
Insight into Degrading Effects of Two Fungi on Polyurethane Coating Failure in a Simulated Atmospheric Environment
Abstract
:1. Introduction
2. Experimental Section
2.1. T. funiculosus and P. chrysosporium Spore Suspension
2.2. Growth Curve
2.3. Analysis of Organic Acid Metabolites
2.4. PU Coating Preparation
2.5. Fungi Treatment Process
2.6. EIS Test
3. Results and Discussion
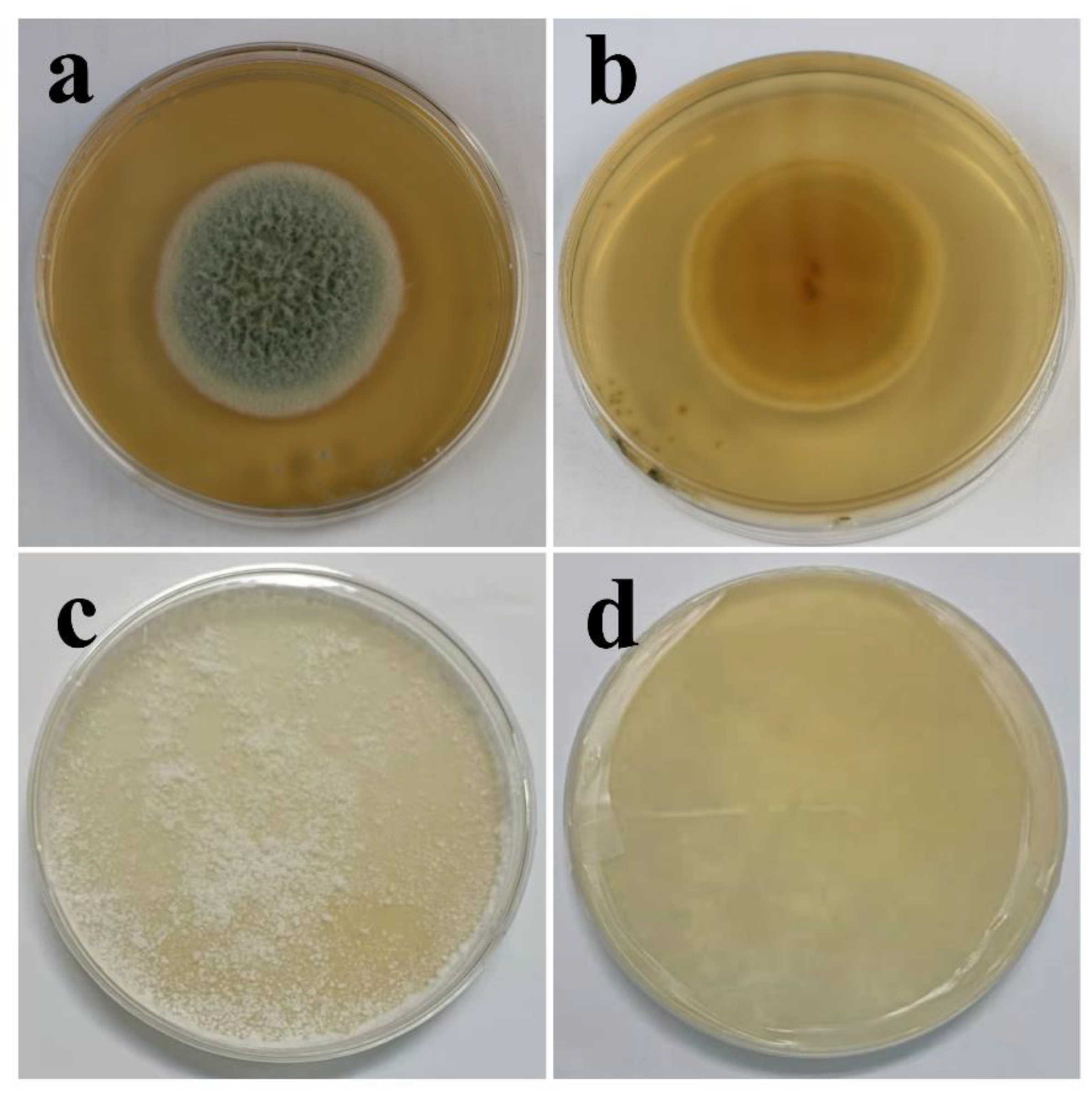
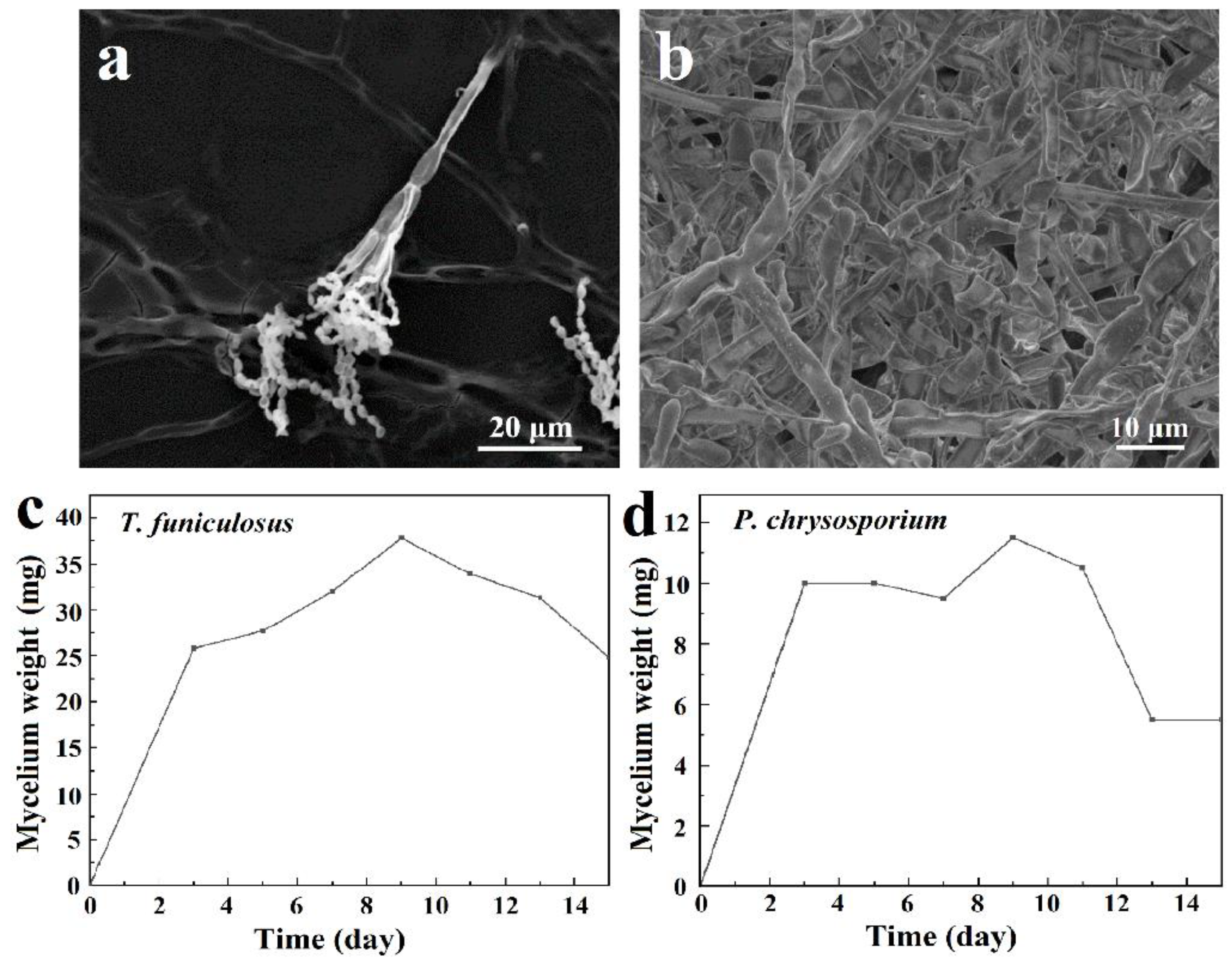
3.1. Morphologies and Growth Curve of T. funiculosus and P. chrysosporium
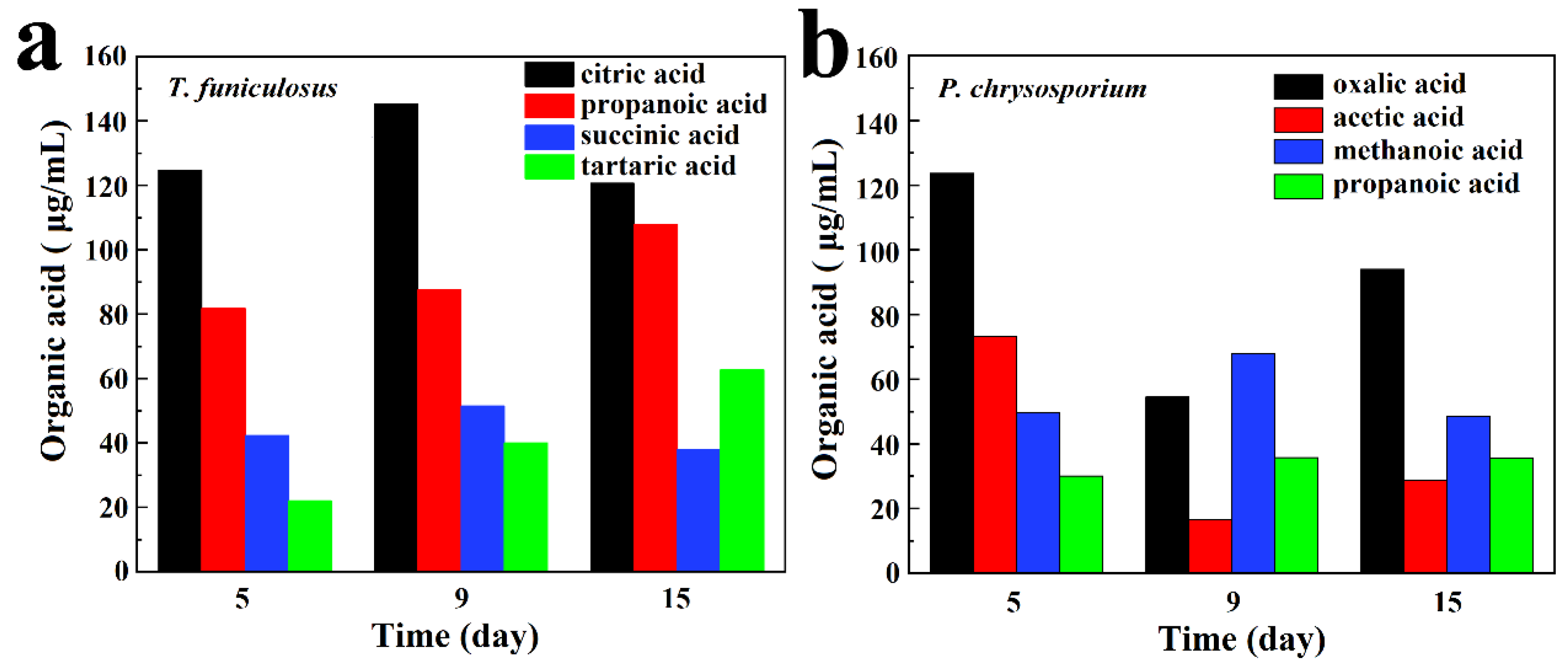
3.2. Analysis of Organic Acids Metabolites
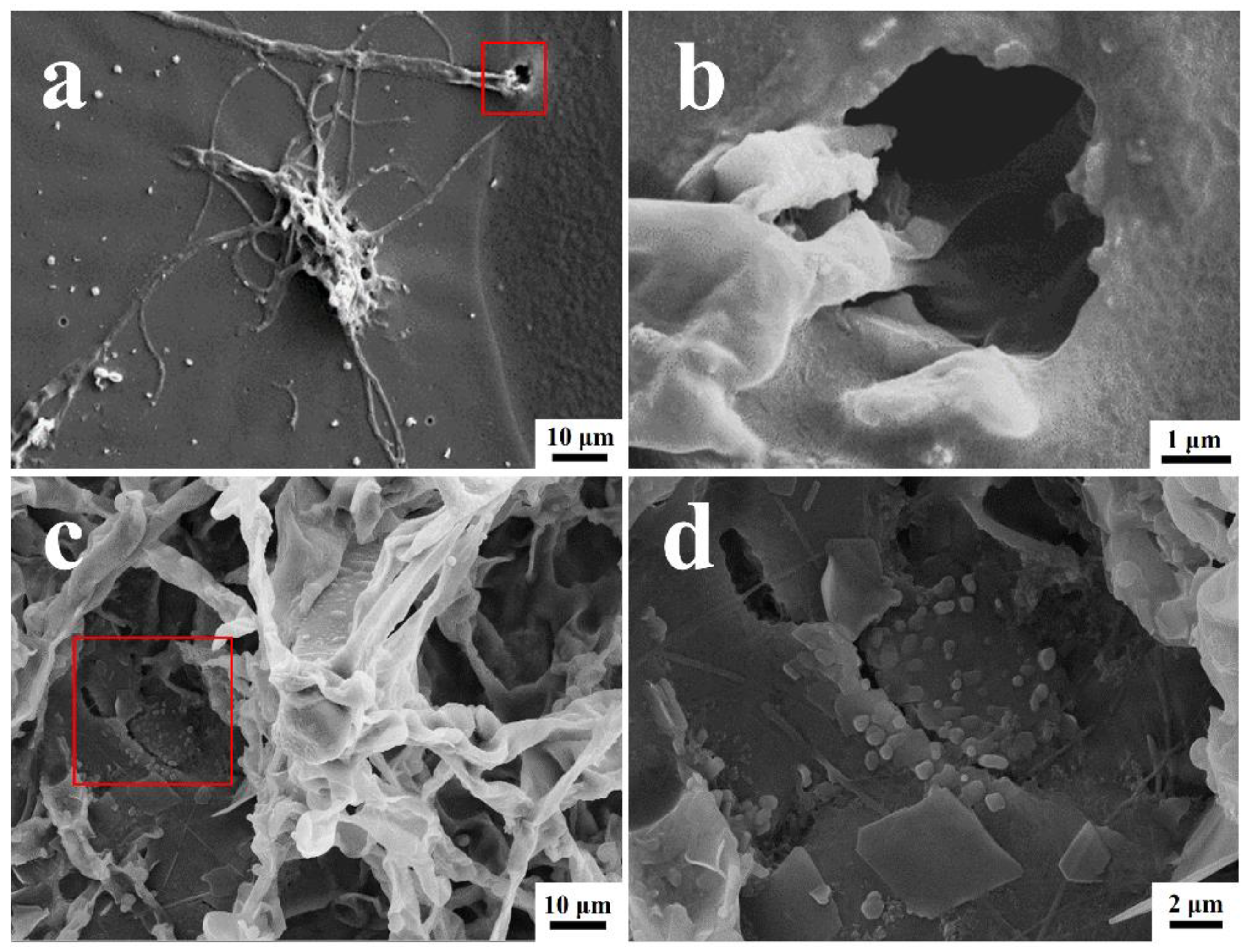
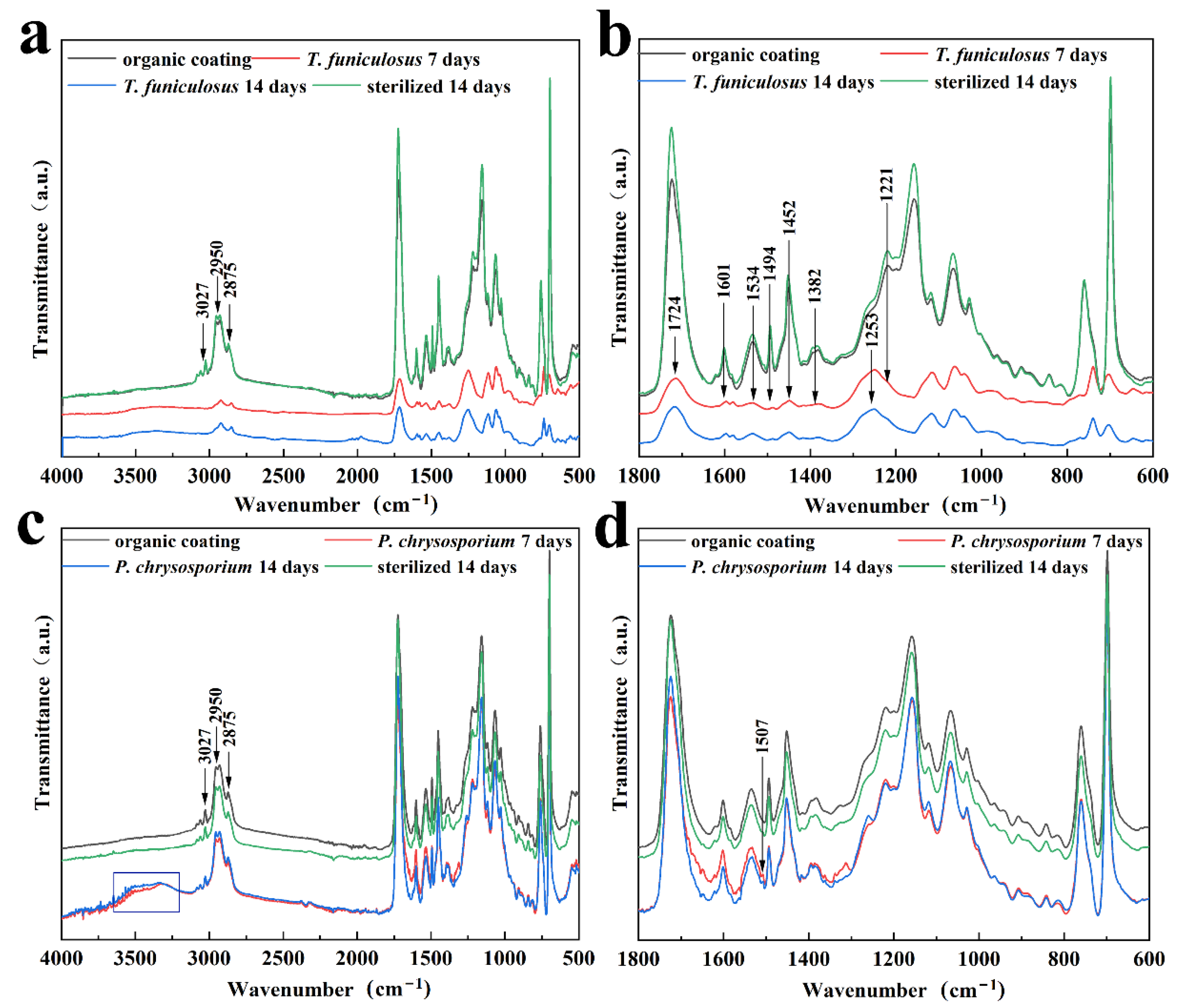
3.3. Morphological Analysis of Coating Surfaces
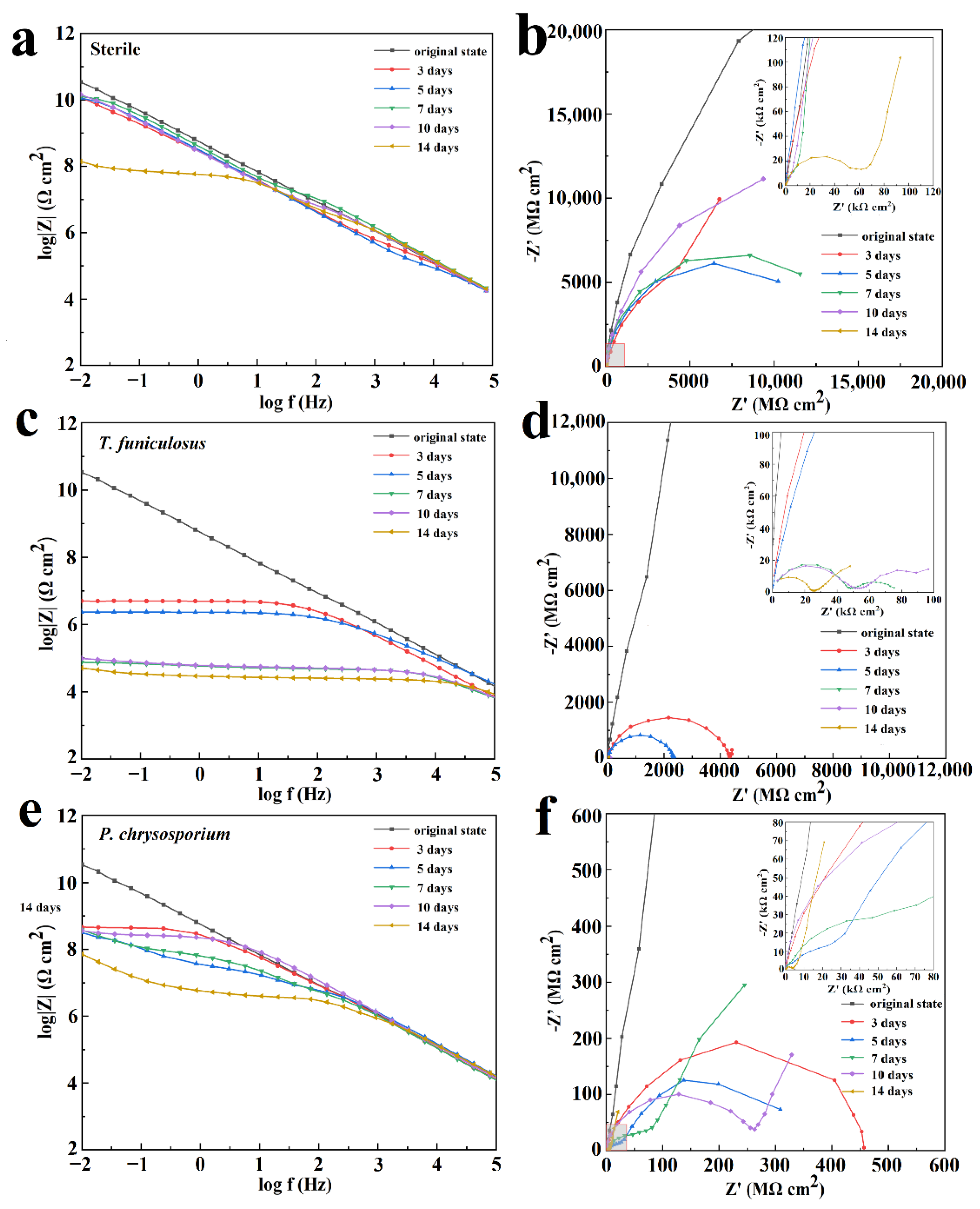
3.4. EIS Analysis
3.5. Features of Coating Failure by Two Fungi
3.5.1. Different Behaviors of Two Fungi
- (1)
- The mycelium stage of fungi and their quantity are determinants of their attack capability on the coating. The maximum mycelium weight of T. funiculosus and P. chrysosporium was about 37.8 mg and 11.5 mg, respectively, which means that the former possesses stronger abilities to obtain nutrients from coatings as well as to destroy the coating than the latter in atmospheric environments. Generally, the impact of fungi on the coating failure is proportional to their concentration, especially when PU is the only available carbon resource to support fungi metabolism in an atmospheric environment [36]. Moreover, better-developed mycelia could secret more organic acids, which was demonstrated by the higher total concentration of secreted organic acids by T. funiculosus.
- (2)
- The types and total amount of organic acids secreted by T. funiculosus and P. chrysosporium were the key factors in the biodegradation of the PU coatings. The major four organic acids secreted by T. funiculosus were carboxylic acids, which can accelerate the hydrolysis of ester and urethane bonds. Comparatively, oxalic acid was the major organic acid secreted by P. chrysosporium, which was toxic to its metabolism, thus inhibiting or even decreasing the growth and reproduction of fungi during long incubation periods. In addition, ester and urethane bonds were barely affected, indicating that this fungus generated less damage to the chemical structure of the coating. Interestingly, the total amount of organic acids secreted by T. funiculosus increased constantly during the 2 weeks incubation time instead of remaining constant as did P. chrysosporium, which led to severe degradation of the PU coatings by T. funiculosus.
- (3)
- The corrosion processes induced by these two fungi were different. According to the EIS results, the barrier properties of the coatings worsened after colonization by T. funiculosus than by P. chrysosporium, although the corrosion processes eventually occurred on the metal surface in both experimental groups. The cross-linked structures were destroyed by T. funiculosus colonization, decreasing the anti-corrosion strength of the coating. Further, the coatings were not able to resist the mycelium attack leading to perforations of the coating. Hence, the coatings’ failure not only occurs on the coatings’ surface caused by secreted organic acids but also in the deeper coatings caused by the mycelium attack. Moreover, the mycelium penetrating the coating can transport organic acids to the metal-coating interface and accelerate the process of corrosion. Hence, the EIS results showed that |Z|0.01Hz decreased continuously, and the Warburg impedance was not present during the 14 days of treatment with T. funiculosus. However, after colonization by P. chrysosporium, the coating developed cracks and sank because atmospheric moisture permeated into the coatings and formed alkaline blisters [26], although the molecular structure inside the coating was intact. The |Z|0.01Hz value remained stable from day 3 days to day 10 of colonization, and the Warburg impedance appeared, indicating that the corrosion products were accumulated under the coatings. During this process, although corrosion occurred at the metal-coating interface, the accumulated corrosion products inhibited the corrosion process of the metal substrate.
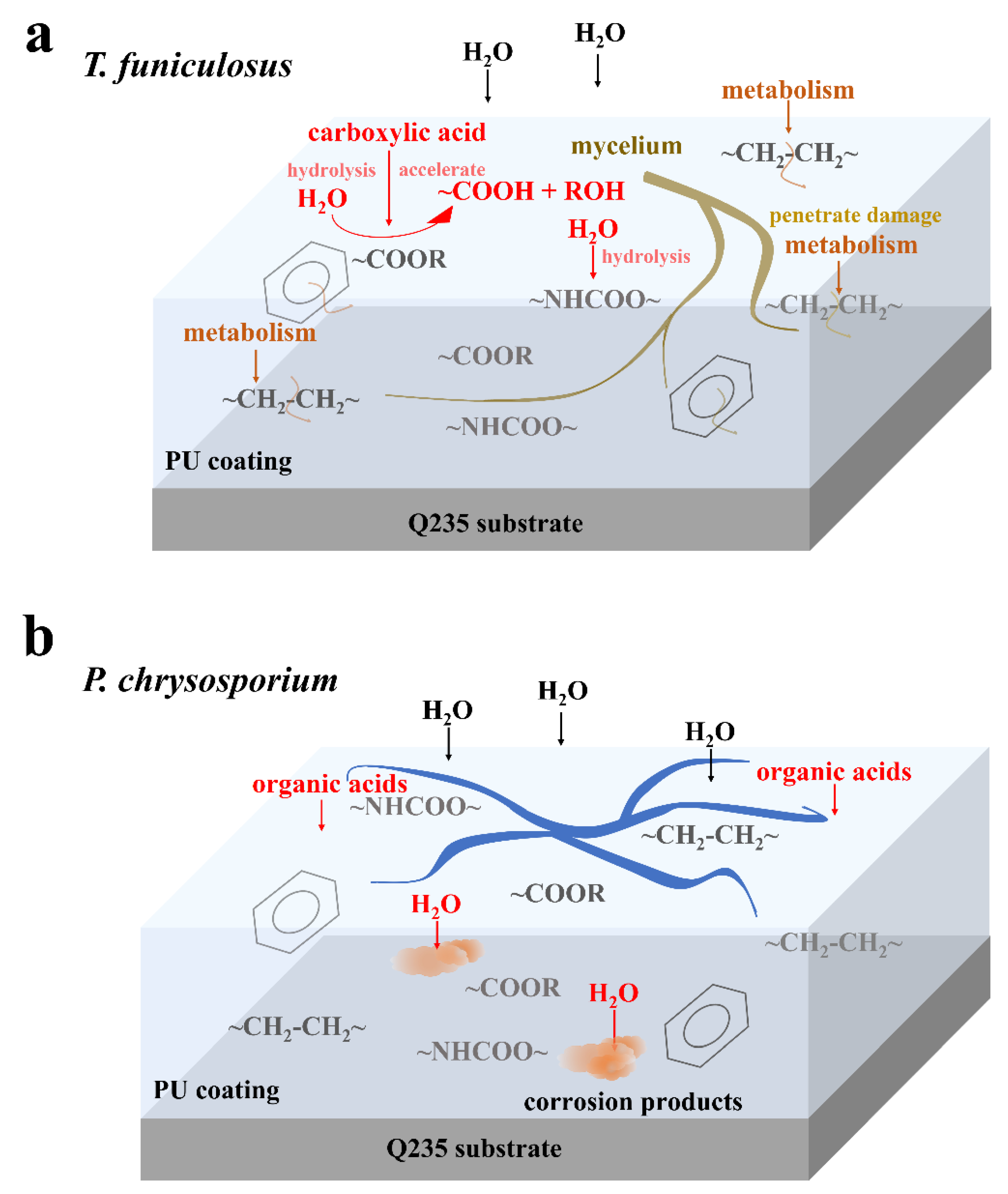
3.5.2. Coating Failure Processes Associated with Two Fungi
4. Conclusions
- (1)
- During the colonization of T. funiculosus, the mycelium penetrated the interface through microholes, while only depressions and cracks were observed in the P. chrysosporium colonized area.
- (2)
- The total concentration of metabolic organic acid secreted by T. funiculosus and P. chrysosporium was about 412.50 µg/mL and 346.34 µg/mL, respectively. Citric acid, propanoic acid, succinic acid, and tartaric acid had the highest content in the metabolic organic acids secreted by T. funiculosus, while P. chrysosporium secreted oxalic acid, acetic acid, methanolic acid, and propanoic acid.
- (3)
- The carboxylic acids secreted by T. funiculosus accelerated the hydrolysis of the ester and urethane bonds and damaged the carbon chain and benzene rings of the PU coating skeleton. Comparatively, acids secreted by P. chrysosporium had less impact on the degradation of the coating.
- (4)
- The PU coatings almost lost the barrier properties after colonization by T. funiculosus for 14 days with the |Z|0.01Hz value reaching about 5.1 × 104 Ω·cm2. However, their corrosion inhibition properties were not affected by the colonization of P. chrysosporium for the same time and the |Z|0.01Hz value observed was 7.2 × 107 Ω·cm2.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hou, B.; Li, X.; Ma, X.; Du, C.; Zhang, D.; Zheng, M.; Xu, W.; Lu, D.; Ma, F. The cost of corrosion in China. NPJ Mat. Degrad. 2017, 1, 4. [Google Scholar] [CrossRef]
- Xavier, J. Electrochemical and dynamic mechanical properties of polyurethane nanocomposite reinforced with functionalized TiO2–ZrO2 nanoparticles in automobile industry. Appl. Nanosci. 2022, 12, 1763–1778. [Google Scholar] [CrossRef]
- Ahmed, M.; Chi, Y.T.; Hung, Y.; Reyes, L.; Yeh, J. UV-cured electroactive polyurethane acrylate coatings with superhydrophobic surface structure of biomimetic peacock feather for anticorrosion application. Prog. Org. Coat. 2022, 165, 106679. [Google Scholar] [CrossRef]
- Shen, Y.; Wu, Z.; Tao, J.; Jia, Z.; Chen, H.; Liu, S.; Jiang, J.; Wang, Z. Spraying preparation of eco-friendly superhydrophobic coatings with ultralow water adhesion for effective anticorrosion and antipollution. ACS Appl. Mater. Interfaces 2020, 12, 25484–25493. [Google Scholar] [CrossRef]
- Wang, H.; Li, X.; Ji, Y.; Xu, J.; Ye, Z.; Wang, S.; Du, X. Highly transparent, mechanical, and self-adhesive zwitterionic conductive hydrogels with polyurethane as a cross-linker for wireless strain sensors. J. Mater. Chem. B 2022, 10, 2933–2943. [Google Scholar] [CrossRef]
- Zhang, W.Y.; Gao, W.; Lin, F. Preparation of solvent-free moisture curable heavy anti corrosion PU coating. Trans. Tech. Publ. Ltd. 2014, 898, 80–83. [Google Scholar] [CrossRef]
- Dacuan, C.N.; Abellana, V.Y.; Canseco, H.; Cañete, M.Q. Hydrophobic polyurethane coating against corrosion of reinforced concrete structures exposed to marine environment. Civil Eng. Archit. 2021, 9, 721–736. [Google Scholar] [CrossRef]
- Geun, U.J. Development of Highly Anti-Corrosive and Mechanically Improved Graphene-Based Polyurethane Nanocomposites for the Application of Pipeliners in Oil Sands Transportation, Canada. 2020. Available online: https://uwspace.uwaterloo.ca/handle/10012/15661 (accessed on 20 February 2020).
- Jothi, V.; Adesina, A.Y.; Rahman, M.M.; Kumar, A.M.; Ram, J.S.N. Performance, Improved adhesion and corrosion resistant performance of polyurethane coatings on anodized Mg alloy for aerospace applications. J. Mater. Eng. Perform. 2020, 29, 2586–2596. [Google Scholar] [CrossRef]
- Nadeau, L.J.; Barlow, D.E.; Hung, C.S.; Biffinger, J.C.; Crouch, A.L.; Hollomon, J.L.; Ecker, C.D.; Russell, J.N., Jr.; Crookes-Goodson, W.J. Colonization and degradation of polyurethane coatings by Pseudomonas protegens biofilms is promoted by PueA and PueB hydrolases. Int. Biodeter. Biodegr. 2021, 156, 105121. [Google Scholar] [CrossRef]
- Tikhani, F.; Hadavand, B.; Bafghi, H.; Jouyandeh, M.; Vahabi, H.; Formela, K.; Hosseini, H.; Paran, S.; Esmaeili, A.; Mohaddespour, A.; et al. Polyurethane/silane-functionalized ZrO2 nanocomposite powder coatings: Thermal degradation kinetics. Coatings 2020, 10, 413. [Google Scholar] [CrossRef]
- Duong, N.; An, T.; Oanh, V.; Vu, P.; Thao, P.; Hang, T. Degradation of polyurethane coating containing graphene oxide under effect of UV radiation, Vietnam. J. Chem. 2020, 58, 333–337. [Google Scholar] [CrossRef]
- Lu, L.; Ma, Z.; Liu, Q.; Zhang, D. Pollutant-accelerated aging behaviors of fluorocarbon coating in tropical marine atmosphere. Prog. Org. Coat. 2020, 139, 105447. [Google Scholar] [CrossRef]
- Filip, Z. Polyurethane as the sole nutrient source for Aspergillus niger and Cladosporium herbarum. Eur. J. Appl. Microbiol. Biotechnol. 1979, 7, 277–280. [Google Scholar] [CrossRef]
- García, G.; Velasco-Téllez, A.; Rodríguez-Herrera, R.; Aguilar, C. Polyurethane as substrate for fungal strains. Adv. Biosci. Biotechnol. 2011, 2, 52–58. [Google Scholar] [CrossRef] [Green Version]
- Rodrigo, O.; Daynet, S.; Jaime, N.; Nora, F.; Alejandro, A.; Jhonny, V.; Milton, A. Analysis of the degradation of polyethylene, polystyrene and polyurethane mediated by three filamentous fungi isolated from the Antarctica. Afr. J. Biotechnol. 2021, 20, 66–76. [Google Scholar] [CrossRef]
- Jehanno, C.; Pérez-Madrigal, M.; Demarteau, J.; Sardon, H.; Dove, A. Organocatalysis for depolymerization. Polym. Chem. 2019, 10, 172–186. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, H.; Akinari, Y.; Kaiso, K.; Kamimura, A. Efficient depolymerization and chemical conversion of polyamide 66 to 1,6-hexanediol. J. Mater. Cycles Waste Manag. 2017, 19, 326–331. [Google Scholar] [CrossRef]
- Shukla, S.; Harad, A.M.; Mahato, D. Depolymerization of nylon 6 waste fibers. J. Appl. Polym. Sci. 2006, 100, 186–190. [Google Scholar] [CrossRef]
- Hepburn, C. Polyurethane Elastomers; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012. [Google Scholar] [CrossRef]
- Gewert, B.; Plassmann, M.; MacLeod, M. Pathways for degradation of plastic polymers floating in the marine environment. Environ. Sci. Process. Impacts 2015, 17, 1513–1521. [Google Scholar] [CrossRef]
- Nakajima-Kambe, T.; Shigeno-Akutsu, Y.; Nomura, N.; Onuma, F.; Nakahara, T. Microbial degradation of polyurethane, polyester polyurethanes and polyether polyurethanes. Appl. Microbiol. Biot. 1999, 51, 134–140. [Google Scholar] [CrossRef]
- Lamba, N.; Woodhouse, K.; Cooper, S. Polyurethanes in Biomedical Applications; Routledge: New York, NY, USA, 2017. [Google Scholar] [CrossRef]
- Szociński, M.; Darowicki, K.; Schaefer, K. Application of impedance imaging to evaluation of organic coating degradation at a local scale. J. Coat. Technol. Res. 2013, 10, 65–72. [Google Scholar] [CrossRef] [Green Version]
- Sørensen, P.A.; Kiil, S.; Dam-Johansen, K.; Weinell, C. Anticorrosive coatings: A review. J. Coat. Technol. Res. 2009, 6, 135–176. [Google Scholar] [CrossRef]
- Koehler, E.L. The influence of contaminants on the failure of protective organic coatings on steel. Corrosion 1977, 33, 209–217. [Google Scholar] [CrossRef]
- Floyd, F.L.; Groseclose, R.G.; Frey, C.M. Mechanistic model for corrosion protection via paint. J. Oil. Col. Chem. Assoc. 1983, 66, 329. [Google Scholar]
- Pommersheim, J.M.; Nguyen, T.; Zhang, Z.; Hubbard, J.B. Degradation of organic coatings on steel: Mathematical models and predictions. Prog. Org. Coat. 1994, 25, 23–41. [Google Scholar] [CrossRef]
- Lu, L.; Liu, Q. Synergetic effects of photo-oxidation and biodegradation on failure behavior of polyester coating in tropical rain forest atmosphere. J. Mater. Sci. Technol. 2021, 64, 195–202. [Google Scholar] [CrossRef]
- Little, B.; Pope, R.; Ray, R. An evaluation of fungal-influenced corrosion of aircraft operating in marine tropical environments. Mar. Corros. Trop. Environ. Fla. 2000, 13, 257–269. [Google Scholar] [CrossRef]
- Rangel, D.N.E.; Anderson, A.J.; Roberts, D.W. Evaluating physical and nutritional stress during mycelial growth as inducers of tolerance to heat and UV-B radiation in Metarhizium anisopliae conidia. Mycol. Res. 2008, 112, 1362–1372. [Google Scholar] [CrossRef]
- Dias, L.P.; Araújo, C.A.S.; Pupin, B.; Ferreira, P.C.; Braga, G.Ú.L.; Rangel, D.E.N. The Xenon Test Chamber Q-SUN® for testing realistic tolerances of fungi exposed to simulated full spectrum solar radiation. Fungal Biol. 2018, 122, 592–601. [Google Scholar] [CrossRef]
- Gu, J.G.; Gu, J.D. Methods currently used in testing microbiological degradation and deterioration of a wide range of polymeric materials with various degree of degradability: A review. J. Polym. Environ. 2005, 13, 65–74. [Google Scholar] [CrossRef]
- Haque, M.; Chowdhury, R.; Islam, K.; Akbar, M.A. Propionic acid is an alternative to antibiotics in poultry diet. Bang. J. Anim. Sci. 2009, 38, 115–122. [Google Scholar] [CrossRef]
- Xu, P.; Leng, Y.; Zeng, G.; Huang, D.; Lai, C.; Zhao, M.; Cheng, M. Cadmium induced oxalic acid secretion and its role in metal uptake and detoxification mechanisms in Phanerochaete chrysosporium. Appl. Microbiol. Biot. 2015, 99, 435–443. [Google Scholar] [CrossRef]
- Seeley, M.E.; Song, B.; Passie, R.; Hale, R.C. Microplastics affect sedimentary microbial communities and nitrogen cycling. Nat. Commun. 2020, 1, 2372. [Google Scholar] [CrossRef] [PubMed]
- Nikje, M.M.A.; Nikrah, M. Chemical recycling and liquefaction of rigid polyurethane foam wastes through microwave assisted glycolysis process. J. Macromol. Sci A 2007, 44, 613–617. [Google Scholar] [CrossRef]
- Srikanth, M.; Sandeep, T.; Sucharitha, K.; Godi, S. Bioprocessing, Biodegradation of plastic polymers by fungi: A brief review. Bioresour. Bioprocess. 2022, 9, 42. [Google Scholar] [CrossRef]
- Madaj, R.; Sroczyński, W.; Sójka, M.; Olejnik, T.P.; Sobiecka, E.J.P. Batch mode reactor for 3, 5-dinitrosalicylic acid degradation by Phanerochaete chrysosporium. Processes 2021, 9, 105. [Google Scholar] [CrossRef]
- Skender, B.; Vaculová, A.; Slavík, J.; Machala, M.; Kozubík, A.; Hofmanová, J. Growth regulation activity of glycosylceramides and their metabolites in basidial fungus Flammulina velutipes. Chem. Phys. Lipids 2011, 164S, S32–S37. [Google Scholar] [CrossRef]
- Wu, W.; Hao, W.K.; Liu, Z.Y.; Li, X.G.; Du, C.W.; Liao, W.J. Corrosion behavior of E690 high-strength steel in alternating wet-dry marine environment with different pH values. J. Mater. Eng. Perform. 2015, 24, 4636–4646. [Google Scholar] [CrossRef]
- Aghaie, E.; Pazouki, M.; Hosseini, M.R.; Ranjbar, M.; Ghavipanjeh, F. Response surface methodology (RSM) analysis of organic acid production for Kaolin beneficiation by Aspergillus niger. Chem. Eng. J. 2009, 147, 245–251. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Organic Acid | Content-1 (µg/mL) | Content-2 (µg/mL) | Content-3 (µg/mL) |
|---|---|---|---|---|
| 1 | citric acid | 124.75 | 145.31 | 120.62 |
| 2 | propanoic acid | 81.85 | 87.55 | 107.96 |
| 3 | succinic acid | 42.36 | 51.50 | 37.89 |
| 4 | tartaric acid | 22.04 | 40.05 | 62.71 |
| 5 | malonic acid | 24.52 | 2.72 | 24.81 |
| 6 | malic acid | 13.69 | 14.51 | 23.68 |
| 7 | acetic acid | 10.99 | 11.85 | 11.62 |
| 8 | lactic acid | 0.87 | 11.89 | 11.34 |
| 9 | methanoic acid | 6.81 | 7.63 | 7.50 |
| 10 | oxalic acid | 3.49 | 3.74 | 4.38 |
| Total | 331.36 | 376.75 | 412.50 | |
| No. | Organic Acid | Content-1 (µg/mL) | Content-2 (µg/mL) | Content-3 (µg/mL) |
|---|---|---|---|---|
| 1 | oxalic acid | 123.65 | 54.50 | 93.92 |
| 2 | acetic acid | 73.17 | 16.45 | 28.77 |
| 3 | methanoic acid | 49.56 | 67.90 | 48.60 |
| 4 | propanoic acid | 30.09 | 35.78 | 35.57 |
| 5 | lactic acid | 19.43 | 45.31 | 28.86 |
| 6 | malonic acid | 22.62 | 20.90 | 2.17 |
| 7 | tartaric acid | 1.13 | 14.22 | 15.78 |
| 8 | malic acid | 0.00 | 4.05 | 38.41 |
| 9 | citric acid | 4.21 | 16.29 | 4.25 |
| 10 | succinic acid | 0.00 | 60.73 | 0.00 |
| Total | 323.84 | 336.13 | 346.34 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hao, X.; Yang, K.; Zhang, D.; Lu, L. Insight into Degrading Effects of Two Fungi on Polyurethane Coating Failure in a Simulated Atmospheric Environment. Polymers 2023, 15, 328. https://doi.org/10.3390/polym15020328
Hao X, Yang K, Zhang D, Lu L. Insight into Degrading Effects of Two Fungi on Polyurethane Coating Failure in a Simulated Atmospheric Environment. Polymers. 2023; 15(2):328. https://doi.org/10.3390/polym15020328
Chicago/Turabian StyleHao, Xiangping, Kexin Yang, Dawei Zhang, and Lin Lu. 2023. "Insight into Degrading Effects of Two Fungi on Polyurethane Coating Failure in a Simulated Atmospheric Environment" Polymers 15, no. 2: 328. https://doi.org/10.3390/polym15020328