1. Introduction
Pollination is one of the most important issues concerning flowering plants, both from the point of view of conservation biology [
1] and agriculture [
2]. A practical problem in greenhouses is that the maximal number of adult pollinators maintained by greenhouse crops is less than what is needed for an economically level of fructification [
3]. For instance, a problem in the tomato flower is the absence of nectar [
4]. Thus, tomato plants are apparently of little or no value in attracting bees [
2]. In [
5,
6] stated that the blossom “contains little or no nectar.” It is reported that bumble bees “gathered chiefly pollen’’ from tomato flowers [
7]. Thus, if nectar is produced, a question that should be settled, it is of little significance in the relation of insect pollination of tomatoes. The pollen is more attractive to wild bees than honey bees. There are two possible solutions: (a) Release of pollinators other than honey-bees and bumblebees, such as syrphids [
8], since the flowers of tomato are widely visited by hoverfly species [
9,
10]. (b) Supplying of additional food to pollinators in order to maintain more adult insects than what is maintained by greenhouse crops [
11,
12].
Syrphids, commonly known as hoverflies or flower flies, make up one of the largest families of Diptera. To date, the family has 6674 species in 284 genera, with most species distributed in the Neotropical, Nearctic, and Palaeotropical regions of the world [
13]. The adults of hoverflies, as their name implies, are usually found at flowers or hovering in the bright sunlight. They require pollen, nectar, and honeydew during the adult stage [
14,
15,
16]. Haslett [
14] studied the role of flower nectar and pollen, by dissecting the adult females of
Rhyngia campestris Meigen (Diptera: Syrphidae) captured in the field. Thus, this author found that nectar seems to be necessary at the beginning and at the end of the females’ ovarian development. On the contrary, pollen is essential for such ovarian development and subsequent oviposition. However, unpublished data [
17] seem to indicate that adult females and males of
Eristalis tenax (L.) (Diptera: Syrphidae), under laboratory conditions, fed with different combinations of honey, pollen, food supplements, and vitamins, did not show significant differences in adult longevity, either in fertility and fecundity of females, when pollen, alone or in combination, was present in the diet of adults [
17]. All the above discussion seems to indicate that pollen is a fundamental feeding factor for adult longevity and fecundity. On the contrary, the immature stages of Syrphidae species present different behaviors, i.e., there are species in which the larvae are predatory, and others present a phytophagous or mycophagous diet; finally, there is a wide group of Syrphidae species that are saprophagous. Therefore, this last group needs a specific food substrate [
18]. In addition, several species of this family of insects have demonstrated their effectiveness as pollen vectors in the main horticultural crops in greenhouses [
19,
20,
21].
The present study is motivated by the pollination of tomato by
Eristalis tenax. The adults of this species feed on flowers, while their immatures are saprophagous. In this paper, we will use a theoretical approach, namely, mathematical models, and will start out from the models of theoretical ecology (e.g., [
22,
23,
24]). In fact, to make crop pollination economic and efficient, a dynamic optimization approach is necessary, which on its turn is necessarily based on a dynamic mathematical model (differential equations) available in theoretical ecology. In this way, the optimal control of pollination can be applied to quantify the amount of released agents necessary for a sufficient pollination. Since the classical theoretical models (e.g., [
22]) do not take account of some important factors of our problem, we have to develop models that, as far as possible, involve such details of this practical problem (e.g., longevity of adults and different habitat for larvae and adults). Our method addresses the above practical problem by developing models that, as far as possible, involve the details of the crop pollination situation. We address two basic questions.
The first one is: How many flies are needed for the desired level of pollination? (Given an expected economic yield, the necessary pollination level can be experimentally determined.) To answer this question, we will apply a Markov model, which has been already used to study pollination (e.g., [
25,
26]). The Markov model is one of the simplest mathematical tools to handle the general pollination process, when during a day the flowers are emptied. (The number of nonvisited flowers decreases, but due to the repeated visits, new pollination takes place with smaller and smaller probability.)
The second question is: In which way can we maintain this desired pollinator density (e.g., [
23,
27])? Here, instead of the extremely inefficient continuous release of adult flies in the greenhouse, we propose another way: Initially we release adult flies. Then, we maintain the hoverfly population at the desired density by supplementary feeding (sugar and pollen), ensuring the reproduction of hoverfly. We will study the case when in the greenhouse, we also release larvae supplied with special larval substrate provided in containers distributed throughout the greenhouse. We suppose the amount of food for the juveniles is sufficient both for the juveniles released and for the offspring of the adult released in the greenhouse. Hence, we will need a two-stage model, including the juvenile stage. On the other hand, the supplementary food supply can increase both fecundity and longevity of adult flies, but it does not increase the survival of the hoverfly larvae. Furthermore, in our case, supplementary food increases their longevity. The longevity already plays a crucial role in the models of host–parasite systems (e.g., [
28,
29]), which is quite different from pollination. In particular, we introduce a nonadditive model to take into account the increased longevity due to supplementary food [
17] (see
Section 2.3).
In our approach, the effect of feeding on the population dynamics of the hoverfly, will be described in terms of an optimal control model. Such models have already been applied to the optimization of release strategies in biological pest control, see e.g., [
30,
31]. In the present case, the optimal control model will be used to keep the adult hoverfly density above, but close to the desired level.
The main aim of this paper is to face at least two basic questions, as we already mentioned: 1. How many hoverflies are necessary for an economically suitable level of pollination of a given amount of crop plants? 2. In which way can we maintain a required pollinator density higher than what is maintained by the greenhouse? Now, in the present paper, we deal with the first phase of our research project, building up a conceptual model based on mathematical methods, addressing these questions.
Finally, we emphasize that this is a theoretical work, and literature data are used only to show the potentialities of the model (and may be helpful for a sensitivity analysis too).
2. Materials and Methods
At this first stage, we develop a conceptual theoretical model that will help the design of appropriate experiments, and only at the second stage of the research, we will carry out experiments. Therefore, in the present paper for the simulations, we will use only illustrative data, based on general experience, in part on data of the literature.
The species considered for this work was
Eristalis tenax (L.) (Diptera, Syrphidae: Eristalinae). Data on its biology and ecology have been collected from the available literature on this species and other nearby species in the same genus (e.g., [
14,
27,
32,
33,
34,
35,
36]).
Our aim is to maintain the hoverfly population at a density necessary for a sufficient pollination, at minimum cost. To this end, we have to go through the following three steps. (A) We introduce a stochastic model to calculate the pollinator density needed for the economically successful pollination of a given number of plants, during a fixed time interval (see
Section 2.1). (B) We set up a stage-specific deterministic population dynamics for the hoverfly (see
Section 2.2). (C) In these dynamics, we introduce the effect of time-dependent quantity of supplementary food intended for the stable maintenance of the economically desired pollinator density. This leads to a deterministic optimal control model for the hoverfly with added supplementary food (See
Section 2.3). These three models are built upon each other in the sense that the desired density of pollinator is calculated by a stochastic model and then, based on a deterministic population dynamics, an optimal control model is set up.
2.1. How Many Pollinators Are Needed for an Economically Successful Pollination of Crop Plants?
In our simplified model, there are flowers in a greenhouse waiting for pollinations. For our conceptual stochastic model, we use some simplifying assumptions about the flowers. We suppose that there is no intraflower variability in the probability of being visited (in reality, some flowers could be exposed at the top of the plant, and others could be hidden behind leaves). Moreover, we also assume that M is fixed during flowering period.
In our model, pollination agents are insects (e.g., pollinators of a single species), there are of them (). Pollinators are continuously searching flowers during their daily active period (when pollinators feed intensively), denoted by T. During this active period, we consider the following events, since the pollinator does not know if a flower is already exhausted by another pollinator: (a) searching: a pollinator stops feeding and starts searching. When a pollinator finds a flower, it faces the following possibilities: (b) pollinate an unfertilized flower. A simplifying assumption of our conceptual stochastic model on pollination is that a single visit by a pollinator is enough for the pollination of a flower. (c) Find an unoccupied but pollinated flower, and still spend a certain amount of time in it. (d) Find a flower occupied by another adult, and start a new search. In other words, there is no interference between pollinators during feeding (i.e., there is no fighting between pollinators). We emphasize that our model takes into account that at the beginning of the activity period T, more pollination is carried out than at the end of T, since the number of pollinated flowers increases with time, consequently more and more times, the pollinators find flowers that are already pollinated. In other words, during each day, the pollinators find less and less virgin flowers, because they have already visited a part of the flowers before, repeatedly. We also assume that during T, all flies are active. This simplifying assumption based on the fact that for the desired pollination rate, we need more flies than the maximal number of pollinators maintained by the tomato, thus the overwhelming majority of flies have not fed (are not full), thus they will not stop visiting flowers. For the simplicity, we also do not take into account other activities during the feeding period of a day, such as mating, fighting, and displaying.
Both searching and handling/pollinating take time. Searching times are supposed to be independent and exponentially distributed random variables with expectation
. The same holds for handling times with mean
. For the sake of simplicity, we suppose that the flower found at the end of the searching time is random with uniform distribution, though, in fact, closer flowers are found with greater probability. We also suppose that the mean time a pollinator spends in an exhausted flower and that in a flower waiting for pollination are the same, though feeding presumably requires more time. We note, our technical assumption on exponential distribution makes our investigation easier, but we note that [
37] fitted gamma distribution to the handling time for the herbivore. Observe, that at this starting stage of our model, we only concentrate on the pollination, so we do not consider other activities of the pollinator (like rest on a flower during feeding, interaction between pollinators, e.g., mating and fighting, or egg laying, etc.), which also take time, so these other activities can decrease the efficiency of pollination.
Our object is to find an approximate answer to the question “Given a fixed, relatively long time interval (a day, say) and a small positive number , how many pollinators are needed so that the proportion of unpollinated flowers would fall below by the end of the given time period?” We assume and are dwarfed by .
A naïve estimation can easily be derived. Since the number of pollinators is much smaller than the number of flowers, it is of small probability that a pollinator finds an occupied flower. Neglecting this unlikely possibility, we can say that
pollinators perform approximately
turns of searching and handling during time
. Considering them independent, the chance that a given flower has not been visited by the end of the monitoring period
is
The justification of this naïve estimation in a more elaborated stochastic model is presented in
Appendix A.1.
The temperature has not been included among these parameters. Although insects are ectotherms, we have considered that environmental factors in greenhouse crops allow us to simplify the model.
2.2. A Population Dynamic Model for Pollinator
As in
Section 2, we suppose that in the greenhouse, the density of monocultural crops does not change. We deal with monocultural crops, so we suppose that both the number of flowers and the quantity of nectar and pollen produced in unit time are constant during the flowering period of the plant. Clearly, this simplifying assumption does not take into account that the proportion of plants and flowers on plants flowering, and hence nectar and pollen availability, will vary through the lifetime of a crop.
This assumption simplifies the modelling, because we do not have to take into consideration how fertility, due to the pollinators, can affect the plant density throughout generations. In mathematical terms, this also means that the plant dynamics can be neglected, since we concentrate on greenhouses. Another simplifying consequence of the constant plant density is that, in the dynamics of pollinator density, the numerical response (i.e., α in our model below) can be considered constant. Therefore, here we will consider the following deterministic model for the density change of hoverflies.
Let
and
be the density of juvenile and adult pollinators, respectively,
the number of juveniles produced by an adult hoverfly in unit time (egg number),
the rate of leaving juvenile stage (by either death or development),
and
the death rates of juvenile and adult, respectively (
);
K the coefficient of the density-dependent competition. Then the dynamic model is
Observe that in the above model, there is no competition for food between juveniles and adults.
Explanation of the first equation: what happens to the juveniles in unit time? Using the above introduced parameters, the juvenile population is increased by the egg laying of adults (i.e., by ) and decreased by leaving the juvenile stage (i.e., by ). The food competition between larvae does not appear in Equation (1), since larvae are supplied with sufficient food.
Explanation of the second equation: what happens to an adult in unit time? We suppose two types of issues:
- (a)
The survival rate of a “newcomer” adult is , they are not sensitive to either the quality or the quantity of food, since they have some reserve from their larva stage, collected from the ground (see the first term of the right-hand side of (2)).
- (b)
An adult emerged (see the second term of the right-hand side of (2)) may die for two reasons:
by competition for food among them, implying a density-dependent death rate: ;
by ageing, this is not density-dependent, it is characterized just by a death rate . Here, the individuals are supposed to be “ever-young” (i.e., the imago’s death rate does not depend on age), e.g., there is an exponentially distributed survival. In the latter case, the life span expected at emergence (shortly also often called longevity) is .
In order to survive, an individual must survive both the death causes in unit time, which occurs with probability
, thus the survival probability of an individual in unit time is
and hence the resulting death rate (the proportion of individuals dying in unit time) is
Therefore, the number of individuals died in unit time is
Remark 1. The death rate of the adult can also be written as An explanation to the latter may be the following: suppose that an individual can face death risk either by starving or by aging or both. If for example 10, individuals died only by starving, 10 only by aging, 5 faced both death risks, 15 individuals died by starving, and 15 by aging, but in this way, those having faced both death causes, are counted twice, so the latter should be subtracted to obtain the actual number of individuals that died. As a consequence, if there are two death causes then the sum of the two death rates is an upper estimate of the actual death rate.
In
Appendix A.2, we show that under a simple condition on the model parameters, an asymptotically stable equilibrium of dynamics (1)–(2) is guaranteed. Namely, equilibrium
x* with
is positive, if
Remark 2. Condition (3) is obviously fulfilled if the fertility rate is high enough. Furthermore, this condition also implies that x* is locally asymptotically stable (see Appendix A.2). If the convergence is quick enough, the insect population reaches its stable density during a short time respect to the crop season. If not, the asymptotically stable equilibrium would not help to reach the desired number of adults. 2.3. A Deterministic Optimal Control Model for the Hoverfly with Added Supplementary Food
A first idea might be a continuous release of adult flies in the greenhouse, considering the dynamics of the adult without reproduction, obtained from systems (1)–(2):
the purely adult population would die out very quickly, therefore, a control by the release of adults according to the control equation
would be extremely inefficient.
2.4. Controlling Pollination Level by Supplementary Food
A common practice in both pollinating and entomophagous insects is the application of nutritional supplements to increase their activity in crop pollination or application of biological agents in crops and/or greenhouses (e.g., [
15]). In the case of pollinating insects, applications of food and nutritional supplements would be applied in water by spraying on plants (e.g., [
38]) or better placing feeders in different sites inside the greenhouse. Evidently, solar radiation, air oxygen, and other factors can degrade the food supplement. Therefore, it will be necessary to re-spray the food supplement repeatedly. Remember, the juveniles need different supplementary food.
The supplementary food added by spray has three possible effects:
Decreases the density-dependent food competition between adults (e.g., [
39]): Let us denote by
, the increment of the coefficient
K-nek due to the supplementary food for the hoverfly given in unit time by the farmer.
Increases longevity (expected life span) of the adult (e.g., [
16]): Let
denote the effect of supplementary food decreasing the death rate of adult hoverfly, 0
. The pollinator mainly needs nectar for food [
17].
Increases fecundity (number of eggs of a female) (e.g., [
14]): Let
be the effect of supplementary food increasing fecundity.
We can consider a “feeding function” depending on the added sugar and pollen
,
This function is based on the physiology of the insect [
17]. The exact forms of functions
are unknown, in our dynamic model. We can consider the following simplest theoretical feeding functions based on biological intuition and experimental practice.
- (a)
Density-dependent food competition function: the pollinators mainly need nectar for survival; thus, the supplementary sugar decreases the starvation of adults. We assume that this function is linear.
where
is a positive constant.
- (b)
The longevity function displays a saturation with respect to
, since the death rate cannot decrease arbitrarily.
where
and
are positive constants. Since the latter function is increasing and is saturated at value
if condition
holds.
- (c)
The fertility function may be
where
and
are positive constants. This function, for each fixed
, is a saturation function of
.
Since now, the pollinator population has two types of food at disposal, the optimal foraging strategy of the pollinator could also be taken into consideration (e.g., [
40]). For the sake of simplicity, we will not consider the optimal foraging behavior in our present conceptual models (4) and (5) below. This simplifying condition can be a good approximation, if the quantity of food sprayed on the plants can be considered constant between two actions (e.g., the added food does not oxidize or deteriorate in the meanwhile). Then, even if the hoverfly consumes the added food and also visits the flowers, being the densities of both resources fixed, the optimal foraging is also fixed, hence in the model describing the dynamics of pollinator density, the food-dependent parameters are also fixed (and the optimal foraging preference is included in the interaction parameters).
Now inserting the above functions
into dynamics (1)–(2), we have the following control dynamics:
Hence, the optimal control problem to be solved is the following: We start with initial release of juvenile and adult flies
, with
, where
is the number of adult hoverfly necessary for the satisfactory pollination, calculated according to the reasoning of
Section 2.1. Then, with time-dependent food supply
as control in a time interval
], with control dynamics (4)–(5), we will solve the problem of minimization of the total costs of the food supply under the condition that
. For mathematical details, see
Appendix A.3.
The above optimal control problem has been solved with a toolbox developed for MATLAB in [
41,
42]. The results are plotted in the Results section.
4. Discussion
In this paper, our main objective is to show that with the aid of a pilot theoretical study, a deeper and more detailed insight can be gained into our concrete problem on pollination. However, in general, we suggest the introduction of a new approach, where in the first step, based on previous knowledge, a mathematical model is set up, the behavior of which would be the basis for certain experimental design. Then in the light of the future experimental results, we may refine the model, and design further experiments, and so forth. In this sense, the present paper is a modelling-methodological development, to be followed by the next stage when the necessary parameters will be experimentally measured, and the model itself might be also refined.
4.1. Discussion of Operational Issues
Our conceptual model provides the following predictions:
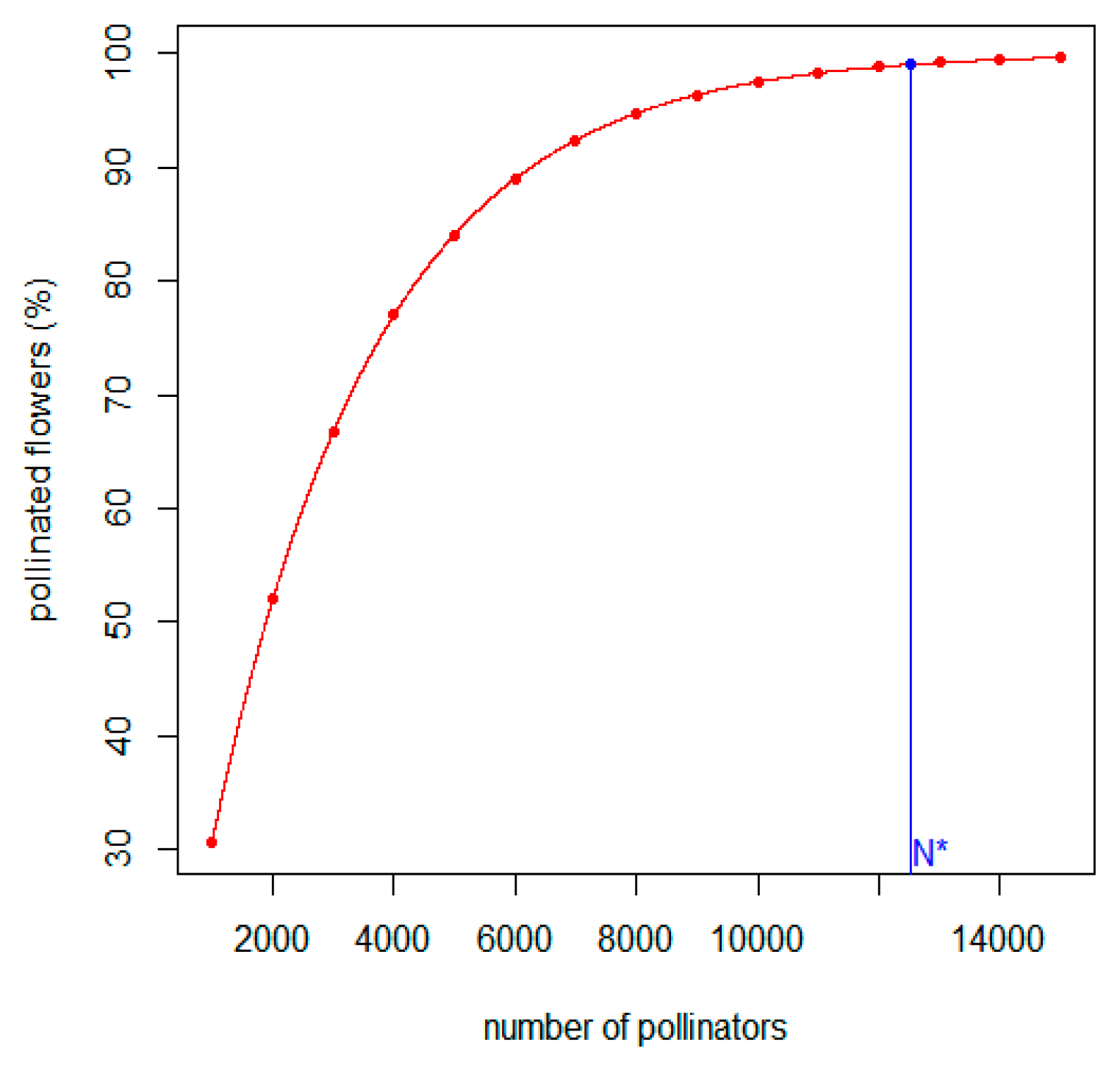
Figure 1 calls the attention to the fact that with increasing number of pollinators, the proportion of pollinated flowers saturates. For example, from 65% to reach 92%, the number of pollinators must be doubled. In practice, this means that the number of pollinators necessary for a high percentage pollination may exceed the maximum number of adult pollinators maintained by the crops. This nonlinear dependence will also remain in more sophisticated models, because the pollinators visit the already pollinated flowers again and again, thus for a higher pollination, substantially more pollinators are needed. Therefore, food supply may facilitate pollination, since it helps to maintain a substantially higher adult pollinator population. Our optimal control model also makes it possible to calculate the necessary food supply, minimizing its cost, once we have obtained the real model parameters in the second stage of our project. Further model calculations should be interpreted similarly.
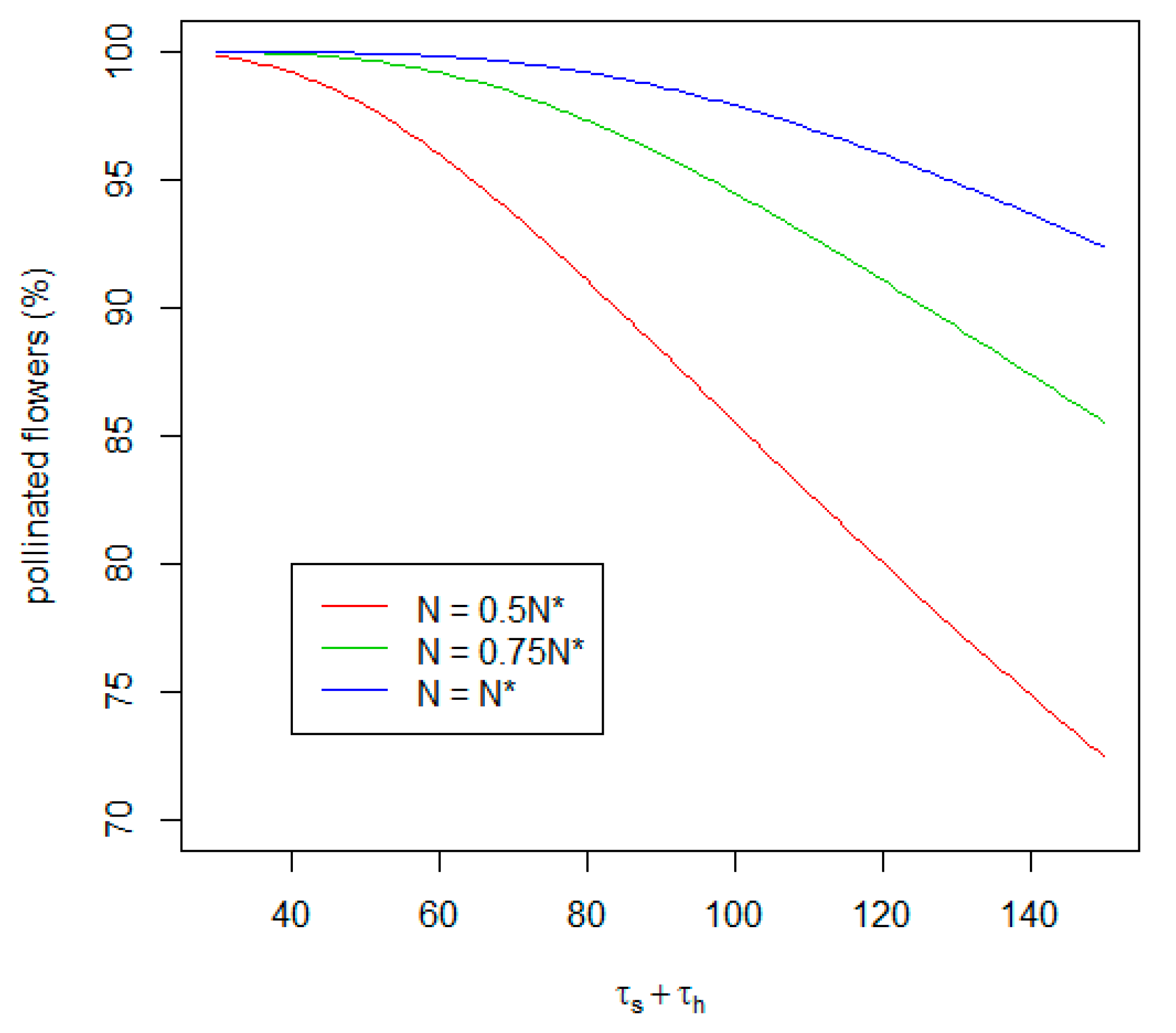
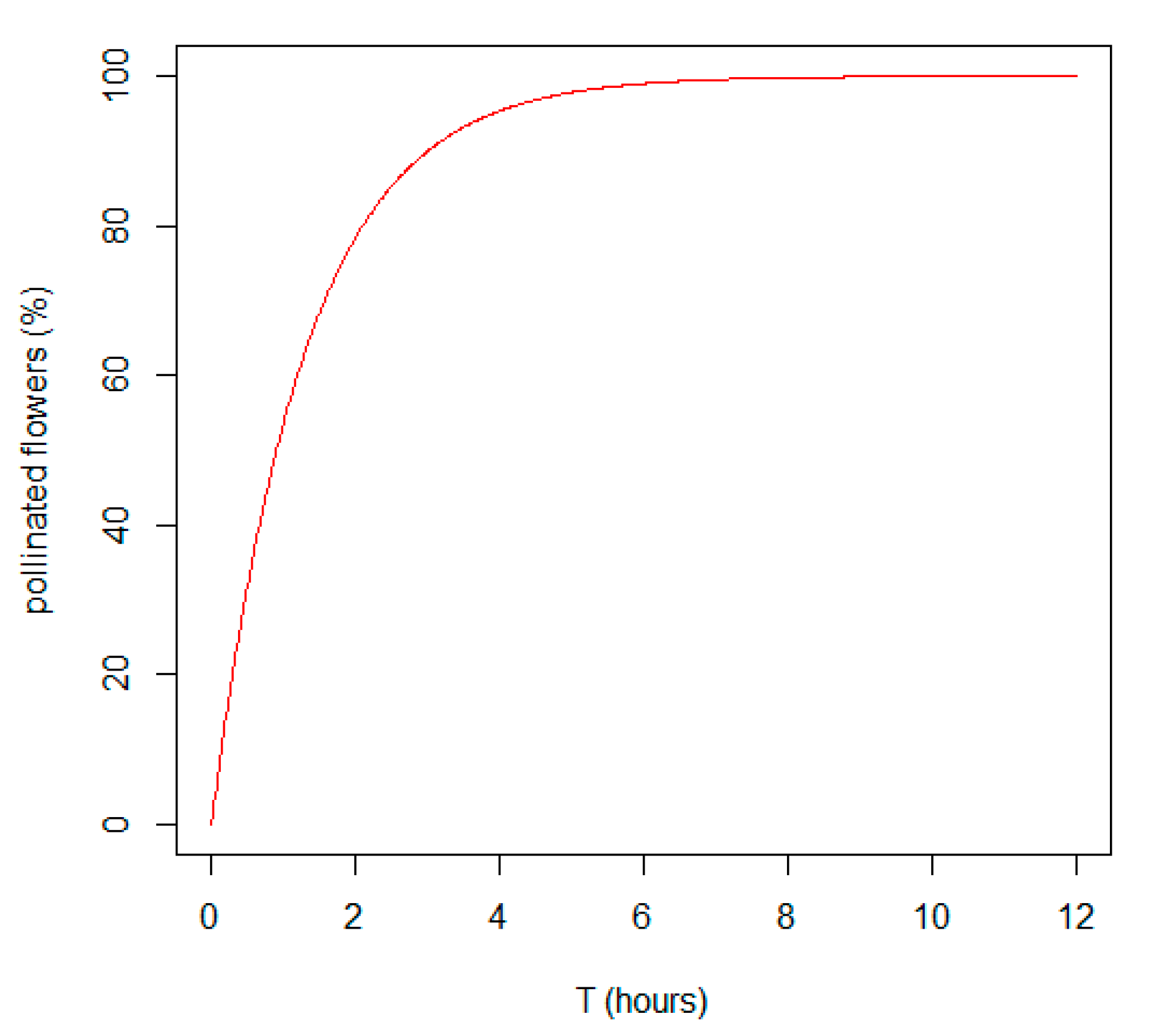
Our model has also shown to what extend the decrease in searching and handling time (see
Figure 2) and the increase in activity time (see
Figure 3) would increase the efficiency of the pollinators. This knowledge may facilitate the choice of new pollinator agents.
Hoverflies, like E. tenax and species with similar size and behavior, are already being applied for pollination in greenhouses. According to the protocol in practice, adult flies are released twice. Larvae feed on substrate placed on the ground, implying no hygienic risk. Therefore, we would suggest to release also juvenile at the beginning, with mold added.
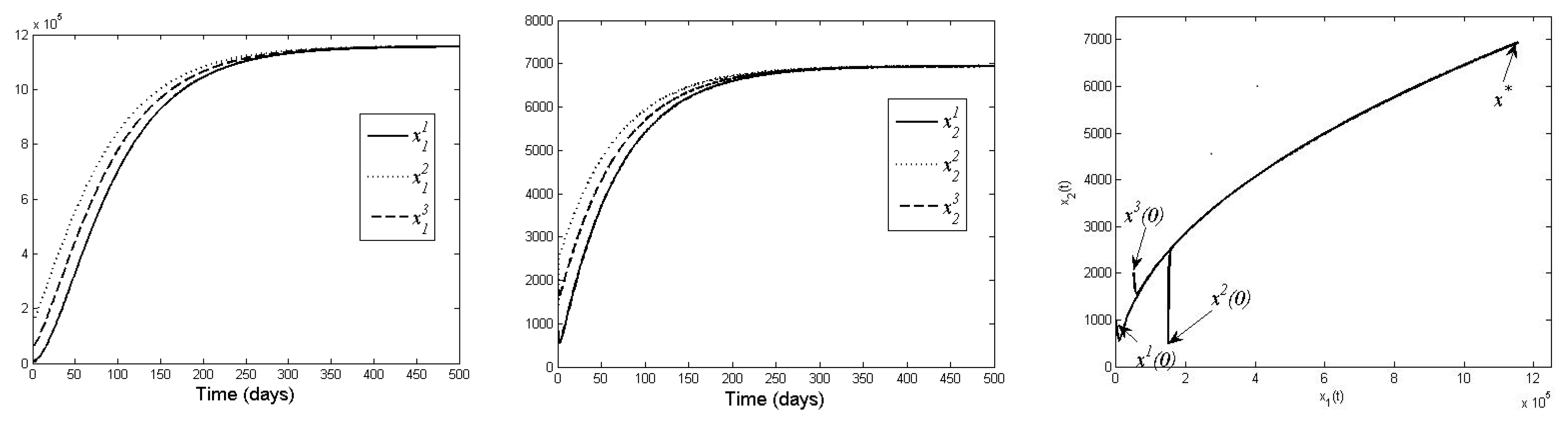
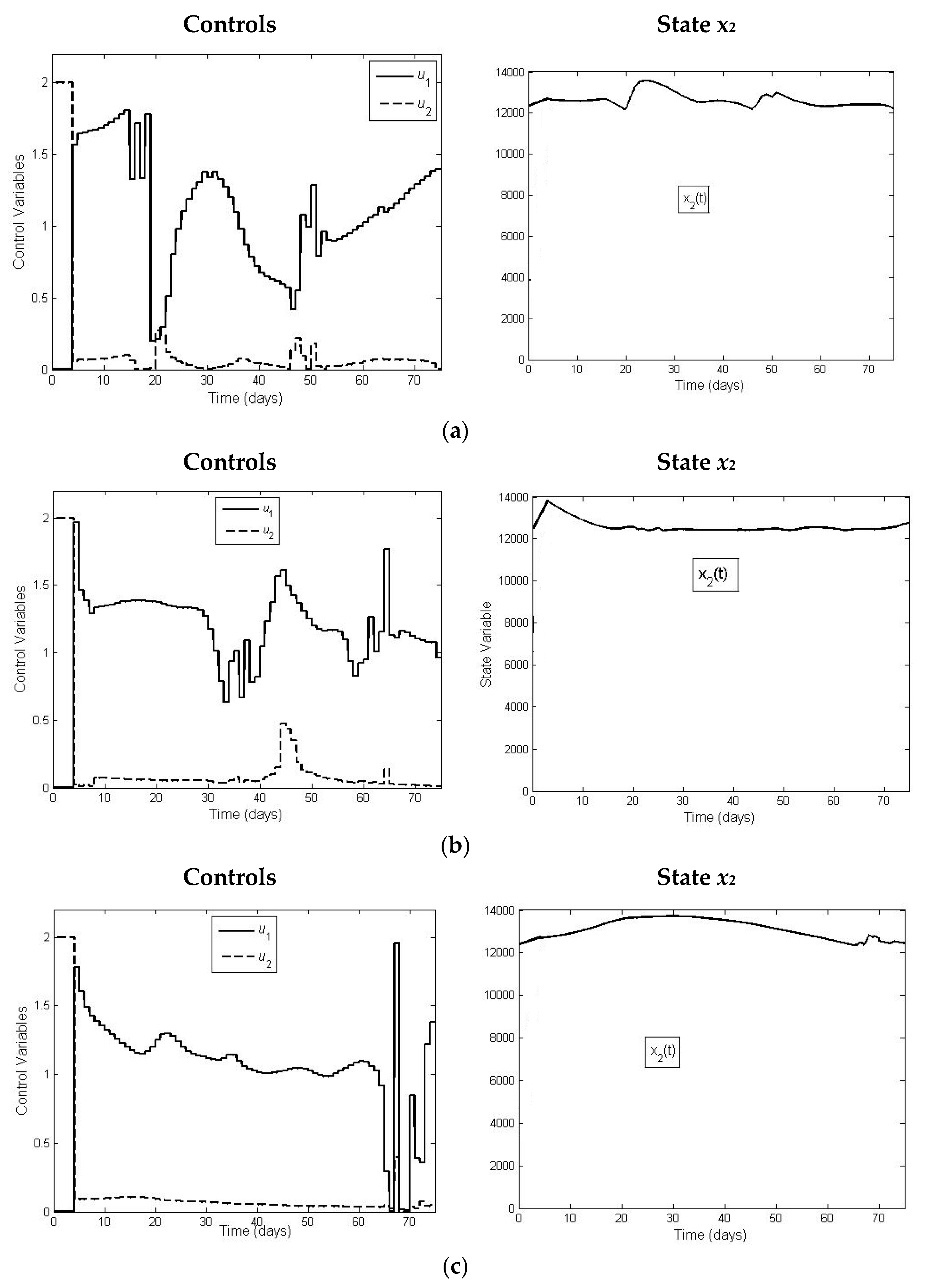
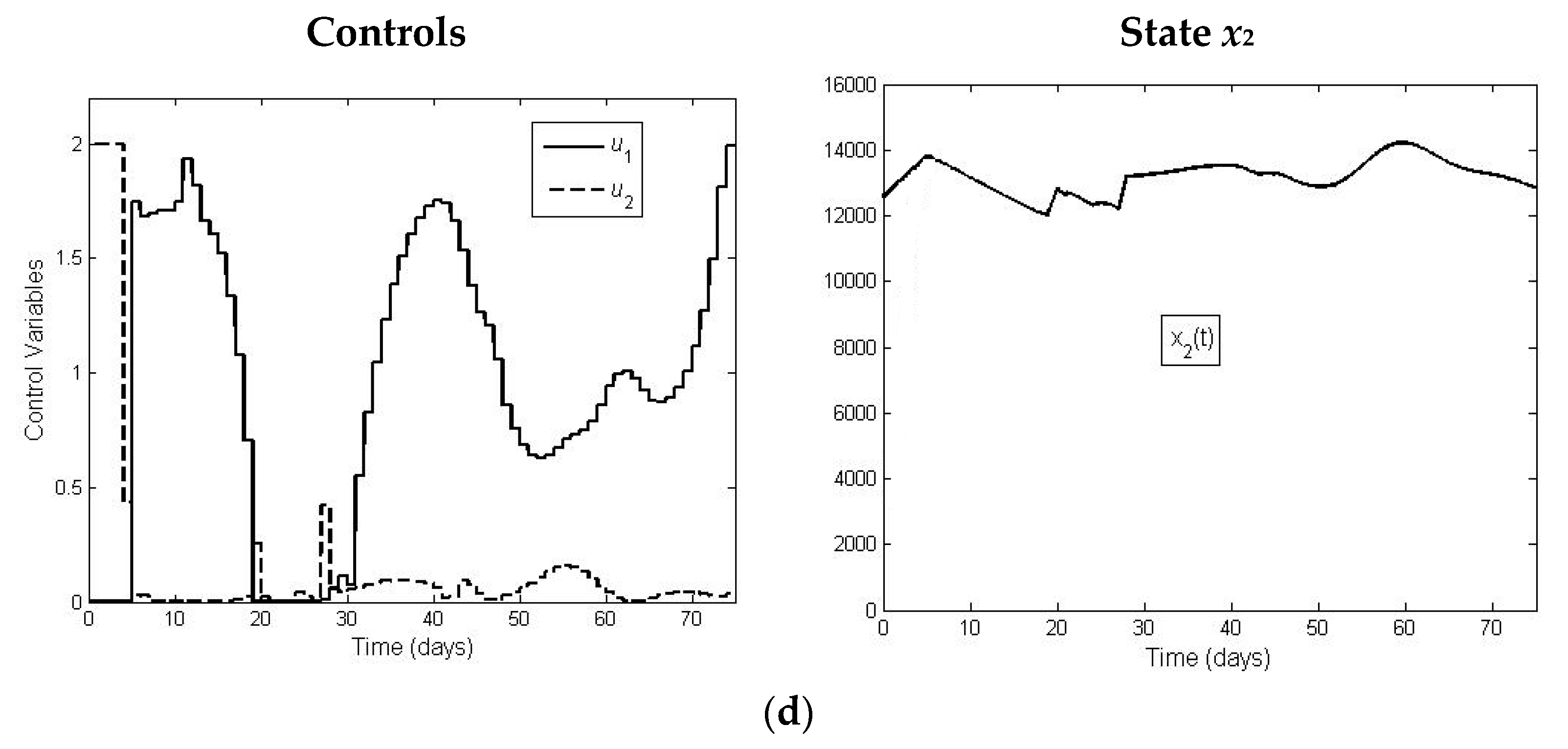
The results of our optimal control model show that by increasing initial release of juvenile, the necessary supplementary food can be reduced, implying the decrease in costs. Concerning the control model, we also notice that for the application of the corresponding toolbox, we had to apply piecewise constant control functions, changing the added food on daily basis (see
Figure 5). If for technical convenience we want to intervene less frequently, we can apply a less fine model, applying food supply with longer time intervals of constancy for the control functions.
4.2. Discussion of Theoretical Issues
In this paper, we start out from a practical problem of beneficial insects in greenhouses, in particular, insect pollinators. Observe that for the presented conceptual model, a combination of different mathematical methods was necessary. We emphasize, we use Markov model to handle the general pollination process. After the mathematical study of our model, we were also surprised that when we used some data on hoverfly from the literature [
43], the prediction obtained from the Markov model matched surprisingly well with the density suggested by the agent producing company. That is why we rely on Markov processes as a good approximation in this context. Moreover, the investigation of our practical problem can raise some interesting questions from the point of view of theoretical population ecology, as well. Namely, in our population dynamic model, we have to take account the fact that the density does depend not only on the carrying capacity but also on the life span of the insect.
The theoretical foundations of applied sciences using mathematical models have the following advantages:
First advantage: during the model building, the modeler has to list the assumptions of the model in question. These assumptions can be considered as starting hypotheses for the corresponding experiments and are either validated or confuted by the experiments. In the latter case, we need a new model, and so on. For instance, in our model, we formulated concrete biological hypotheses that can be tested by trials also appropriate to estimate the model parameters, including those of the feeding functions (see, (a–c)) functions in
Section 2.4). With these trials, we may also point out whether certain changes in the model are necessary. In this way, from the original
conceptual model, we will obtain an
executable model. The final result of our project should be the optimal control of the economically successful pollination by appropriate maintenance of the pollinator population.
Moreover, from the modelling point of view, the efficiency of adding food to enhance pollination of plants, as a matter of fact, depends on the optimal foraging of the pollinator. On the one hand, one of the well-known results of optimal foraging theory is the zero-one rule which claims that: If the more valuable food is abundant enough then the foragers accept only this type, while if the more valuable food is not abundant enough, the foragers accept both types of food [
45]. Therefore, if the added abundant food is more attractive for the pollinator than the plant flowers, then the additional food can decrease the economically success of pollination. Another issue is that we have no information on the searching and handling times of the additional food. In order to build up a well-detailed density-dependent optimal foraging model and dynamics, these data would also be a prerequisite, since the possible optimal foraging strategy-dependent functional response needs these parameters [
46]. On the other hand, if the crop plants offer no sufficient food for the pollinator, then a small quantity of added food (e.g., some vitamin) can increase the pollinator density, so Liebig’s law of the minimum may play an important role (see e.g., [
47]). Obviously, the numerical response of pollinators will depend, at the same time, on the nectar and pollen produced by the plant, and on the additional food. Clearly, the numerical response can be measured by experiment.
Second advantage: when we have a model with the right assumptions, then the model parameters will be estimated by trials (e.g., searching and handling times, all feeding functions in our case). We emphasize these experiments can be carried out in laboratory, which are less expensive than field experiments.
Third advantage: a theoretical model, with appropriate assumptions and estimated parameters, can also give some hints to the design of field experiments, as a result, the number of field experiments, and hence their costs can be reduced.
Furthermore, the present conceptual model raises not only experimental problems but also theoretical ones. For example, we have shown how pollination of flowers by insects can be described by appropriate stochastic models. In the future, this kind of model can predict how many flowers are visited by the pollinator only once, twice, and so on. The number of visits has an effect on the successful pollination (e.g., [
48]). Furthermore, since the nectar of a flower can be depleted by the insect, the stochastic model can predict the nectar and pollen collected by the insect, so this kind of model provides the numerical response of the insect, as well.
Moreover, appropriate deterministic population models can predict the density dynamics of the pollinator. We note that in order to maintain the desired pollination level, an option could have been a control model for tracking the constant desired value of adult hoverfly as reference trajectory. In our optimal control model instead, dealing with an inequality constraint for the adult hoverfly density, the minimization of the feeding costs also keeps the adult density near (and above) the required level.
Of course, more realistic population dynamics may also consider the optimal foraging behavior of the pollinator (when the searching and handling times are measured in a trial). Furthermore, the optimal control model used here can calculate the optimal regime of added food for more complicated population dynamics, as well.
According to the results found for the mathematical model developed even without the support of an experimental part, first, we can calculate the hypothetical density of adult hover flies needed for a successful pollination of tomato crop and second, the deterministic control model allows establishing the strategy of supplementary food supply for an optimal achievement of pollination of the crop. Of course, for different parameter values, the result of the model calculation would be different. However, at the present stage, we only wanted to show how the optimal strategy can be calculated, and not the concrete illustrative numerical result is the achievement. We can take into account, on the one hand, the need to carry out experimental trials that allow us to validate the two stages of the models and achieve a better adjustment to the real pollination data in tomato crops in greenhouses. On the other hand, these experimental trials should also be extended to the other 7 most important crops in Mediterranean greenhouses, in addition to tomato, pepper, eggplant, cucumber, zucchini, melon, watermelon, and beans. Its relevance derives from the total area of these crops (201,000 ha), which represents 10.6% of the overall area of greenhouses worldwide [
49]. It is considered that this can be very useful as the theoretical models developed can be flexible enough to be extended to these crops. This is because not all plant species are equally good for pollinators. Some of them supply both nectar and pollen abundantly when in bloom, and these are often called honey plants, because they are best suited for honey production. Plants producing nectar but little or no pollen are also considered to be honey plants. Other plants, however, may yield pollen but little or no nectar [
50]. This can be extended to the previously described greenhouse crops [
4].

 and
and 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}