
Transcriptomic Analysis of Differentially Expressed Genes in Arabidopsis thaliana Overexpressing BnMYB2 from Boehmeria nivea under Cadmium Stress

Abstract
:1. Introduction
2. Results
2.1. The Quality Analysis of Transcriptome
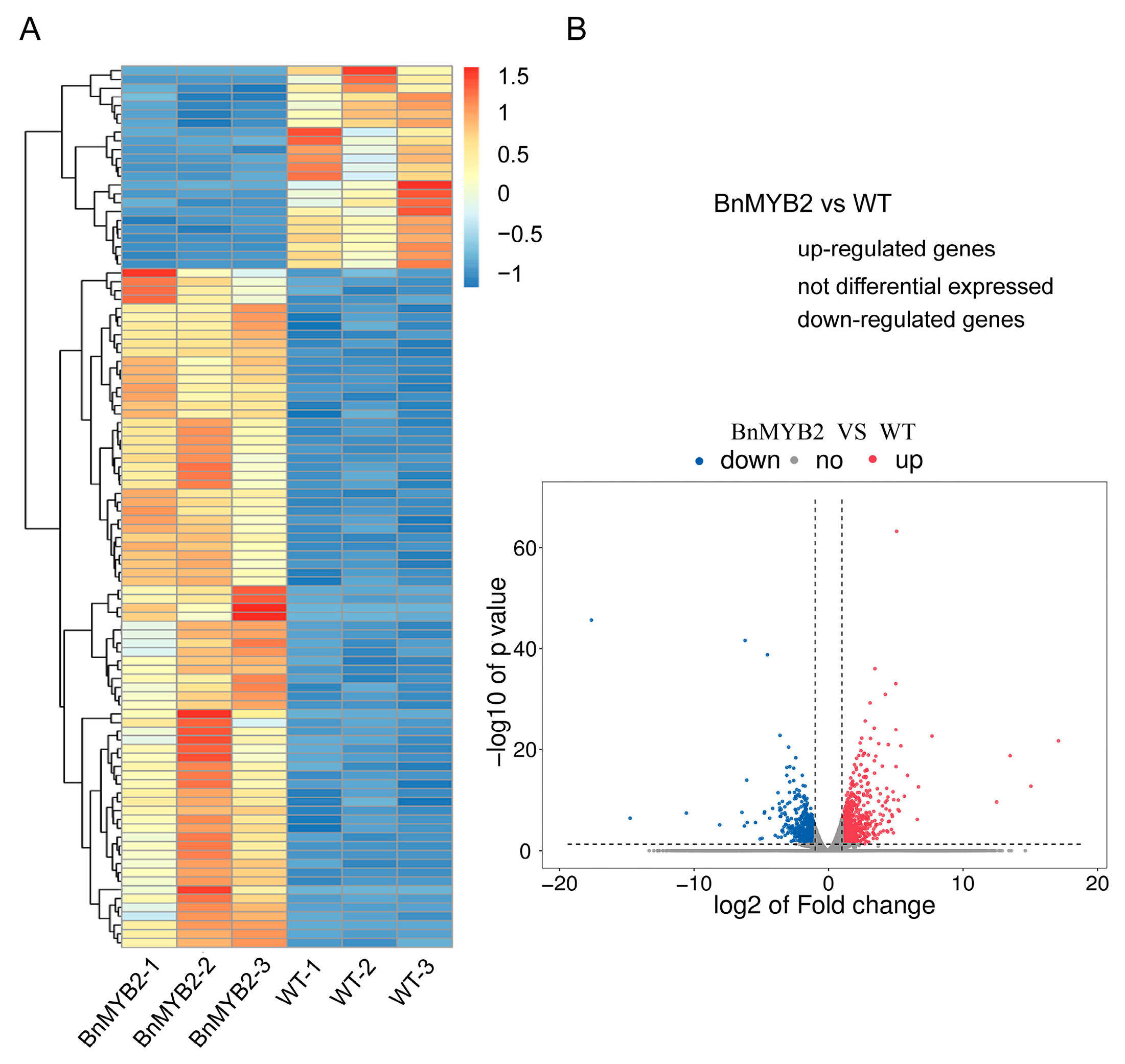
2.2. Differential Expression Genes (DEGs) Analysis
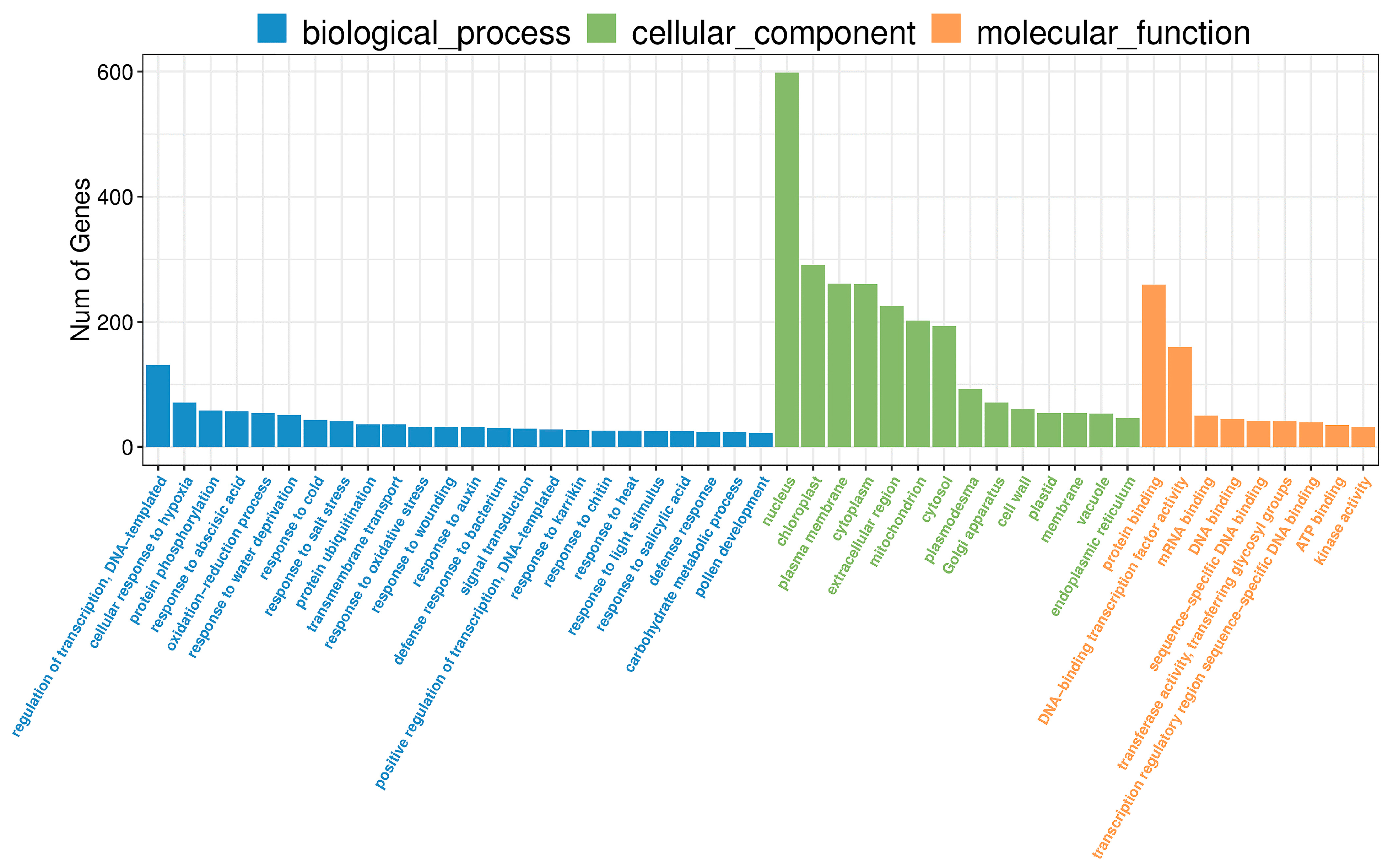
2.3. GO Analysis of DEGs
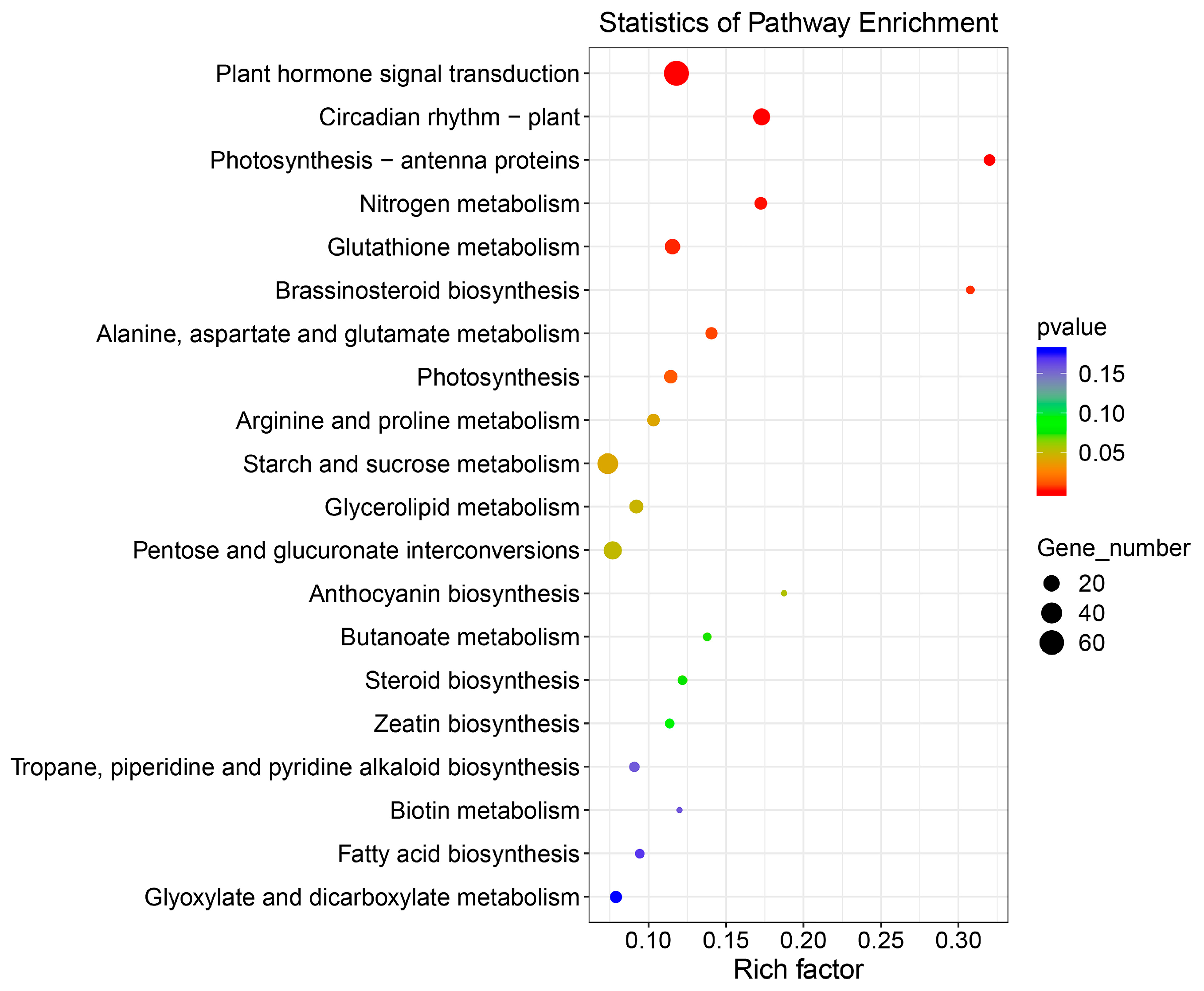
2.4. KEGG Pathway Analysis of DEGs
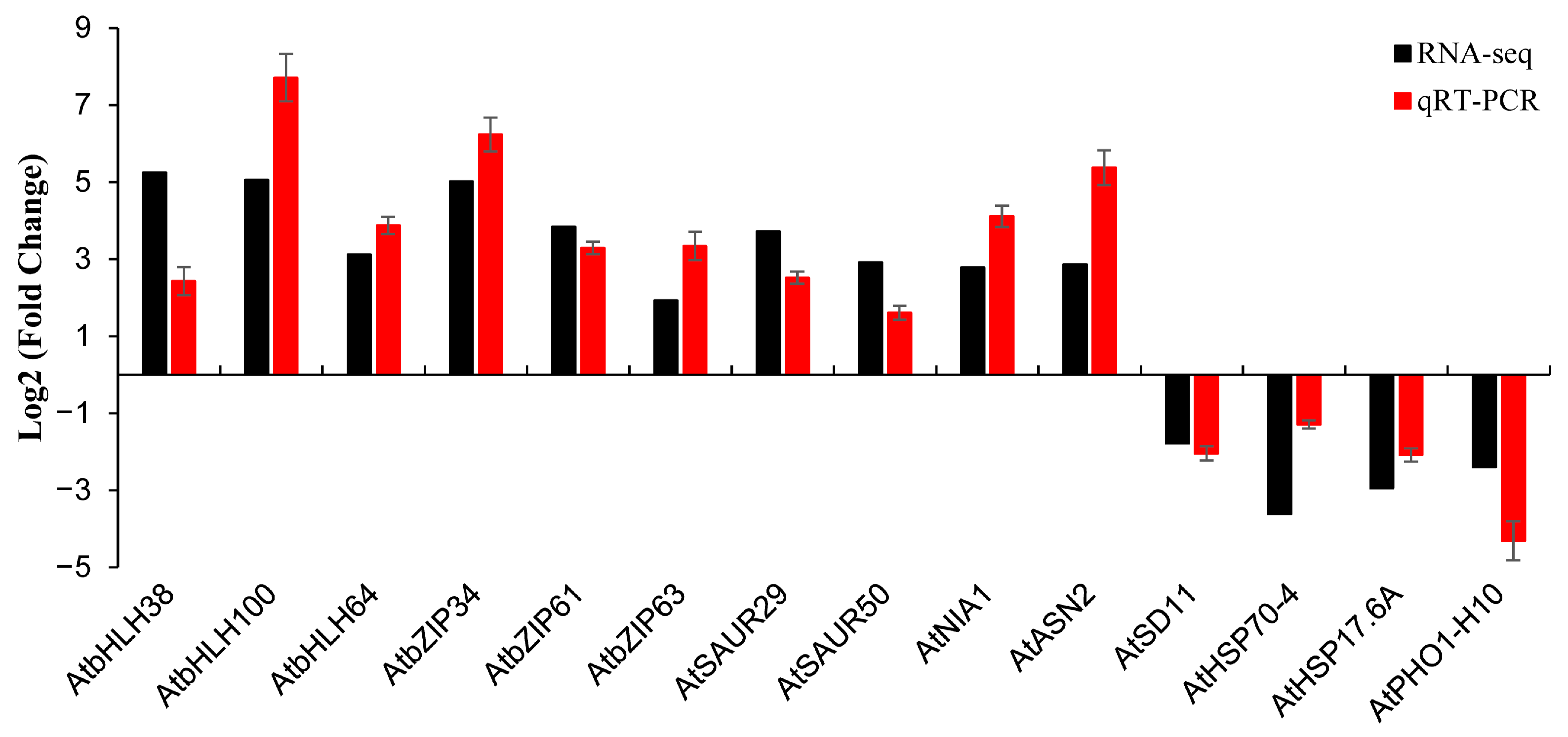
2.5. Verification of RNA-Seq Data
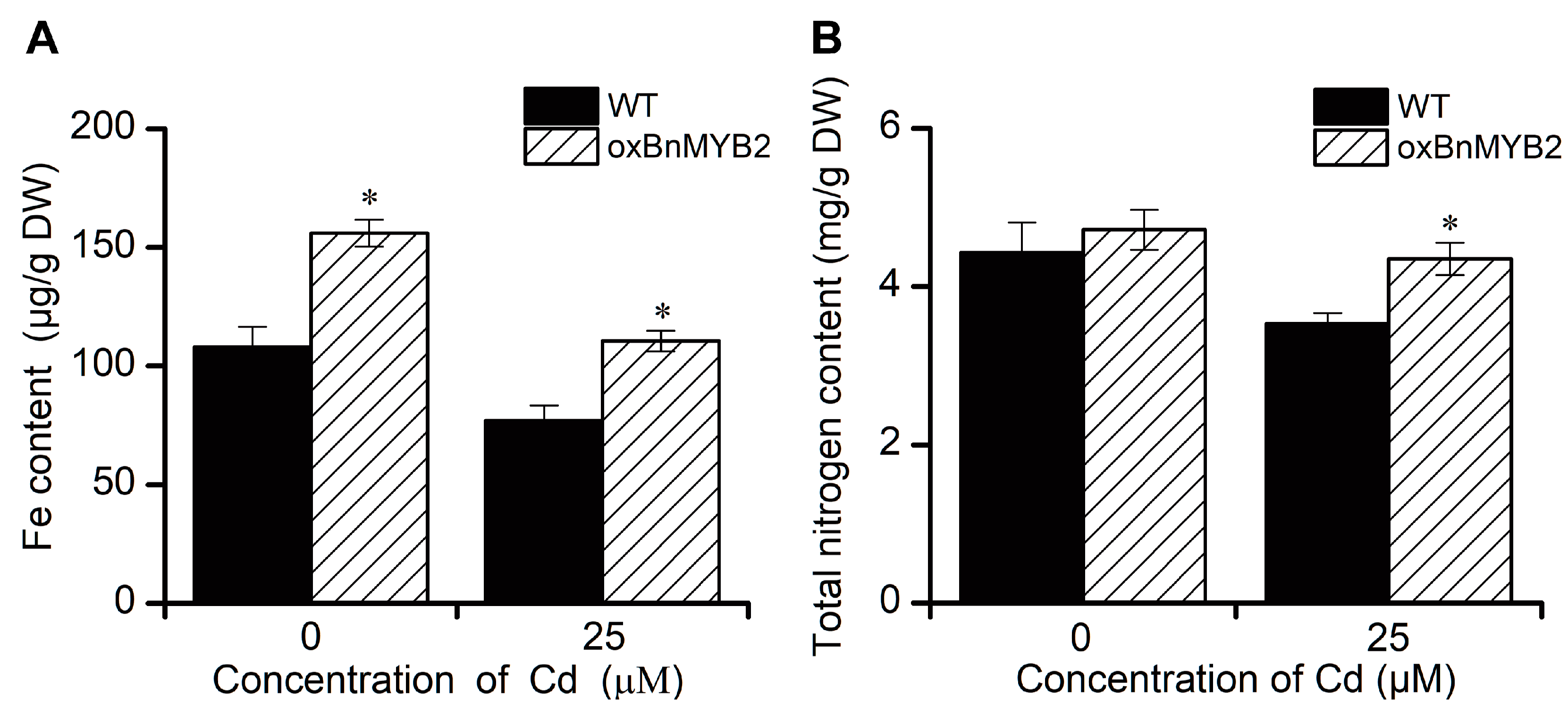
2.6. BnMYB2 Transgenic Arabidopsis Plants Showed Improved Fe and Nitrogen Content in Shoots
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Condition
4.2. RNA Sequencing and Differential Gene Expression Analysis
4.3. Validation of DEGs by qRT-PCR
4.4. Determination of Iron and Nitrogen Content
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Xia, X.; Wu, S.; Zhou, Z.; Wang, G. Microbial Cd(II) and Cr(VI) resistance mechanisms and application in bioremediation. J. Hazard. Mater. 2021, 401, 123685. [Google Scholar] [CrossRef] [PubMed]
- Clemens, S.; Aarts, M.G.; Thomine, S.; Verbruggen, N. Plant science: The key to preventing slow cadmium poisoning. Trends Plant Sci. 2013, 18, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhu, Y.; Yu, L.; Yang, M.; Zou, X.; Yin, C.; Lin, Y. Research Advances in Cadmium Uptake, Transport and Resistance in Rice (Oryza sativa L.). Cells 2022, 11, 569. [Google Scholar] [CrossRef]
- Dubos, C.; Stracke, R.; Grotewold, E.; Weisshaar, B.; Martin, C.; Lepiniec, L. MYB transcription factors in Arabidopsis. Trends Plant Sci. 2010, 15, 573–581. [Google Scholar] [CrossRef]
- Ai, T.N.; Naing, A.H.; Yun, B.-W.; Lim, S.H.; Kim, C.K. Overexpression of RsMYB1 Enhances Anthocyanin Accumulation and Heavy Metal Stress Tolerance in Transgenic Petunia. Front. Plant Sci. 2018, 9, 1388. [Google Scholar] [CrossRef] [Green Version]
- Sapara, K.K.; Khedia, J.; Agarwal, P.; Gangapur, D.R.; Agarwal, P.K. SbMYB15 transcription factor mitigates cadmium and nickel stress in transgenic tobacco by limiting uptake and modulating antioxidative defence system. Funct. Plant Biol. 2019, 46, 702–714. [Google Scholar] [CrossRef]
- Zhang, P.; Wang, R.; Ju, Q.; Li, W.; Tran, L.-S.P.; Xu, J. The R2R3-MYB Transcription Factor MYB49 Regulates Cadmium Accumulation. Plant Physiol. 2019, 180, 529–542. [Google Scholar] [CrossRef]
- Zhu, S.J.; Shi, W.J.; Jie, Y.C.; Zhou, Q.M. Cloning and expression analysis of cadmium-responsive transcription factor gene BnMYB1 from ramie (Boehmeria nivea). J. Agric. Biotechnol. 2018, 26, 774–783. [Google Scholar]
- Zhu, S.; Shi, W.; Jie, Y.; Zhou, Q.; Song, C. A MYB transcription factor, BnMYB2, cloned from ramie (Boehmeria nivea) is involved in cadmium tolerance and accumulation. PLoS ONE 2020, 15, e0233375. [Google Scholar] [CrossRef]
- Zhu, S.J.; Shi, W.J. Cloning and expression pattern analysis of BnMYB3 transcription factor gene in ramie. Acta Bot. Boreali-Occident. Sin. 2019, 39, 422–429. [Google Scholar]
- Lei, M.; Yue, Q.; Chen, T.; Huang, Z.; Liao, X.; Liu, Y.; Zheng, G.; Chang, Q. Heavy metal concentrations in soils and plants around Shizhuyuan mining area of Hu’nan province. Acta Ecol. Sin. 2005, 25, 1146–1151. [Google Scholar]
- Zhu, S.; Shi, W.; Zhang, J. Effect of different ramie (Boehmeria nivea L. Gaud) cultivars on the adsorption of heavy metal ions cadmium and lead in the remediation of contaminated farmland soils. Open Chem. 2022, 20, 444–454. [Google Scholar] [CrossRef]
- She, W.; Jie, Y.; Xing, H.; Lu, Y.; Kang, W.; Wang, D. Heavy metal concentrations and bioaccumulation of ramie (Boehmeria nivea) growing on 3 mining areas in Shimen, Lengshuijiang and Liuyang of Hunan province. Acta Ecol. Sin. 2011, 31, 874–881. [Google Scholar]
- Sandalio, L.; Dalurzo, H.; Gómez, M.; Romero-Puertas, M.; del Río, L. Cadmium-induced changes in the growth and oxidative metabolism of pea plants. J. Exp. Bot. 2001, 52, 2115–2126. [Google Scholar] [CrossRef]
- Zhang, G.; Fukami, M.; Sekimoto, H. Influence of cadmium on mineral concentrations and yield components in wheat genotypes differing in Cd tolerance at seedling stage. Field Crops Res. 2002, 77, 93–98. [Google Scholar] [CrossRef]
- Liu, J.; Liang, J.; Li, K.; Zhang, Z.; Yu, B.; Lu, X.; Yang, J.; Zhu, Q. Correlations between cadmium and mineral nutrients in absorption and accumulation in various genotypes of rice under cadmium stress. Chemosphere 2003, 52, 1467–1473. [Google Scholar] [CrossRef]
- Wu, H.; Chen, C.; Du, J.; Liu, H.; Cui, Y.; Zhang, Y.; He, Y.; Wang, Y.; Chu, C.; Feng, Z.; et al. Co-Overexpression FIT with AtbHLH38 or AtbHLH39 in Arabidopsis-Enhanced Cadmium Tolerance via Increased Cadmium Sequestration in Roots and Improved Iron Homeostasis of Shoots. Plant Physiol. 2012, 158, 790–800. [Google Scholar] [CrossRef] [Green Version]
- Shao, G.; Chen, M.; Wang, W.; Mou, R.; Zhang, G. Iron nutrition affects cadmium accumulation and toxicity in rice plants. Plant Growth Regul. 2007, 53, 33–42. [Google Scholar] [CrossRef]
- Yuan, Y.; Wu, H.; Wang, N.; Li, J.; Zhao, W.; Du, J.; Wang, D.; Ling, H.-Q. FIT interacts with AtbHLH38 and AtbHLH39 in regulating iron uptake gene expression for iron homeostasis in Arabidopsis. Cell Res. 2008, 18, 385–397. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Cui, Y.; Liu, Y.; Fan, H.; Du, J.; Huang, Z.; Yuan, Y.; Wu, H.; Ling, H.-Q. Requirement and Functional Redundancy of Ib Subgroup bHLH Proteins for Iron Deficiency Responses and Uptake in Arabidopsis thaliana. Mol. Plant 2013, 6, 503–513. [Google Scholar] [CrossRef] [Green Version]
- Feller, A.; Machemer, K.; Braun, E.L.; Grotewold, E. Evolutionary and comparative analysis of MYB and bHLH plant transcription factors. Plant J. 2011, 66, 94–116. [Google Scholar] [CrossRef] [PubMed]
- Millard, P.S.; Weber, K.; Kragelund, B.B.; Burow, M. Specificity of MYB interactions relies on motifs in ordered and disordered contexts. Nucleic Acids Res. 2019, 47, 9592–9608. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Chen, X.; Chai, S.; Sheng, H.; Sha, L.; Fan, X.; Zeng, J.; Kang, H.; Zhang, H.; Xiao, X.; et al. TpIRT1 from Polish wheat (Triticum polonicum L.) enhances the accumulation of Fe, Mn, Co, and Cd in Arabidopsis. Plant Sci. 2021, 312, 111058. [Google Scholar] [CrossRef] [PubMed]
- Nishida, S.; Tsuzuki, C.; Kato, A.; Aisu, A.; Yoshida, J.; Mizuno, T. AtIRT1, the Primary Iron Uptake Transporter in the Root, Mediates Excess Nickel Accumulation in Arabidopsis thaliana. Plant Cell Physiol. 2011, 52, 1433–1442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, L.; Qu, M.; Zhu, Y.; Peng, C.; Wang, J.; Gao, D.; Chen, C. ZINC TRANSPORTER5 and ZINC TRANSPORTER9 Function Synergistically in Zinc/Cadmium Uptake. Plant Physiol. 2020, 183, 1235–1249. [Google Scholar] [CrossRef]
- Inaba, S.; Kurata, R.; Kobayashi, M.; Yamagishi, Y.; Mori, I.; Ogata, Y.; Fukao, Y. Identification of putative target genes of bZIP19, a transcription factor essential for Arabidopsis adaptation to Zn deficiency in roots. Plant J. 2015, 84, 323–334. [Google Scholar] [CrossRef] [Green Version]
- Assunção, A.G.L.; Herrero, E.; Lin, Y.-F.; Huettel, B.; Talukdar, S.; Smaczniak, C.; Immink, R.G.H.; Van Eldik, M.; Fiers, M.; Schat, H.; et al. Arabidopsis thaliana transcription factors bZIP19 and bZIP23 regulate the adaptation to zinc deficiency. Proc. Natl. Acad. Sci. USA 2010, 107, 10296–10301. [Google Scholar] [CrossRef] [Green Version]
- Xian, J.; Wang, Y.; Niu, K.; Ma, H.; Ma, X. Transcriptional regulation and expression network responding to cadmium stress in a Cd-tolerant perennial grass Poa Pratensis. Chemosphere 2020, 250, 126158. [Google Scholar] [CrossRef]
- Chen, S.; Xie, M.; Cui, M.; Li, W.; Xu, Z.; Jia, C.; Yang, G. Identification of Broussonetia papyrifera transcription Factor BpbZIP1 and analysis of its response to cadmium stress. Bull. Bot. Res. 2022, 42, 394–402. [Google Scholar]
- Huang, C.; Zhou, J.; Jie, Y.; Xing, H.; Zhong, Y.; Yu, W.; She, W.; Ma, Y.; Liu, Z.; Zhang, Y. A Ramie bZIP transcription factor BnbZIP2 is involved in drought, salt, and heavy metal stress response. DNA Cell Biol. 2016, 35, 776–786. [Google Scholar] [CrossRef]
- Jakoby, M.; Weisshaar, B.; Dröge-Laser, W.; Vicente-Carbajosa, J.; Tiedemann, J.; Kroj, T.; Parcy, F. bZIP transcription factors in Arabidopsis. Trends Plant Sci. 2002, 7, 106–111. [Google Scholar] [CrossRef]
- Joo, J.; Lee, Y.H.; Song, S.I. Overexpression of the rice basic leucine zipper transcription factor OsbZIP12 confers drought tolerance to rice and makes seedlings hypersensitive to ABA. Plant Biotechnol. Rep. 2014, 8, 431–441. [Google Scholar] [CrossRef]
- Hobo, T.; Kowyama, Y.; Hattori, T. A bZIP factor, TRAB1, interacts with VP1 and mediates abscisic acid-induced transcription. Proc. Natl. Acad. Sci. USA 1999, 96, 15348–15353. [Google Scholar] [CrossRef] [Green Version]
- Lu, G.; Gao, C.; Zheng, X.; Han, B. Identification of OsbZIP72 as a positive regulator of ABA response and drought tolerance in rice. Planta 2009, 229, 605–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, N.; Zhang, H.; Li, X.; Xiao, J.; Xiong, L. Constitutive Activation of Transcription Factor OsbZIP46 Improves Drought Tolerance in Rice. Plant Physiol. 2012, 158, 1755–1768. [Google Scholar] [CrossRef] [Green Version]
- Xiang, Y.; Tang, N.; Du, H.; Ye, H.; Xiong, L. Characterization of OsbZIP23 as a Key Player of the Basic Leucine Zipper Transcription Factor Family for Conferring Abscisic Acid Sensitivity and Salinity and Drought Tolerance in Rice. Plant Physiol. 2008, 148, 1938–1952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matiolli, C.C.; Tomaz, J.P.; Duarte, G.T.; Prado, F.M.; Del Bem, L.E.V.; Silveira, A.B.; Gauer, L.; Corrêa, L.G.G.; Drumond, R.D.; Viana, A.J.C.; et al. The Arabidopsis bZIP Gene AtbZIP63 Is a Sensitive Integrator of Transient Abscisic Acid and Glucose Signals. Plant Physiol. 2011, 157, 692–705. [Google Scholar] [CrossRef] [Green Version]
- Mathur, P.; Mukherjee, S. Molecular mechanisms of auxin mediated regulation of heavy metal and metalloid stress in plants. Environ. Exp. Bot. 2022, 196, 104796. [Google Scholar] [CrossRef]
- Matthes, M.S.; Best, N.B.; Robil, J.M.; Malcomber, S.; Gallavotti, A.; McSteen, P. Auxin EvoDevo: Conservation and Diversification of Genes Regulating Auxin Biosynthesis, Transport, and Signaling. Mol. Plant 2018, 12, 298–320. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Xu, C.-B.; Sun, X.-J.; Hu, Z.; Fan, S.-J.; Jiang, Q.-Y.; Zhang, H. TaSAUR78 enhances multiple abiotic stress tolerance by regulating the interacting gene TaVDAC1. J. Integr. Agric. 2019, 18, 2682–2690. [Google Scholar] [CrossRef]
- He, Y.; Liu, Y.; Li, M.; Lamin-Samu, A.T.; Yang, D.; Yu, X.; Izhar, M.; Jan, I.; Ali, M.; Lu, G. The Arabidopsis SMALL AUXIN UP RNA32 Protein Regulates ABA-Mediated Responses to Drought Stress. Front. Plant Sci. 2021, 12, 625493. [Google Scholar] [CrossRef]
- Chen, F.; Wang, R.-J.; Wu, C.-J.; Lin, M.; Yan, H.-W.; Xiang, Y. SAUR8, a small auxin-up RNA gene in poplar, confers drought tolerance to transgenic Arabidopsis plants. Gene 2022, 837, 146692. [Google Scholar] [CrossRef] [PubMed]
- Qiu, T.; Qi, M.; Ding, X.; Zheng, Y.; Zhou, T.; Chen, Y.; Han, N.; Zhu, M.; Bian, H.; Wang, J. The SAUR41 subfamily of SMALL AUXIN UP RNA genes is abscisic acid inducible to modulate cell expansion and salt tolerance in Arabidopsis thaliana seedlings. Ann. Bot. 2020, 125, 805–819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chauhan, R.; Awasthi, S.; Indoliya, Y.; Chauhan, A.S.; Mishra, S.; Agrawal, L.; Srivastava, S.; Dwivedi, S.; Singh, P.C.; Mallick, S.; et al. Transcriptome and proteome analyses reveal selenium mediated amelioration of arsenic toxicity in rice (Oryza sativa L.). J. Hazard. Mater. 2020, 390, 122122. [Google Scholar] [CrossRef] [PubMed]
- Meng, L.; Yang, Y.; Ma, Z.; Jiang, J.; Zhang, X.; Chen, Z.; Cui, G.; Yin, X. Integrated physiological, transcriptomic and metabolomic analysis of the response of Trifolium pratense L. to Pb toxicity. J. Hazard. Mater. 2022, 436, 129128. [Google Scholar] [CrossRef]
- Hernandez, L.E.; Gárate, A.; Carpena-Ruiz, R. Effects of cadmium on the uptake, distribution and assimilation of nitrate in Pisum sativum. Plant Soil 1997, 189, 97–106. [Google Scholar] [CrossRef]
- Khan, M.I.R.; Iqbal, N.; Masood, A.; Mobin, M.; Anjum, N.A.; Khan, N.A. Modulation and significance of nitrogen and sulfur metabolism in cadmium challenged plants. Plant Growth Regul. 2016, 78, 1–11. [Google Scholar] [CrossRef]
- He, F.; Zhao, Q.; Huang, J.-L.; Niu, M.-X.; Feng, H.-C.; Shi, Y.-J.; Zhao, K.-J.; Cui, X.-L.; Wu, X.-L.; Mi, J.-X.; et al. External application of nitrogen alleviates toxicity of cadmium on poplars via starch and sucrose metabolism. Tree Physiol. 2021, 41, 2126–2141. [Google Scholar] [CrossRef]
- Huang, J.; Wu, X.; Tian, F.; Chen, Q.; Luo, P.; Zhang, F.; Wan, X.; Zhong, Y.; Liu, Q.; Lin, T. Changes in Proteome and Protein Phosphorylation Reveal the Protective Roles of Exogenous Nitrogen in Alleviating Cadmium Toxicity in Poplar Plants. Int. J. Mol. Sci. 2019, 21, 278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balotf, S.; Kavoosi, G.; Kholdebarin, B. Nitrate reductase, nitrite reductase, glutamine synthetase, and glutamate synthase expression and activity in response to different nitrogen sources in nitrogen-starved wheat seedlings. Biotechnol. Appl. Biochem. 2016, 63, 220–229. [Google Scholar] [CrossRef]
- Campbell, W.H. Nitrate reductase and its role in nitrate assimilation in plants. Physiol. Plant. 2006, 74, 214–219. [Google Scholar] [CrossRef]
- Singh, P.K.; Indoliya, Y.; Chauhan, A.S.; Singh, S.P.; Singh, A.P.; Dwivedi, S.; Tripathi, R.D.; Chakrabarty, D. Nitric oxide mediated transcriptional modulation enhances plant adaptive responses to arsenic stress. Sci. Rep. 2017, 7, 3592. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; You, J.; Liang, X. Nitrate reductase-mediated nitric oxide production is involved in copper tolerance in shoots of hulless barley. Plant Cell Rep. 2015, 34, 367–379. [Google Scholar] [CrossRef]
- Emamverdian, A.; Ding, Y.; Barker, J.; Mokhberdoran, F.; Ramakrishnan, M.; Liu, G.; Li, Y. Nitric Oxide Ameliorates Plant Metal Toxicity by Increasing Antioxidant Capacity and Reducing Pb and Cd Translocation. Antioxidants 2021, 10, 1981. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Yin, H.; Li, Y.; Liu, X. Nitric Oxide Is Associated with Long-Term Zinc Tolerance in Solanum nigrum. Plant Physiol. 2010, 154, 1319–1334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gill, S.S.; Hasanuzzaman, M.; Nahar, K.; Macovei, A.; Tuteja, N. Importance of nitric oxide in cadmium stress tolerance in crop plants. Plant Physiol. Biochem. 2013, 63, 254–261. [Google Scholar] [CrossRef]
- Liu, X.; Hu, B.; Chu, C. Nitrogen assimilation in plants: Current status and future prospects. J. Genet. Genom. 2022, 49, 394–404. [Google Scholar] [CrossRef]
- Lynch, J.M.; Barbano, D.M. Kjeldahl Nitrogen Analysis as a Reference Method for Protein Determination in Dairy Products. J. AOAC Int. 1999, 82, 1389–1398. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Raw Reads (bp) | Clean Reads (bp) | Clean Bases (Gb) | Valid Ratio (%) | Mapped Reads (%) | Q20 (%) | Q30 (%) |
|---|---|---|---|---|---|---|---|
| WT-1 | 42150324 | 39690828 | 5.95 | 94.16 | 97.40 | 99.99 | 98.59 |
| WT-2 | 50317342 | 47695500 | 7.15 | 94.79 | 97.43 | 99.99 | 98.62 |
| WT-3 | 47497386 | 45386300 | 6.81 | 95.56 | 97.39 | 99.99 | 98.67 |
| BnMYB2-1 | 50881806 | 48696066 | 7.30 | 95.70 | 96.87 | 99.99 | 98.57 |
| BnMYB2-2 | 54783260 | 52579700 | 7.89 | 95.98 | 97.04 | 99.99 | 98.68 |
| BnMYB2-3 | 51195792 | 48957544 | 7.34 | 95.63 | 97.03 | 99.99 | 98.65 |
| Gene ID | Symbol | Fold Change | log2 (FC) | p-Value | Regulation |
|---|---|---|---|---|---|
| bHLH transcription factors | |||||
| AT3G56970 | AtbHLH38 | 38.15 | 5.25 | 1.45 × 10−8 | up |
| AT2G41240 | AtbHLH100 | 33.40 | 5.06 | 6.17 × 10−64 | up |
| AT3G56980 | AtbHLH39 | 32.14 | 5.01 | 1.21 × 10−24 | up |
| AT2G18300 | AtbHLH64 | 8.70 | 3.12 | 7.06 × 10−23 | up |
| AT5G50915 | AtbHLH137 | 7.74 | 2.95 | 5.45 × 10−7 | up |
| AT4G01460 | AtbHLH57 | 3.26 | 1.71 | 3.30 × 10−4 | up |
| AT5G04150 | AtbHLH101 | 2.54 | 1.34 | 2.04 × 10−3 | up |
| AT3G07340 | AtbHLH62 | 2.54 | 1.34 | 2.37 × 10−5 | up |
| AT2G31730 | AtbHLH154 | 2.30 | 1.20 | 9.79 × 10−4 | up |
| AT1G02340 | AtbHLH26 | 2.10 | 1.07 | 2.34 × 10−3 | up |
| bZIP transcription factors | |||||
| AT2G42380 | AtbZIP34 | 32.44 | 5.02 | 2.49 × 10−17 | up |
| AT3G58120 | AtbZIP61 | 14.33 | 3.84 | 6.07 × 10−12 | up |
| AT5G28770 | AtbZIP63 | 3.80 | 1.93 | 1.06 × 10−17 | up |
| Gene ID | Symbol | Fold Change | log2 (FC) | p-Value | Regulation |
|---|---|---|---|---|---|
| Plant hormone signal transduction | |||||
| AT3G03820 | AtSAUR29 | 13.21 | 3.72 | 1.61 × 10−10 | up |
| AT4G34760 | AtSAUR50 | 7.59 | 2.92 | 3.21 × 10−10 | up |
| AT2G21200 | AtSAUR7 | 6.61 | 2.72 | 2.32 × 10−7 | up |
| AT2G21210 | AtSAUR6 | 6.21 | 2.63 | 6.31 × 10−20 | up |
| AT5G18020 | AtSAUR20 | 5.34 | 2.42 | 5.94 × 10−7 | up |
| AT5G18060 | AtSAUR23 | 5.13 | 2.36 | 3.46 × 10−5 | up |
| AT5G18030 | AtSAUR21 | 5.07 | 2.34 | 1.17 × 10−4 | up |
| AT4G38860 | AtSAUR16 | 4.18 | 2.06 | 9.38 × 10−17 | up |
| Nitrogen metabolism | |||||
| AT1G77760 | AtNIA1 | 6.89 | 2.78 | 4.14 × 10−7 | up |
| AT1G37130 | AtNIA2 | 2.50 | 1.32 | 1.34 × 10−7 | up |
| AT2G15620 | AtNIR1 | 2.41 | 1.27 | 3.83 × 10−6 | up |
| Alanine, aspartate and glutamate metabolism | |||||
| AT5G65010 | AtASN2 | 7.26 | 2.86 | 1.47 × 10−16 | up |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, S.; Li, W.; Yan, S.; Shi, W. Transcriptomic Analysis of Differentially Expressed Genes in Arabidopsis thaliana Overexpressing BnMYB2 from Boehmeria nivea under Cadmium Stress. Catalysts 2023, 13, 662. https://doi.org/10.3390/catal13040662
Zhu S, Li W, Yan S, Shi W. Transcriptomic Analysis of Differentially Expressed Genes in Arabidopsis thaliana Overexpressing BnMYB2 from Boehmeria nivea under Cadmium Stress. Catalysts. 2023; 13(4):662. https://doi.org/10.3390/catal13040662
Chicago/Turabian StyleZhu, Shoujing, Wenyang Li, Suhui Yan, and Wenjuan Shi. 2023. "Transcriptomic Analysis of Differentially Expressed Genes in Arabidopsis thaliana Overexpressing BnMYB2 from Boehmeria nivea under Cadmium Stress" Catalysts 13, no. 4: 662. https://doi.org/10.3390/catal13040662