Polyplexes of Functional PAMAM Dendrimer/Apoptin Gene Induce Apoptosis of Human Primary Glioma Cells In Vitro

Abstract
:
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Synthesis of PAMAM-FHR
2.3. Plasmids and Cell Culture
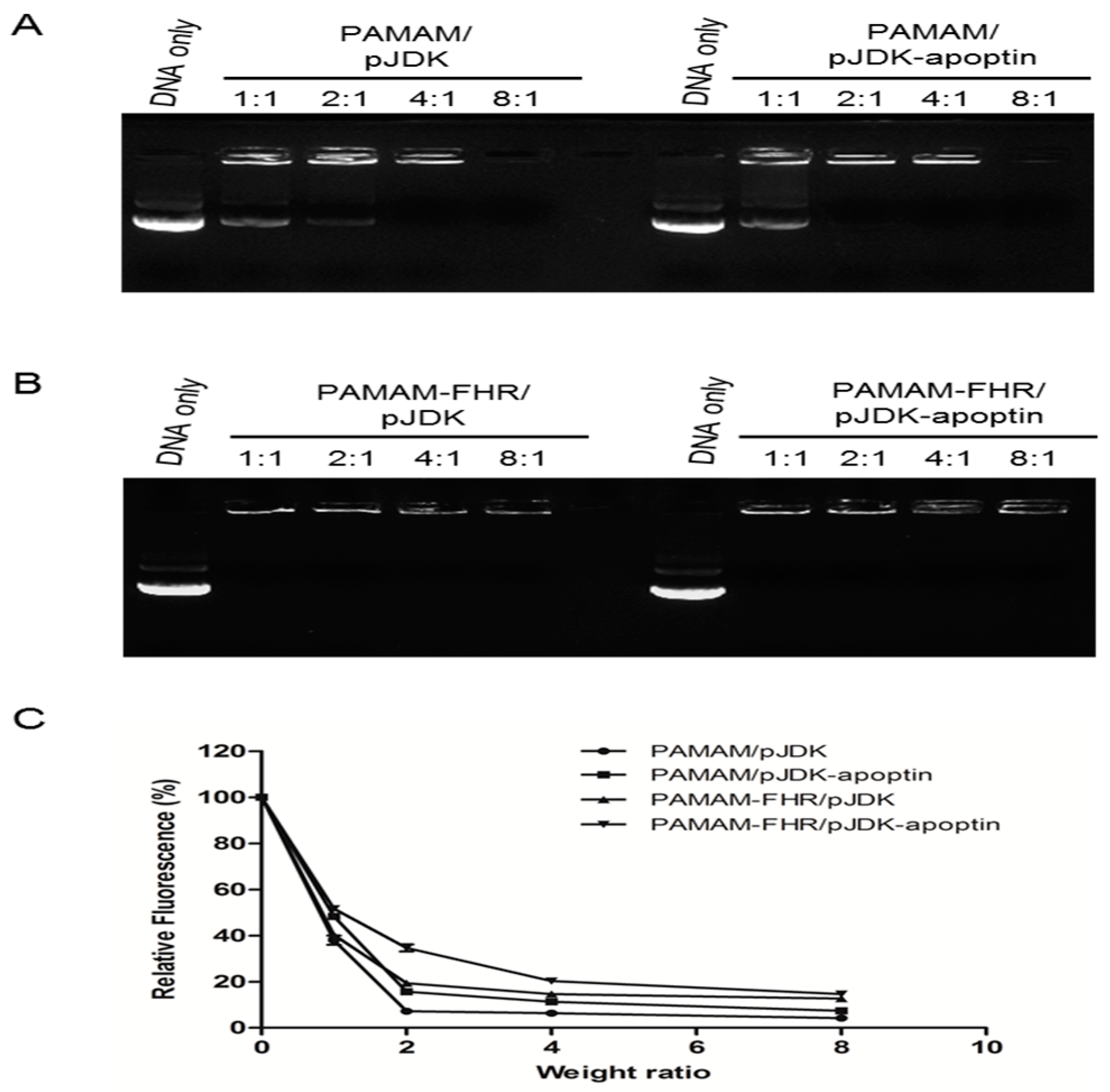
2.4. Gel Retardation Assay
2.5. PicoGreen Assay
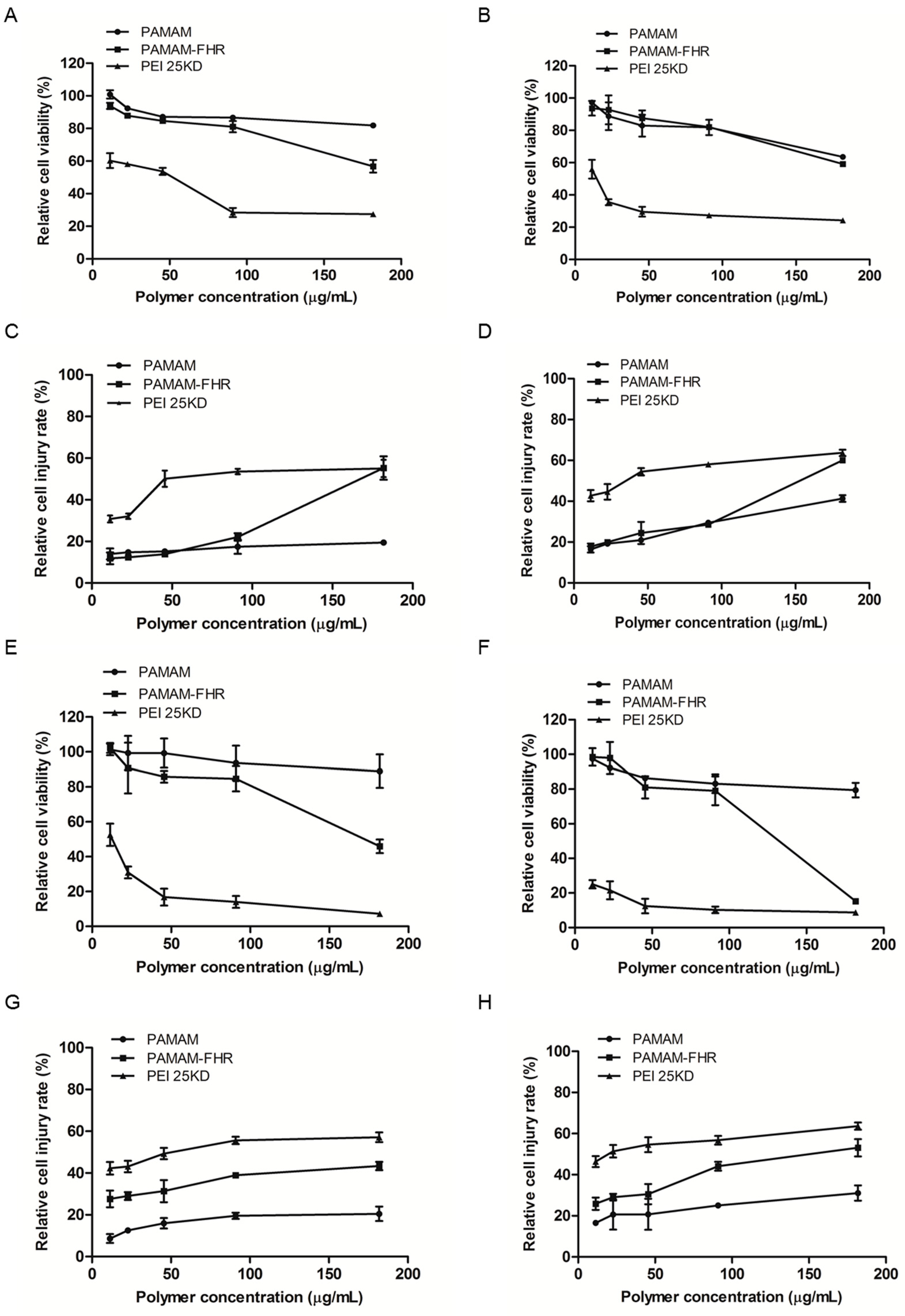
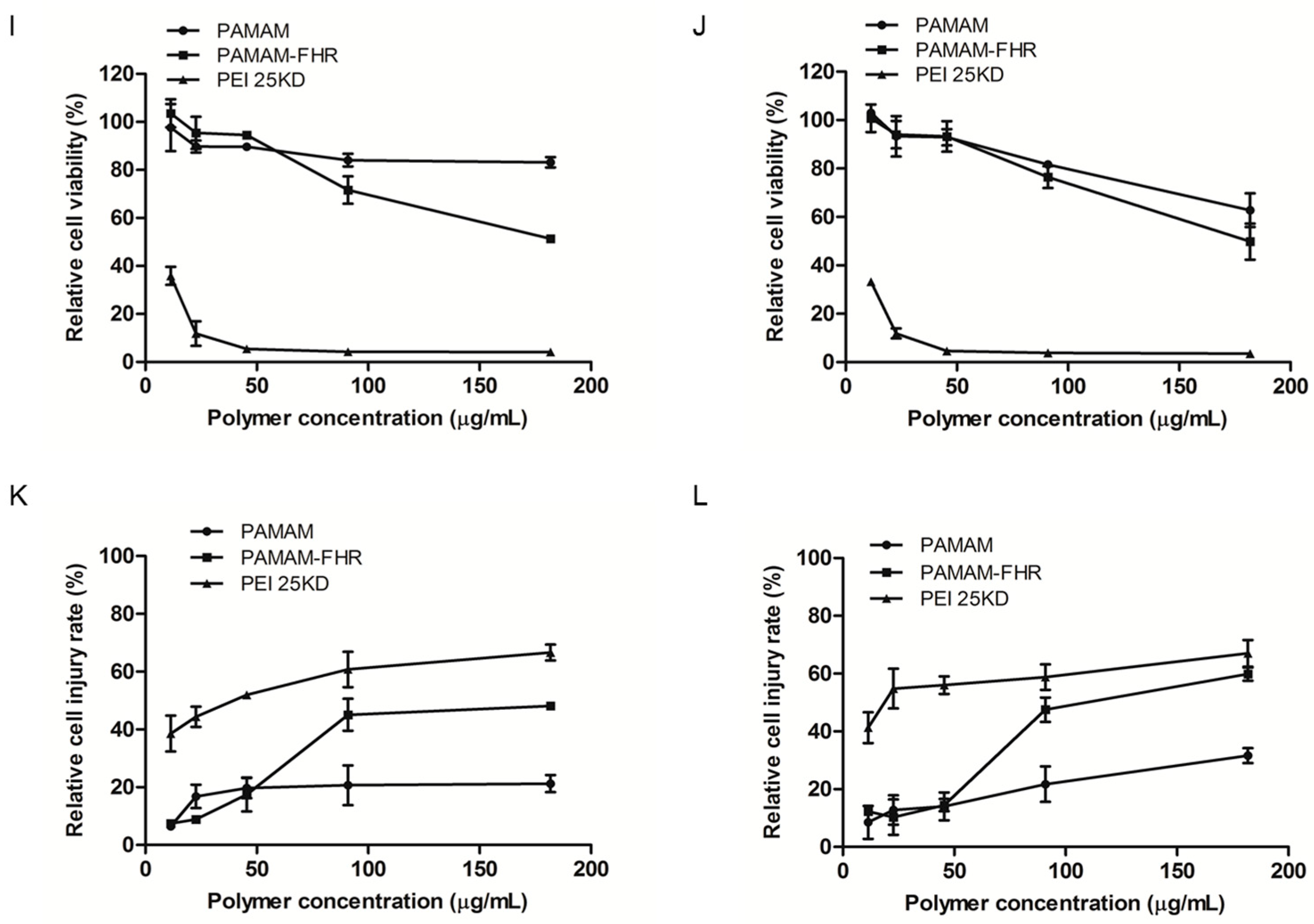
2.6. Cytotoxicity Assay
2.7. Cellular Uptake Imaging
2.8. In Vitro Transfection Assay
2.9. Expression of Green Fluorescent Protein (GFP)
2.10. Cell Cycle Analysis
2.11. Imaging of Intracellular Trafficking
2.12. Mitochondrial Membrane Potential (MMP) Assay
2.13. Annexin V Staining
2.14. RNA Extract and Real-Time-PCR Quantification
2.15. Measurement of Dynamic Light Scattering (DLS) and Zeta Potential
2.16. Statistical Analysis
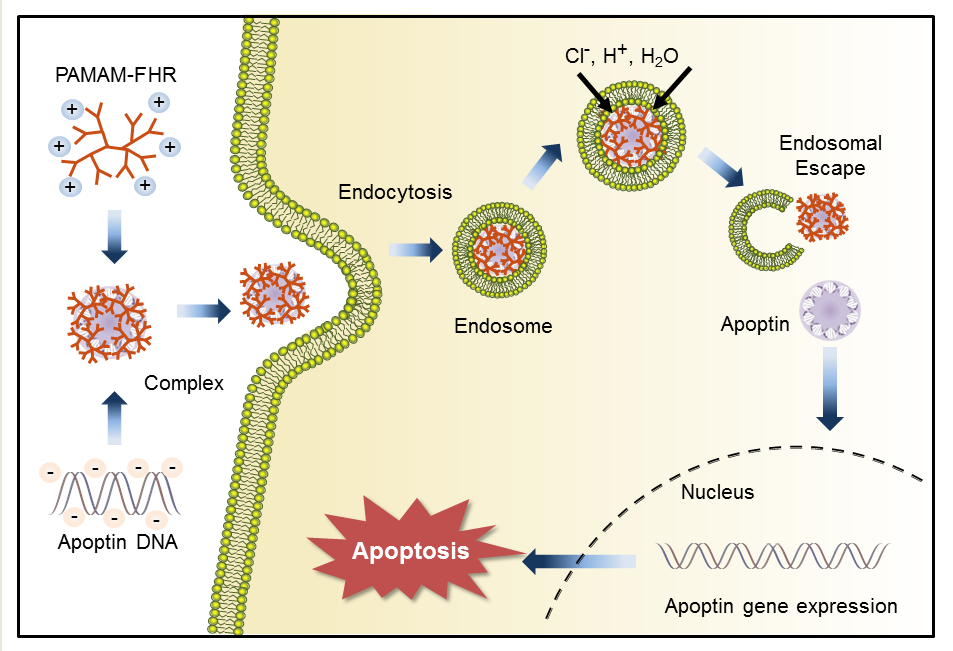
3. Results and Discussion
3.1. Characteristics of PAMAM-FHR and Complexes with Plasmid DNA
3.2. Characterization of Complexes
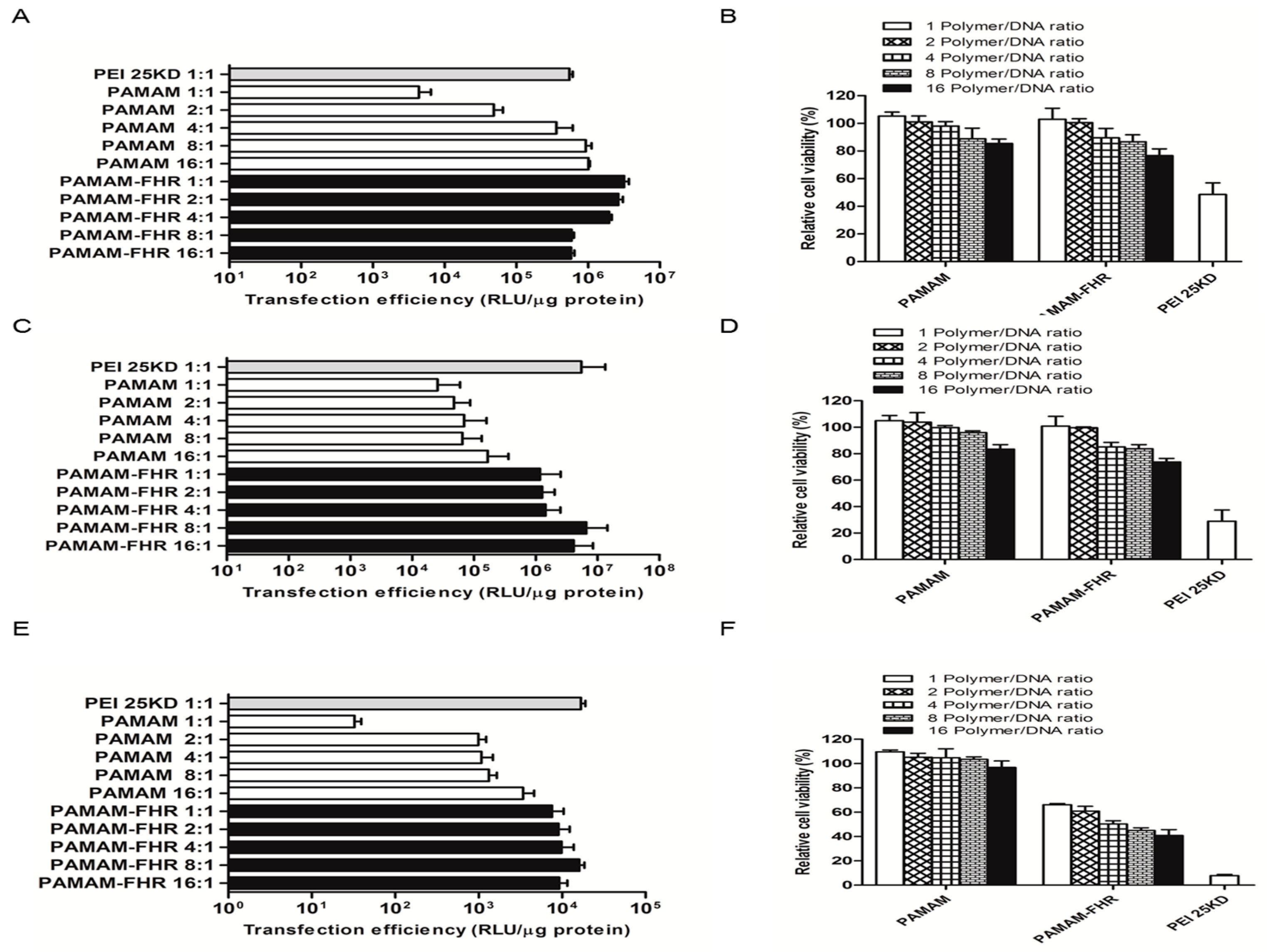
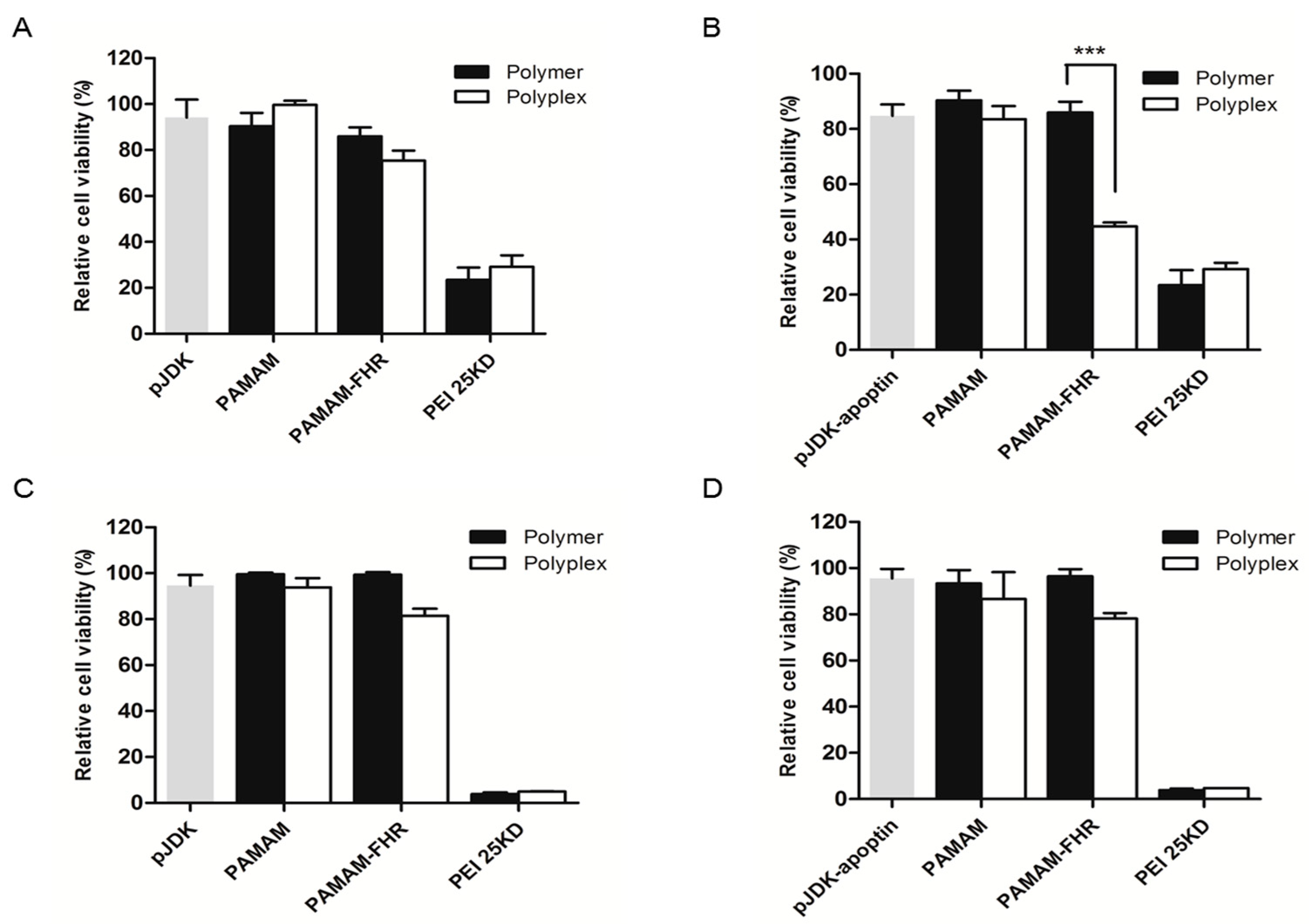
3.3. Cytotoxicity Assay
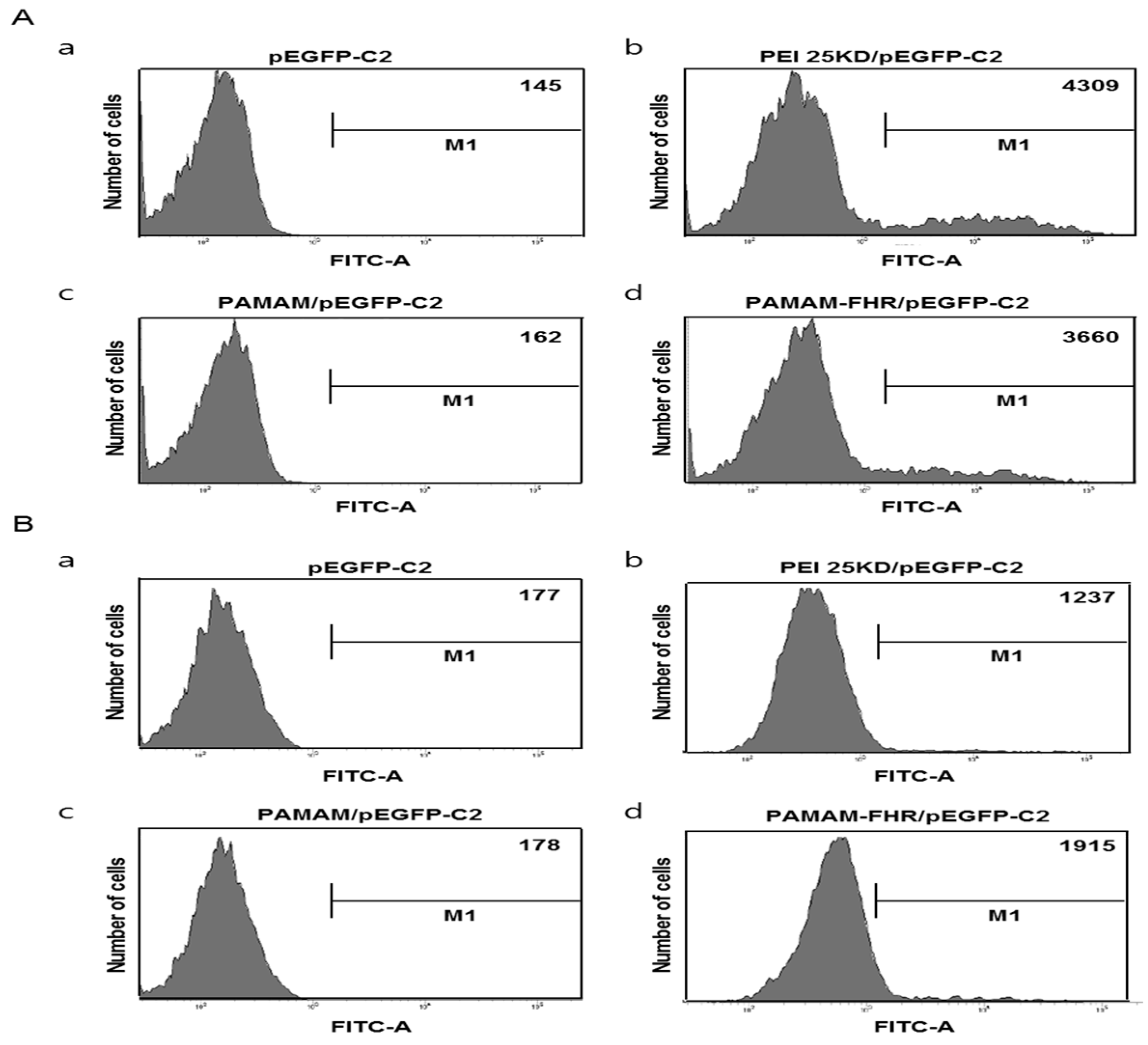
3.4. Transfection Efficiency In Vitro
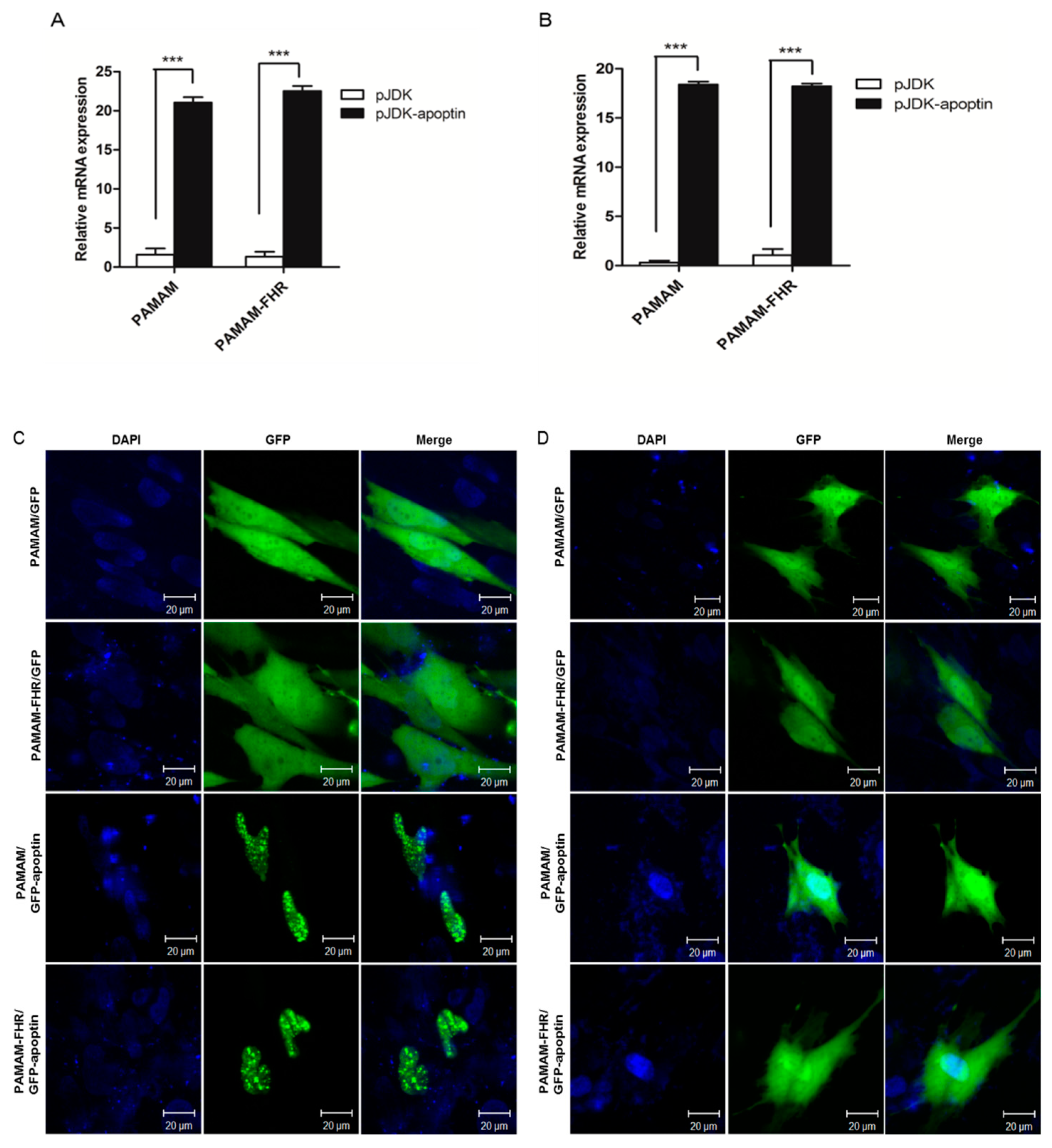
3.5. Expression of Apoptin in Cells Treated with PAMAM-FHR/pJDK-Apoptin Complexes
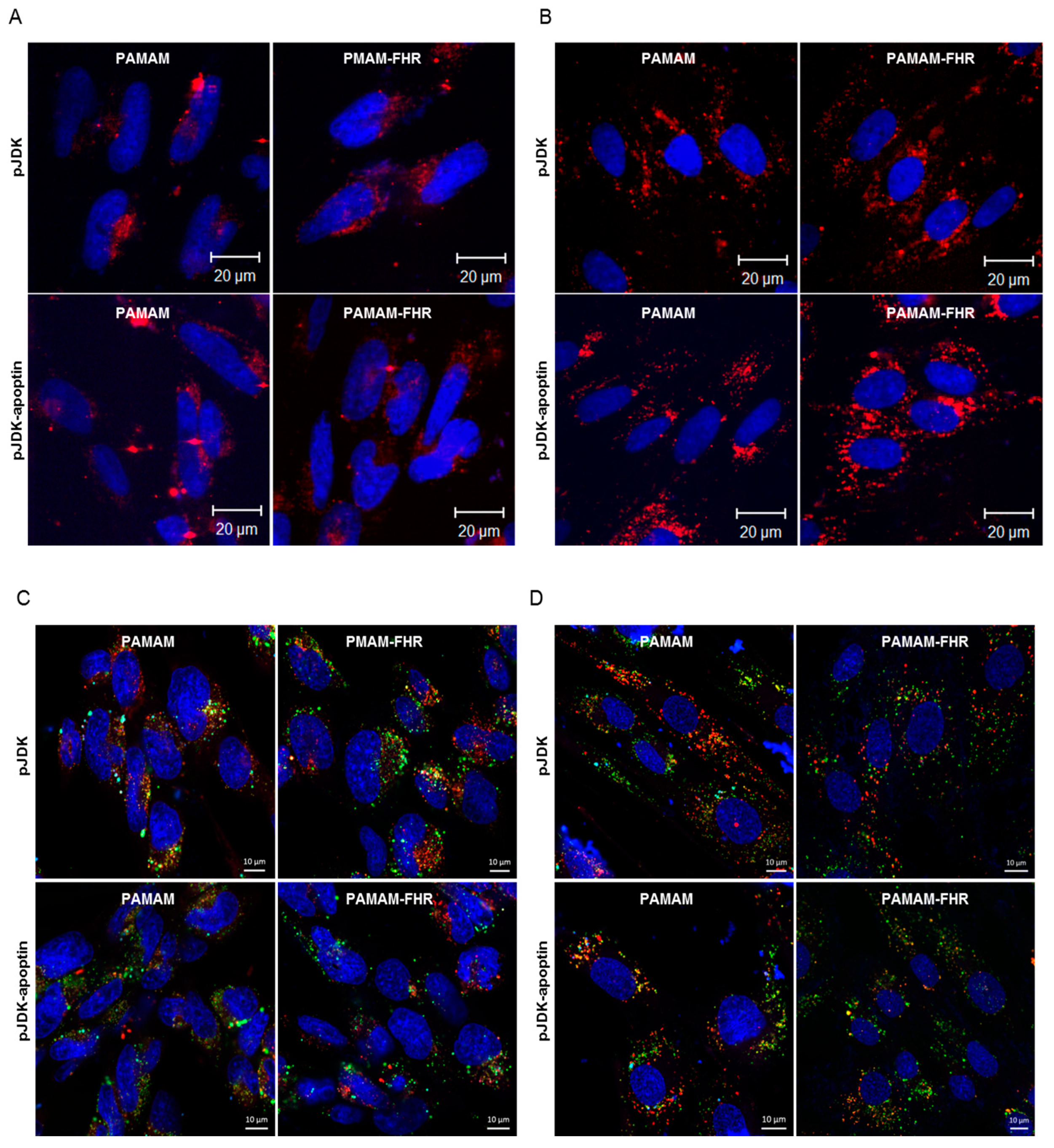
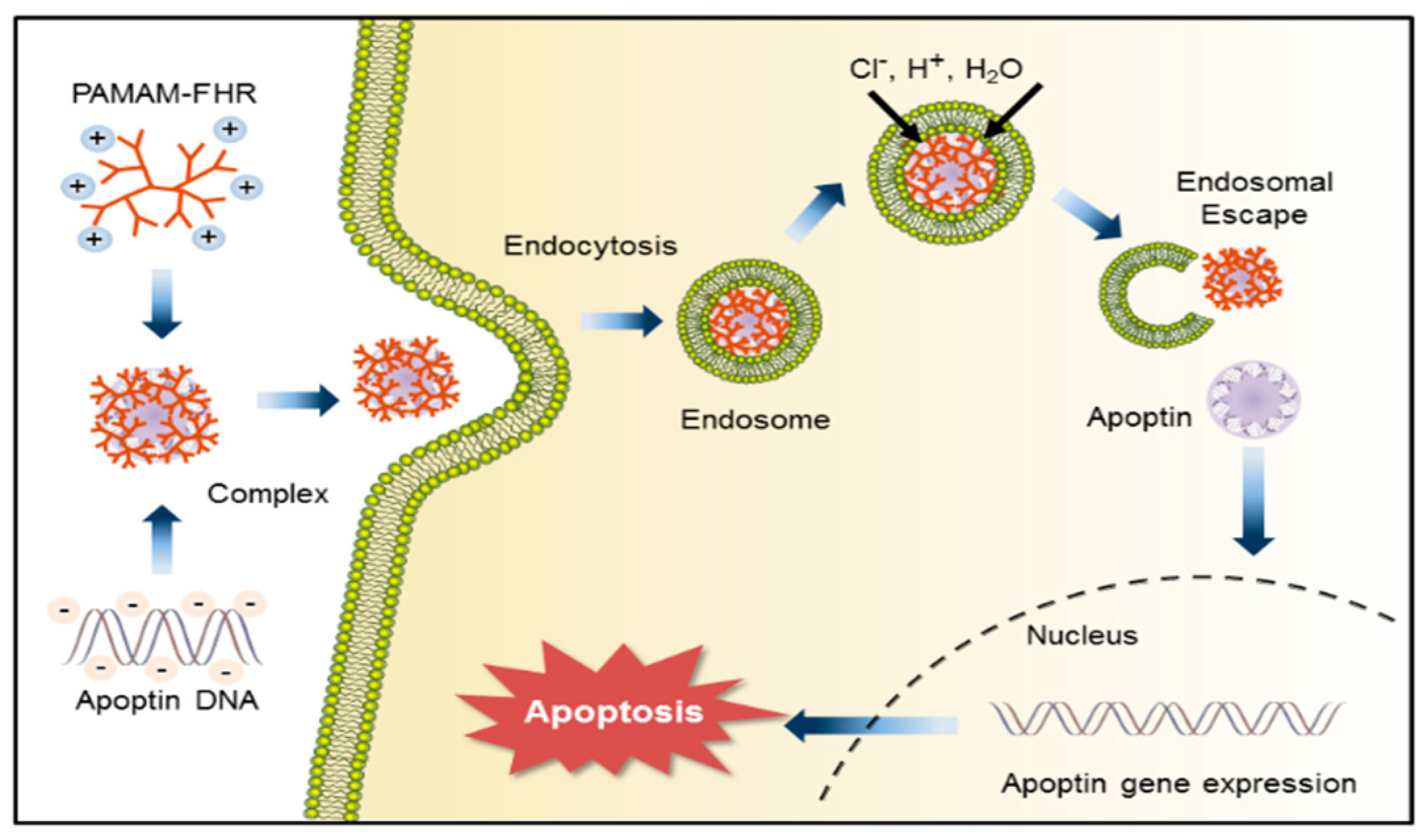
3.6. Intracellular Traffic of PAMAM-FHR/Apoptin Complexes
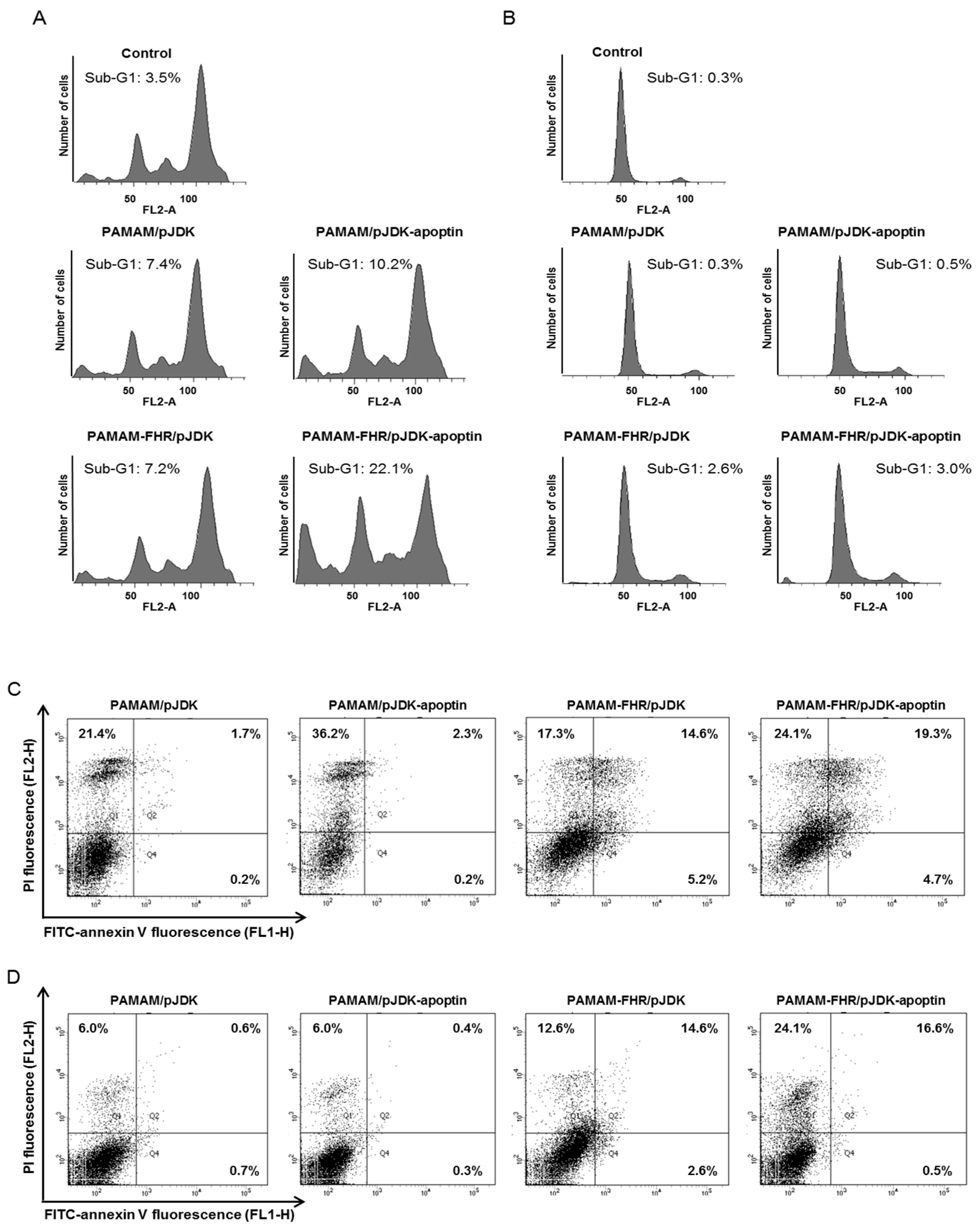
3.7. PAMAM-FHR/pJDK-Apoptin Complexes Induce Apoptosis In Vitro
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Assi, H.; Candolfi, M.; Baker, G.; Mineharu, Y.; Lowenstein, P.R.; Castro, M.G. Gene therapy for brain tumors: Basic developments and clinical implementation. Neurosci. Lett. 2012, 527, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Juratli, T.A.; Schackert, G.; Krex, D. Current status of local therapy in malignant gliomas--a clinical review of three selected approaches. Pharmacol. Ther. 2013, 139, 341–358. [Google Scholar] [CrossRef] [PubMed]
- Reardon, D.A.; Wen, P.Y. Therapeutic advances in the treatment of glioblastoma: Rationale and potential role of targeted agents. Oncologist 2006, 11, 152–164. [Google Scholar] [CrossRef] [PubMed]
- Kanu, O.O.; Mehta, A.; Di, C.; Lin, N.; Bortoff, K.; Bigner, D.D.; Yan, H.; Adamson, D.C. Glioblastoma multiforme: A review of therapeutic targets. Expert Opin. Ther. Targets 2009, 13, 701–718. [Google Scholar] [CrossRef] [PubMed]
- Pourgholi, F.; Hajivalili, M.; Farhad, J.N.; Kafil, H.S.; Yousefi, M. Nanoparticles: Novel vehicles in treatment of glioblastoma. Biomed. Pharmacother. 2016, 77, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Wegscheid, M.L.; Morshed, R.A.; Cheng, Y.; Lesniak, M.S. The art of attraction: Applications of multifunctional magnetic nanomaterials for malignant glioma. Expert Opin. Drug Deliv. 2014, 11, 957–975. [Google Scholar] [CrossRef] [PubMed]
- Engelhard, H.H. Gene therapy for brain tumors: The fundamentals. Surg. Neurol. 2000, 54, 3–9. [Google Scholar] [CrossRef]
- Kane, J.R.; Miska, J.; Young, J.S.; Kanojia, D.; Kim, J.W.; Lesniak, M.S. Sui generis: Gene therapy and delivery systems for the treatment of glioblastoma. Neuro-Oncology 2015, 17 (Suppl. 2), ii24–ii36. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Hu, M.; Wang, D.; Wang, G.; Zhu, X.; Yan, D.; Sun, J. Multifunctional hyperbranched glycoconjugated polymers based on natural aminoglycosides. Bioconjugate Chem. 2012, 23, 1189–1199. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, Z.; Zhu, X.; Chen, M. A smart gene delivery platform: Cationic oligomer. Eur. J. Pharm. Sci. 2017, 105, 33–40. [Google Scholar] [CrossRef]
- Lin, G.; Zhang, H.; Huang, L. Smart polymeric nanoparticles for cancer gene delivery. Mol. Pharm. 2015, 12, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Al-Dosari, M.S.; Gao, X. Nonviral gene delivery: Principle, limitations, and recent progress. AAPS J. 2009, 11, 671–681. [Google Scholar] [CrossRef] [PubMed]
- Cloninger, M.J. Biological applications of dendrimers. Curr. Opin. Chem. Biol. 2002, 6, 742–748. [Google Scholar] [CrossRef]
- Jones, C.H.; Chen, C.K.; Ravikrishnan, A.; Rane, S.; Pfeifer, B.A. Overcoming nonviral gene delivery barriers: Perspective and future. Mol. Pharm. 2013, 10, 4082–4098. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.C.; MacKay, J.A.; Frechet, J.M.; Szoka, F.C. Designing dendrimers for biological applications. Nat. Biotechnol. 2005, 23, 1517–1526. [Google Scholar] [CrossRef] [PubMed]
- Bielinska, A.U.; Kukowska-Latallo, J.F.; Baker, J.R., Jr. The interaction of plasmid DNA with polyamidoamine dendrimers: Mechanism of complex formation and analysis of alterations induced in nuclease sensitivity and transcriptional activity of the complexed DNA. Biochim. Biophys. Acta 1997, 1353, 180–190. [Google Scholar] [CrossRef]
- Bae, Y.; Green, E.S.; Kim, G.Y.; Song, S.J.; Mun, J.Y.; Lee, S.; Park, J.I.; Park, J.S.; Ko, K.S.; Han, J.; et al. Dipeptide-functionalized polyamidoamine dendrimer-mediated apoptin gene delivery facilitates apoptosis of human primary glioma cells. Int. J. Pharm. 2016, 515, 186–200. [Google Scholar] [CrossRef] [PubMed]
- Thuy le, T.; Mallick, S.; Choi, J.S. Polyamidoamine (pamam) dendrimers modified with short oligopeptides for early endosomal escape and enhanced gene delivery. Int. J. Pharm. 2015, 492, 233–243. [Google Scholar] [CrossRef]
- Truong, N.P.; Jia, Z.; Burgess, M.; Payne, L.; McMillan, N.A.; Monteiro, M.J. Self-catalyzed degradable cationic polymer for release of DNA. Biomacromolecules 2011, 12, 3540–3548. [Google Scholar] [CrossRef]
- Whitfield, R.; Anastasaki, A.; Truong Phuoc, N.; Cook, A.; Omedes-Pujol, M.; Loczenski Rose, V.; Nguyen, T.; Burns, J.A.; Perrier, S.; Davis, T.; et al. Efficient binding, protection, and self-release of dsrna in soil by linear and star cationic polymers. ACS Macro Lett. 2018, 7, 909–915. [Google Scholar] [CrossRef]
- Gu, W.; Jia, Z.; Truong, N.P.; Prasadam, I.; Xiao, Y.; Monteiro, M.J. Polymer nanocarrier system for endosome escape and timed release of sirna with complete gene silencing and cell death in cancer cells. Biomacromolecules 2013, 14, 3386–3389. [Google Scholar] [CrossRef] [PubMed]
- Truong, N.P.; Gu, W.; Prasadam, I.; Jia, Z.; Crawford, R.; Xiao, Y.; Monteiro, M.J. An influenza virus-inspired polymer system for the timed release of sirna. Nat. Commun. 2013, 4, 1902. [Google Scholar] [CrossRef] [PubMed]
- Los, M.; Panigrahi, S.; Rashedi, I.; Mandal, S.; Stetefeld, J.; Essmann, F.; Schulze-Osthoff, K. Apoptin, a tumor-selective killer. Biochim. Biophys. Acta 2009, 1793, 1335–1342. [Google Scholar] [CrossRef] [PubMed]
- Rollano Penaloza, O.M.; Lewandowska, M.; Stetefeld, J.; Ossysek, K.; Madej, M.; Bereta, J.; Sobczak, M.; Shojaei, S.; Ghavami, S.; Los, M.J. Apoptins: Selective anticancer agents. Trends Mol. Med. 2014, 20, 519–528. [Google Scholar] [CrossRef] [PubMed]
- Tavassoli, M.; Guelen, L.; Luxon, B.A.; Gaken, J. Apoptin: Specific killer of tumor cells? Apoptosis 2005, 10, 717–724. [Google Scholar] [CrossRef] [PubMed]
- Nie, Y.Q.; Leng, X.X.; Jiang, Y.X.; Chai, S.G.; Zhang, J.Z.; Zou, Q.C. Influence of reactive poss and ddp on thermal stability and flame retardance of upr nanocomposites. E-Polymers 2017, 17, 463–470. [Google Scholar] [CrossRef]
- An, S.; Nam, K.; Choi, S.; Bai, C.Z.; Lee, Y.; Park, J.S. Nonviral gene therapy in vivo with pam-rg4/apoptin as a potential brain tumor therapeutic. Int. J. Nanomed. 2013, 8, 821–834. [Google Scholar]
- Bae, Y.; Rhim, H.S.; Lee, S.; Ko, K.S.; Han, J.; Choi, J.S. Apoptin gene delivery by the functionalized polyamidoamine dendrimer derivatives induces cell death of u87-mg glioblastoma cells. J. Pharm. Sci. 2017, 106, 1618–1633. [Google Scholar] [CrossRef]
- Paleos, C.M.; Tziveleka, L.A.; Sideratou, Z.; Tsiourvas, D. Gene delivery using functional dendritic polymers. Expert Opin. Drug Deliv. 2009, 6, 27–38. [Google Scholar] [CrossRef]
- Hunter, A.C. Molecular hurdles in polyfectin design and mechanistic background to polycation induced cytotoxicity. Adv. Drug Deliv. Rev. 2006, 58, 1523–1531. [Google Scholar] [CrossRef]
- Holder, A.L.; Goth-Goldstein, R.; Lucas, D.; Koshland, C.P. Particle-induced artifacts in the mtt and ldh viability assays. Chem. Res. Toxicol. 2012, 25, 1885–1892. [Google Scholar] [CrossRef] [PubMed]
- Freeman, E.C.; Weiland, L.M.; Meng, W.S. Modeling the proton sponge hypothesis: Examining proton sponge effectiveness for enhancing intracellular gene delivery through multiscale modeling. J. Biomater. Sci. Polym. Ed. 2013, 24, 398–416. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.S.; Zhu, X.Y.; Yan, D.Y. A controlled release system for simultaneous promotion of gene transfection and antitumor effects. RSC Adv. 2014, 4, 64596–64600. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, Z.P.; Chen, M.S. The length of hydrophobic chain in amphiphilic polypeptides regulates the efficiency of gene delivery. Polymers 2018, 10, 379. [Google Scholar] [CrossRef]
- Liu, Z.; Zhang, Z.; Zhou, C.; Jiao, Y. Hydrophobic modifications of cationic polymers for gene delivery. Prog. Polym. Sci. 2010, 35, 1144–1162. [Google Scholar] [CrossRef]
- Zhang, S.; Gao, H.; Bao, G. Physical principles of nanoparticle cellular endocytosis. ACS Nano 2015, 9, 8655–8671. [Google Scholar] [CrossRef]
- Santos, J.L.; Pandita, D.; Rodrigues, J.; Pego, A.P.; Granja, P.L.; Balian, G.; Tomas, H. Receptor-mediated gene delivery using pamam dendrimers conjugated with peptides recognized by mesenchymal stem cells. Mol. Pharm. 2010, 7, 763–774. [Google Scholar] [CrossRef]
- Miyamoto, S.; Howes, A.L.; Adams, J.W.; Dorn, G.W., 2nd; Brown, J.H. Ca2+ dysregulation induces mitochondrial depolarization and apoptosis: Role of na+/ca2+ exchanger and akt. J. Biol. Chem. 2005, 280, 38505–38512. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Complexes | Size (nm) | Polydispersity (PDI) | Zeta Potential (mV) |
|---|---|---|---|
| PAMAM/pJDK | 284.6 ± 1.6 | 0.13 | 37.8 ± 3.3 |
| PAMAM-FHR/pJDK | 185.7 ± 6.3 | 0.21 | 40.9 ± 2.7 |
| PAMAM/pJDK-apoptin | 239.2 ± 0.7 | 0.20 | 36.3 ± 1.3 |
| PAMAM-FHR/pJDK-apoptin | 178.7 ± 1.2 | 0.23 | 38.5 ± 3.5 |

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bae, Y.; Thuy, L.T.; Lee, Y.H.; Ko, K.S.; Han, J.; Choi, J.S. Polyplexes of Functional PAMAM Dendrimer/Apoptin Gene Induce Apoptosis of Human Primary Glioma Cells In Vitro. Polymers 2019, 11, 296. https://doi.org/10.3390/polym11020296
Bae Y, Thuy LT, Lee YH, Ko KS, Han J, Choi JS. Polyplexes of Functional PAMAM Dendrimer/Apoptin Gene Induce Apoptosis of Human Primary Glioma Cells In Vitro. Polymers. 2019; 11(2):296. https://doi.org/10.3390/polym11020296
Chicago/Turabian StyleBae, Yoonhee, Le Thi Thuy, Young Hwa Lee, Kyung Soo Ko, Jin Han, and Joon Sig Choi. 2019. "Polyplexes of Functional PAMAM Dendrimer/Apoptin Gene Induce Apoptosis of Human Primary Glioma Cells In Vitro" Polymers 11, no. 2: 296. https://doi.org/10.3390/polym11020296