Isolation and Characterization of Plant Growth Promoting Endophytic Bacteria from Desert Plants and Their Application as Bioinoculants for Sustainable Agriculture

 , ,
, ,  , , ,
, , ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Sampling and Study Area
2.2. Isolation of Bacterial Endophytes
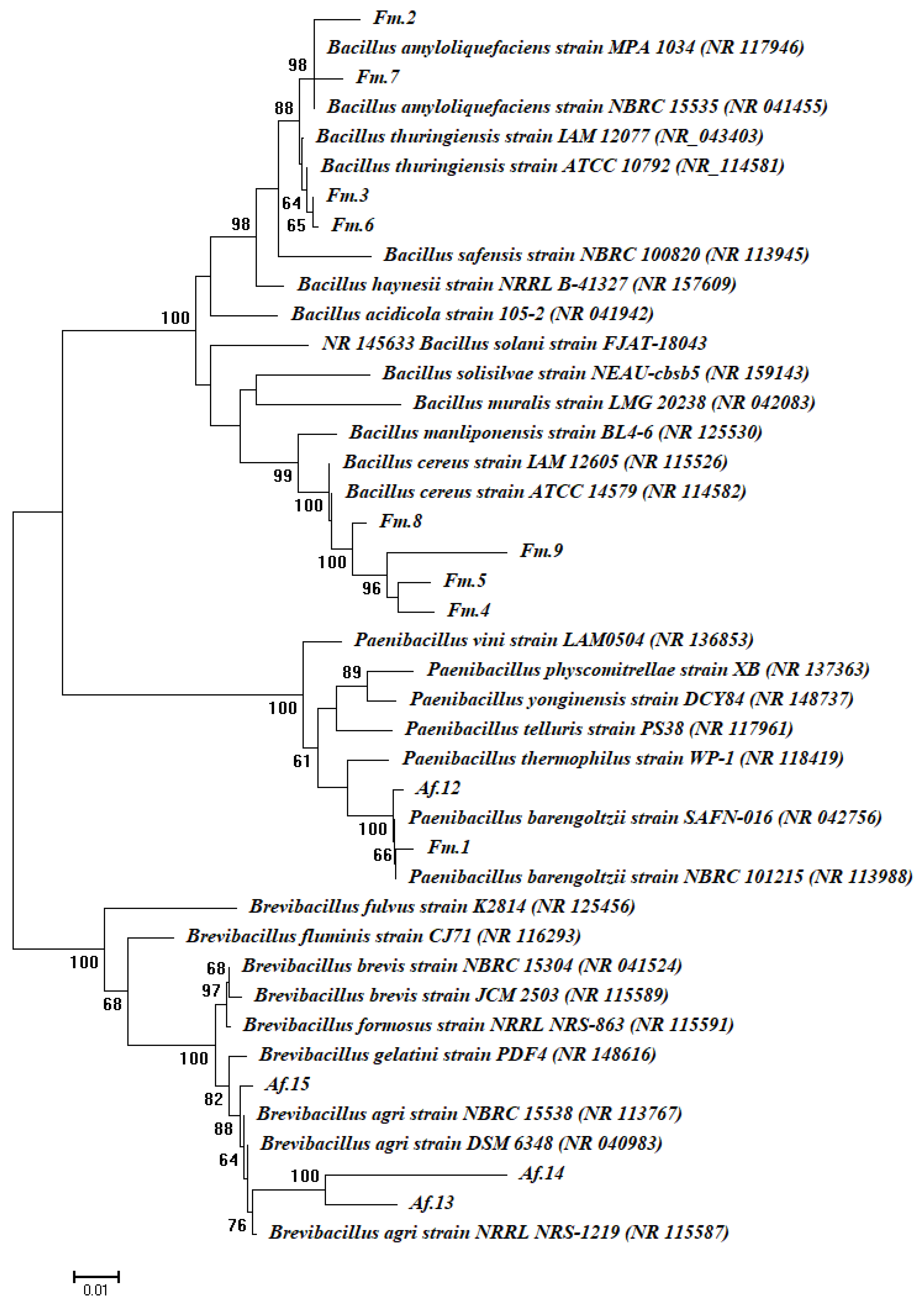
2.3. Molecular Identification of Bacterial Endophytes
2.4. Screening the Extracellular Enzymatic Activities of Bacterial Endophytes
2.5. Antimicrobial Activity of Bacterial Endophytes
2.6. Screening for In Vitro Plant Growth Promoting (PGP) Traits
2.6.1. Phosphate Solubilization
2.6.2. Ammonia Production
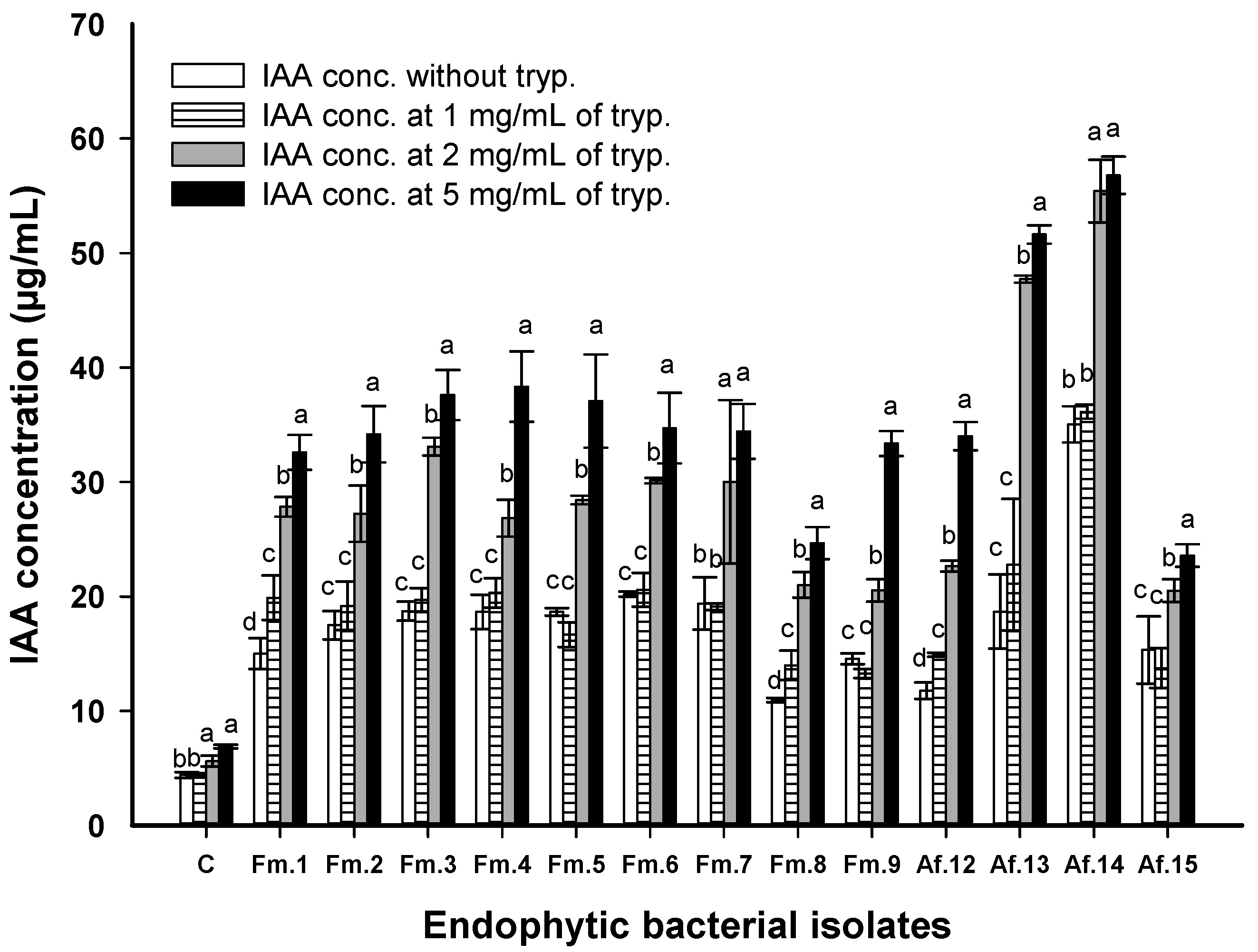
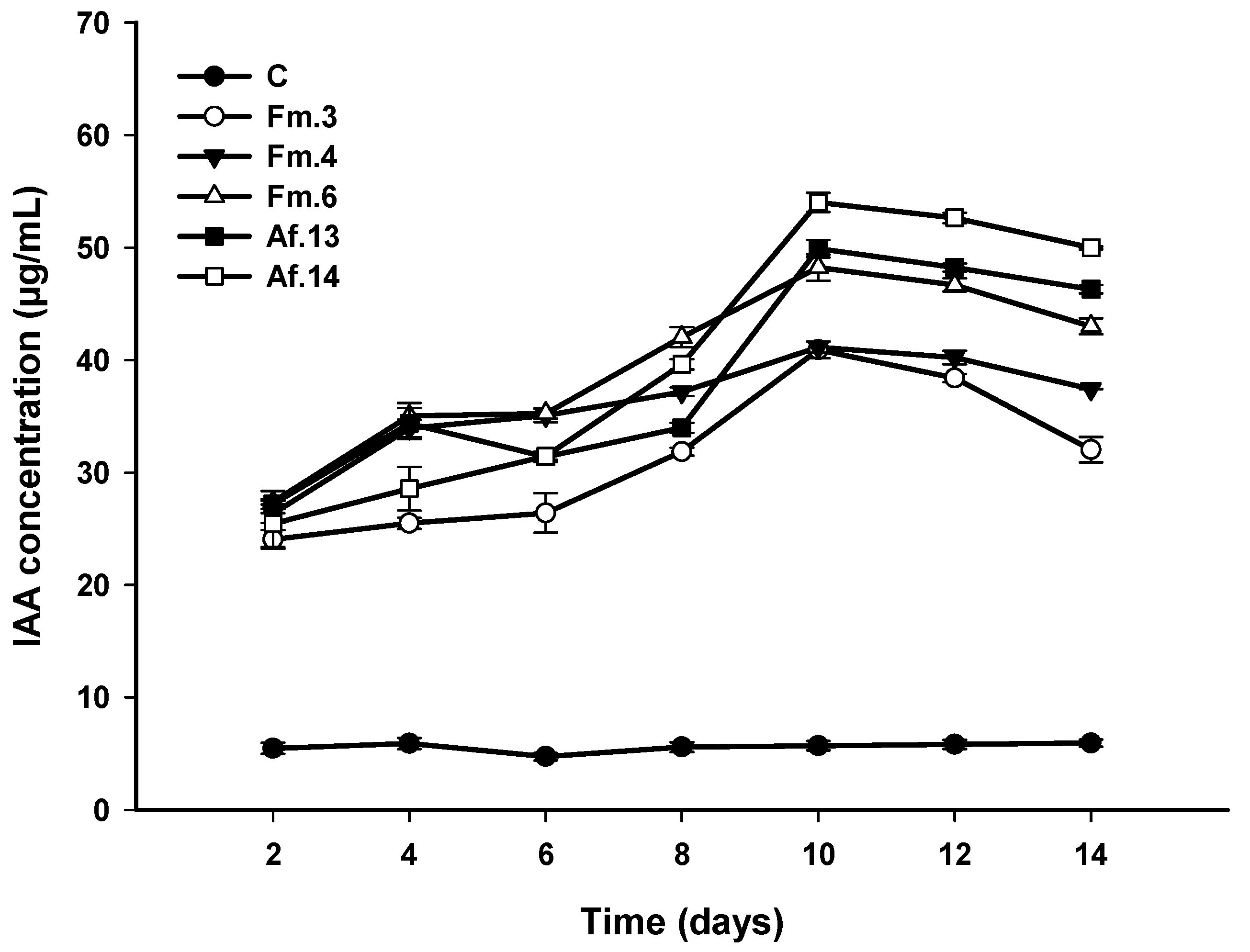
2.7. Quantitative Screening for Indole-3-acetic acid (IAA) Production
2.8. Effect of Bacterial Isolates on Zea mays L. Growth
2.8.1. Experimental Design
2.8.2. Culture Conditions
2.9. Plant Tissue Analysis
2.10. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sun, R.; Zhang, X.; Guo, X.; Wang, D.; Chu, H. Bacterial diversity in soils subjected to long-term chemical fertilization can be more stably maintained with the addition of livestock manure than wheat straw. Soil Biol. Biochem. 2015, 88, 9–18. [Google Scholar] [CrossRef]
- Zhou, J.; Jiang, X.; Zhou, B.; Zhao, B.; Ma, M.; Guan, D.; Li, J.; Chen, S.; Cao, F.; Shen, D.; et al. Thirty four years of nitrogen fertilization decreases fungal diversity and alters fungal community composition in black soil in northeast China. Soil Biol. Biochem. 2016, 95, 135–143. [Google Scholar] [CrossRef]
- Rafi, M.M.; Krishnaveni, M.S.; Charyulu, P.B.B.N. Phosphate-Solubilizing Microorganisms and Their Emerging Role in Sustainable Agriculture. In Recent Developments in Applied Microbiology and Biochemistry; Buddolla, V., Ed.; Academic Press: Cambridge, MA, USA; Elsevier: Amsterdam, The Netherlands, 2019; Volume 17, pp. 223–233. [Google Scholar] [CrossRef]
- De La, T.; Neyser, V.R.; Clara, I.R.; Martha, R.; Carlos, A.; Federico, A.G.; Héctor, P.; Reiner, R. Effect of plant growth-promoting bacteria on the growth and fructan production of Agave americana L. Brazilian J. Microbiol. 2016, 47, 587–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gamez, R.; Cardinale, M.; Montes, M.; Ramirez, S.; Schnell, S.; Rodriguez, F. Screening, plant growth promotion and root colonization pattern of two rhizobacteria (Pseudomonas fluorescens Ps006 and Bacillus amyloliquefaciens Bs006) on banana cv. Williams (Musa acuminata Colla). Microbiol. Res. 2019, 220, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Abdelaal, K.A.A.; Tawfik, S.F. Response of Sugar Beet Plant (Beta vulgaris L.) to Mineral Nitrogen Fertilization and Bio-Fertilizers. Int. J. Curr. Microbiol. Appl. Sci. 2015, 4, 677–688. [Google Scholar]
- Abdelaal, K.A.A. Pivotal Role of Bio and Mineral Fertilizer Combinations on Morphological, Anatomical and Yield Characters of Sugar Beet Plant (Beta vulgaris L.). Middle East J. Agric. Res. 2015, 4, 717–734. [Google Scholar]
- Abdelaal, K.A.A.; Badawy, S.A.; Abdel Aziz, R.M.; Neana, S.M.M. Effect of mineral nitrogen levels and PGPR on morphophysiological characters of three sweet sorghum varieties (Sorghum bicolor L. Moench). J. Plant Prod. 2015, 6, 189–203. [Google Scholar] [CrossRef]
- Hassan, S.E.; Salem, S.S.; Fouda, A.; Awad, M.A.; El-Gamal, M.S.; Abdo, A.M. New approach for antimicrobial activity and bio-control of various pathogens by biosynthesized copper nanoparticles using endophytic actinomycetes. J. Radiation Res. Appl. Sci. 2018, 11, 262–270. [Google Scholar] [CrossRef] [Green Version]
- Hassan, S.E. Plant growth-promoting activities for bacterial and fungal endophytes isolated from medicinal plant of Teucrium polium L. J. Adv. Res. 2017, 8, 687–695. [Google Scholar] [CrossRef]
- Naseem, H.; Ahsan, M.; Shahid, M.A.; Khan, N. Exopolysaccharides producing rhizobacteria and their role in plant growth and drought tolerance. J. Basic Microbiol. 2018, 58, 1009–1022. [Google Scholar] [CrossRef]
- Taktek, S.; St-Arnaud, M.; Yves Piché, J.; Fortin, A.; Antoun, H. Igneous phosphate rock solubilization by biofilm-forming mycorrhizobacteria and hyphobacteria associated with Rhizoglomus irregulare DAOM 197198. Mycorrhiza 2017, 27, 13–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbamondi, G.R.; Tommonaro, G.; Weyens, N.; Thijs, S.; Sillen, W.; Gkorezis, P.; Iodice, C.; Rangel, M.; Nicolaus, B.; Vangronsveld, J. Plant growth-promoting effects of rhizospheric and endophytic bacteria associated with different tomato cultivars and new tomato hybrids. Chem. Biol. Technol. Agric. 2016, 3, 1. [Google Scholar] [CrossRef] [Green Version]
- Hafez, Y.M.; Attia, K.A.; Kamel, S.; Alamery, S.; El-Gendy, S.; Al-Dosse, A.; Mehiar, F.; Ghazy, A.; Abdelaal, K.A.A. Bacillus subtilis as a bio-agent combined with nano molecules can control powdery mildew disease through histochemical and physiobiochemical changes in cucumber plants. Physiol. Mol. Plant Path. 2020, 111, 101489. [Google Scholar] [CrossRef]
- Hafez, Y.; Emeran, A.; Esmail, S.; Mazrou, Y.; Abdrabbo, D.; Abdelaal, K.A.A. Alternative treatments improve physiological characters, yield and tolerance of wheat plants under leaf rust disease stress. Fresenius Environ. Bull. 2020, 29, 4738–4748. [Google Scholar]
- St-Arnaud, M.; Vujanovic, V. Effect of the arbuscular mycorrhizal symbiosis on plant diseases and pests. In Mycorrhizae in Crop Production; Hamel, C., Plenchette, C., Eds.; Haworth Press: Binghampton, NY, USA, 2007; pp. 67–122. [Google Scholar]
- Hafez, Y.M.; Abdelaal, K.A.A.; Badr, M.M.; Esmaeil, R.A. Control of Puccinia triticina the causal agent of wheat leaf rust disease using safety resistance inducers correlated with endogenously antioxidant enzymes up-regulation. Egyptian J. Biol. Pest Control 2017, 27, 1–10. [Google Scholar]
- Fouda, A.; Abdel-Maksoud, G.; Abdel-Rahman, M.A.; Eid, A.M.; Barghoth, M.G.; El-Sadany, M.A. Monitoring the effect of biosynthesized nanoparticles against biodeterioration of cellulose-based materials by Aspergillus niger. Cellulose 2019, 26, 6583–6597. [Google Scholar] [CrossRef]
- Fouda, A.; Abdel-Maksoud, G.; Abdel-Rahman, M.A.; Salem, S.S.; Hassan, S.E.; El-Sadany, M.A. Eco-friendly approach utilizing green synthesized nanoparticles for paper conservation against microbes involved in biodeterioration of archaeological manuscript. Int. Biodet. Biodegr. 2019, 142, 160–169. [Google Scholar] [CrossRef]
- Fouda, A.; Hassan, S.E.; Salem, S.S.; Shaheen, T.I. In-Vitro cytotoxicity, antibacterial, and UV protection properties of the biosynthesized Zinc oxide nanoparticles for medical textile applications. Microb. Pathog. 2018, 125, 252–261. [Google Scholar] [CrossRef]
- Cherif, H.; Marasco, R.; Rolli, E.; Ferjani, R.; Fusi, M.; Soussi, A.; Mapelli, F.; Blilou, I.; Borin, S.; Boudabous, A.; et al. Oasis desert farming selects environment-specific date palm root endophytic communities and cultivable bacteria that promote resistance to drought. Environ. Microb. Rep. 2015, 7, 668–678. [Google Scholar] [CrossRef] [Green Version]
- Khan, N.; Bano, A.; Rahman, M.A.; Rathinasabapathi, B.; Babar, M.A. UPLC-HRMS-based untargeted metabolic profiling reveals changes in chickpea (Cicer arietinum) metabolome following long-term drought stress. Plant Cell Environ. 2019, 42, 115–132. [Google Scholar] [CrossRef] [Green Version]
- Köberl, M.; Müller, H.; Ramadan, E.M.; Berg, G. Desert Farming Benefits from Microbial Potential in Arid Soils and Promotes Diversity and Plant Health. PLoS ONE 2011, 6, e24452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, N.; Bano, A.; Babar, M.A. The stimulatory effects of plant growth promoting rhizobacteria and plant growth regulators on wheat physiology grown in sandy soil. Arch. Microbiol. 2019, 201, 769–785. [Google Scholar] [CrossRef] [PubMed]
- Hanna, A.L.; Youssef, H.H.; Amer, W.M.; Monib, M.; Fayez, M.; Hegazi, N.A. Diversity of bacteria nesting the plant cover of north Sinai deserts, Egypt. J. Adv. Res. 2013, 4, 13–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selim, K.A.; El-Beih, A.A.; AbdEl-Rahman, T.M.; El-Diwany, A.I. Biodiversity and antimicrobial activity of endophytes associated with Egyptian medicinal plants. Mycosphere 2011, 2, 669–678. [Google Scholar] [CrossRef]
- ALKahtani, M.D.F.; Attia, K.A.; Hafez, Y.M.; Khan, N.; Eid, A.M.; Ali, M.A.M.; Abdelaal, K.A.A. Chlorophyll Fluorescence Parameters and Antioxidant Defense System Can Display Salt Tolerance of Salt Acclimated Sweet Pepper Plants Treated with Chitosan and Plant Growth Promoting Rhizobacteria. Agronomy 2020, 10, 1180. [Google Scholar] [CrossRef]
- Kiarie, S.; Nyasani, J.O.; Gohole, L.S.; Maniania, N.K.; Subramanian, S. Impact of Fungal Endophyte Colonization of Maize (Zea mays L.) on Induced Resistance to Thrips and Aphid-Transmitted Viruses. Plants 2020, 9, 416. [Google Scholar] [CrossRef] [Green Version]
- Abd el Fatah, H.Y.; Mohamed, E.A.; Hassan, M.B.; Mohamed, K.A. An Economic Analysis for Maize Market in Egypt. Middle East J. Agric. Res. 2015, 4, 873–878. [Google Scholar]
- Berg, G.; Grube, M.; Schloter, M.; Smalla, K. Unraveling the plant microbiome: Looking back and future perspectives. Front. Microbiol. 2014, 5, 148. [Google Scholar] [CrossRef] [Green Version]
- Arora, S.; Patel, P.N.; Vanza, M.J.; Rao, G.G. Isolation and characterization of endophytic bacteria colonizing halophyte and other salt tolerance plant species from coastal Gujarta. Afr. J. Microb. Res. 2014, 8, 1779–1788. [Google Scholar]
- Miller, D.N.; Bryant, J.E.; Madsen, E.L.; Ghiorse, W.C. Evaluation and optimization of DNA extraction and purification procedures for soil and sediment samples. Appl. Environ. Microbiol. 1999, 65. [Google Scholar] [CrossRef] [Green Version]
- Lane, D.J. 16S/23S rRNA sequencing. In Nucleic Acids Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; John Wiley & Sons: Chichester, UK, 1991. [Google Scholar]
- Lv, Y.; Zhang, F.; Chen, J.; Cui, J.; Xing, Y.; Li, X.; Guo, S. Diversity and antimicrobial activity of endophytic fungi associated with the alpine plant Saussurea involucrata. Biol. Pharm. Bull. 2010, 33, 1300–1306. [Google Scholar] [CrossRef] [Green Version]
- Yadav, R.; Chauhan, N.; Chouhan, A.S.; Soni, V.K.; Omray, L. Antimicrobial screening of various extracts of Aphanmixis polystachya stems bark. Int. J. Adv. Pharm. Sci. 2010, 1, 147–150. [Google Scholar]
- Mohamed, A.A.; Fouda, A.; Abdel-Rahman, M.A.; Hassan, S.E.; El-Gamal, M.S.; Salem, S.S.; Shaheen, T.I. Fungal strain impacts the shape, bioactivity and multifunctional properties of green synthesized zinc oxide nanoparticles. Biocat. Agric. Biotech. 2019, 19, 101103. [Google Scholar] [CrossRef]
- Sharaf, O.M.; Al-Gamal, M.S.; Ibrahim, G.A.; Dabiza, N.M.; Salem, S.S.; El-ssayad, M.F.; Youssef, A.M. Evaluation and characterization of some protective culture metabolites in free and nano-chitosan-loaded forms against common contaminants of Egyptian cheese. Carbohyd. Polym. 2019, 223, 115094. [Google Scholar] [CrossRef] [PubMed]
- Alsharif, S.M.; Salem, S.S.; Abdel-Rahman, M.A.; Fouda, A.; Eid, A.M.; Hassan, S.E.; Awad, M.A.; Mohamed, A.A. Multifunctional properties of spherical silver nanoparticles fabricated by different microbial taxa. Heliyon 2020, 6, 3943. [Google Scholar] [CrossRef]
- Fouda, A.; Hassan, S.E.-D.; Abdo, A.M.; El-Gamal, M.S. Antimicrobial, Antioxidant and Larvicidal Activities of Spherical Silver Nanoparticles Synthesized by Endophytic Streptomyces spp. Biol. Trace Element Res. 2020, 195, 707–724. [Google Scholar] [CrossRef]
- Jasim, B.; John, C.; Jyothis, M.; Radhakrishnan, E.K. Plant growth promoting potential of endophytic bacteria isolated from Piper nigrum. Plant Growth Regul 2013, 71, 1–11. [Google Scholar] [CrossRef]
- Singh, P.; Kumar, V.; Agrawal, S. Evaluation of phytase producing bacteria for their plant growth promoting activities. Intern. J. Microbiol. 2014, 426483. [Google Scholar] [CrossRef] [Green Version]
- Marag, P.S.; Suman, A. Growth stage and tissue specific colonization of endophytic bacteria having plant growth promoting traits in hybrid and composite maize (Zea mays L.). Microbiol. Rese 2018, 214, 101–113. [Google Scholar] [CrossRef]
- AOAC. International Official Methods of Analysis of AOAC International. In Official Methods of Analysis of AOAC International; George, W., Ed.; Association of Official Analytical Chemists: Rockville, MD, USA, 2016; p. 3172. [Google Scholar]
- Rice, E.W.; Baird, R.B.; Eaton, A.D. Standard Methods for the Examination of Water and Wastewater, 23rd ed.; American Public Health Association, American Water Works Association: Washington, DC, USA, 2017. [Google Scholar]
- Eida, A.A.; Ziegler, M.; Lafi, F.F.; Michell, C.T.; Voolstra, C.R.; Hirt, H.; Saad, M.M. Desert plant bacteria reveal host influence and beneficial plant growth properties. PLoS ONE 2018, 13, e0208223. [Google Scholar] [CrossRef] [PubMed]
- Eid, A.M.; Salim, S.S.; Hassan, S.S.; Ismail, M.A.; Fouda, A. Role of Endophytes in Plant Health and Abiotic Stress Management. In Microbiome in Plant Health and Disease: Challenges and Opportunities; Kumar, V., Prasad, R., Kumar, M., Choudhary, D.K., Eds.; Springer: Singapore, 2019; pp. 119–144. [Google Scholar]
- Fouda, A.; Hassan, S.E.; Eid, A.M.; Ewais, E.E. The Interaction Between Plants and Bacterial Endophytes Under Salinity Stress. In Endophytes and Secondary Metabolites; Jha, S., Ed.; Springer Nature: Cham, Switzerland, 2019; pp. 591–607. [Google Scholar]
- Choi, Y.W.; Hodgkiss, I.; Hyde, K. Enzyme production by endophytes of Brucea javanica. J. Agric. Tech. 2005, 1, 55–66. [Google Scholar]
- Glick, B.R. Plant growth-promoting bacteria: Mechanisms and applications. Scientifica 2012, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro, R.A.; Quecine, M.C.; Lacava, P.T.; Batista, B.D.; Luvizotto, D.M.; Marcon, J.; Ferreira, A.; Melo, I.S.; Azevedo, J.L. Isolation and enzyme bioprospection of endophytic bacteria associated with plants of Brazilian mangrove ecosystem. SpringerPlus 2014, 3, 382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vurukonda, S.S.P.; Giovanardi, D.; Stefani, E. Plant growth promoting and biocontrol activity of Streptomyces spp. as endophytes. Intern. J. Mol. Sci. 2018, 19, 952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chow, Y.Y.; Rahman, S.; Ting, A.S.Y. Understanding colonization and proliferation potential of endophytes and pathogen in planta via plating, polymerase chain reaction, and ergosterol assay. J. Adv. Res. 2017, 8, 13–21. [Google Scholar] [CrossRef] [Green Version]
- Khan, N.; Zandi, P.; Ali, S.; Mehmood, A.; Adnan Shahid, M.; Yang, J. Impact of salicylic acid and PGPR on the drought tolerance and phytoremediation potential of Helianthus annus. Front. Microbiol. 2018, 9, 2507. [Google Scholar] [CrossRef] [Green Version]
- Asraful, I.; Md, S.; Math, R.K.; Kim, J.M.; Yun, M.G.; Cho, J.J.; Kim, E.J.; Lee, Y.H.; Yun, H.D. Effect of plant age on endophytic bacterial diversity of balloon flower (Platycodon grandiflorum) root and their antimicrobial activities. Curr. Microbiol. 2010, 61, 346–356. [Google Scholar] [CrossRef]
- Sun, L.; Lu, Z.; Bie, X.; Lu, F.; Yang, S. Isolation and characterization of a co-producer of fengycins and surfactins, endophytic Bacillus amyloliquefaciens ES-2, from Scutellaria baicalensis Georgi. World J. Microbiol. Biotechnol. 2006, 22, 1259–1266. [Google Scholar] [CrossRef]
- Canova, S.P.; Petta, T.; Reyes, L.F.; Zucchi, T.; Moraes, L.A.; Melo, I. Characterization of lipopeptides from Paenibacillus sp. (IIRAC30) suppressing Rhizoctonia solani. World J. Microbiol. Biotechnol. 2010, 26, 2241–2247. [Google Scholar] [CrossRef]
- Biasolo, G.; Kucmanski, D.A.; Salamoni, S.P.; Gardin, J.P.; Minotto, E.; Baratto, C.M. Isolation, characterization and selection of bacteria that promote plant growth in grapevines (Vitis sp.). J. Agric. Sci. 2017, 9, 184–194. [Google Scholar] [CrossRef]
- Li, X.; Geng, X.; Xie, R.; Fu, L.; Jiang, J.; Gao, L.; Sun, J. The endophytic bacteria isolated from elephant grass (Pennisetum purpureum Schumach) promote plant growth and enhance salt tolerance of hybrid Pennisetum. Biotech. Biofuels 2016, 9, 190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banik, A.; Dash, G.K.; Swain, P.; Kumar, U.; Mukhopadhyay, S.K.; Dangar, T.K. Application of rice (Oryza sativa L.) root endophytic diazotrophic Azotobacter sp. strain Avi2 (MCC 3432) can increase rice yield under green house and field condition. Microbiol. Res. 2019, 219, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Bano, A.; Curá, J.A. Role of Beneficial Microorganisms and Salicylic Acid in Improving Rainfed Agriculture and Future Food Safety. Microorganisms 2020, 8, 1018. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, A.A.; Forzani, M.V.; Soares, R.; Sibov, S.T.; Vieira, J. Isolation and selection of plant growth-promoting bacteria associated with sugarcane. Agric. Res. Tropics 2016, 46, 149–158. [Google Scholar] [CrossRef] [Green Version]
- Lin, L.; Zhengyi, L.; Chunjin, H.; Zhang, X.; Chang, S.; Yang, L.; Yangrui, L.; Qianli, A. Plant growth-promoting nitrogen-fixing Enterobacteria are in association with sugarcane plants growing in Guangxi, China. Microbes Environ. 2012, 27, 391–398. [Google Scholar] [CrossRef] [Green Version]
- Fouda, A.; Hassan, S.E.; Eid, A.M.; Ewais, E.E. Biotechnological applications of fungal endophytes associated with medicinal plant Asclepias sinaica (Bioss). Ann. Agric. Sci. 2015, 60, 95–104. [Google Scholar] [CrossRef] [Green Version]
- Hassan, S.E.; Fouda, A.; Radwan, A.A.; Salem, S.S.; Barghoth, M.G.; Awad, M.A.; Abdo, A.M.; El-Gamal, M.S. Endophytic actinomycetes Streptomyces spp mediated biosynthesis of copper oxide nanoparticles as a promising tool for biotechnological applications. JBIC 2019, 24, 377–393. [Google Scholar] [CrossRef]
- Bokhari, A.; Essack, M.; Lafi, F.F.; Andres-Barrao, C.; Jalal, R.; Alamoudi, S.; Saad, M.M. Bioprospecting desert plant Bacillus endophytic strains for their potential to enhance plant stress tolerance. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef]
- Orozco, M.; Ma del, C.; Ma del, C.R.; Bernard, R.G.; Gustavo, S. Microbiome engineering to improve biocontrol and plant growth-promoting mechanisms. Microbiol. Res. 2018, 208, 25–31. [Google Scholar] [CrossRef]
- Khan, N.; Bano, A.; Babar, M.A. The root growth of wheat plants, the water conservation and fertility status of sandy soils influenced by plant growth promoting rhizobacteria. Symbiosis 2017, 72, 195–205. [Google Scholar] [CrossRef]
- Gyaneshwar, P.; Naresh, K.G.; Parekh, L.J.; Poole, P.S. Role of soil microorganisms in improving P nutrition of plants. Plant Soil 2002, 245, 83–93. [Google Scholar] [CrossRef]
- Hijri, M. Analysis of a large dataset of mycorrhiza inoculation field trials on potato shows highly significant increases in yield. Mycorrhiza 2016, 26, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Read, A.; Louise, F.; Taylor, H. The Ecology of Genetically Diverse Infections. Science 2001, 292, 1099. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Soil Analysis |
|---|---|
| Soil Texture | Loamy Sand |
| Physical characters (%) | |
| Sand | 76.8 |
| Silt | 10.9 |
| Clay | 12.2 |
| Chemical characters (mg kg−1) | |
| P | 24 |
| K | 14.075 |
| Na | 186.44 |
| Ca | 27.25 |
| Cl | 134.35 |
| Plant Species | Bacterial Strain Code | Homologue Sequences (Sequence Identity %) | NCBI Accession Numbers |
|---|---|---|---|
| Fagonia mollis | Fm.1 | Paenibacillus barengoltzii (99%) | NR_042756 |
| Fm.2 | Bacillus amyloliquefaciens (98%) | NR_117946 | |
| Fm.3 | Bacillus thuringiensis (97%) | NR_043403 | |
| Fm.4 | Bacillus cereus (98%) | NR_115526 | |
| Fm.5 | Bacillus cereus (99%) | NR_115526 | |
| Fm.6 | Bacillus thuringiensis (98%) | NR_114581 | |
| Fm.7 | Bacillus amyloliquefaciens (97%) | NR_117946 | |
| Fm.8 | Bacillus cereus (97%) | NR_115526 | |
| Fm.9 | Bacillus cereus (96%) | NR_115526 | |
| Achillea fragrantissima | Af.12 | Paenibacillus barengoltzii (99%) | NR_113988 |
| Af.13 | Brevibacillus agri (95%) | NR_113767 | |
| Af.14 | Brevibacillus agri (93%) | NR_113767 | |
| Af.15 | Brevibacillus agri (99%) | NR_113767 |
| Bacterial Strains 1 | Diameter of Clear Zones (mm) 2 | |||||
|---|---|---|---|---|---|---|
| Amylase | Pectinase | CMCase 3 | Cellulase | Xylanase | Gelatinase | |
| C | 0 d | 0 d | 0 e | 0 d | 0 f | 0 f |
| Fm.1 | 17.6 ± 1.2 a | 16.3 ± 0.3 b | 18.6 ± 0.6 b | 20.0 ± 1.7 b | 17.0 ± 1.0 b | 21.0 ± 0.5 a |
| Fm.2 | 17.3 ± 0.8 a | 17.6 ± 0.6 a | 20.6 ± 0.6 a | 18.6 ± 0.8 c | 19.6 ± 0.3 a | 22.0 ± 0.5 a |
| Fm.3 | 17.0 ± 1.5 a | 17.0 ± 0.5 a | 18.0 ± 1.0 b | 18.6 ± 1.2 c | 16.0 ± 00.0 b | 22.3 ± 1.4 a |
| Fm.4 | 17.6 ± 0.8 a | 17.0 ± 0.5 a | 19.0 ± 0.5 b | 19.6 ± 1.7 b | 14.6 ± 0.3 c | 20.6 ± 0.6 b |
| Fm.5 | 17.6 ± 0.8 a | 16.3 ± 0.8 b | 21.3 ± 1.2 a | 22.0 ± 1.1 a | 18.6 ± 0.3 a | 21.6 ± 0.8 a |
| Fm.6 | 17.0 ± 1.1 a | 15.6 ± 0.3 c | 18.3 ± 0.8 b | 16.3 ± 0.3d | 16.3 ± 0.8b | 17.0 ± 1.5 d |
| Fm.7 | 17.3 ± 0.6 a | 17.0 ± 0.5 a | 15.6 ± 0.8 c | 16.6 ± 1.2 d | 14.3 ± 0.6 c | 15.0 ± 1.5 e |
| Fm.8 | 17.6 ± 0.6 a | 14.6 ± 0.3 c | 18.0 ± 1.15 b | 18.3 ± 0.3 c | 14.3 ± 0.6 c | 18.3 ± 1.6 c |
| Fm.9 | 15.3 ± 1.7 b | 15.6 ± 0.3 c | 18.6 ± 0.3 b | 17.6 ± 0.8 c | 9.0 ± 1.1 e | 18.0 ± 1.5 c |
| Af.12 | 9.3 ± 0.3 c | 0 d | 19.3 ± 0.3 b | 20.6 ± 0.6 b | 13.6 ± 0.3 d | 0 f |
| Af.13 | 0 d | 0 d | 0 e | 0 d | 0 f | 19.0 ± 1.5 c |
| Af.14 | 0 d | 14.0 ± 0.2 d | 0 e | 0 d | 0 f | 0 f |
| Af.15 | 0 d | 17.6 ± 0.3 a | 11.0 ± 1.0 d | 17.33 ± 1.76 c | 0 f | 20.3 ± 2.3 b |
| Bacterial Strains 1 | Diameter of Clear Zone (mm) | |||||
|---|---|---|---|---|---|---|
| P. aeruginosa | S. typhi | E. coli | S. aureus | B. subtilis | C. albicans | |
| C | − | − | − | − | − | − |
| Fm.1 | 17 | − | − | − | − | − |
| Fm.2 | 15 | − | − | − | − | 15 |
| Fm.3 | 15 | − | − | − | − | − |
| Fm.4 | 15 | − | − | − | − | − |
| Fm.5 | 17 | − | − | − | − | − |
| Fm.6 | 20 | − | − | − | − | − |
| Fm.7 | 22 | − | 18 | − | − | − |
| Fm.8 | 30 | 15 | 18 | − | − | − |
| Fm.9 | 25 | − | 20 | − | − | − |
| Af.12 | 15 | − | − | − | − | − |
| Af.13 | 15 | 11 | 11 | − | 17 | 18 |
| Af.14 | 20 | − | − | − | − | − |
| Af.15 | 15 | − | − | − | − | − |
| Bacterial Strains 1 | Ammonia Production 2 | P solubilization Diameter of Clear Zone (mm) 3 |
|---|---|---|
| C | - | 0 d |
| Fm.1 | − | 0 d |
| Fm.2 | + | 8.6 ± 0.3 b |
| Fm.3 | ++ | 9.0 ± 0 a |
| Fm.4 | ‒ | 7.6 ± 0.3 c |
| Fm.5 | ++ | 9.6 ± 0.3 a |
| Fm.6 | + | 8.6 ± 0.3 b |
| Fm.7 | + | 8.3 ± 0.3 b |
| Fm.8 | ‒ | 9.3 ± 0.3 a |
| Fm.9 | ++ | 9.3 ± 0.3 a |
| Af.12 | ++ | 9.3 ± 0.3 a |
| Af.13 | ++ | 0 d |
| Af.14 | ++ | 0 d |
| Af.15 | ++ | 0 d |
| Bacterial Strains 1 | Dry Weight (mg) 2 | Shoot Nutrients Content (mg) | |||
|---|---|---|---|---|---|
| Shoot | Root | P | N | K | |
| C | 82 ± 3.56 c | 252.8 ± 19.5 a | 0.42 ± 0.01 c | 2.2 ± 0.17 c | 8.70 ± 0.03 b |
| Fm.3 | 110.3 ± 5.5 a | 322.64 ± 16.0 a | 1.05 ± 0.07 a | 3.9 ± 0.19 b | 11.35 ± 0.92 a |
| Fm.4 | 81.8 ± 4 c | 315.8 ± 27.9 a | 0.72 ± 0.03 b | 3.1 ± 0.60 bc | 8.80 ± 0.62 b |
| Fm.6 | 103.2 ± 1.8 a | 295.66 ± 30.1 a | 0.76 ± 0.02 b | 4.4 ± 0.18 b | 11.11 ± 0.31 a |
| Af.13 | 108.2 ± 1.05 a | 286.86 ± 19.3 a | 0.44 ± 0.01 c | 7.1 ± 0.18 a | 10.57 ± 0.60 ab |
| Af.14 | 90.78 ± 4.55 b | 303.8 ± 27.3 a | 0.40 ± 0.02 c | 7.2 ± 0.43 a | 9.97 ± 0.13 ab |
| Mix | 95.2 ± 2.8 b | 249.6 ± 19.7 a | 0.40 ± 0.01 c | 3.4 ± 0.27 bc | 10.45 ± 0.36 ab |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
ALKahtani, M.D.F.; Fouda, A.; Attia, K.A.; Al-Otaibi, F.; Eid, A.M.; Ewais, E.E.-D.; Hijri, M.; St-Arnaud, M.; Hassan, S.E.-D.; Khan, N.; et al. Isolation and Characterization of Plant Growth Promoting Endophytic Bacteria from Desert Plants and Their Application as Bioinoculants for Sustainable Agriculture. Agronomy 2020, 10, 1325. https://doi.org/10.3390/agronomy10091325
ALKahtani MDF, Fouda A, Attia KA, Al-Otaibi F, Eid AM, Ewais EE-D, Hijri M, St-Arnaud M, Hassan SE-D, Khan N, et al. Isolation and Characterization of Plant Growth Promoting Endophytic Bacteria from Desert Plants and Their Application as Bioinoculants for Sustainable Agriculture. Agronomy. 2020; 10(9):1325. https://doi.org/10.3390/agronomy10091325
Chicago/Turabian StyleALKahtani, Muneera D. F., Amr Fouda, Kotb A. Attia, Fahad Al-Otaibi, Ahmed M. Eid, Emad El-Din Ewais, Mohamed Hijri, Marc St-Arnaud, Saad El-Din Hassan, Naeem Khan, and et al. 2020. "Isolation and Characterization of Plant Growth Promoting Endophytic Bacteria from Desert Plants and Their Application as Bioinoculants for Sustainable Agriculture" Agronomy 10, no. 9: 1325. https://doi.org/10.3390/agronomy10091325