
Biochemical and Spectroscopic Characterization of a Recombinant Laccase from Thermoalkaliphilic Bacillus sp. FNT with Potential for Degradation of Polycyclic Aromatic Hydrocarbons (PAHs)

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
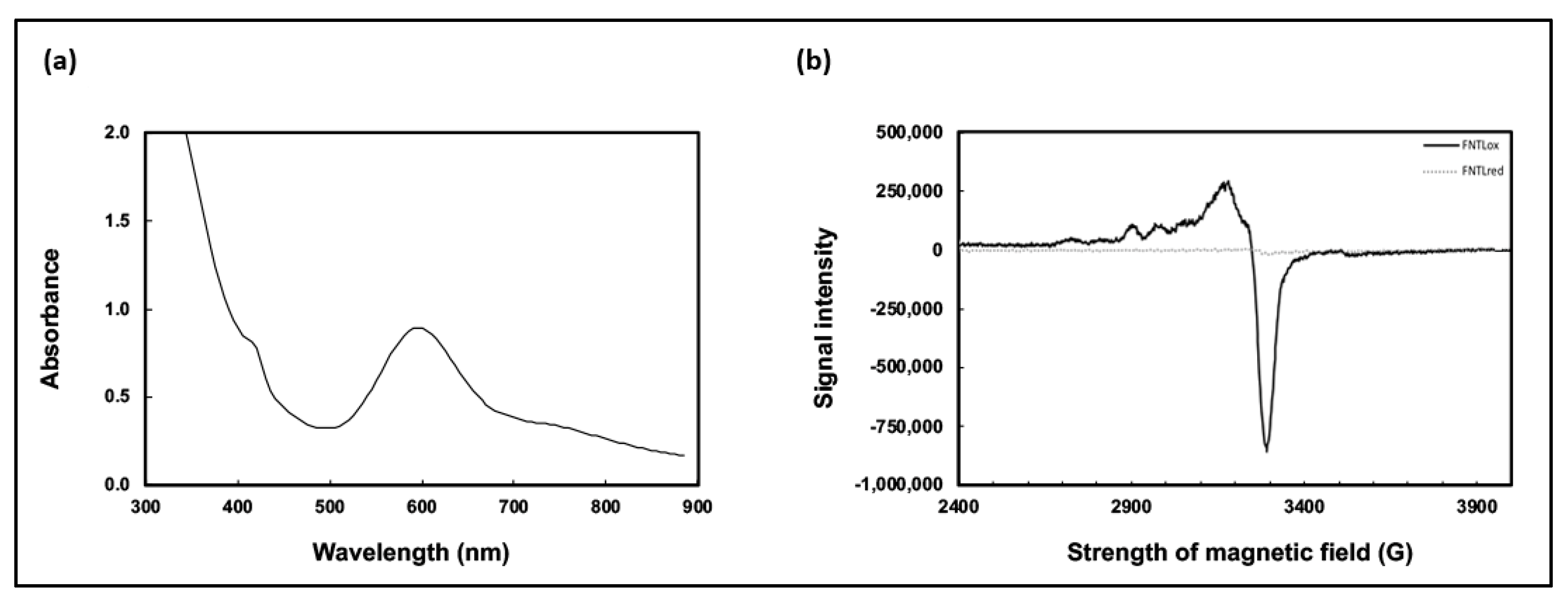
2.1. Further Characterization of Recombinant FNT Laccase (FNTL)
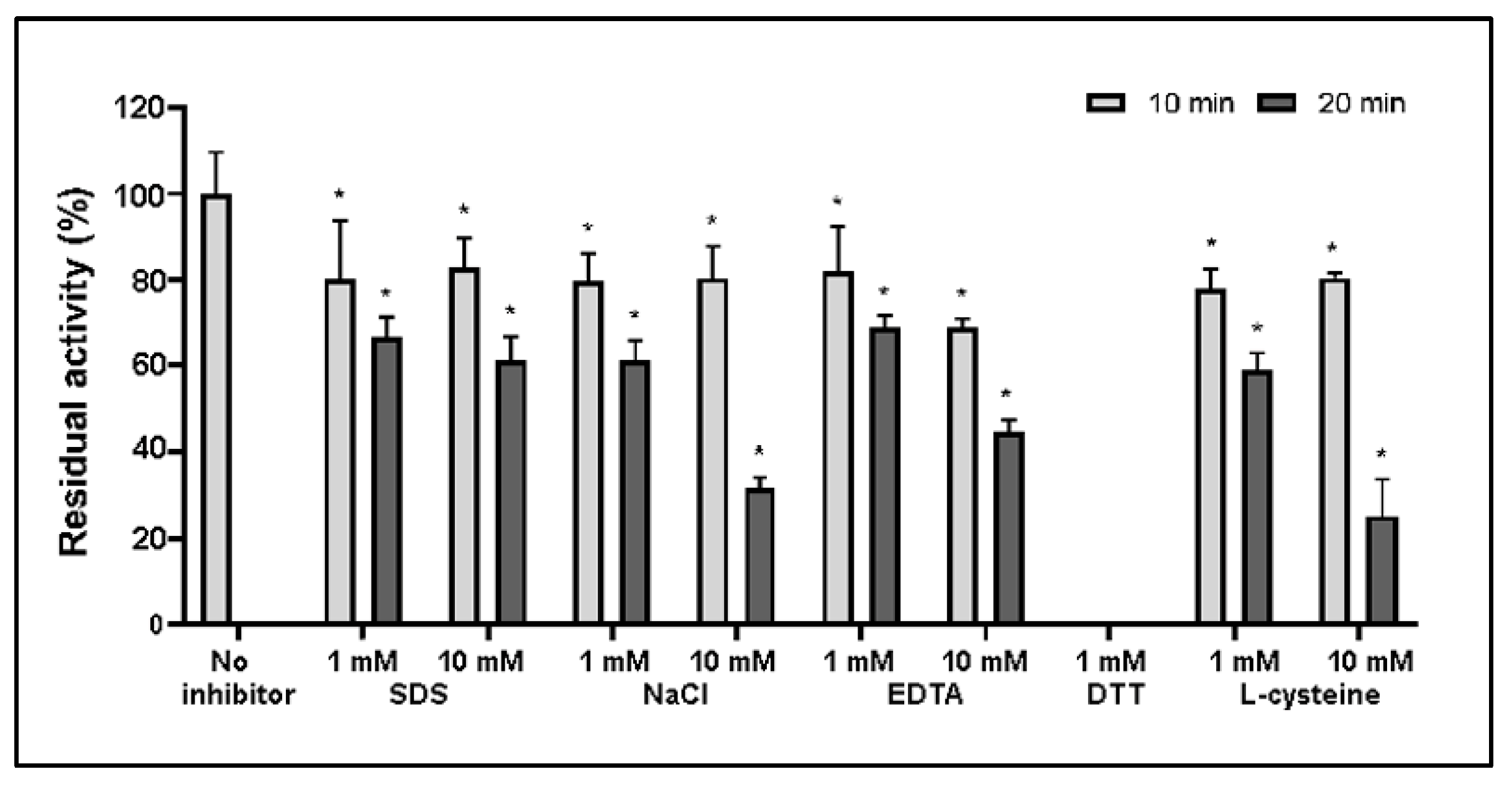
2.2. Effect of Inhibitors on FNTL
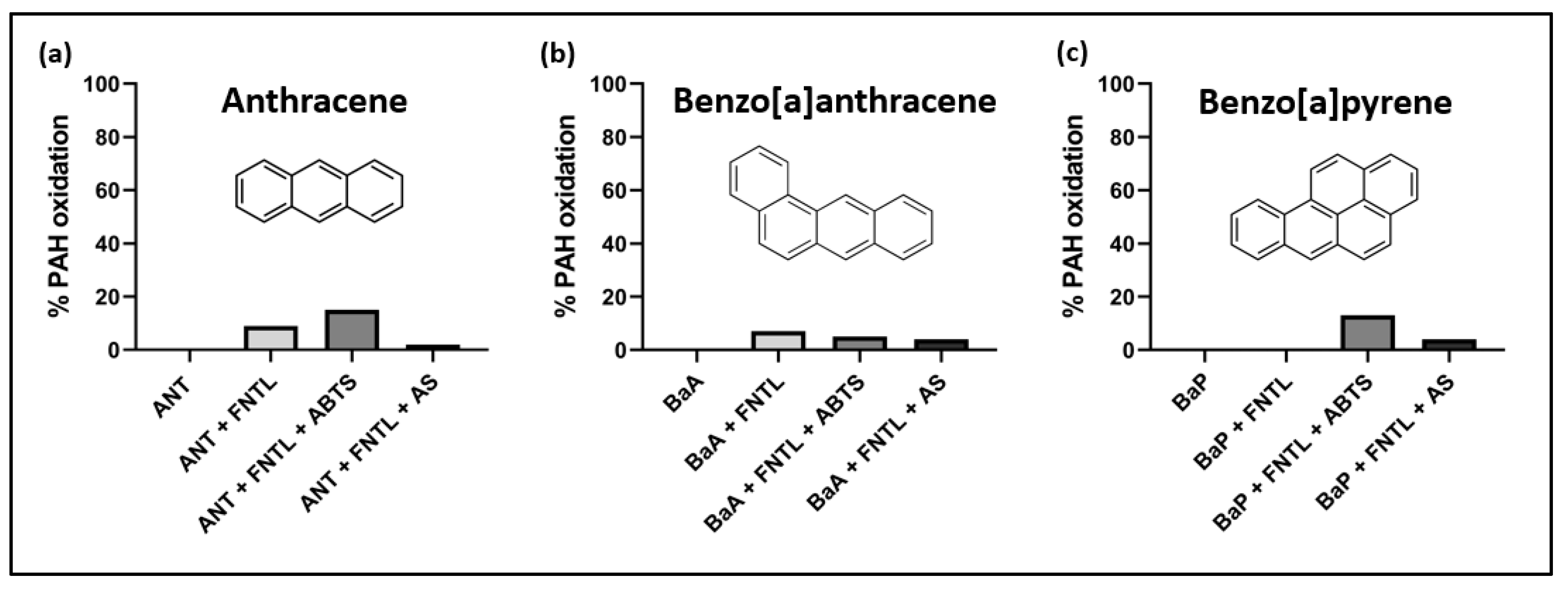
2.3. Evaluation of PAHs Oxidation
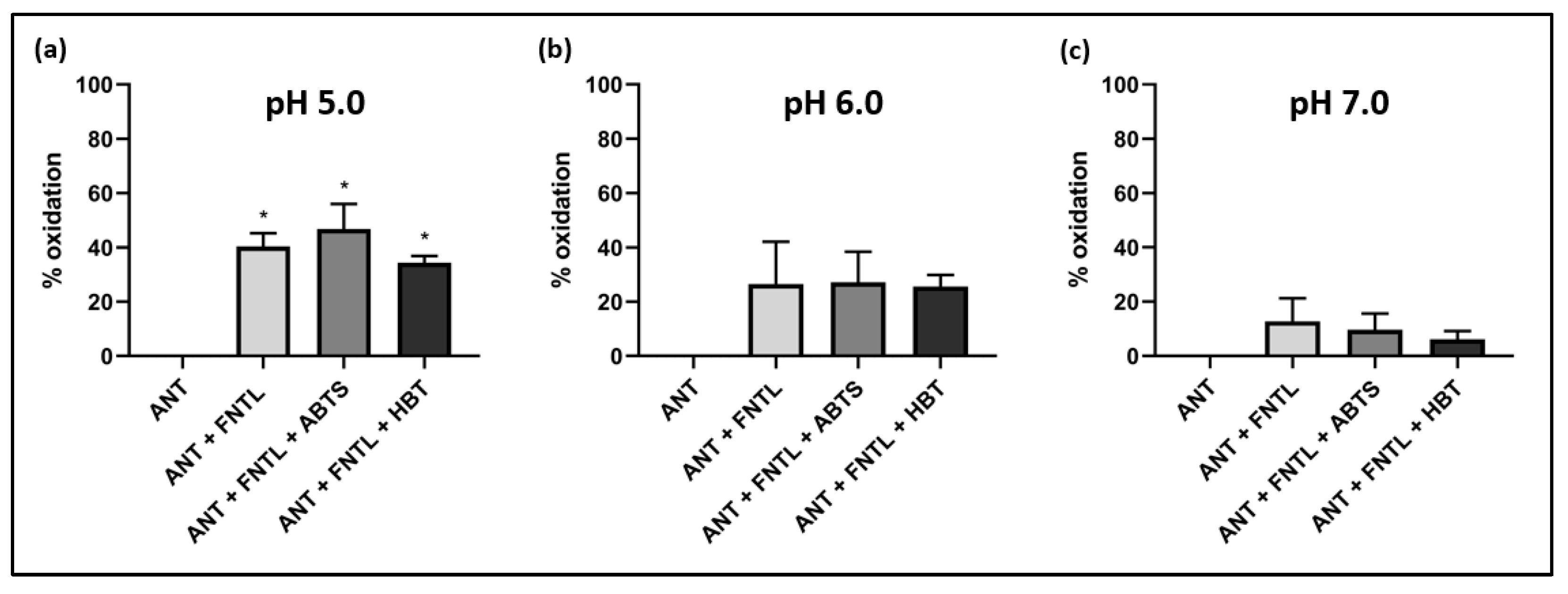
2.3.1. Evaluation of ANT Oxidation and Effect of Different pH
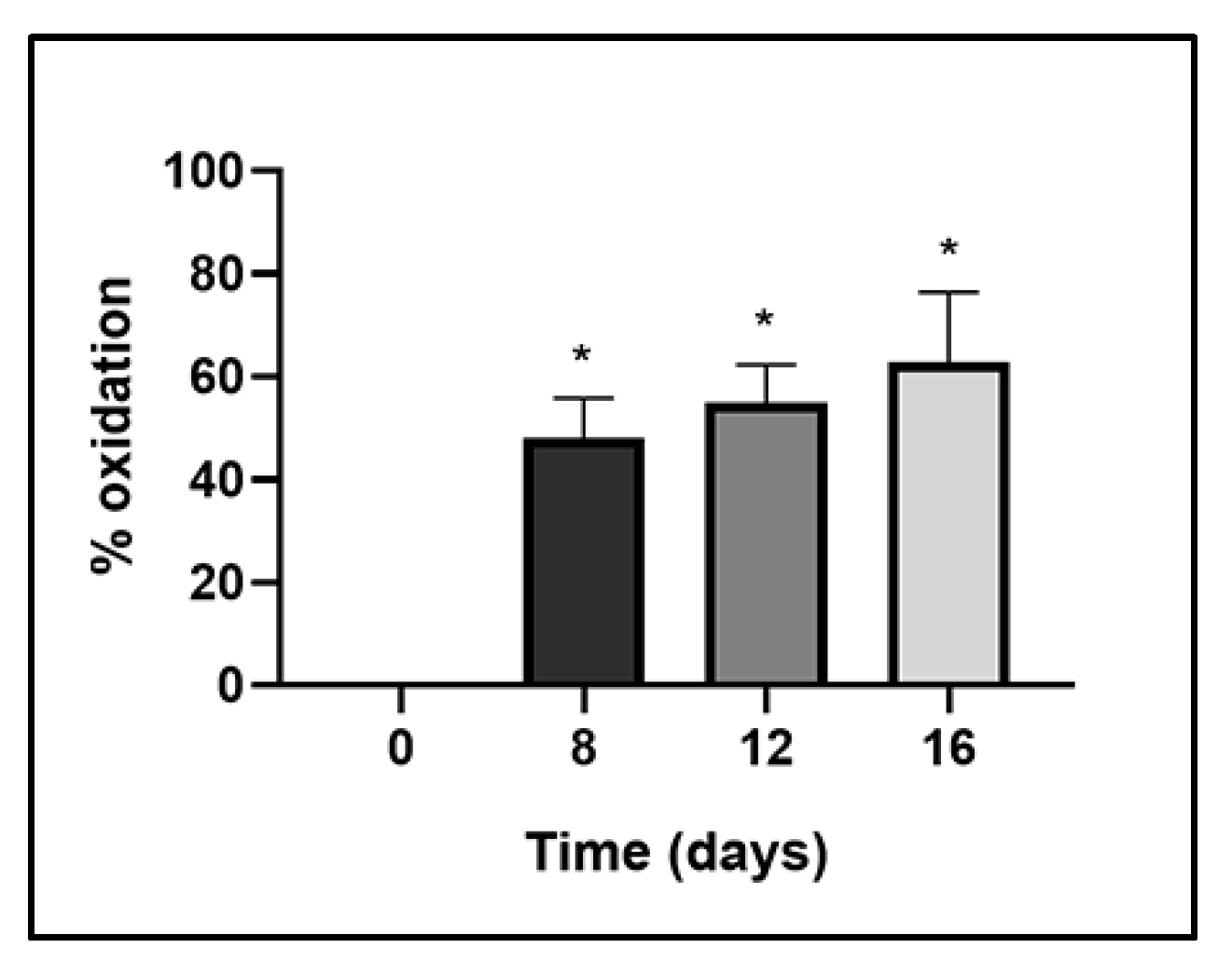
2.3.2. Evaluation of ANT Oxidation in Time
3. Materials and Methods
3.1. Recombinant Expression
3.2. Enzyme Assay
3.3. Spectroscopic Analysis
3.4. Effect of Inhibitors
3.5. Oxidation of Polycyclic Aromatic Hydrocarbons by FNTL
Effect of pH and Incubation Time on Anthracene Oxidation by FNTL
3.6. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arregui, L.; Ayala, M.; Gómez-Gil, X.; Gutiérrez-Soto, G.; Hernández-Luna, C.E.; Herrera De Los Santos, M.; Levin, L.; Rojo-Domínguez, A.; Romero-Martínez, D.; Saparrat, M.C.N.; et al. Laccases: Structure, Function, and Potential Application in Water Bioremediation. Microb. Cell Factories 2019, 18, 1–33. [Google Scholar] [CrossRef] [PubMed]
- Alcalde, M. Laccases: Biological Functions, Molecular Structure and Industrial Applications. In Industrial Enzymes Structure, Function and Applications; Polaina, J., MacCabe, A.P., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 461–476. [Google Scholar] [CrossRef]
- Singh, G.; Arya, S.K. Utility of Laccase in Pulp and Paper Industry: A Progressive Step towards the Green Technology. Int. J. Biol. Macromol. 2019, 134, 1070–1084. [Google Scholar] [CrossRef] [PubMed]
- Unuofin, J.O.; Falade, A.O.; Aladekoyi, O.J. Applications of Microbial Laccases in Bioremediation of Environmental Pollutants: Potential Issues, Challenges, and Prospects. In Bioremediation for Environmental Sustainability; Elsevier: Amsterdam, The Netherlands, 2021; pp. 519–540. [Google Scholar] [CrossRef]
- Viswanath, B.; Rajesh, B.; Janardhan, A.; Kumar, A.P.; Narasimha, G. Fungal Laccases and Their Applications in Bioremediation. Enzyme Res. 2014, 2014, 163242. [Google Scholar] [CrossRef] [PubMed]
- Kudanga, T.; le Roes-Hill, M. Laccase Applications in Biofuels Production: Current Status and Future Prospects. Appl. Microbiol. Biotechnol. 2014, 98, 6525–6542. [Google Scholar] [CrossRef]
- Guan, Z.B.; Luo, Q.; Wang, H.R.; Chen, Y.; Liao, X.R. Bacterial Laccases: Promising Biological Green Tools for Industrial Applications. Cell. Mol. Life Sci. 2018, 75, 3569–3592. [Google Scholar] [CrossRef]
- Loi, M.; Glazunova, O.; Fedorova, T.; Logrieco, A.F.; Mulè, G. Fungal Laccases: The Forefront of Enzymes for Sustainability. J. Fungi 2021, 7, 1048. [Google Scholar] [CrossRef]
- Alegbeleye, O.O.; Opeolu, B.O.; Jackson, V.A. Polycyclic Aromatic Hydrocarbons: A Critical Review of Environmental Occurrence and Bioremediation. Environ. Manag. 2017, 60, 758–783. [Google Scholar] [CrossRef]
- Kuśmierz, M.; Oleszczuk, P.; Kraska, P.; Pałys, E.; Andruszczak, S. Persistence of Polycyclic Aromatic Hydrocarbons (PAHs) in Biochar-Amended Soil. Chemosphere 2016, 146, 272–279. [Google Scholar] [CrossRef]
- Punnapayak, H.; Prasongsuk, S.; Messner, K.; Danmek, K.; Lotrakul, P. Polycyclic Aromatic Hydrocarbons (PAHs) Degradation by Laccase from a Tropical White Rot Fungus Ganoderma Lucidum. Afr. J. Biotechnol. 2009, 8, 5897–5900. [Google Scholar] [CrossRef]
- Zeng, J.; Lin, X.; Zhang, J.; Li, X.; Wong, M.H. Oxidation of Polycyclic Aromatic Hydrocarbons by the Bacterial Laccase CueO from E. coli. Appl. Microbiol. Biotechnol. 2011, 89, 1841–1849. [Google Scholar] [CrossRef]
- Sharma, N.; Leung, I.K.H. Novel Thermophilic Bacterial Laccase for the Degradation of Aromatic Organic Pollutants. Front. Chem. 2021, 9, 711345. [Google Scholar] [CrossRef]
- Bai, Y.; Ali, S.; Liu, S.; Zhou, J.; Tang, Y. Characterization of Plant Laccase Genes and Their Functions. Gene 2023, 852, 147060. [Google Scholar] [CrossRef]
- Laufer, Z.; Beckett, R.P.; Minibayeva, F.V.; Luthje, S.; Bottger, M. Diversity of Laccases from Lichens in Suborder Peltigerineae. Bryologist 2009, 112, 418–426. [Google Scholar] [CrossRef]
- Li, Q.; Wang, X.; Korzhev, M.; Schröder, H.C.; Link, T.; Tahir, M.N.; Diehl-Seifert, B.; Müller, W.E.G. Potential Biological Role of Laccase from the Sponge Suberites Domuncula as an Antibacterial Defense Component. Biochim. Biophys. Acta 2015, 1850, 118–128. [Google Scholar] [CrossRef]
- Asano, T.; Seto, Y.; Hashimoto, K.; Kurushima, H. Mini-Review an Insect-Specific System for Terrestrialization: Laccase-Mediated Cuticle Formation. Insect Biochem. Mol. Biol. 2019, 108, 61–70. [Google Scholar] [CrossRef]
- Akram, F.; Ashraf, S.; Haq, I.U.; Shah, F.I.; Aqeel, A. Eminent Industrial and Biotechnological Applications of Laccases from Bacterial Source: A Current Overview. Appl. Biochem. Biotechnol. 2022, 194, 2336–2356. [Google Scholar] [CrossRef]
- Chauhan, P.S.; Goradia, B.; Saxena, A. Bacterial Laccase: Recent Update on Production, Properties and Industrial Applications. 3 Biotech 2017, 7, 323. [Google Scholar] [CrossRef]
- Liu, H.; Cheng, Y.; Du, B.; Tong, C.; Liang, S.; Han, S.; Zheng, S.; Lin, Y. Overexpression of a Novel Thermostable and Chloride-Tolerant Laccase from Thermus Thermophilus SG0.5JP17-16 in Pichia Pastoris and Its Application in Synthetic Dye Decolorization. PLoS ONE 2015, 10, e0119833. [Google Scholar] [CrossRef]
- Wang, T.N.; Zhao, M. A Simple Strategy for Extracellular Production of CotA Laccase in Escherichia coli and Decolorization of Simulated Textile Effluent by Recombinant Laccase. Appl. Microbiol. Biotechnol. 2017, 101, 685–696. [Google Scholar] [CrossRef]
- Fang, Z.; Li, T.; Wang, Q.; Zhang, X.; Peng, H.; Fang, W.; Hong, Y.; Ge, H.; Xiao, Y. A Bacterial Laccase from Marine Microbial Metagenome Exhibiting Chloride Tolerance and Dye Decolorization Ability. Appl. Microbiol. Biotechnol. 2011, 89, 1103–1110. [Google Scholar] [CrossRef]
- Guan, Z.B.; Shui, Y.; Song, C.M.; Zhang, N.; Cai, Y.J.; Liao, X.R. Efficient Secretory Production of CotA-Laccase and Its Application in the Decolorization and Detoxification of Industrial Textile Wastewater. Environ. Sci. Pollut. Res. Int. 2015, 22, 9515–9523. [Google Scholar] [CrossRef] [PubMed]
- Enguita, F.J.; Martins, L.O.; Henriques, A.O.; Carrondo, M.A. Crystal Structure of a Bacterial Endospore Coat Component: A Laccase with Enhanced Thermostability Properties. J. Biol. Chem. 2003, 278, 19416–19425. [Google Scholar] [CrossRef] [PubMed]
- Prins, A.; Kleinsmidt, L.; Khan, N.; Kirby, B.; Kudanga, T.; Vollmer, J.; Pleiss, J.; Burton, S.; le Roes-Hill, M. The Effect of Mutations near the T1 Copper Site on the Biochemical Characteristics of the Small Laccase from Streptomyces coelicolor A3(2). Enzyme Microb. Technol. 2015, 68, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Basheer, S.; Rashid, N.; Ashraf, R.; Akram, M.S.; Siddiqui, M.A.; Imanaka, T.; Akhtar, M. Identification of a Novel Copper-Activated and Halide-Tolerant Laccase in Geobacillus thermopakistaniensis. Extremophiles 2017, 21, 563–571. [Google Scholar] [CrossRef] [PubMed]
- Brander, S.; Mikkelsen, J.D.; Kepp, K.P. TtMCO: A highly thermostable laccase-like multicopper oxidase from the thermophilic Thermobaculum terrenum. J. Mol. Catal. 2015, 112, 59–65. [Google Scholar] [CrossRef]
- Chen, C.Y.; Huang, Y.C.; Wei, C.M.; Meng, M.; Liu, W.H.; Yang, C.H. Properties of the newly isolated extracellular thermo-alkali-stable laccase from thermophilic actinomycetes, Thermobifida fusca and its application in dye intermediates oxidation. AMB Express 2013, 3, 49. [Google Scholar] [CrossRef]
- Miyazaki, K. A Hyperthermophilic Laccase from Thermus thermophilus HB27. Extremophiles 2005, 9, 415–425. [Google Scholar] [CrossRef]
- Navas, L.E.; Martínez, F.D.; Taverna, M.E.; Fetherolf, M.M.; Eltis, L.D.; Nicolau, V.; Estenoz, D.; Campos, E.; Benintende, G.B.; Berretta, M.F. A Thermostable Laccase from Thermus sp. 2.9 and Its Potential for Delignification of Eucalyptus Biomass. AMB Express 2019, 9, 24. [Google Scholar] [CrossRef]
- Verma, A.; Shirkot, P. Purification and Characterization of Thermostable Laccase from Thermophilic Geobacillus thermocatenulatus MS5 and Its Application in Removal of Textiles Dyes. Sch. Acad. J. Biosci. 2014, 2, 479–485. [Google Scholar]
- Rai, R.; Bibra, M.; Chadha, B.S.; Sani, R.K. Enhanced Hydrolysis of Lignocellulosic Biomass with Doping of a Highly Thermostable Recombinant Laccase. Int. J. Biol. Macromol. 2019, 137, 232–237. [Google Scholar] [CrossRef]
- Tripathy, S.; Maiti, N.K. Construction of Geobacillus thermoglucosidasius cDNA Library and Analysis of Genes Expressed in Response to Heat Stress. Mol. Biol. Rep. 2014, 41, 1639–1644. [Google Scholar] [CrossRef]
- Rosano, G.L.; Ceccarelli, E.A. Recombinant Protein Expression in Escherichia coli: Advances and Challenges. Front. Microbiol. 2014, 5, 172. [Google Scholar] [CrossRef]
- Espina, G.; Cáceres-Moreno, P.; Mejías-Navarrete, G.; Ji, M.; Sun, J.; Blamey, J.M. A Novel and Highly Active Recombinant Spore-Coat Bacterial Laccase, Able to Rapidly Biodecolorize Azo, Triarylmethane and Anthraquinonic Dyestuffs. Int. J. Biol. Macromol. 2021, 170, 298–306. [Google Scholar] [CrossRef]
- Jones, S.M.; Solomon, E.I. Electron Transfer and Reaction Mechanism of Laccases. Cell. Mol Life Sci. 2015, 72, 869–883. [Google Scholar] [CrossRef]
- Dedeyan, B.; Klonowska, A.; Tagger, S.; Tron, T.; Iacazio, G.; Gil, G.; Le Petit, J. Biochemical and Molecular Characterization of a Laccase from Marasmius quercophilus. Appl. Environ. Microbiol. 2000, 66, 925–929. [Google Scholar] [CrossRef]
- Koschorreck, K.; Richter, S.M.; Ene, A.B.; Roduner, E.; Schmid, R.D.; Urlacher, V.B. Cloning and Characterization of a New Laccase from Bacillus licheniformis Catalyzing Dimerization of Phenolic Acids. Appl. Microbiol. Biotechnol. 2008, 79, 217–224. [Google Scholar] [CrossRef]
- Siroosi, M.; Amoozegar, M.A.; Khajeh, K. Purification and Characterization of an Alkaline Chloride-Tolerant Laccase from a Halotolerant Bacterium, Bacillus sp. strain WT. J. Mol. Catal. 2016, 134, 89–97. [Google Scholar] [CrossRef]
- Couto, S.; Herrera, L. Inhibitors of Laccases: A Review. Curr. Enzym. Inhib. 2006, 2, 343–352. [Google Scholar] [CrossRef]
- Sharma, B.; Dangi, A.K.; Shukla, P. Contemporary Enzyme Based Technologies for Bioremediation: A Review. J. Environ. Manag. 2018, 210, 10–22. [Google Scholar] [CrossRef]
- Brander, S.; Mikkelsen, J.D.; Kepp, K.P. Characterization of an Alkali- and Halide-Resistant Laccase Expressed in E. coli: CotA from Bacillus clausii. PLoS ONE 2014, 9, e99402. [Google Scholar] [CrossRef]
- Lu, L.; Wang, T.N.; Xu, T.F.; Wang, J.Y.; Wang, C.L.; Zhao, M. Cloning and Expression of Thermo-Alkali-Stable Laccase of Bacillus licheniformis in Pichia pastoris and Its Characterization. Bioresour. Technol. 2013, 134, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wei, Z.; Zhang, M.; Peng, X.; Yu, G.; Teng, M.; Gong, W. Crystal Structures of E. coli Laccase CueO at Different Copper Concentrations. Biochem. Biophys. Res. Commun. 2007, 354, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Johannes, C.; Majcherczyk, A. Laccase Activity Tests and Laccase Inhibitors. J. Biotechnol. 2000, 78, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Zhang, S.; Diao, H.; Zhao, H.; Zhu, X.; Lu, F.; Lu, Z. Purification and characterization of a temperature-and pH-stable laccase from the spores of Bacillus vallismortis fmb-103 and its application in the degradation of malachite green. J. Agric. Food Chem. 2013, 61, 5468–5473. [Google Scholar] [CrossRef]
- Wang, C.; Zhao, M.; Lu, L.; Wei, X.; Li, T. Characterization of spore laccase from Bacillus subtilis WD23 and its use in dye decolorization. Afr. J. Biotechnol. 2011, 10, 2186–2192. [Google Scholar]
- Durão, P.; Chen, Z.; Fernandes, A.T.; Hildebrandt, P.; Murgida, D.H.; Todorovic, S.; Pereira, M.M.; Melo, E.P.; Martins, L.O. Copper Incorporation into Recombinant CotA Laccase from Bacillus subtilis: Characterization of Fully Copper Loaded Enzymes. J. Biol. Inorg. Chem. 2008, 13, 183–193. [Google Scholar] [CrossRef]
- Johannes, C.; Majcherczyk, A.; Hüttermann, A. Degradation of Anthracene by Laccase of Trametes versicolor in the Presence of Different Mediator Compounds. Appl. Microbiol. Biotechnol. 1996, 46, 313–317. [Google Scholar] [CrossRef]
- Zeng, J.; Zhu, Q.; Wu, Y.; Lin, X. Oxidation of Polycyclic Aromatic Hydrocarbons Using Bacillus subtilis CotA with High Laccase Activity and Copper Independence. Chemosphere 2016, 148, 1–7. [Google Scholar] [CrossRef]
- Wang, L.; Tan, Y.; Sun, S.; Zhou, L.; Wu, G.; Shao, Y.; Wang, M.; Xin, Z. Improving Degradation of Polycyclic Aromatic Hydrocarbons by Bacillus atrophaeus Laccase Fused with Vitreoscilla Hemoglobin and a Novel Strong Promoter Replacement. Biology 2022, 11, 1129. [Google Scholar] [CrossRef]
- Alcalde, M.; Bulter, T.; Arnold, F.H. Colorimetric Assays for Biodegradation of Polycyclic Aromatic Hydrocarbons by Fungal Laccases. J. Biomol. Screen. 2002, 7, 547–553. [Google Scholar] [CrossRef]
- Pozdnyakova, N.N.; Rodakiewicz-Nowak, J.; Turkovskaya, O.V.; Haber, J. Oxidative Degradation of Polyaromatic Hydrocarbons Catalyzed by Blue Laccase from Pleurotus ostreatus D1 in the Presence of Synthetic Mediators. Enzyme Microb. Technol. 2006, 39, 1242–1249. [Google Scholar] [CrossRef]
- Xu, P.; Du, H.; Peng, X.; Tang, Y.; Zhou, Y.; Chen, X.; Fei, J.; Meng, Y.; Yuan, L. Degradation of Several Polycyclic Aromatic Hydrocarbons by Laccase in Reverse Micelle System. Sci. Total Environ. 2020, 708, 134970. [Google Scholar] [CrossRef]
- Peng, X.; Yuan, X.Z.; Liu, H.; Zeng, G.M.; Chen, X.H. Degradation of Polycyclic Aromatic Hydrocarbons (PAHs) by Laccase in Rhamnolipid Reversed Micellar System. Appl. Biochem. Biotechnol. 2015, 176, 45–55. [Google Scholar] [CrossRef]
- Deng, J.; Wang, H.; Zhan, H.; Wu, C.; Huang, Y.; Yang, B.; Mosa, A.; Ling, W. Catalyzed Degradation of Polycyclic Aromatic Hydrocarbons by Recoverable Magnetic Chitosan Immobilized Laccase from Trametes versicolor. Chemosphere 2022, 301, 134753. [Google Scholar] [CrossRef]
- Cañas, A.I.; Alcalde, M.; Plou, F.; Martínez, M.J.; Martínez, Á.T.; Camarero, S. Transformation of Polycyclic Aromatic Hydrocarbons by Laccase Is Strongly Enhanced by Phenolic Compounds Present in Soil. Environ. Sci. Technol 2007, 41, 2964–2971. [Google Scholar] [CrossRef]
- Ike, P.T.L.; Birolli, W.G.; dos Santos, D.M.; Porto, A.L.M.; Souza, D.H.F. Biodegradation of Anthracene and Different PAHs by a Yellow Laccase from Leucoagaricus gongylophorus. Environ. Sci. Pollut. Res. Int. 2019, 26, 8675–8684. [Google Scholar] [CrossRef]
- Hu, X.; Zhang, Y.; Zhao, X.; Hwang, H.M. Biodegradation of Benzo[a]Pyrene with Immobilized Laccase: Genotoxicity of the Products in HaCaT and A3 Cells. Environ. Mol. Mutagen 2007, 48, 106–113. [Google Scholar] [CrossRef]
- Kadri, T.; Rouissi, T.; Kaur Brar, S.; Cledon, M.; Sarma, S.; Verma, M. Biodegradation of Polycyclic Aromatic Hydrocarbons (PAHs) by Fungal Enzymes: A Review. J. Environ. Sci. 2017, 51, 52–74. [Google Scholar] [CrossRef]
- Espina, G.; Muñoz-Ibacache, S.A.; Cáceres-Moreno, P.; Amenabar, M.J.; Blamey, J.M. From the Discovery of Extremozymes to an Enzymatic Product: Roadmap Based on Their Applications. Front. Bioeng. Biotechnol. 2022, 9, 1271. [Google Scholar] [CrossRef]
- Gallagher, S.; Sasse, J. Protein Analysis by SDS-PAGE and Detection by Coomassie Blue or Silver Staining. Curr. Protoc. Pharmacol. 2001, 2, A.3B.1–A.3B.10. [Google Scholar] [CrossRef]
- Bradford, M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Lante, A.; Crapisi, A.; Krastanov, A.; Spettoli, P. Biodegradation of Phenols by Laccase Immobilised in a Membrane Reactor. Process Biochem. 2000, 36, 51–58. [Google Scholar] [CrossRef]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein Identification and Analysis Tools on the ExPASy Server. In The Proteomics Protocols Handbook; Walker, J.M., Ed.; Humana Press: New York, NY, USA, 2005; pp. 571–607. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bueno-Nieto, C.; Cortés-Antiquera, R.; Espina, G.; Atalah, J.; Villanueva, J.; Aliaga, C.; Zuñiga, G.E.; Blamey, J.M. Biochemical and Spectroscopic Characterization of a Recombinant Laccase from Thermoalkaliphilic Bacillus sp. FNT with Potential for Degradation of Polycyclic Aromatic Hydrocarbons (PAHs). Catalysts 2023, 13, 763. https://doi.org/10.3390/catal13040763
Bueno-Nieto C, Cortés-Antiquera R, Espina G, Atalah J, Villanueva J, Aliaga C, Zuñiga GE, Blamey JM. Biochemical and Spectroscopic Characterization of a Recombinant Laccase from Thermoalkaliphilic Bacillus sp. FNT with Potential for Degradation of Polycyclic Aromatic Hydrocarbons (PAHs). Catalysts. 2023; 13(4):763. https://doi.org/10.3390/catal13040763
Chicago/Turabian StyleBueno-Nieto, Constanza, Rodrigo Cortés-Antiquera, Giannina Espina, Joaquín Atalah, Javiera Villanueva, Carolina Aliaga, Gustavo E. Zuñiga, and Jenny M. Blamey. 2023. "Biochemical and Spectroscopic Characterization of a Recombinant Laccase from Thermoalkaliphilic Bacillus sp. FNT with Potential for Degradation of Polycyclic Aromatic Hydrocarbons (PAHs)" Catalysts 13, no. 4: 763. https://doi.org/10.3390/catal13040763