
Role of Rhizosphere Soil Microbes in Adapting Ramie (Boehmeria nivea L.) Plants to Poor Soil Conditions through N-Fixing and P-Solubilization

1
College of Agronomy, Hunan Agricultural University, Changsha 410128, China
2
Orient Science & Technology College of Hunan Agricultural University, Changsha 410128, China
*
Author to whom correspondence should be addressed.
Agronomy 2021, 11(11), 2096; https://doi.org/10.3390/agronomy11112096
Submission received: 30 September 2021
/
Revised: 16 October 2021
/
Accepted: 17 October 2021
/
Published: 20 October 2021
(This article belongs to the Special Issue Social-Ecologically More Sustainable Agricultural Production)
Abstract
:The N-fixing and P-solubilization functions of soil microbes play a vital role in plant adaptation to nutrient-deficiency conditions. However, their exact roles toward the adaptation of ramie to poor soil conditions are still not clear. To fill this research gap, the N-fixing and P-solubilization efficiencies of soils derived from the rhizosphere of several ramie genotypes with different levels of poor soil tolerance were compared. Correlations between the N-fixing, P-solubilization efficiency, and the poor soil tolerable index were analyzed to quantify their contributions towards the adaptation of ramie plants to poor soil conditions. To explore how the microorganisms affected the potential of N-fixing/P-solubilization, the activities of the nutrients related the soil enzymes were also tested and compared. The results of this study confirm the existence of N-fixing and P-solubilization bacteria in the ramie rhizosphere of the soil. The number of N-fixing bacteria varied from 3010.00 to 46,150.00 c.f.u. per gram dry soil for the ramie treatment, while it was only 110.00 c.f.u. per gram dry soil for treatment without ramie cultivation. The average P-solubilization efficiency of ramie treatment was almost five times higher than that of the control soil (0.65 vs. 0.13 mg mL−1). The significant correlations between the poor soil tolerance index and the N-fixing bacteria number (r = 0.829)/nitrogenase activity (r = 0.899) suggest the significantly positive role of N-fixing function in the adaptation of ramie plants to poor soil. This is also true for P-solubilization, as indicated by the significant positively correlation coefficients between the ramie poor soil tolerance index and P-solubilization efficiency (0.919)/acid phosphatase activity (0.846). These characteristics would accelerate the application of “holobiont” breeding for improving ramie nutrient use efficiency.
1. Introduction
Ramie (Boehmeria nivea L.) is a perennial fiber yielding crop, which generally requires high inputs to achieve potential yield and maintain a good fiber quality [1]. The nutrient requirements for ramie production are generally two to four times higher than that of the normal field crops. For example, the recommended demands of N, P2O5, and K2O for the production of 100 kg ramie fiber are 8.00 kg, 2.00 kg, and 9.00 kg [2], respectively, while they are only 2.56 kg, 0.77 kg, and 2.53 kg for the production of 100 kg corn grains [3]. Actually, during the farmer’s production process, higher amounts of fertilizer are usually applied as most Chinese farmers believe the higher the fertilizer application, the higher the crop yield. Over-fertilization exists in almost all the agricultural production fields in China, which results in the crops’ nutrient uptake efficiencies being generally lower than 30% or even 20% [4]. This over-fertilization has caused a series of problems such as surface and groundwater pollution, as well as increasing greenhouse gas emissions. To address these problems, the Chinese government has implemented a strategy of “agricultural transformation and upgrading”. One important task of the strategy is to establish a sustainable agricultural production system with special emphasis on low input cropping systems.
When implementing such a system in ramie production, the key aspect is to minimize the use of production inputs, especially use of synthetic fertilizers [5]. One way to reduce fertilizer input is through optimization of the cultivation techniques [2]. However, it is still advised to breed new ramie varieties that are characterized by high NUE (nutrient utilization efficiency). Traditionally, plant breeding was conducted by altering the plant’s own genomic information. Recently, a new perspective of “holobiont” breeding has emerged and been accepted [6], which claims “plant breeding goes microbial” [7]. The holobiont considerers the plant and its associated microbiome as an evolutionary unit, which together transmit the genetic information to next generation [8]. Therefore, future breeding should set the selection targets not only in terms of plant materials, but also for the associated microbes.
Soil microorganisms play a vital role in nutrient recycling [9]. During our previous research, the helpful roles of soil microbes in the adaptation of the ramie to poor soil conditions were confirmed [10]. This is generally achieved through enrichment of the beneficial bacteria and through a reduction of harmful fungi simultaneously. The current results indicate that ramie’s NUE can be improved through holobiont breeding. However, the premise for achieving this potential lies in understanding how the microorganisms improve the ramie’s NUE.
Therefore, the current study is designed with an overall aim to explore the role of soil microbes for improving the NUE of the ramie plant. The improvement in NUE by microorganisms is mainly evaluated through their potential for increasing the amount of nutrients available for plant uptake and subsequent utilization [9]. For example, in legume species, the high NUE is attributed to the N-fixing characteristic, which is also true for some non-legume species such as miscanthus [11,12]. Phosphate solubilizing microorganisms (PSM) can convert immobilized inorganic P to soluble organic P (i.e., P-solubilization), which increases the bioavailable P for plant uptake and utilization [13]. Our previous work already detected the N-fixing bacteria of Bradyrhizobium from the rhizosphere soil of ramie using sequencing technology [10]. However, the exact N-fixing potential is not clear. Therefore, the first objective (Objective I) of this study is to quantitatively assess the N-fixing potential of the ramie rhizosphere soil and to evaluate the contribution of the N-fixing characteristic in the poor soil tolerance of the ramie plant. Next to nitrogen, phosphorus is the second most important element for plant growth. Thus, the second objective (Objective II) of this study is to explore the role of P-solubilization, which is similar to N-fixing, and its potential contribution towards adaptation of the ramie plant to poor soil conditions. Another key aspect is the role of soil enzymes, which act as a bridge between the microorganism and N-fixing/P-solubilization [14]. To explore how the microorganisms affect the potential of N-fixing/P-solubilization (Objective III), the activities of the nutrient-related soil enzymes are also monitored and compared as a part of this study. Furthermore, based on the outcomes of the former research work [10], it is revealed that the harmful fungal communities (e.g., Cladosporium) can be enriched in the ramie rhizosphere soil, which can kill the beneficial bacteria and limit the N-fixing potential. Therefore, the fourth objective (Objective IV) is to verify the inhibitory effect of fungal communities on N-fixing bacterial communities in the rhizosphere.
2. Materials and Methods
The N-fixing and P-solubilization potential of soils derived from the rhizosphere of several ramie genotypes have different tolerance levels to poor soil conditions. The tolerance level is expressed by the overall plant field performance under poor soil conditions, as a higher field performance will have a stronger poor soil tolerance ability. The overall field performance was normalized (detail calculation process shown in [10]) and then expressed to a field performance index (NFPI). The N-fixing potential was quantified by the number of nitrogen-fixing bacteria in the soil and the related nitrogenase activity. The P-solubilization quantity was measured as the available phosphorus (AP) content in the TCP (tricalcium phosphate) liquid medium after microbial cultivation. Other than nitrogenase, the activity of acid phosphatase (S-ACP), urease (S-UE), and sucrase (S-SC) were also tested and compared in this study. Their contributions for helping the ramie plant adapt to poor soil conditions were evaluated through the correlation analysis between the soil enzyme activity and the corresponding NFPI value. Quantification of the anti-N-fixing-bacteria activity by fungal communities was conducted using the inhibition-zone assay.
2.1. Materials and Sampling Strategies
All of the above-described tests were conducted using the fresh soil samples collected in December 2020. Soil samples of four ramie genotypes and one blank control CK (i.e., without ramie cultivation) were compared. The four ramie genotypes, namely Xiangzhu XB (XZ-XB), Zhongzhu 1 (ZZ-1), Xiangzhu X2 (XZ-X2), and Xiangzhu 3 (XZ-3), were established in 2010 at the experimental station of Huarong (29°32′46″ N, 112°39′57″ E, 73 m a.s.l.). The CK was set as the weed free bare ground, which was adjacent to the ramie cultivation block. Without ramie cultivation, the CK block was dominated by the species of Chrysanthemum indicum L. and Humulus scandens (Lour.) Merr. The field soil is a poor sandy red soil that was found to have a total nitrogen content of 0.69 g/kg, available phosphorus content of 9.62 mg/kg, and exchangeable potassium content of 56.53 mg/kg. According to the field evaluation results [10], the poor soil tolerance ability of the four tested genotypes were shown as XZ-XB (NFPI = 0.953) > ZZ-1(NFPI = 0.701) > XZ-X2(NFPI = 0.452) > XZ-3 (NFPI = 0.000). More detailed information about the experiment design and the field performance of the genotypes can be found in our previously published work [10]. The rhizosphere soil samples of the ramie plants were collected in the field and transported to the laboratory in an ice box. The rhizosphere soil samples, defined as the soil remaining attached to the roots after shaking plants vigorously [15], were collected according to the “air shaking method”. The rhizosphere soil samples of five plants within one block were randomly selected in an “S” pattern and then mixed to one composite rhizosphere sample. Five soil cores (0–20-cm depth) within the CK block were collected according to the “S” pattern and were mixed to be the composite CK sample. All the samples were sieved through 50-mesh sieves and stored at 4 °C until the start of the test (less than 30 days here).
2.2. Quantification of the Number of Soil N-Fixing Bacteria
The number of the N-fixing bacteria was quantified using the spread plate method, with a detailed procedure comprised of following steps:
(a) Preparation of the soil suspension by adding 10.00 g of the fresh soil sample to 90 mL sterile distilled water and then shaking at 30 °C at 150 rpm for 30 min. (b) Preparation of the diluted soil suspension by 10-, 102-, 103-, 104-, and 105-fold dilutions of the suspension, which was prepared by adding 1 mL previous-level-fold suspension to 9 mL of sterile distilled water. (c) Determination of the N-fixing bacteria number of 103-, 104-, and 105-fold diluted suspensions were plated onto Petri plates containing Ashby nitrogen-free solid medium. This was followed by 3 d incubation at 30 °C and the number of N-fixing bacteria in each plate was counted. Afterwards, the counted number was converted to the comparable number, expressed as log c.f.u. per gram of dry soil.
The above steps of each soil sample were repeated four times. The composition of the Ashby nitrogen-free solid medium included 10.00 g mannitol, 0.20 g KH2PO4, 0.20 g MgSO4, 0.20 g NaCl, 0.30 g K2SO4, 5.0 g CaCO3, 1000.00 mL distilled water, and 18.00 g agar. At first, these components were mixed and adjusted to a pH of 7.0, followed by sterilization at 121 °C for 30 min.
2.3. Quantitative Assay to Determine the P Solubilization Efficiency of Rhizosphere Microbes
In this assay, each 250 mL Erlenmeyer flasks containing 90.00 mL TCP liquid medium was inoculated with 10.00 mL soil suspension (see Step A in Section 2.2) and then shook at 30 °C at 150 rpm for 72 h. To eliminate the background effect, a control treatment was carried out simultaneously without the soil suspension and adding only the corresponding sterilized (at 121 °C for 30 min) suspension. At the end of incubation time, 5 mL cultures were sampled and centrifuged at 12,000 rpm for 5 min. Afterwards, the supernatants were used to determine the AP contents by the molybdenum blue method. The net P-solubilization quantity of each sample was the difference of AP content between the normal treatment and control treatment. Each soil sample had four replications. The composition of the TCP liquid medium included 0.30 g NaCl, 0.30 g MgSO4·7H2O, 0.50 g (NH4)2SO4, 0.30 g KCl, 0.03 g FeSO4·7H2O, 0.03 g MnSO4·4H2O, 5.00 g Ca3(PO4)2, 10.00 g glucose, and 1000.00 mL distilled water. In addition, prior to microbial cultivation, the pH of the aforementioned medium was adjusted to 7.0 and sterilized at 121 °C for 30 min.
2.4. Quantitative Estimation of the Soil Enzyme Activity
The nitrogenase activity was measured using an acetylene reduction assay (ARA) and was expressed by the conversion efficiency of acetylene (C2H2) to ethylene (C2H4). The detailed description of ARA method is as follows: (1) adding 0.50 mL soil suspension to each serum bottle (100 mL) containing 50.00 mL nitrogen-free Ashby liquid medium; (2) sealing the bottle by cotton plugs and incubation at 30 °C and 150 rpm for 72 h; (3) replacing cotton plugs with air tight serum stopper; (4) removing 5.00 mL atmospheric air from the tube and injecting same volume of acetylene; and (5) afterwards, continue with incubation for another 72 h and draw 1 mL gas sample from the tube to measure the C2H4 concentration using gas chromatography. Based on this measured concentration, the amount of C2H4 produced per gram of dry soil per 24 h is the nitrogenase activity, which was calculated according to the method described in the study of Haskett [16].
The activities of S-ACP, S-UE, and S-SC were determined using the Solarbio detection kits (Solarbio Technology Co., Ltd., Beijing, China) of BC0140, BC0120, and BC0240, respectively, as per the manufacturer’s instructions. The S-UE activity was expressed as the amount of NH3-N, the S-ACP activity as the amount of phenol, and the S-SC activity as the amount of reducing sugar produced per gram of dry soil after 24 h at 37 °C.
2.5. Antagonistic Test to Quantify the Effect of Fungal Communities on N-Fixing Bacteria
Quantification of the anti-N-fixing-bacteria activity by fungal communities was conducted using an inhibition-zone assay. In this assay, the dot culture of the fungal colony on the N-fixing bacteria grown in Petri plates was carried out. The inhibition efficiency was calculated as halo zone diameter/colony diameter (HD/CD). The procedure comprised the following: (a) enrichment of the N-fixing bacteria by adding 5.00 mL soil suspension to 95.00 mL sterilized liquid N-fixing bacteria enrichment medium (15.00 g glucose, 0.80 g KH2PO4, 0.20 g MgSO4, 0.20 g NaCl, 1.00 g CaCO3, 1.00 mL Na2MoO4 (mass fraction of 1%), 1.00 mL H3BO3 (mass fraction of 1%), 1.00 mL MnSO4 (mass fraction of 1%), 1.00 mL FeSO4·7H2O (mass fraction of 1%) and 1000.00 mL distilled water) and then incubated at 30 °C and 150 rpm for 3 days, then taking 5.00 mL of the bacterial culture and enriching again by following the same procedure. (b) Preparation of the N-fixing bacteria to be cultured in petri plates by plating 1.00 mL dule-enriched N-fixing bacterial culture to the surface of petri plates containing a solid LB (Luria-Bertani) medium followed by incubation at 30 °C for 3 days. (c) Preparation of the fungal colony by plating 1 mL ten-fold diluted (10−1) soil suspension on the surface of Petri plates containing a solid PAD medium, followed by incubation at 28 °C until the whole surface was covered by mycelium. Afterwards, a fungal colony was cut using a 5 mm cork borer. (d) Co-incubation of the fungal colony and N-fixing bacteria in Petri plates by spotting three fungal colonies and one control colony (prepared according to the same method for fungal colony with replacing 1 mL soil suspension to 1 mL distilled water) on the surface of N-fixing bacteria grown at four equidistant points near the Petri center in four directions, followed by co-incubation of the paired plates at 30 °C for 3 days. (e) At the end of incubation, the diameter of the halo and the colony were measured.
All of the above steps for each sample were carried out in four replications.
2.6. Statistical Analysis
Data are presented as mean ± SD. Differences in terms of the tested parameters as described above among the different ramie genotypes were analyzed using one-way ANOVA (analysis of variance) in SAS 9.4 software (SAS Institute, Cary, NC, USA). The mean values of these parameters were compared using Duncan’s multiple range tests at both a p < 0.05 and p < 0.01 level. If 0.01 < p < 0.05, the significance was marked by p < 0.05, otherwise by p < 0.01. A correlation analysis was performed between the activities of the four tested soil enzymes and the poor soil tolerance ability of ramie plants (expressed by the NFPI).
3. Results
3.1. Comparison of the N-Fixing and P Solubilization Efficiencies between Different Ramie Germplasms
The results shown in Figure 1 indicate that the N-fixing and P-solubilization efficiency of the ramie rhizosphere soil was significantly stronger (p < 0.05) than that in the blank control CK soil without ramie. The number of N-fixing bacteria in the ramie rhizosphere soil, which was pooled of 30,822.00 c.f.u. per gram dry soil for all the four genotypes, was 280-times higher than that of the CK (110.00 c.f.u. per gram dry soil) without ramie cultivation (Figure 1a). This was also true for the P-solubilization efficiency, as the AP content (Figure 1b) in the incubation culture of the ramie rhizosphere (four genotypes pooled) was almost five-times higher than that of the CK soil (0.65 vs. 0.13 mg mL−1). To compare the tested genotypes, there is a general trend that genotypes with a better adaptability to poor soil conditions are characterized by a high N-fixing and P-solubilization efficiency. For example, XZ-XB, as the most tolerable genotype, had the second highest N-fixing bacteria number (46,150.00 c.f.u. per gram dry soil), which was just slightly lower than that (53,060.00 c.f.u. per gram dry soil) of XZ-X2 (p > 0.01), but significantly (p < 0.01) higher than that of ZZ-1 and XZ-3. In particular, XZ-XB showed 15.3-times more N-fixing bacteria (46,150.00 vs. 3010.00 c.f.u.) compared with the poorest tolerable genotype (XZ-3). The correlation results show that the N-fixing bacteria number was significantly positively correlated with the poor soil tolerance index NFPI (r = 0.829, p < 0.01). In terms of differences in the P-solubilization efficiency, the genotypes from a high to low P solubilization ability were XZ-XB (AP content of 0.85 mg mL−1), ZZ-1 (0.85 mg mL−1), XZ-3 (0.47 mg mL−1), and XZ-X2 (0.41 mg mL−1). Although the order of XZ-3 and XZ-X2 for P solubilization ability was against that of NFPI, the difference in the AP content between XZ-3 and XZ-X2 did not reach a significant level (p > 0.01). The positive correlation (r = 0.919) between the P-solubilization efficiency and NFPI was also significant (p < 0.01).
3.2. Effects of Fungal Communities on N-Fixing Bacteria
The results of this study confirm that the ramie rhizosphere soil derived fungal communities have an inhibitory effect on N-fixing bacteria. The halo zone was observed for all of the ramie treatments except for the CK treatment (Figure 1c). This inhibitory effect weakened the ramie poor soil tolerance ability, as indicated by the significantly negative relationship (r = −0.995, p < 0.01) between HD/CD and NFPI. More precisely, the highest HD/CD ratio of 3.42 was observed in the rhizosphere soil of XZ-3 (the worst performing genotypes under poor soil condition), followed by XZ-X2 (2.39), ZZ-1 (2.00), and XZ-XB (1.72). This trend was opposite to the order of the poor soil tolerance ability. Additionally, the halo zone boundary of the XZ-XB treatment was not fixed and some sporadic N-fixing bacteria colonies were observed within the halo zone. This also indicates that the fungal communities from XZ-XB rhizosphere soil had a relatively weak inhibitory effect on the growth of N-fixing bacteria. In contrast, XZ-3 treatment had a large halo zone and obvious boundary, suggesting a strong inhibitory effect. However, the poor ramie soil tolerance was not fully attributed to the inhibitory effect of the fungal communities on the N-fixing bacteria. For example, the genotype of XZ-X2 had the second-highest HD/CD, but also the highest number for the N-fixing bacteria. Furthermore, the number of N-fixing bacteria in XZ-X2 was 2.5-times higher than that of ZZ-1, despite no significant differences in terms of the HD/CD ratio between these two genotypes.
3.3. Comparison of the Soil Enzyme Activity between Different Ramie Germplasms
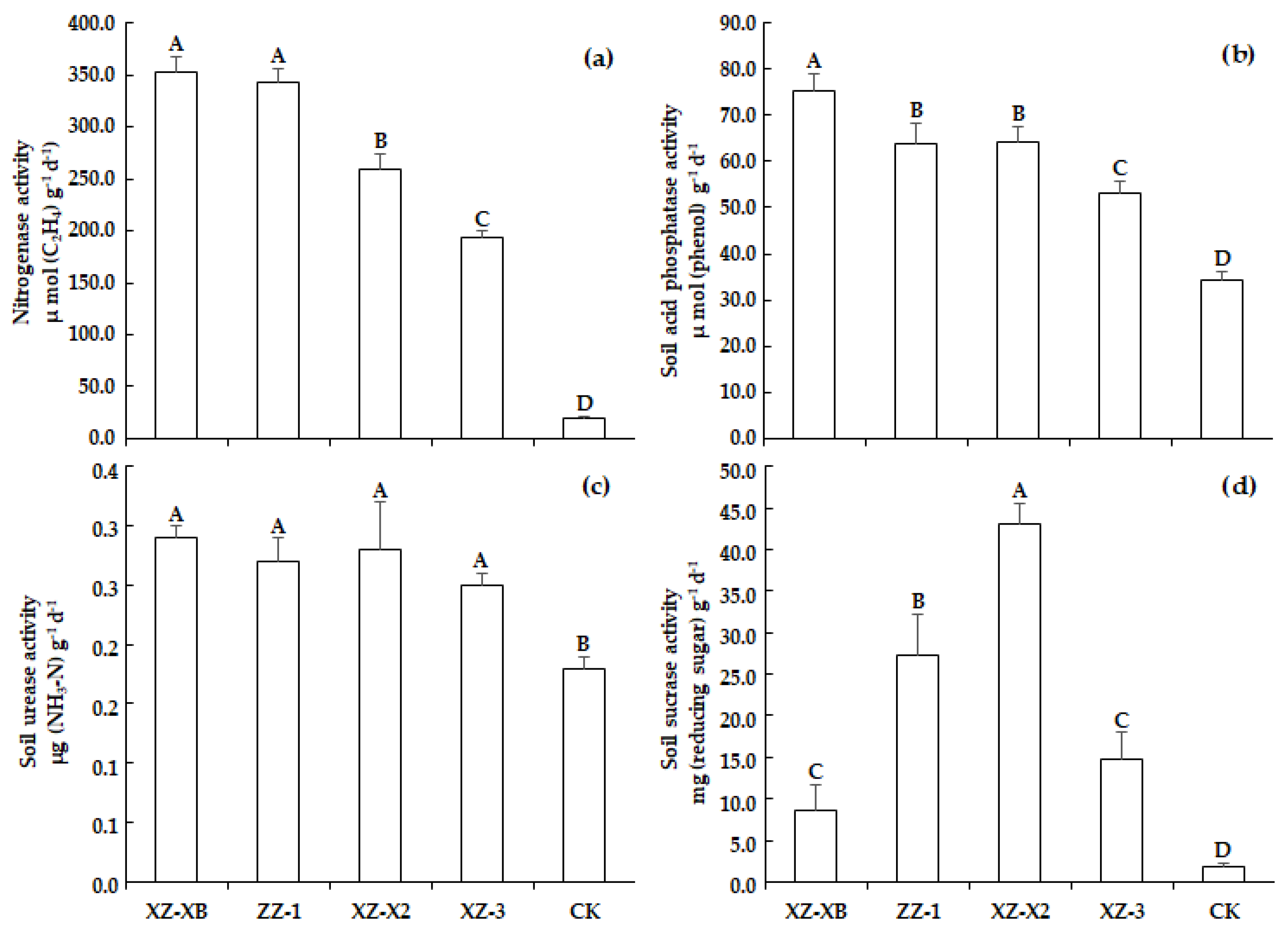
The results presented in Figure 2 compare the differences in terms of the activities of nitrogenase, S-ACP, S-UE, and S-SC among the soils cultivated with different ramie genotypes and the control without ramie cultivation. The activities of the tested enzymes in the ramie cultivation treatments were all higher than that of the CK treatment. In particular, the difference in terms of nitrogenase activity was the most significant, as the average activity in ramie cultivation treatment (287.60 μ mol (C2H4) g−1 d−1) was 15-times higher than that of the CK (19.13 μ mol (C2H4) g−1 d−1), whereas the S-SC activity (23.39 mg (reducing sugar) g−1 d−1) was 13.4-times higher than the CK treatment (1.75 mg (reducing sugar) g−1 d−1). The S-ACP activity only had a difference of 1.86-times (64.19 vs. 34.41 μ mol (phenol) g−1 d−1). For the activity of S-UE, the ramie treatment (four genotypes pooled) was only 34.3% higher (0.27 vs. 0.18 μg (NH3-N) g−1 d−1) than that of CK.
To compare the ramie genotypes, significant (p < 0.01) differences were observed in terms of nitrogenase, S-ACP, and S-SC activities, but not in case of the S-UE activity (p > 0.05). The nitrogenase activity showed a completely consistent trend with the poor soil tolerance ability of the ramie plant. The nitrogenase activity (Figure 2a) in the XZ-XB treatment (353.30 μ mol (C2H4) g−1 d−1) was significantly (p < 0.05) higher than that of XZ-X2 treatment (259.33 μ mol (C2H4) g−1 d−1) and the XZ-3 treatment (194.10 μ mol (C2H4) g−1 d−1), whereas the difference was not significant for the ZZ-1 treatment (343.47 μ mol (C2H4) g−1 d−1). The differences in S-ACP activity (Figure 2b) among the genotypes were generally consistent with the ramie’s poor soil tolerance ability. The best performing XZ-XB treatment had the highest S-ACP activity of 75.45 μ mol (phenol) g−1 d−1, whereas the lowest activity of 53.28 μ mol (phenol) g−1 d−1 was recorded in the worst performing XZ-3. In the better performing ZZ-1 treatment, the S-ACP activity was lower than the XZ-X2 treatment (63.72 vs. 64.30 μ mol (phenol) g−1 d−1), but these differences were not significant. The S-UE activities (Figure 2c) of the four genotypes ranged from 0.25 to 0.29 μg (NH3-N) g−1 d−1, which were not significantly different. For the S-SC activity (Figure 2d), the XZ-X2 treatment had the highest activity of 43.02 mg (reducing sugar) g−1 d−1, which was 36.50% higher (p < 0.01) than that of the second-highest treatment of ZZ-1 (27.34 mg (reducing sugar) g−1 d−1). However, the best-performing XZ-XB had the lowest S-SC activity of only 8.51 mg (reducing sugar) g−1 d−1.
The contribution of the soil enzyme to improve the ability of ramie to tolerate poor soil conditions was expressed by the relationship between the soil enzyme activity and the poor soil tolerance index NFPI. The results (Table 1) show that the adaptability of ramie to poor soil was significantly and positively correlated with the nitrogenase activity, S-ACP activity, and S-UE activity, but not with the S-SC activity (r = 0.256, p = 0.421). According to the Pearson correlation coefficient, the adaptability of the ramie plant to poor soil was mostly correlated with the nitrogenase activity, as indicated by the highest correlation coefficient of 0.899 (p < 0.001), followed by the S-ACP activity (r = 0.846, p = 0.005) and then the S-UE activity (r = 0.698, p = 0.012). Besides, a significantly negative correlation (r = −0.995, p < 0.001) was observed between the nitrogenase activity and HD/CD, indicating an inhibitory effect of the fungal communities on the N-fixing bacterial activity.
4. Discussion
Historically, research in terms of N-fixing microbes was only conducted on legume species due to the specific characteristics of the rhizobia. However, recently, an increasing number of studies have also shown the existence of N-fixing bacteria on non-legume species, such as corn [17], miscanthus [12], and sugarcane [18]. N-fixing bacteria can reduce N2 in the air to a form (mainly NH4+) that can be absorbed and utilized by plants, especially under the N-deficiency condition [19]. This explains the positive correlation between the poor soil tolerance ability of the ramie plant and the N-fixing efficiency of the rhizosphere microbes. The N-fixing efficiency is co-contributed by the N-fixing bacteria number and nitrogenase activity. The different N-fixing bacteria number between genotypes can be explained by the plant that controls the N-fixing bacteria number by controlling the type and amount of root exudates [20,21]. Besides, the results of present study indicate the harmful fungal communities is also a factor in determining the N-fixing bacteria number. Additionally, the N-fixing efficiency is more related with the nitrogenase activity. Wang [22] found bacterial strains with nearly 10-times difference of nitrogenase activity in potato rhizosphere soil. Although the N-fixing potential is confirmed by ramie, there is still a large gap compared with legume species. For example, the highest N-fixing potential in this study is only 10–20% of that by soybean [23]. In the future, more studies are required to close the gap. Moreover, this paper only generally describes the existence of N-fixing bacteria in the ramie rhizosphere soil and its positive contribution in help ramie plant adapting to the poor soil condition. To apply this characteristic in future holobiont breeding, it is necessary to identify the specific species of the N-fixing bacteria and then create a stable genetic holobiont group of ramie plant-nitrogen fixing bacteria. For nitrogen cycling, urease also plays an important role in affecting the hydrolysis process of urea [24]. However, there is no significant difference in the urease activity among the tested genotypes, indicating that ramie plants did not adapt to a poor soil environment by affecting the utilization of urea.
This study also found that the P-solubilization by soil microorganisms makes a positive contribution to the adaptation of the ramie to poor soil. The soil microorganisms can secrete organic acids, protons, polysaccharides, and other substances. These substances could accelerate the conversion of insoluble P (e.g., rock P) to soluble form, which prevents the phosphorus availability and absorption by plants [25,26]. This explains the positive role of soil microorganisms in promoting the growth of ramie under poor soil conditions. The P-solubilization efficiency of the microorganism is mainly controlled by the generated phosphatase activity [27]. In this paper, the P-solubilization efficiency of the microorganism is consistent with the corresponding acid phosphatase activity, which also proves the applicability of this view in ramie plants. This study also finds that although there are significant differences in sucrase activity among different ramie genotypes, it is not significantly correlated with the adaptability of the ramie to poor soil. As an important material for catalyzing the decomposition of organic matter, the sucrase activity is closely related to the soil fertility [28]. However, the soil fertility is mainly increased in terms of organic matter, but not in the mineral elements such as N, P, and K.
The results of this study confirm the N-fixing and P-solubilization potential of the ramie rhizosphere soil microbes, especially from the poor soil tolerable genotype. In addition, the negative effects of the rhizosphere soil fungal community on the N-fixing bacterial are confirmed. These results are summarized based on the artificial experiments that were conducted in the sterilized conditions with only one or a few microbial strains. However, in reality, soil microbes grown in an unsterilized condition encounter a more diverse microbial community. The diverse condition suggests a more serious interaction potential between different microbes [29]. This could weaken or also strengthen the microbe’s effect tested in the artificial conditions. For this reason, a more realistic test of the N-fixing and P-solubilization potential of the ramie rhizosphere soil microbes is required. This can be conducted by isolating the N-fixing, P-solubilization microbes firstly, then adding the isolation inoculum to unsterilized soil and evaluating their potential on the growth and NUE improvement of the ramie plant. In future isolation research, the effect of the cultivation medium should be taken into consideration to get high selectivity and reliability. In this study, the Ashby medium was used in the N-fixing cultivation as it is one of the most common and suitable media for diazotrophs co-cultivation [30,31,32]. Actually, diazotroph is not only one kind of prokaryote, but includes several different kinds such as Rhizobium, Ensifer, Azospirillum [33,34]. Each kind of diazotroph has its own most suitable media as, in general, Ashby for Azotobacter [35] and yeast extract mannitol agar (YMA) for Rhizobium [31]. The exact species of the ramie rhizosphere contented N-fixing bacteria are still not confirmed. To isolate the N-fixing bacteria more effectively in the future study, different cultivation mediums, e.g., Beijerinckia medium and Derxia medium, should be compared.
5. Conclusions
This study confirms the existence of N-fixing and P-solubilization in the rhizosphere soil of ramie plants. These characteristics of rhizosphere soil microbes help ramie plants adapt to poor soil conditions. The N-fixing efficiency is co-contributed by the N-fixing bacteria number and strong nitrogenase activity. One reason for the low N-fixing efficiency of intolerable genotypes is that the fungal communities in the corresponding rhizosphere soil strongly reduce the nitrogenase activity, also in terms of N-fixing bacteria number.
Author Contributions
S.W.: Methodology, Investigation, Formal analysis, Writing—Original Draft Preparation; H.J.: Resources, Formal analysis, Visualization; Y.J.: Conceptualization, Supervision, Project Administration. All authors have read and agreed to the published version of the manuscript.
Funding
This study was financially supported by the National Natural Science Foundation of China (32071940), China’s National Key R&D Program (2019YFD1002205-3 & 2017FY100604-02), Foundation for the Construction of Innovative Hunan (2020NK2028) and Research Project Founded by Hunan Education Department (20C0957).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We appreciate Yu Dai, Qiao Liu and Jing Xiao for their help in the process of experiment implementation and chemical analysis. We would also like to thank Shuai Xue for his assistance in experiment design and Yasir Iqbal for the language editing of this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Tatar, Ö.; Ilker, E. Impact of different nitrogen and potassium application on yield and fiber quality of ramie (Boehmeria nivea). Int. J. Agric. Biol. 2010, 14, 369–372. [Google Scholar]
- Liu, L.J.; Chen, H.Q. Effect of planting density and fertilizer application on fiber yield of ramie (Boehmeria nivea). J. Integr. Agric. 2012, 11, 1199–1206. [Google Scholar] [CrossRef]
- Wang, P.; Wang, Y.M. Effects of different fertilizations on maize nutrient uptake in Hemi-dry-land. Chin. J. Soil Sci. 2009, 40, 1135–1138. [Google Scholar]
- Miao, Y.; Stewart, B.A.; Zhang, F. Long-term experiments for sustainable nutrient management in China. A review. Agron. Sustain. Dev. 2011, 31, 397–414. [Google Scholar] [CrossRef] [Green Version]
- Jiao, X.Q.; Mongol, N. The transformation of agriculture in China: Looking back and looking forward. J. Integr. Agric. 2018, 17, 755–764. [Google Scholar] [CrossRef] [Green Version]
- Gopal, M.; Gupta, A. Microbiome selection could spur next-generation plant breeding strategies. Front. Microbiol. 2016, 7, 1971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Z.; Jousset, A. Plant breeding goes microbial. Trends Plant Sci. 2012, 22, 555–558. [Google Scholar] [CrossRef] [PubMed]
- Kroll, S.; Agler, M.T. Genomic dissection of host-microbe and microbe-microbe interactions for advanced plant breeding. Curr. Opin. Plant Biol. 2017, 36, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Baligar, V.C.; Fageria, N.K. Nutrient use efficiency in plants. Commun. Soil Sci. Plan. 2001, 32, 921–950. [Google Scholar] [CrossRef]
- Wu, S.; Xue, S. Seasonal nutrient cycling and enrichment of nutrient-related soil microbes aid in the adaptation of ramie (Boehmeria nivea L.) to nutrient-deficient conditions. Front. Plant Sci. 2021, 12, 644904. [Google Scholar] [CrossRef]
- Xu, Y.; Zheng, C. Quantitative assessment of the potential for soil improvement by planting Miscanthus on saline-alkaline soil and the underlying microbial mechanism. GCB Bioenergy 2021, 3, 1191–1205. [Google Scholar] [CrossRef]
- Liu, Y.; Ludewig, U. Nitrogen-dependent bacterial community shifts in root, rhizome and rhizosphere of nutrient-efficient Miscanthus x giganteus from long-term field trials. GCB Bioenergy 2019, 11, 1334–1347. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.B.; Sayyed, R.Z. Phosphate solubilizing microbes: Sustainable approach for managing phosphorus deficiency in agricultural soils. SpringerPlus 2013, 2, 587. [Google Scholar] [CrossRef] [Green Version]
- Nannipieri, P.; Trasar-Cepeda, C. Soil enzyme activity: A brief history and biochemistry as a basis for appropriate interpretations and meta-analysis. Biol. Fertil. Soils 2018, 54, 11–19. [Google Scholar] [CrossRef]
- Chaudhary, D.R.; Gautam, R.K. Nutrients, microbial community structure and functional gene abundance of rhizosphere and bulk soils of halophytes. Appl. Soil Ecol. 2015, 91, 16–26. [Google Scholar] [CrossRef]
- Haskett, T.L.; Knights, H.E. A simple in situ assay to assess plant-associative bacterial nitrogenase activity. Front. Microbiol. 2021, 12, 690439. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Bai, Y. Maize aerial roots fix atmospheric N2 by interacting with nitrogen fixing bacteria. Sci. Sin. Vitae 2019, 49, 89–90. [Google Scholar]
- de Lima, D.R.M.; dos Santos, I.B. Genetic diversity of N-fixing and plant growth-promoting bacterial community in different sugarcane genotypes, association habitat and phenological phase of the crop. Arch. Microbiol. 2021, 203, 1089–1105. [Google Scholar] [CrossRef]
- Rocha, K.F.; Kuramae, E.E. Microbial N-cycling gene abundance is affected by cover crop specie and development stage in an integrated cropping system. Arch. Microbiol. 2020, 202, 2005–2012. [Google Scholar] [CrossRef] [PubMed]
- Dennis, P.G.; Miller, A.J. Are root exudates more important than other sources of rhizodeposits in structuring rhizosphere bacterial communities? FEMS Microbiol. Ecol. 2010, 72, 313–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Pan, F. Response of symbiotic and asymbiotic nitrogen-fixing microorganisms to nitrogen fertilizer application. J. Soils Sediment. 2019, 19, 1948–1958. [Google Scholar] [CrossRef]
- Wang, W.; Yang, S. Isolation and characterization of nitrogen fixing and phosphate solubilizing bacteria from potato rhizosphere of reclaimed cropland. Nat. Sci. Ed. 2019, 47, 127–133. [Google Scholar]
- Wang, J.; Xu, Y. Effects of rhizobium, soybean variety, soil type on nitrogenase activity. J. Northeast Agric. Univ. 2008, 39, 36–39. [Google Scholar]
- Fu, Q.; Abadie, M. Effects of urease and nitrification inhibitors on soil N, nitrifier abundance and activity in a sandy loam soil. Biol. Fertil. Soils 2020, 56, 185–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, A.; Saha, T.N. Efficient microorganism compost benefits plant growth and improves soil health in Calendula and Marigold. Hortic. Plant J. 2017, 3, 67–72. [Google Scholar] [CrossRef]
- Adetunji, A.T.; Lewu, F.B. The biological activities of β-glucosidase, phosphatase and urease as soil quality indicators: A review. J. Plant Nutr. Soil Sci. 2017, 17, 794–807. [Google Scholar] [CrossRef] [Green Version]
- Widdig, M.; Schleuss, P.M. Nitrogen and phosphorus additions alter the abundance of phosphorus-solubilizing bacteria and phosphatase activity in grassland soils. Front. Environ. Sci. 2019, 7, 185. [Google Scholar] [CrossRef] [Green Version]
- Guo, Q.Q.; Xiao, M.R. Polyester microfiber and natural organic matter impact microbial communities, carbon-degraded enzymes, and carbon accumulation in a clayey soil. J. Hazard. Mater. 2021, 405, 124701. [Google Scholar] [CrossRef] [PubMed]
- Ordoñez, Y.M.; Fernandez, B.R. Bacteria with phosphate solubilizing capacity alter mycorrhizal fungal growth both inside and outside the root and in the presence of native microbial communities. PLoS ONE 2016, 11, e0154438. [Google Scholar] [CrossRef]
- Stella, M.; Suhaimi, M. Selection of suitable growth medium for free-living diazotrophs isolated from compost. J. Trop. Agric. Food Sci. 2010, 38, 211–219. [Google Scholar]
- Ramírez, M.D.A.; España, M. Genetic diversity and characterization of symbiotic bacteria isolated from endemic Phaseolus cultivars located in contrasting agroecosystems in Venezuela. Microbes Environ. 2021, 36, ME20157. [Google Scholar] [CrossRef]
- Wickramasinghe, W.R.K.D.W.K.V.; Girija, D. Multi-phasic nitrogen fixing plant growth promoting rhizobacteria as biofertilizer for rice cultivation. Res. J. Agric. Sci. 2021, 12, 399–404. [Google Scholar]
- Soumare, A.; Diedhiou, A.G. Exploiting biological nitrogen fixation: A route towards a sustainable agriculture. Plants 2020, 9, 1011. [Google Scholar] [CrossRef] [PubMed]
- Gopalakrishnan, S.; Srinivas, V. Nitrogen fixation, plant growth and yield enhancements by diazotrophic growth-promoting bacteria in two cultivars of chickpea (Cicero arietinum L.). Biocatal. Agric. Biotechnol. 2017, 11, 116–123. [Google Scholar] [CrossRef]
- Jiménez, D.J.; Montaña, J.S. Characterization of free nitrogen fixing bacteria of the genus Azotobacter in organic vegetable-grown Colombian soils. Braz. J. Microbiol. 2011, 42, 846–858. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Variations of the rhizosphere soil content nitrogen-fixing microbe numbers (a), P-solubilization efficiency of the soil microbes (b) and anti-N-fixing-bacteria activity by fungal communities (c) between different ramie germplasms. Different capital letters within each measured trait indicate the least significant differences at a p < 0.01 level. N/A means not available. XZ-XB, Xiangzhu XB; ZZ-1, Zhongzhu 1; XZ-X2, Xiangzhu X2; XZ-3, Xiangzhu 3. CK is the control without ramie cultivation. HD/CD represents the halo zone diameter/colony diameter that measured in the antagonistic test.
Figure 1.
Variations of the rhizosphere soil content nitrogen-fixing microbe numbers (a), P-solubilization efficiency of the soil microbes (b) and anti-N-fixing-bacteria activity by fungal communities (c) between different ramie germplasms. Different capital letters within each measured trait indicate the least significant differences at a p < 0.01 level. N/A means not available. XZ-XB, Xiangzhu XB; ZZ-1, Zhongzhu 1; XZ-X2, Xiangzhu X2; XZ-3, Xiangzhu 3. CK is the control without ramie cultivation. HD/CD represents the halo zone diameter/colony diameter that measured in the antagonistic test.

Figure 2.
Comparison of the soil enzyme activity of nitrogenase (a), acid phosphatase (b), urease (c), and sucrose (d) between different ramie germplasms. Different capital letters within each measured trait indicate least significant differences at p < 0.01 level. XZ-XB, Xiangzhu XB; ZZ-1, Zhongzhu 1; XZ-X2, Xiangzhu X2; XZ-3, Xiangzhu 3. CK is the control without ramie cultivation.
Figure 2.
Comparison of the soil enzyme activity of nitrogenase (a), acid phosphatase (b), urease (c), and sucrose (d) between different ramie germplasms. Different capital letters within each measured trait indicate least significant differences at p < 0.01 level. XZ-XB, Xiangzhu XB; ZZ-1, Zhongzhu 1; XZ-X2, Xiangzhu X2; XZ-3, Xiangzhu 3. CK is the control without ramie cultivation.


{kind=link}
{kind=link}
Table 1.
Correlations between the soil enzyme activity (nitrogenase, acid phosphatase, urease, and sucrase) and the ability of ramie to tolerate poor soil, expressed by the normalized field performance index (NFPI).
Table 1.
Correlations between the soil enzyme activity (nitrogenase, acid phosphatase, urease, and sucrase) and the ability of ramie to tolerate poor soil, expressed by the normalized field performance index (NFPI).
| Nitrogenase Activity | Acid Phosphatase Activity | Urease Activity | Sucrase Activity | |
|---|---|---|---|---|
| r | 0.899 | 0.846 | 0.698 | 0.256 |
| p | <0.001 | 0.005 | 0.012 | 0.421 |
Note: Detail information of NFPI is shown in the published study of [10]; r: Pearson’s coefficients; p: p-values of the Pearson’s coefficients.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wu, S.; Jie, H.; Jie, Y. Role of Rhizosphere Soil Microbes in Adapting Ramie (Boehmeria nivea L.) Plants to Poor Soil Conditions through N-Fixing and P-Solubilization. Agronomy 2021, 11, 2096. https://doi.org/10.3390/agronomy11112096
AMA Style
Wu S, Jie H, Jie Y. Role of Rhizosphere Soil Microbes in Adapting Ramie (Boehmeria nivea L.) Plants to Poor Soil Conditions through N-Fixing and P-Solubilization. Agronomy. 2021; 11(11):2096. https://doi.org/10.3390/agronomy11112096
Chicago/Turabian StyleWu, Shenglan, Hongdong Jie, and Yucheng Jie. 2021. "Role of Rhizosphere Soil Microbes in Adapting Ramie (Boehmeria nivea L.) Plants to Poor Soil Conditions through N-Fixing and P-Solubilization" Agronomy 11, no. 11: 2096. https://doi.org/10.3390/agronomy11112096
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.