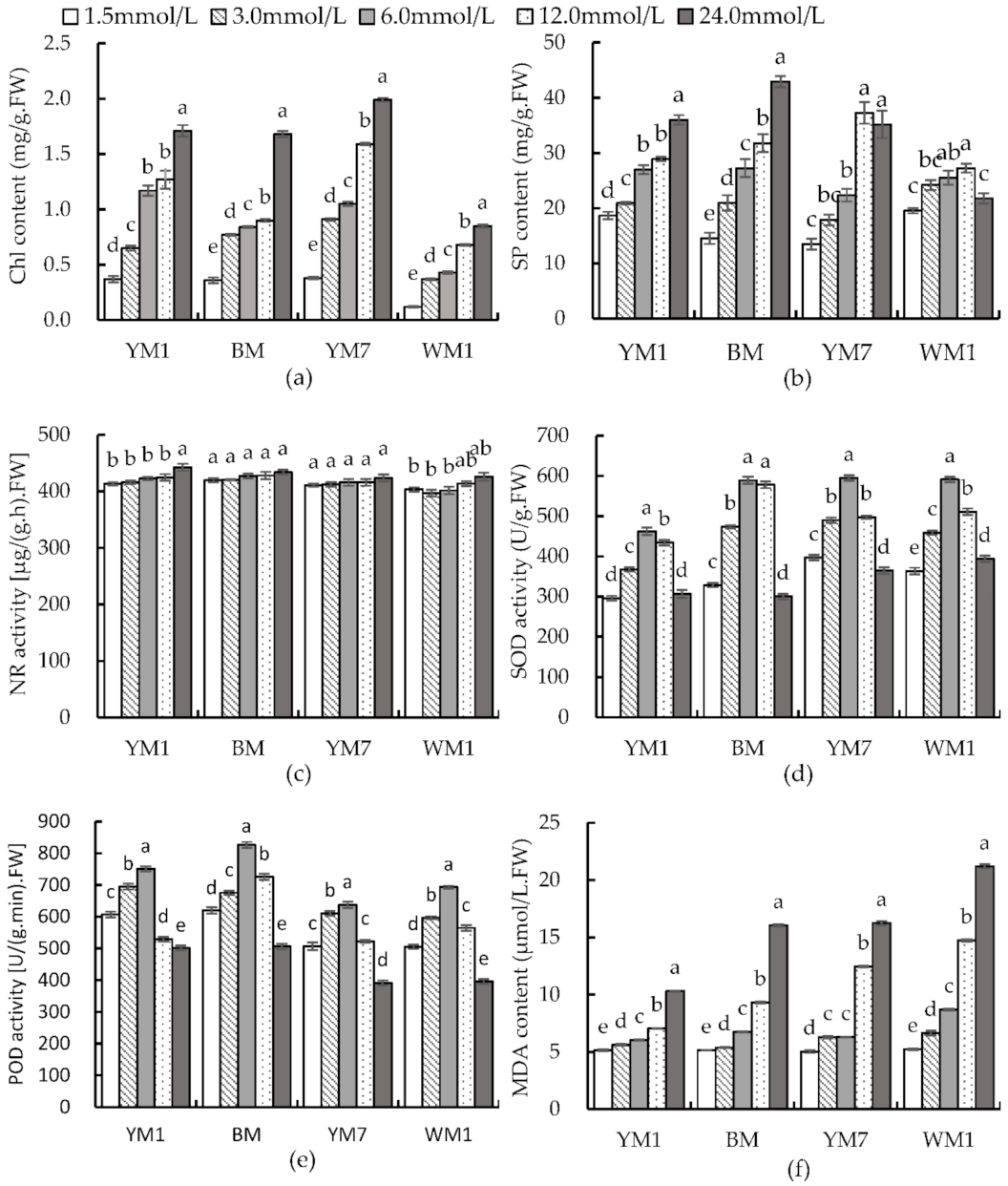
3.4. Effect of N Supply on Chl, SP, and NR in Hemp Leaf
3.5. Effect of N Supply on SOD, POD, and MDA in Hemp Leaf
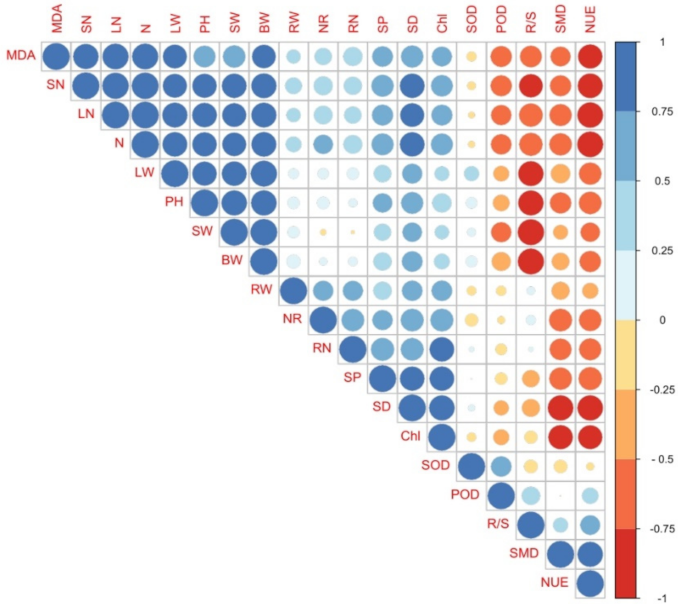
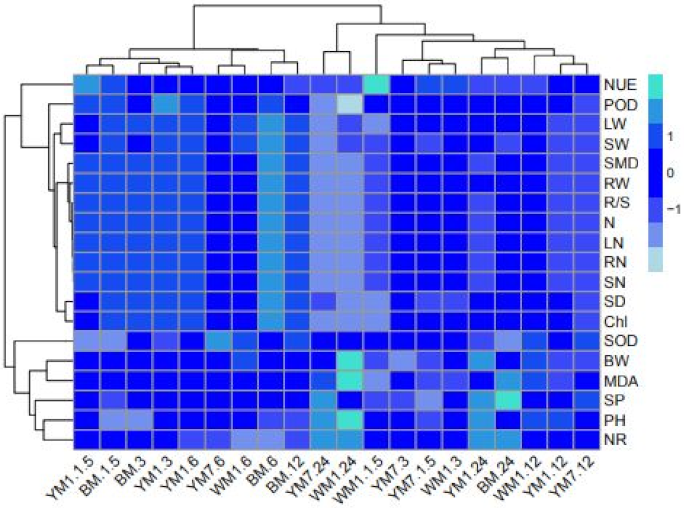
3.6. Relationships
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, Q.; Wu, K.; Fu, X.; Li, S.; Liu, X.; Gao, X. Sustainable crop yields from the coordinated modulation of plant growth and nitrogen metabolism. Chin. Sci. Bull. 2019, 64, 2633–2640. [Google Scholar] [CrossRef] [Green Version]
- Asif, I.; Dong, Q.; Wang, Z.; Wang, X.G.; Gui, H.P.; Zhang, H.H.; Pang, N.C.; Zhang, X.L.; Song, M.Z. Growth and nitrogen metabolism are associated with nitrogen-use efficiency in cotton genotypes. Plant Physiol. Biochem. 2020, 149, 61–74. [Google Scholar] [CrossRef]
- Ahmed, M.; Rauf, M.; Mukhtar, Z.; Saeed, N.A. Excessive use of nitrogenous fertilizers: An unawareness causing serious threats to environment and human health. Environ. Sci. Pollut. Res. 2017, 24, 26983–26987. [Google Scholar] [CrossRef]
- Harm, V.B.; Jake, M.S.; Atina, G.C.; Carling, M.T.; Andrew, G.S.; Timothy, R.H.; Jonathan, E.P. The Draft Genome and Transcriptome of Cannabis sativa. Genome Biol. 2011, 12, R102. Available online: http://genomebiology.com/2011/12/10/R102 (accessed on 12 November 2021).
- Farinon, B.; Molinari, R.; Costantini, L.; Merendino, N. The seed of industrial hemp (Cannabis sativa L.): Nutritional Quality and Potential Functionality for Human Health and Nutrition. Nutrients 2020, 12, 1935. [Google Scholar] [CrossRef]
- Ranalli, P.; Venturi, G. Hemp as a raw material for industrial applications. Euphytica 2004, 140, 1–6. [Google Scholar] [CrossRef]
- Salentijn, E.M.J.; Petit, J.; Trindade, L.M. The Complex Interactions Between Flowering Behavior and Fiber Quality in Hemp. Front. Plant Sci. 2019, 10, 614. [Google Scholar] [CrossRef]
- Liu, F.H.; Du, G.H.; Yang, Y.; Deng, G.; Tang, K.L. Green and Efficient Cultivation Techniques for Industrial Hemp of Flower Heads and Leaf Uses, 1st ed.; Yunnan University Press: Kunming, China, 2020; pp. 15–18. [Google Scholar]
- Deng, G.; Du, G.; Yang, Y.; Bao, Y.; Liu, F. Planting Density and Fertilization Evidently Influence the Fiber Yield of Hemp (Cannabis sativa L.). Agronomy 2019, 9, 368. [Google Scholar] [CrossRef] [Green Version]
- Kakabouki, I.; Kousta, A.; Folina, A.; Karydogianni, S.; Zisi, C.; Kouneli, V.; Papastylianou, P. Effect of Fertilization with Urea and Inhibitors on Growth, Yield and CBD Concentration of Hemp (Cannabis sativa L.). Sustainability 2021, 13, 2157. [Google Scholar] [CrossRef]
- Forrest, C.; Young, J.P. The Effects of Organic and Inorganic Nitrogen Fertilizer on the Morphology and Anatomy of Cannabis sativa “Fédrina” (Industrial Fibre Hemp) Grown in Northern British Columbia, Canada. J. Ind. Hemp 2006, 11, 3–24. [Google Scholar] [CrossRef]
- Papastylianou, P.; Kakabouki, I.; Travlos, I. Effect of Nitrogen Fertilization on Growth and Yield of Industrial Hemp (Cannabis sativa L.). Not. Bot. Hort. Agrobot. Cluj-Napoca 2017, 46, 197–201. [Google Scholar] [CrossRef] [Green Version]
- Aubin, M.; Seguin, P.; Vanasse, A.; Tremblay, G.F.; Mustafa, A.F.; Charron, J. Industrial Hemp Response to Nitrogen, Phosphorus, and Potassium Fertilization. CFTM 2015, 1, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Sausserde, R.; Adamovics, A. Effect of nitrogen fertilizer rates on industrial hemp (Cannabis sativa L.) biomass production. In Proceedings of the International Multidisciplinary Scientific Geo Conference, Varna, Bulgaria, 16–22 June 2013; Volume 1, pp. 339–346. [Google Scholar]
- Amaducci, S.; Errani, M.; Venturi, G. Response of hemp to plant population and nitrogen fertilization. Ital. J. Agron. 2002, 6, 103–111. [Google Scholar]
- Wylie, S.E.; Ristvey, A.G.; Fiorellino, N.M. Fertility management for industrial hemp production: Current knowledge and future research needs. GCB Bioenergy 2021, 13, 517–524. [Google Scholar] [CrossRef]
- Li, B.; Xin, W.; Sun, S.; Shen, Q.; Xu, G. Physiological and Molecular Responses of Nitrogen-starved Rice Plants to Re-supply of Different Nitrogen Sources. Plant Soil 2006, 287, 145–159. [Google Scholar] [CrossRef]
- Wang, X.K. Experimental Principle and Technology of Plant Physiology and Biochemistry, 3rd ed.; Higher Education Press: Beijing, China, 2015; pp. 50–105. [Google Scholar]
- Silveira, J.; Matos, J.; Cecatto, V.; Viegas, R.; Oliveira, J. Nitrate reductase activity, distribution, and response to nitrate in two contrasting Phaseolus species inoculated with Rhizobium spp. Environ. Exp. Bot. 2001, 46, 37–46. [Google Scholar] [CrossRef]
- Islam, M.S.; Peng, S.; Visperas, R.M.; Ereful, N.; Bhuiya, M.S.U.; Julfiquar, A. Lodging-related morphological traits of hybrid rice in a tropical irrigated ecosystem. Field Crop. Res. 2007, 101, 240–248. [Google Scholar] [CrossRef]
- Sperling, O.; Karunakaran, R.; Erel, R.; Yasuor, H.; Klipcan, L.; Yermiyahu, U. Excessive nitrogen impairs hydraulics, limits photosynthesis, and alters the metabolic composition of almond trees. Plant Physiol. Biochem. 2019, 143, 265–274. [Google Scholar] [CrossRef]
- Wu, X.R.; Zhang, W.J.; Wu, L.M.; Wang, F.; Li, G.H.; Liu, Z.H.; Tang, S.; Ding, C.G.; Wang, S.H.; Ding, Y.F. Characteristics of lodging resistance of super-Hybrid indica rice and its response to nitrogen. Sci. Agric. Sin. 2015, 48, 2705–2717. [Google Scholar] [CrossRef]
- Zhang, J.; Li, G.-H.; Song, Y.-P.; Zhang, W.-J.; Yang, C.-D.; Wang, S.-H.; Ding, Y.-F. Lodging Resistance of Super-Hybrid Rice Y Liangyou 2 in Two Ecological Regions. Acta Agron. Sin. 2013, 39, 682. [Google Scholar] [CrossRef]
- Song, M.D.; Li, Z.P.; Feng, H. Effects of irrigation and nitrogen regimes on dry matter dynamic accumulation and yield of winter wheat. Trans. Chin. Soc. Agric. Eng. 2016, 32, 119–126. [Google Scholar] [CrossRef]
- Grasso, R.; de Souza, R.; Peña-Fleitas, M.T.; Gallardo, M.; Thompson, R.B.; Padilla, F.M. Root and crop responses of sweet pepper (Capsicum annuum) to increasing N fertilization. Sci. Hortic. 2020, 273, 109645. [Google Scholar] [CrossRef]
- Garnett, T.; Conn, V.; Kaiser, B.N. Root based approaches to improving nitrogen use efficiency in plants. Plant Cell Environ. 2009, 32, 1272–1283. [Google Scholar] [CrossRef]
- Drew, M.C.; Saker, L.R.; Ashley, T.W. Nutrient Supply and the Growth of the Seminal Root System in Barley. J. Exp. Bot. 1973, 24, 1189–1202. [Google Scholar] [CrossRef]
- Anderson, E.L. Tillage and N fertilization effects on maize root growth and root:shoot ratio. Plant Soil 1988, 108, 245–251. [Google Scholar] [CrossRef]
- Ju, X.T.; Gu, B.J. Status-quo, problem and trend of nitrogen fertilization in China. J. Plant Nutr. Fertil. 2014, 20, 783–795. [Google Scholar] [CrossRef]
- Qu, L.B.; Gu, H.Y.; Liu, J.J.; Qing, G.J. Plant Biology, 1st ed; The Science Press: Beijing, China, 2012; pp. 303–320. [Google Scholar]
- Ren, B.; Dong, S.; Zhao, B.; Liu, P.; Zhang, J. Responses of Nitrogen Metabolism, Uptake and Translocation of Maize to Waterlogging at Different Growth Stages. Front. Plant Sci. 2017, 8, 1216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.P.; Dong, H.L.; Liu, A.Z.; Liu, J.R.; Li, R.Y.; Sun, M.; Li, Y.B.; Mao, S.C. Effects of nitrogen application rates on physiological characteristics of functional leaves, nitrogen use efficiency and yield of cotton. J. Plant Nutr. Fertil. 2015, 21, 81–91. [Google Scholar] [CrossRef]
- Jian, L.L.; Han, L.S.; Han, X.R.; Zhan, X.M.; Zuo, R.H.; Wu, Z.C.; Yuan, C. Effects of nitrogen on growth, root morphological traits, nitrogen uptake and utilization efficiency of maize seedlings. J. Plant Nutr. Fertil. 2011, 17, 247–253. [Google Scholar] [CrossRef]
- Iqbal, A.; Dong, Q.; Wang, X.; Gui, H.; Zhang, H.; Zhang, X.; Song, M. Variations in Nitrogen Metabolism are Closely Linked with Nitrogen Uptake and Utilization Efficiency in Cotton Genotypes under Various Nitrogen Supplies. Plants 2020, 9, 250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krapp, A.; Berthomé, R.; Orsel, M.; Mercey-Boutet, S.; Yu, A.; Castaings, L.; Elftieh, S.; Major, H.; Renou, J.-P.; Daniel-Vedele, F. Arabidopsis Roots and Shoots Show Distinct Temporal Adaptation Patterns toward Nitrogen Starvation. Plant Physiol. 2011, 157, 1255–1282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, B.-M.; Wang, Z.-H.; Li, S.-X.; Wang, G.-X.; Song, H.-X.; Wang, X.-N. Effects of nitrate supply on plant growth, nitrate accumulation, metabolic nitrate concentration and nitrate reductase activity in three leafy vegetables. Plant Sci. 2004, 167, 635–643. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Bowler, C.; Montagu, M.V.; Inze, D. Superoxide Dismutase and Stress Tolerance. Annu. Rev. Plant Biol. 1992, 43, 83–116. [Google Scholar] [CrossRef]
- Willekens, H.; Chamnongpol, S.; Davey, M.; Schraudner, M.; Langebartels, C.; Van Montagu, M.; Inzé, D.; Van Camp, W. Catalase is a sink for H2O2 and is indispensable for stress defence in C3 plants. EMBO J. 1997, 16, 4806–4816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, L.; Xie, Y.; Hu, L.; Si, J.; Wang, Z. Excessive nitrogen application dampens antioxidant capacity and grain filling in wheat as revealed by metabolic and physiological analyses. Sci. Rep. 2017, 7, 43363. [Google Scholar] [CrossRef]
- Mitsuhara, I.; Malik, K.A.; Miura, M.; Ohashi, Y. Animal cell-death suppressors Bcl-xL and Ced-9 inhibit cell death in tobacco plants. Curr. Biol. 1999, 9, 775–778. [Google Scholar] [CrossRef] [Green Version]
- Ishida, H.; Anzawa, D.; Kokubun, N.; Makino, A.; Mae, T. Direct evidence for non-enzymatic fragmentation of chloroplastic glutamine synthetase by a reactive oxygen species. Plant Cell Environ. 2002, 25, 625–631. [Google Scholar] [CrossRef]
- Domínguez-Valdivia, M.D.; Aparicio-Tejo, P.M.; Lamsfus, C.; Cruz, C.; Martins-Loução, M.A.; Moran, J.F. Nitrogen nutrition and antioxidant metabolism in ammonium-tolerant and -sensitive plants. Physiol. Plant. 2008, 132, 359–369. [Google Scholar] [CrossRef]
- Liu, H.W.; Zhang, R.R.; Liu, Y.L.; Xie, C.H.; Lin, H.; Lin, Z.X.; Ye, W.H. Effect of Nitrogen on Pennisetum sp. Seeding Stage Growth and Photosynthetic Physiological Characteristic. Northern Hortic. 2016, pp. 133–138. Available online: http://bfyy.paperonce.org/oa/DArticle.aspx?type=view&id=201608037 (accessed on 12 November 2021).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Nitrogen (mmol/L) | Plant Height (cm) | Stem Diameter (mm) | Root/Shoot Ratio | Stem Mass Density (g/cm3) |
|---|---|---|---|---|---|
| YM1 | 1.5 | 62 ± 8 c | 3.3 ± 0.2 c | 0.37 ± 0.03 a | 0.90 ± 0.09 a |
| 3.0 | 75 ± 5 b | 3.5 ± 0.3 c | 0.36 ± 0.03 ab | 0.78 ± 0.09 b | |
| 6.0 | 89 ± 2 a | 4.6 ± 0.4 b | 0.33 ± 0.02 b | 0.40 ± 0.01 c | |
| 12.0 | 94 ± 5 a | 5.0 ± 0.5 b | 0.28 ± 0.01 c | 0.36 ± 0.02 cd | |
| 24.0 | 95 ± 1 a | 6.1 ± 0.5 a | 0.28 ± 0.02 c | 0.29 ± 0.01 d | |
| BM | 1.5 | 45 ± 2 d | 3.9 ± 0.2 b | 0.43 ± 0.03 a | 0.82 ± 0.04 a |
| 3.0 | 54 ± 2 c | 4.2 ± 0.5 b | 0.39 ± 0.06 ab | 0.73 ± 0.23 b | |
| 6.0 | 68 ± 1 b | 4.7 ± 0.5 ab | 0.35 ± 0.07 abc | 0.50 ± 0.21 b | |
| 12.0 | 70 ± 4 ab | 4.7 ± 0.5 ab | 0.31 ± 0.02 bc | 0.49 ± 0.16 b | |
| 24.0 | 72 ± 1 a | 5.2 ± 0.7 a | 0.30 ± 0.06 c | 0.43 ± 0.18 b | |
| YM7 | 1.5 | 58 ± 4 d | 3.1 ± 0.4 d | 0.36 ± 0.02 a | 1.14 ± 0.09 a |
| 3.0 | 71 ± 7 c | 4.0 ± 0.5 c | 0.32 ± 0.02 b | 0.61 ± 0.05 b | |
| 6.0 | 81 ± 3 b | 4.4 ± 0.4 c | 0.29 ± 0.01 c | 0.51 ± 0.04 c | |
| 12.0 | 87 ± 4 ab | 5.2 ± 0.4 b | 0.28 ± 0.01 c | 0.40 ± 0.01 d | |
| 24.0 | 92 ± 1 a | 6.1 ± 0.4 a | 0.28 ± 0.01 c | 0.30 ± 0.03 e | |
| WM1 | 1.5 | 69 ± 6 c | 3.2 ± 0.2 d | 0.27 ± 0.02 a | 1.42 ± 0.04 a |
| 3.0 | 76 ± 2 c | 3.5 ± 0.2 c | 0.26 ± 0.01 ab | 0.96 ± 0.17 b | |
| 6.0 | 90 ± 4 b | 4.6 ± 0.2 b | 0.24 ± 0.02 b | 0.55 ± 0.05 c | |
| 12.0 | 98 ± 6 ab | 4.8 ± 0.3 ab | 0.21 ± 0.02 c | 0.50 ± 0.06 c | |
| 24.0 | 104 ± 4 a | 5.0 ± 0.1 a | 0.21 ± 0.01 c | 0.47 ± 0.02 c | |
| YM1 | 1.5 | 62 ± 8 c | 3.3 ± 0.2 c | 0.37 ± 0.03 a | 0.90 ± 0.09 a |
| 3.0 | 75 ± 5 b | 3.5 ± 0.3 c | 0.36 ± 0.03 ab | 0.78 ± 0.09 b | |
| 6.0 | 89 ± 2 a | 4.6 ± 0.4 b | 0.33 ± 0.02 b | 0.40 ± 0.01 c | |
| 12.0 | 94 ± 5 a | 5.0 ± 0.5 b | 0.28 ± 0.01 c | 0.36 ± 0.02 cd | |
| 24.0 | 95 ± 1 a | 6.1 ± 0.5 a | 0.28 ± 0.02 c | 0.29 ± 0.01 d |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.; Zha, W.; Tang, K.; Deng, G.; Du, G.; Liu, F. Effect of Nitrogen Supply on Growth and Nitrogen Utilization in Hemp (Cannabis sativa L.). Agronomy 2021, 11, 2310. https://doi.org/10.3390/agronomy11112310
Yang Y, Zha W, Tang K, Deng G, Du G, Liu F. Effect of Nitrogen Supply on Growth and Nitrogen Utilization in Hemp (Cannabis sativa L.). Agronomy. 2021; 11(11):2310. https://doi.org/10.3390/agronomy11112310
Chicago/Turabian StyleYang, Yang, Wenxin Zha, Kailei Tang, Gang Deng, Guanghui Du, and Feihu Liu. 2021. "Effect of Nitrogen Supply on Growth and Nitrogen Utilization in Hemp (Cannabis sativa L.)" Agronomy 11, no. 11: 2310. https://doi.org/10.3390/agronomy11112310