
Engineering Strategies for Efficient Bioconversion of Glycerol to Value-Added Products by Yarrowia lipolytica

Abstract
:1. Introduction
2. Glycerol-Based Biomanufacturing Using Yarrowia
2.1. Bioconversion of Glycerol to Lipids
2.1.1. Lipogenesis in Oleaginous Microorganisms
- The allosteric inhibition of the lipid biosynthetic pathway by saturated FA [49];
- The maximum lipid accumulation capacity;
- The level of dissolved oxygen which has to sustain specific growth rate and cell density at a stable productive state but also prevent the downregulation of lipid biosynthesis [44];
2.1.2. Pure and Crude Glycerol as Carbon Source for Lipid Production by Yarrowia lipolytica
2.2. Bioconversion of Glycerol to Various Bio-Products
3. Pathway Metabolic Engineering Strategies to Improve Glycerol-Based Bioprocesses
3.1. Lipids
3.1.1. TAG Biosynthetic Pathway
3.1.2. Precursor Metabolites
3.1.3. Engineering Redox and Energy Equivalents
- The NADP+-dependent GPD (cofactor switch by using NADP+-dependent GPDs i.e., GapC from Clostridium acetobutylicum and GPD1 from Kluyveromyces lactis);
- The pyruvate/oxaloacetate/malate which that converts 1 mol NADH to 1 mol NADPH at a cost of 1 mol ATP (by using the NADP+-dependent malic enzyme MCE2 from Mucor circinelloides);
- The non-oxidative glycolytic pathway which yields 3 mol acetyl-CoA from 1 mol glucose, bypassing the Embden-Meyerhof-Parnas (EMP) pathway (by co-expressing phosphoketolase from Leuconostoc mesenteroides and phosphate acetyltransferase from Clostridium kluyveri).
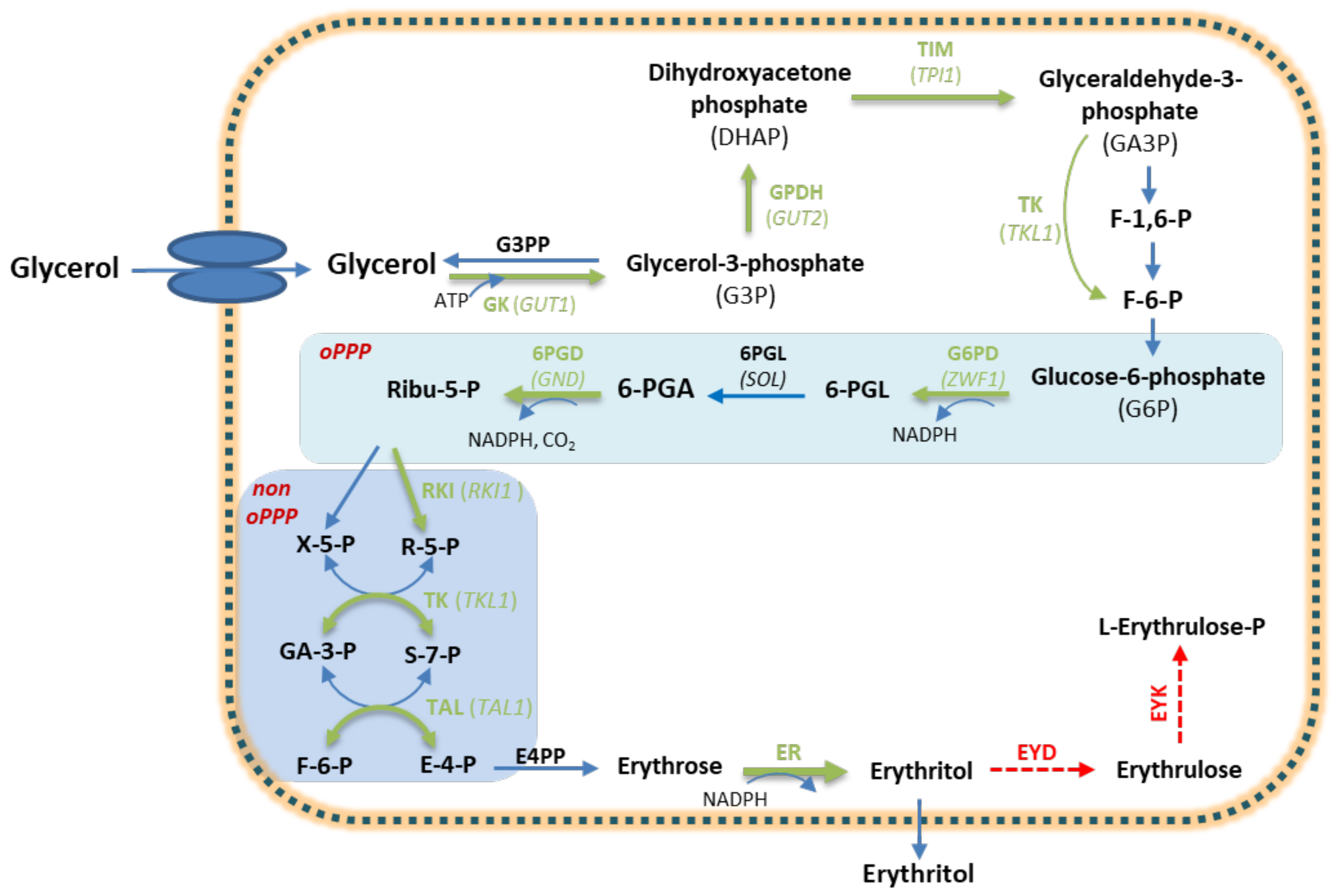
3.2. Polyols
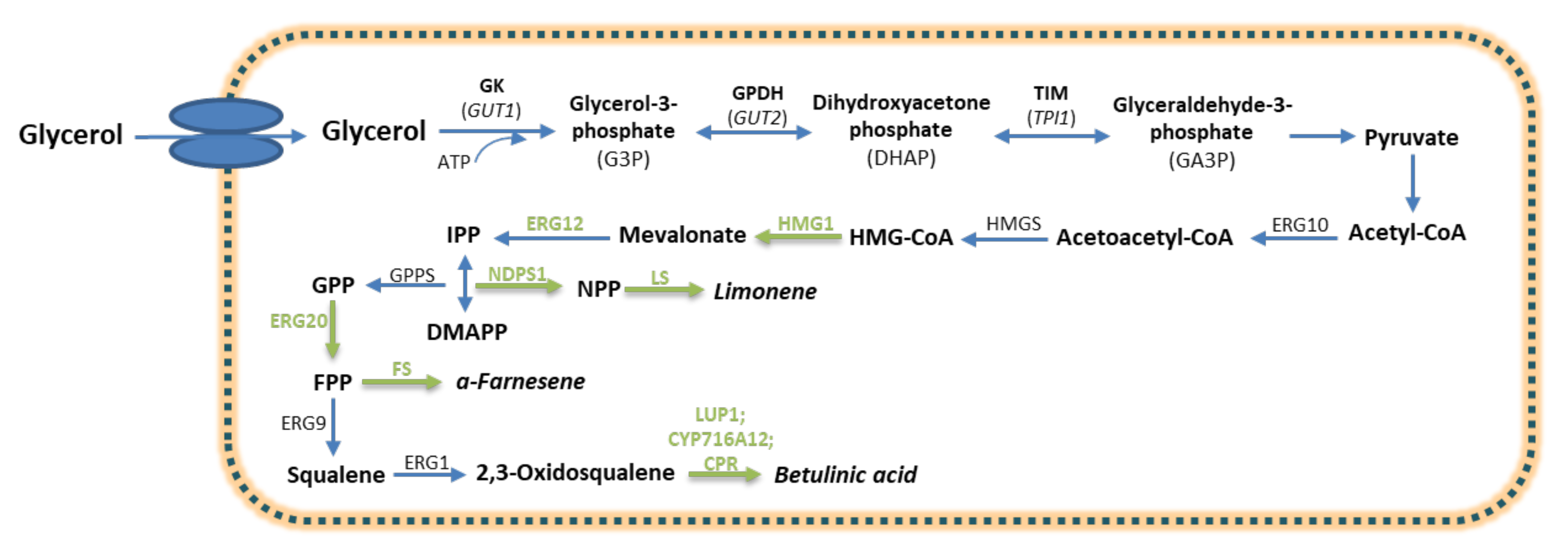
3.3. Terpenoids
3.4. TCA Cycle Intermediates

{kind=link}
{kind=link}
{kind=link}
| Strain | Carbon Source 1 | Product | Engineering 2 | Strategy | Set-Up 3 | Data from [Ref.] |
|---|---|---|---|---|---|---|
| VKM Y-2378 | CG | 41 g L−1 pyruvate; a-ketoglutarate | SE; ME; PE | 33 taxonomically different yeast strains; thiamine and biotin concentration; nitrogen source; temperature, aeration, and glycerol concentration; pH 4.5 | B | [78] |
| Po1f | CG | 160 g L−1 succinic acid | SE; PE | Inactivation of SDH5; effect of pH, oxygen supply; pH 6.0 | B | [124] |
| W29 | CG | 81.11 g L−1 citric acid; 24.90 g L−1 malic acid | ME | Optimization of the carbon and nitrogen source; pH 4.0 | B | [72] |
| Po1f | PG | 110.7 g L−1 succinic acid | SE; PE | Deletion of Ylach; overexpression of heterologous ScPCK from S. cerevisiae and native YlSCS2; fed-batch fermentation; pH not regulated | B | [126] |
| Po1f | PG | 45.5 g L−1 succinic acid | SE; ME | Ts-mutants with blocked SDH activity; medium buffering; CaCO3; pH < 3.5 | SF | [123] |
| WSH-Z06 | PG | 66.2 g L−1 a-ketoglutaric acid | ME; PE | Two-stage pH control strategy; constant feeding; pH 3.0 | B | [128] |
| 374/4 | PG | 8.2 g L−1 pyruvic acid | SS; ME | Thiamine-auxotrophic strain; thiamine concentration; periodical addition of glycerol; pH 4.5 | B | [129] |
3.5. Polyphenols
4. Conclusions
5. Future Perspectives
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bankar, A.V.; Kumar, A.R.; Zinjarde, S.S. Environmental and industrial applications of Yarrowia lipolytica. Appl. Microbiol. Biotechnol. 2009, 84, 847–865. [Google Scholar] [CrossRef] [PubMed]
- Barth, G.; Gaillardin, C. Yarrowia lipolytica. In Nonconventional Yeasts in Biotechnology, 1st ed.; Springer: Berlin/Heidelberg, Germany, 1996; pp. 313–388. [Google Scholar] [CrossRef]
- Barth, G.; Gaillardin, C. Physiology and genetics of the dimorphic fungus Yarrowia lipolytica. FEMS Microbiol. Rev. 1997, 19, 219–237. [Google Scholar] [CrossRef] [PubMed]
- Fickers, P.; Benetti, P.H.; Waché, Y.; Marty, A.; Mauersberger, S.; Smit, M.S.; Nicaud, J.M. Hydrophobic substrate utilisation by the yeast Yarrowia lipolytica, and its potential applications. FEMS Yeast Res. 2005, 5, 527–543. [Google Scholar] [CrossRef] [Green Version]
- Wickerham, L.J.; Kurtzman, C.P.; Herman, A.I. Sexual reproduction in Candida lipolytica. Science 1970, 167, 1141. [Google Scholar] [CrossRef]
- Pérez-Campo, F.M.; Domínguez, A. Factors affecting the morphogenetic switch in Yarrowia lipolytica. Curr. Microbiol. 2001, 43, 429–433. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Mawgoud, A.M.; Markham, K.A.; Palmer, C.M.; Liu, N.; Stephanopoulos, G.; Alper, H.S. Metabolic engineering in the host Yarrowia lipolytica. Metab. Eng. 2018, 50, 192–208. [Google Scholar] [CrossRef]
- Beopoulos, A.; Cescut, J.; Haddouche, R.; Uribelarrea, J.L.; Molina-Jouve, C.; Nicaud, J.M. Yarrowia lipolytica as a model for bio-oil production. Prog. Lipid Res. 2009, 48, 375–387. [Google Scholar] [CrossRef]
- Celińska, E.; Grajek, W. A novel multigene expression construct for modification of glycerol metabolism in Yarrowia lipolytica. Microb. Cell Fact. 2013, 12, 102. [Google Scholar] [CrossRef] [Green Version]
- Hatti-Kaul, R.; Törnvall, U.; Gustafsson, L.; Börjesson, P. Industrial biotechnology for the production of bio-based chemicals—A cradle-to-grave perspective. Trends Biotechnol. 2007, 25, 119–124. [Google Scholar] [CrossRef]
- Levinson, W.E.; Kurtzman, C.P.; Kuo, T.M. Characterization of Yarrowia lipolytica and related species for citric acid production from glycerol. Enzyme Microb. Technol. 2007, 41, 292–295. [Google Scholar] [CrossRef]
- Madzak, C. Engineering Yarrowia lipolytica for use in biotechnological applications: A review of major achievements and recent innovations. Mol. Biotechnol. 2018, 60, 621–635. [Google Scholar] [CrossRef] [PubMed]
- Willke, T.; Vorlop, K.D. Industrial bioconversion of renewable resources as an alternative to conventional chemistry. Appl. Microbiol. Biotechnol. 2004, 66, 131–142. [Google Scholar] [CrossRef]
- Workman, M.; Holt, P.; Thykaer, J. Comparing cellular performance of Yarrowia lipolytica during growth on glucose and glycerol in submerged cultivations. AMB Express 2013, 3, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groenewald, M.; Boekhout, T.; Neuvéglise, C.; Gaillardin, C.; van Dijck, P.W.; Wyss, M. Yarrowia lipolytica: Safety assessment of an oleaginous yeast with a great industrial potential. Crit. Rev. Microbiol. 2014, 40, 187–206. [Google Scholar] [CrossRef]
- Markham, K.A.; Alper, H.S. Synthetic biology expands the industrial potential of Yarrowia lipolytica. Trends Biotechnol. 2018, 36, 1085–1095. [Google Scholar] [CrossRef]
- Yan, J.; Han, B.; Gui, X.; Wang, G.; Xu, L.; Yan, Y.; Madzak, C.; Pan, D.; Wang, Y.; Zha, G.; et al. Engineering Yarrowia lipolytica to simultaneously produce lipase and single cell protein from agro-industrial wastes for feed. Sci. Rep. 2018, 8, 758. [Google Scholar] [CrossRef] [Green Version]
- Casaregola, S.; Neuvéglise, C.; Lépingle, A.; Bon, E.; Feynerol, C.; Artiguenave, F.; Wincker, P.; Gaillardin, C. Genomic exploration of the hemiascomycetous yeasts: 17 Yarrowia lipolytica. FEBS Lett. 2000, 487, 95–100. [Google Scholar] [CrossRef] [Green Version]
- Dujon, B.; Sherman, D.; Fischer, G.; Durrens, P.; Casaregola, S.; Lafontaine, I.; De Montigny, J.; Marck, C.; Neuvéglise, C.; Talla, E.; et al. Genome evolution in yeasts. Nature 2004, 430, 35–44. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Muniglia, L.; Chevalot, I.; Aggelis, G.; Marc, I. Accumulation of a cocoa-butter-like lipid by Yarrowia lipolytica cultivated on agro-industrial residues. Curr. Microbiol. 2003, 46, 124–130. [Google Scholar] [CrossRef]
- Ratledge, C.; Wynn, J.P. The biochemistry and molecular biology of lipid accumulation in oleaginous microorganisms. Adv. Appl. Microbiol. 2002, 51, 1–52. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Goodman, J.M. The lipid droplet—A well-connected organelle. Front. Cell Dev. Biol. 2015, 3, 49. [Google Scholar] [CrossRef] [Green Version]
- Beopoulos, A.; Nicaud, J.M.; Gaillardin, C. An overview of lipid metabolism in yeasts and its impact on biotechnological processes. Appl. Microbiol. Biotechnol. 2011, 90, 1193–1206. [Google Scholar] [CrossRef] [PubMed]
- Kerkhoven, E.J.; Pomraning, K.R.; Baker, S.E.; Nielsen, J. Regulation of amino-acid metabolism controls flux to lipid accumulation in Yarrowia lipolytica. NPJ Syst. Biol. Appl. 2016, 2, 16005. [Google Scholar] [CrossRef] [Green Version]
- Wasylenko, T.M.; Ahn, W.S.; Stephanopoulos, G. The oxidative pentose phosphate pathway is the primary source of NADPH for lipid overproduction from glucose in Yarrowia lipolytica. Metab. Eng. 2015, 30, 27–39. [Google Scholar] [CrossRef] [Green Version]
- Klein, M.; Swinnen, S.; Thevelein, J.M.; Nevoigt, E. Glycerol metabolism and transport in yeast and fungi: Established knowledge and ambiguities. Environ. Microbiol. 2017, 19, 878–893. [Google Scholar] [CrossRef] [Green Version]
- Mirończuk, A.M.; Rzechonek, D.A.; Biegalska, A.; Rakicka, M.; Dobrowolski, A. A novel strain of Yarrowia lipolytica as a platform for value-added product synthesis from glycerol. Biotechnol. Biofuels 2016, 9, 180. [Google Scholar] [CrossRef] [Green Version]
- Deckwer, W.D. Microbial conversion of glycerol to 1,3-propanediol. FEMS Microbiol. Rev. 1995, 16, 143–149. [Google Scholar] [CrossRef]
- Dharmadi, Y.; Murarka, A.; Gonzalez, R. Anaerobic fermentation of glycerol by Escherichia coli: A new platform for metabolic engineering. Biotechnol. Bioeng. 2006, 94, 821–829. [Google Scholar] [CrossRef] [PubMed]
- Vivek, N.; Sindhu, R.; Madhavan, A.; Anju, A.J.; Castro, E.; Faraco, V.; Pandey, A.; Binod, P. Recent advances in the production of value added chemicals and lipids utilizing biodiesel industry generated crude glycerol as a substrate—Metabolic aspects, challenges and possibilities: An overview. Bioresour. Technol. 2017, 239, 507–517. [Google Scholar] [CrossRef] [PubMed]
- Demirbas, A.; Bafail, A.; Ahmad, W.; Sheikh, M. Biodiesel production from non-edible plant oils. Energy Explor. Exploit. 2016, 34, 290–318. [Google Scholar] [CrossRef] [Green Version]
- Meher, L.C.; Vidya Sagar, D.; Naik, S.N. Technical aspects of biodiesel production by transesterification—A review. Renew. Sustain. Energy Rev. 2006, 10, 248–268. [Google Scholar] [CrossRef]
- Yang, F.; Hanna, M.A.; Sun, R. Value-added uses for crude glycerol—A byproduct of biodiesel production. Biotechnol. Biofuels 2012, 5, 13. [Google Scholar] [CrossRef] [Green Version]
- Rigouin, C.; Croux, C.; Borsenberger, V.; Ben Khaled, M.; Chardot, T.; Marty, A.; Bordes, F. Increasing medium chain fatty acids production in Yarrowia lipolytica by metabolic engineering. Microb. Cell Fact. 2018, 17, 142. [Google Scholar] [CrossRef]
- Ledesma-Amaro, R.; Nicaud, J.M. Metabolic engineering for expanding the substrate range of Yarrowia lipolytica. Trends Biotechnol. 2016, 34, 798–809. [Google Scholar] [CrossRef] [PubMed]
- Dobrowolski, A.; Drzymała, K.; Mituła, P.; Mirończuk, A.M. Production of tailor-made fatty acids from crude glycerol at low pH by Yarrowia lipolytica. Bioresour. Technol. 2020, 314, 123746. [Google Scholar] [CrossRef] [PubMed]
- Zweytick, D.; Athenstaedt, K.; Daum, G. Intracellular lipid particles of eukaryotic cells. Biochim. Biophys. Acta 2000, 1469, 101–120. [Google Scholar] [CrossRef] [PubMed]
- Czabany, T.; Athenstaedt, K.; Daum, G. Synthesis, storage and degradation of neutral lipids in yeast. Biochim. Biophys. Acta 2007, 1771, 299–309. [Google Scholar] [CrossRef]
- Walther, T.C.; Chung, J.; Farese, R.V. Lipid droplet biogenesis. Annu. Rev. Cell Dev. Biol. 2017, 33, 491–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, C.; Gu, D.; Nambou, K.; Wei, L.; Chen, J.; Imanaka, T.; Hua, Q. Metabolome analysis and pathway abundance profiling of Yarrowia lipolytica cultivated on different carbon sources. J. Biotechnol. 2015, 206, 42–51. [Google Scholar] [CrossRef]
- Xu, P.; Qiao, K.; Ahn, W.S.; Stephanopoulos, G. Engineering Yarrowia lipolytica as a platform for synthesis of drop-in transportation fuels and oleochemicals. Proc. Natl. Acad. Sci. USA 2016, 113, 10848–10853. [Google Scholar] [CrossRef] [Green Version]
- Das, S.; Zarur Coelho, M.A.; Amaral, P.F.F.; Sil, J. Development of nutrient media to increase the accumulation of lipids without genetic modification of a lipogenic microorganism. RSC Adv. 2017, 7, 38149–38154. [Google Scholar] [CrossRef] [Green Version]
- Blazeck, J.; Hill, A.; Liu, L.; Knight, R.; Miller, J.; Pan, A.; Otoupal, P.; Alper, H.S. Harnessing Yarrowia lipolytica lipogenesis to create a platform for lipid and biofuel production. Nat. Commun. 2014, 5, 3131. [Google Scholar] [CrossRef] [Green Version]
- Dourou, M.; Aggeli, D.; Papanikolaou, S.; Aggelis, G. Critical steps in carbon metabolism affecting lipid accumulation and their regulation in oleaginous microorganisms. Appl. Microbiol. Biotechnol. 2018, 102, 2509–2523. [Google Scholar] [CrossRef]
- Gao, Q.; Cao, X.; Huang, Y.Y.; Yang, J.L.; Chen, J.; Wei, L.J.; Hua, Q. Overproduction of fatty acid ethyl esters by the oleaginous yeast Yarrowia lipolytica through metabolic engineering and process optimization. ACS Synth. Biol. 2018, 7, 1371–1380. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhang, L.; Chen, H.; Chen, Y.Q.; Ratledge, C.; Song, Y.; Chen, W. Regulatory properties of malic enzyme in the oleaginous yeast, Yarrowia lipolytica, and its non-involvement in lipid accumulation. Biotechnol. Lett. 2013, 35, 2091–2098. [Google Scholar] [CrossRef]
- Silverman, A.M.; Qiao, K.; Xu, P.; Stephanopoulos, G. Functional overexpression and characterization of lipogenesis-related genes in the oleaginous yeast Yarrowia lipolytica. Appl. Microbiol. Biotechnol. 2016, 100, 3781–3798. [Google Scholar] [CrossRef] [Green Version]
- Park, J.O.; Liu, N.; Holinski, K.M.; Emerson, D.F.; Qiao, K.; Woolston, B.M.; Xu, J.; Lazar, Z.; Islam, M.A.; Vidoudez, C.; et al. Synergistic substrate cofeeding stimulates reductive metabolism. Nat. Metab. 2019, 1, 643–651. [Google Scholar] [CrossRef]
- Qiao, K.; Abidi, S.H.I.; Liu, H.; Zhang, H.; Chakraborty, S.; Watson, N.; Ajikumar, P.K.; Stephanopoulos, G. Engineering lipid overproduction in the oleaginous yeast Yarrowia lipolytica. Metab. Eng. 2015, 29, 56–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magdouli, S.; Guedri, T.; Tarek, R.; Brar, S.K.; Blais, J.F. Valorization of raw glycerol and crustacean waste into value added products by Yarrowia lipolytica. Bioresour. Technol. 2017, 243, 57–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katre, G.; Joshi, C.; Khot, M.; Zinjarde, S.; Ravikumar, A. Evaluation of single cell oil (SCO) from a tropical marine yeast Yarrowia lipolytica NCIM 3589 as a potential feedstock for biodiesel. AMB Express 2012, 2, 36. [Google Scholar] [CrossRef] [Green Version]
- Gajdoš, P.; Nicaud, J.M.; Čertík, M. Glycerol conversion into a single cell oil by engineered Yarrowia lipolytica. Eng. Life Sci. 2016, 17, 325–332. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Aggelis, G. Lipid production by Yarrowia lipolytica growing on industrial glycerol in a single-stage continuous culture. Bioresour. Technol. 2002, 82, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Dobrowolski, A.; Mituła, P.; Rymowicz, W.; Mirończuk, A.M. Efficient conversion of crude glycerol from various industrial wastes into single cell oil by yeast Yarrowia lipolytica. Bioresour. Technol. 2016, 207, 237–243. [Google Scholar] [CrossRef]
- Kuttiraja, M.; Dhouha, A.; Tyagi, R.D. Harnessing the Effect of pH on lipid production in batch cultures of Yarrowia lipolytica SKY7. Appl. Biochem. Biotechnol. 2018, 184, 1332–1346. [Google Scholar] [CrossRef]
- Ram, S.K.; Tyagi, R.D.; Drogui, P. Effect of sludge concentration and crude glycerol matrix as a substrate on the production of single-cell oil by oleaginous yeast Yarrowia lipolytica SKY7. Fermentation 2018, 4, 24. [Google Scholar] [CrossRef] [Green Version]
- Canonico, L.; Ashoor, S.; Taccari, M.; Comitini, F.; Antonucci, M.; Truzzi, C.; Scarponi, G.; Ciani, M. Conversion of raw glycerol to microbial lipids by new Metschnikowia and Yarrowia lipolytica strains. Ann. Microbiol. 2016, 66, 1409–1418. [Google Scholar] [CrossRef]
- Rakicka, M.; Lazar, Z.; Dulermo, T.; Fickers, P.; Nicaud, J.M. Lipid production by the oleaginous yeast Yarrowia lipolytica using industrial by-products under different culture conditions. Biotechnol. Biofuels 2015, 8, 104. [Google Scholar] [CrossRef] [Green Version]
- Dobrowolski, A.; Drzymała, K.; Rzechonek, D.A.; Mituła, P.; Mirończuk, A.M. Lipid production from waste materials in seawater-based medium by the yeast Yarrowia lipolytica. Front. Microbiol. 2019, 10, 547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Zhang, L.; Chen, H.; Chen, Y.Q.; Chen, W.; Song, Y.; Ratledge, C. Enhanced lipid accumulation in the yeast Yarrowia lipolytica by over-expression of ATP: Citrate lyase from Mus musculus. J. Biotechnol. 2014, 192, 78–84. [Google Scholar] [CrossRef]
- Abghari, A.; Chen, S. Engineering Yarrowia lipolytica for enhanced production of lipid and citric Acid. Fermentation 2017, 3, 34. [Google Scholar] [CrossRef] [Green Version]
- Fontanille, P.; Kumar, V.; Christophe, G.; Nouaille, R.; Larroche, C. Bioconversion of volatile fatty acids into lipids by the oleaginous yeast Yarrowia lipolytica. Bioresour. Technol. 2012, 114, 443–449. [Google Scholar] [CrossRef]
- Kuttiraja, M.; Douha, A.; Valéro, J.R.; Tyagi, R.D. Elucidating the effect of glycerol concentration and C/N ratio on lipid production using Yarrowia lipolytica SKY7. Appl. Biochem. Biotechnol. 2016, 180, 1586–1600. [Google Scholar] [CrossRef] [PubMed]
- Samul, D.; Leja, K.; Grajek, W. Impurities of crude glycerol and their effect on metabolite production. Ann. Microbiol. 2014, 64, 891–898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papanikolaou, S.; Muniglia, L.; Chevalot, I.; Aggelis, G.; Marc, I. Yarrowia lipolytica as a potential producer of citric acid from raw glycerol. J. Appl. Microbiol. 2002, 92, 737–744. [Google Scholar] [CrossRef] [PubMed]
- Papanikolaou, S.; Aggelis, G. Biotechnological valorization of biodiesel derived glycerol waste through production of single cell oil and citric acid by Yarrowia lipolytica. Lipid Technol. 2009, 21, 83–87. [Google Scholar] [CrossRef]
- Rzechonek, D.A.; Dobrowolski, A.; Rymowicz, W.; Mirończuk, A.M. Aseptic production of citric and isocitric acid from crude glycerol by genetically modified Yarrowia lipolytica. Bioresour. Technol. 2019, 271, 340–344. [Google Scholar] [CrossRef]
- Tzirita, M.; Kremmyda, M.; Sarris, D.; Koutinas, A.A.; Papanikolaou, S. Effect of salt addition upon the production of metabolic compounds by Yarrowia lipolytica cultivated on biodiesel-derived glycerol diluted with olive-mill wastewaters. Energies 2019, 12, 3649. [Google Scholar] [CrossRef] [Green Version]
- Morgunov, I.G.; Kamzolova, S.V.; Lunina, J.N. The citric acid production from raw glycerol by Yarrowia lipolytica yeast and its regulation. Appl. Microbiol. Biotechnol. 2013, 97, 7387–7397. [Google Scholar] [CrossRef]
- Rywińska, A.; Rymowicz, W.; Marcinkiewicz, M. Valorization of raw glycerol for citric acid production by Yarrowia lipolytica yeast. Electron. J. Biotechnol. 2010, 13, 9–10. [Google Scholar] [CrossRef] [Green Version]
- Rymowicz, W.; Fatykhova, A.R.; Kamzolova, S.V.; Rywińska, A.; Morgunov, I.G. Citric acid production from glycerol-containing waste of biodiesel industry by Yarrowia lipolytica in batch, repeated batch, and cell recycle regimes. Appl. Microbiol. Biotechnol. 2010, 87, 971–979. [Google Scholar] [CrossRef]
- Qian, X.; Xu, N.; Jing, Y.; Song, M.; Zhou, J.; Dong, W.; Xin, F.; Zhang, W.; Jiang, M.; Ochsenreither, K. Valorization of crude glycerol into citric acid and malic acid by Yarrowia lipolytica. Ind. Eng. Chem. Res. 2020, 59, 17165–17172. [Google Scholar] [CrossRef]
- Lazar, Z.; Walczak, E.; Robak, M. Simultaneous production of citric acid and invertase by Yarrowia lipolytica SUC+ transformants. Bioresour. Technol. 2011, 102, 6982–6989. [Google Scholar] [CrossRef]
- Tomaszewska, L.; Rakicka, M.; Rymowicz, W.; Rywińska, A. A comparative study on glycerol metabolism to erythritol and citric acid in Yarrowia lipolytica yeast cells. FEMS Yeast Res. 2014, 14, 966–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rakicka-Pustułka, M.; Miedzianka, J.; Jama, D.; Kawalec, S.; Liman, K.; Janek, T.; Skaradziński, G.; Rymowicz, W.; Lazar, Z. High value-added products derived from crude glycerol via microbial fermentation using Yarrowia clade yeast. Microb. Cell Fact. 2021, 20, 195. [Google Scholar] [CrossRef]
- Rywińska, A.; Marcinkiewicz, M.; Cibis, E.; Rymowicz, W. Optimization of medium composition for erythritol production from glycerol by Yarrowia lipolytica using response surface methodology. Prep. Biochem. Biotechnol. 2015, 45, 515–529. [Google Scholar] [CrossRef]
- Juszczyk, P.; Tomaszewska, L.; Kita, A.; Rymowicz, W. Biomass production by novel strains of Yarrowia lipolytica using raw glycerol, derived from biodiesel production. Bioresour. Technol. 2013, 137, 124–131. [Google Scholar] [CrossRef]
- Kamzolova, S.V.; Morgunov, I.G. Biosynthesis of pyruvic acid from glycerol-containing substrates and its regulation in the yeast Yarrowia lipolytica. Bioresour. Technol. 2018, 266, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.C.; Liu, Z.; Wei, L.J.; Hua, Q. Pathway engineering and medium optimization for α-farnesene biosynthesis in oleaginous yeast Yarrowia lipolytica. J. Biotechnol. 2020, 319, 74–81. [Google Scholar] [CrossRef]
- Ma, Y.R.; Wang, K.F.; Wang, W.J.; Ding, Y.; Shi, T.Q.; Huang, H.; Ji, X.J. Advances in the metabolic engineering of Yarrowia lipolytica for the production of terpenoids. Bioresour. Technol. 2019, 281, 449–456. [Google Scholar] [CrossRef] [PubMed]
- Arnesen, J.A.; Kildegaard, K.R.; Cernuda Pastor, M.; Jayachandran, S.; Kristensen, M.; Borodina, I. Yarrowia lipolytica strains engineered for the production of terpenoids. Front. Bioeng. Biotechnol. 2020, 8, 945. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Wang, H.; Zhang, Z.; Verstrepen, K.J.; Wang, Q.; Dai, Z. Metabolic engineering of Yarrowia lipolytica for terpenoids production: Advances and perspectives. Crit. Rev. Biotechnol. 2022, 42, 618–633. [Google Scholar] [CrossRef] [PubMed]
- Xue, Z.; Sharpe, P.L.; Hong, S.-P.; Yadav, N.S.; Xie, D.; Short, D.R.; Damude, H.G.; A Rupert, R.; E Seip, J.; Wang, J.; et al. Production of omega-3 eicosapentaenoic acid by metabolic engineering of Yarrowia lipolytica. Nat. Biotechnol. 2013, 31, 734–740. [Google Scholar] [CrossRef] [PubMed]
- Sibirny, A.; Madzak, C.; Fickers, P. Genetic engineering of non-conventional yeast for the production of valuable compounds. In Microbial Biotechnology: Progress and Trends, 1st ed.; CRC Press: Boca Raton, FL, USA, 2014. [Google Scholar] [CrossRef]
- Qiao, K.; Wasylenko, T.M.; Zhou, K.; Xu, P.; Stephanopoulos, G. Lipid production in Yarrowia lipolytica is maximized by engineering cytosolic redox metabolism. Nat. Biotechnol. 2017, 35, 173–177. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ledesma-Amaro, R.; Wei, Y.; Ji, B.; Ji, X.J. Metabolic engineering for increased lipid accumulation in Yarrowia lipolytica—A Review. Bioresour. Technol. 2020, 313, 123707. [Google Scholar] [CrossRef]
- Beopoulos, A.; Mrozova, Z.; Thevenieau, F.; Le Dall, M.-T.; Hapala, I.; Papanikolaou, S.; Chardot, T.; Nicaud, J.-M. Control of lipid accumulation in the yeast Yarrowia lipolytica. Appl. Environ. Microbiol. 2008, 74, 7779–7789. [Google Scholar] [CrossRef] [Green Version]
- Dulermo, T.; Nicaud, J.M. Involvement of the G3P shuttle and β-oxidation pathway in the control of TAG synthesis and lipid accumulation in Yarrowia lipolytica. Metab. Eng. 2011, 13, 482–491. [Google Scholar] [CrossRef]
- Yuzbasheva, E.Y.; Mostova, E.B.; Andreeva, N.I.; Yuzbashev, T.V.; Fedorov, A.S.; Konova, I.A.; Sineoky, S.P. A metabolic engineering strategy for producing free fatty acids by the Yarrowia lipolytica yeast based on impairment of glycerol metabolism. Biotechnol. Bioeng. 2018, 115, 433–443. [Google Scholar] [CrossRef]
- Xue, L.L.; Chen, H.H.; Jiang, J.G. Implications of glycerol metabolism for lipid production. Prog. Lipid Res. 2017, 68, 12–25. [Google Scholar] [CrossRef]
- Zhu, Q.; Jackson, E.N. Metabolic engineering of Yarrowia lipolytica for industrial applications. Curr. Opin. Biotechnol. 2015, 36, 65–72. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.Y.; Jian, X.X.; Lv, Y.B.; Nian, K.Q.; Gao, Q.; Chen, J.; Wei, L.J.; Hua, Q. Enhanced squalene biosynthesis in Yarrowia lipolytica based on metabolically engineered acetyl-CoA metabolism. J. Biotechnol. 2018, 281, 106–114. [Google Scholar] [CrossRef]
- Fakas, S. Lipid biosynthesis in yeasts: A comparison of the lipid biosynthetic pathway between the model nonoleaginous yeast Saccharomyces cerevisiae and the model oleaginous yeast Yarrowia lipolytica. Eng. Life Sci. 2017, 17, 292–302. [Google Scholar] [CrossRef] [PubMed]
- Dulermo, T.; Lazar, Z.; Dulermo, R.; Rakicka, M.; Haddouche, R.; Nicaud, J.M. Analysis of ATP-citrate lyase and malic enzyme mutants of Yarrowia lipolytica points out the importance of mannitol metabolism in fatty acid synthesis. Biochim. Biophys. Acta 2015, 1851, 1107–1117. [Google Scholar] [CrossRef] [Green Version]
- Tang, X.; Chen, H.; Chen, Y.Q.; Chen, W.; Garre, V.; Song, Y.; Ratledge, C. Comparison of biochemical activities between high and low lipid-producing strains of Mucor circinelloides: An Explanation for the High Oleaginicity of Strain WJ11. PLoS ONE 2015, 10, e0128396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erian, A.M.; Sauer, M. Utilizing yeasts for the conversion of renewable feedstocks to sugar alcohols—A review. Bioresour. Technol. 2022, 346, 126296. [Google Scholar] [CrossRef]
- Rakicka, M.; Biegalska, A.; Rymowicz, W.; Dobrowolski, A.; Mirończuk, A.M. Polyol production from waste materials by genetically modified Yarrowia lipolytica. Bioresour. Technol. 2017, 243, 393–399. [Google Scholar] [CrossRef]
- Rymowicz, W.; Rywińska, A.; Gładkowski, W. Simultaneous production of citric acid and erythritol from crude glycerol by Yarrowia lipolytica Wratislavia K1. Chem. Pap. 2008, 62, 239–246. [Google Scholar] [CrossRef]
- Rymowicz, W.; Rywińska, A.; Marcinkiewicz, M. High-yield production of erythritol from raw glycerol in fed-batch cultures of Yarrowia lipolytica. Biotechnol. Lett. 2009, 31, 377–380. [Google Scholar] [CrossRef]
- Tomaszewska, L.; Rywińska, A.; Gladkowski, W. Production of erythritol and mannitol by Yarrowia lipolytica yeast in media containing glycerol. J. Ind. Microbiol. Biotechnol. 2012, 39, 1333–1343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatzifragkou, A.; Makri, A.; Belka, A.; Bellou, S.; Mavrou, M.; Mastoridou, M.; Mystrioti, P.; Onjaro, G.; Aggelis, G.; Papanikolaou, S. Biotechnological conversions of biodiesel derived waste glycerol by yeast and fungal species. Energy 2011, 36, 1097–1108. [Google Scholar] [CrossRef]
- Yang, S.; Pan, X.; Wang, Q.; Lv, Q.; Zhang, X.; Zhang, R.; Rao, Z. Enhancing erythritol production from crude glycerol in a wild-type Yarrowia lipolytica by metabolic engineering. Front. Microbiol. 2022, 13, 1054243. [Google Scholar] [CrossRef]
- Mirończuk, A.M.; Biegalska, A.; Dobrowolski, A. Functional overexpression of genes involved in erythritol synthesis in the yeast Yarrowia lipolytica. Biotechnol. Biofuels 2017, 10, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carly, F.; Vandermies, M.; Telek, S.; Steels, S.; Thomas, S.; Nicaud, J.M.; Fickers, P. Enhancing erythritol productivity in Yarrowia lipolytica using metabolic engineering. Metab. Eng. 2017, 42, 19–24. [Google Scholar] [CrossRef]
- Tomaszewska, L.; Rywińska, A.; Rymowicz, W. High selectivity of erythritol production from glycerol by Yarrowia lipolytica. Biomass Bioenergy 2014, 64, 309–320. [Google Scholar] [CrossRef]
- Janek, T.; Dobrowolski, A.; Biegalska, A.; Mirończuk, A.M. Characterization of erythrose reductase from Yarrowia lipolytica and its influence on erythritol synthesis. Microb. Cell Fact. 2017, 16, 118. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Nie, M.Y.; Liu, F.; Chen, J.; Wei, L.J.; Hua, Q. Multiple gene integration to promote erythritol production on glycerol in Yarrowia lipolytica. Biotechnol. Lett. 2021, 43, 1277–1287. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.B.; Dai, X.M.; Zheng, Z.Y.; Zhu, L.; Zhan, X.B.; Lin, C.C. Proteomic analysis of erythritol-producing Yarrowia lipolytica from glycerol in response to osmotic pressure. J. Microbiol. Biotechnol. 2015, 25, 1056–1069. [Google Scholar] [CrossRef]
- Jagtap, S.S.; Bedekar, A.A.; Singh, V.; Jin, Y.S.; Rao, C.V. Metabolic engineering of the oleaginous yeast Yarrowia lipolytica PO1f for production of erythritol from glycerol. Biotechnol. Biofuels 2021, 14, 188. [Google Scholar] [CrossRef] [PubMed]
- Meadows, A.L.; Hawkins, K.M.; Tsegaye, Y.; Antipov, E.; Kim, Y.; Raetz, L.; Dahl, R.H.; Tai, A.; Mahatdejkul-Meadows, T.; Xu, L.; et al. Rewriting yeast central carbon metabolism for industrial isoprenoid production. Nature 2016, 537, 694–697. [Google Scholar] [CrossRef]
- Zhu, F.; Zhong, X.; Hu, M.; Lu, L.; Deng, Z.; Liu, T. In vitro reconstitution of mevalonate pathway and targeted engineering of farnesene overproduction in Escherichia coli. Biotechnol. Bioeng. 2014, 111, 1396–1405. [Google Scholar] [CrossRef]
- Yang, X.; Nambou, K.; Wei, L.; Hua, Q. Heterologous production of α-farnesene in metabolically engineered strains of Yarrowia lipolytica. Bioresour. Technol. 2016, 216, 1040–1048. [Google Scholar] [CrossRef]
- Liu, Y.; Jiang, X.; Cui, Z.; Wang, Z.; Qi, Q.; Hou, J. Engineering the oleaginous yeast Yarrowia lipolytica for production of α-farnesene. Biotechnol. Biofuels 2019, 12, 296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunlop, M.J.; Dossani, Z.Y.; Szmidt, H.L.; Chu, H.C.; Lee, T.S.; Keasling, J.D.; Hadi, M.Z.; Mukhopadhyay, A. Engineering microbial biofuel tolerance and export using efflux pumps. Mol. Syst. Biol. 2011, 7, 470. [Google Scholar] [CrossRef] [PubMed]
- Willrodt, C.; David, C.; Cornelissen, S.; Bühler, B.; Julsing, M.K.; Schmid, A. Engineering the productivity of recombinant Escherichia coli for limonene formation from glycerol in minimal media. Biotechnol. J. 2014, 9, 1000–1012. [Google Scholar] [CrossRef]
- Cheng, S.; Liu, X.; Jiang, G.; Wu, J.; Zhang, J.L.; Lei, D.; Yuan, Y.J.; Qiao, J.; Zhao, G.R. Orthogonal engineering of biosynthetic pathway for efficient production of limonene in Saccharomyces cerevisiae. ACS Synth. Biol. 2019, 8, 968–975. [Google Scholar] [CrossRef]
- Brennan, T.C.; Williams, T.C.; Schulz, B.L.; Palfreyman, R.W.; Krömer, J.O.; Nielsen, L.K. Evolutionary engineering improves tolerance for replacement jet fuels in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2015, 81, 3316–3325. [Google Scholar] [CrossRef] [Green Version]
- Cao, X.; Lv, Y.B.; Chen, J.; Imanaka, T.; Wei, L.J.; Hua, Q. Metabolic engineering of oleaginous yeast Yarrowia lipolytica for limonene overproduction. Biotechnol. Biofuels 2016, 9, 214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, B.Q.; Wei, L.J.; Lv, Y.B.; Chen, J.; Hua, Q. Elevating limonene production in oleaginous yeast yarrowia lipolytica via genetic engineering of limonene biosynthesis pathway and optimization of medium composition. Biotechnol. Bioprocess Eng. 2019, 24, 500–506. [Google Scholar] [CrossRef]
- Sun, J.; Zhang, C.; Nan, W.; Li, D.; Ke, D.; Lu, W. Glycerol improves heterologous biosynthesis of betulinic acid in engineered Yarrowia lipolytica. Chem. Eng. Sci. 2019, 196, 82–90. [Google Scholar] [CrossRef]
- Theodosiou, E.; Tüllinghoff, A.; Toepel, J.; Bühler, B. Exploitation of hetero- and phototrophic metabolic modules for redox-intensive whole-cell biocatalysis. Front. Bioeng. Biotechnol. 2022, 10, 462. [Google Scholar] [CrossRef] [PubMed]
- Jost, B.; Holz, M.; Aurich, A.; Barth, G.; Bley, T.; Müller, R.A. The influence of oxygen limitation for the production of succinic acid with recombinant strains of Yarrowia lipolytica. Appl. Microbiol. Biotechnol. 2015, 99, 1675–1686. [Google Scholar] [CrossRef]
- Yuzbashev, T.V.; Yuzbasheva, E.Y.; Sobolevskaya, T.I.; Laptev, I.A.; Vybornaya, T.V.; Larina, A.S.; Matsui, K.; Fukui, K.; Sineoky, S.P. Production of succinic acid at low pH by a recombinant strain of the aerobic yeast Yarrowia lipolytica. Biotechnol. Bioeng. 2010, 107, 673–682. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Yang, X.; Wang, H.; Rivero, C.P.; Li, C.; Cui, Z.; Qi, Q.; Lin, C.S.K. Robust succinic acid production from crude glycerol using engineered Yarrowia lipolytica. Biotechnol. Biofuels 2016, 9, 179. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Gao, S.; Yang, X.; Lin, C.S.K. Green and sustainable succinic acid production from crude glycerol by engineered Yarrowia lipolytica via agricultural residue based in situ fibrous bed bioreactor. Bioresour. Technol. 2018, 249, 612–619. [Google Scholar] [CrossRef]
- Cui, Z.; Gao, C.; Li, J.; Hou, J.; Lin, C.S.K.; Qi, Q. Engineering of unconventional yeast Yarrowia lipolytica for efficient succinic acid production from glycerol at low pH. Metab. Eng. 2017, 42, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Billerach, G.; Preziosi-Belloy, L.; Lin, C.S.K.; Fulcrand, H.; Dubreucq, E.; Grousseau, E. Impact of nitrogen deficiency on succinic acid production by engineered strains of Yarrowia lipolytica. J. Biotechnol. 2021, 336, 30–40. [Google Scholar] [CrossRef]
- Yu, Z.; Du, G.; Zhou, J.; Chen, J. Enhanced α-ketoglutaric acid production in Yarrowia lipolytica WSH-Z06 by an improved integrated fed-batch strategy. Bioresour. Technol. 2012, 114, 597–602. [Google Scholar] [CrossRef]
- Morgunov, I.G.; Kamzolova, S.V.; Perevoznikova, O.A.; Shishkanova, N.V.; Finogenova, T.V. Pyruvic acid production by a thiamine auxotroph of Yarrowia lipolytica. Process Biochem. 2004, 39, 1469–1474. [Google Scholar] [CrossRef]
- Tomaszewska-Hetman, L.; Rywińska, A.; Lazar, Z.; Juszczyk, P.; Rakicka-Pustułka, M.; Janek, T.; Kuźmińska-Bajor, M.; Rymowicz, W. Application of a new engineered strain of Yarrowia lipolytica for effective production of calcium ketoglutarate dietary supplements. Int. J. Mol. Sci. 2021, 22, 7577. [Google Scholar] [CrossRef]
- He, Q.; Szczepańska, P.; Yuzbashev, T.; Lazar, Z.; Ledesma-Amaro, R. De novo production of resveratrol from glycerol by engineering different metabolic pathways in Yarrowia lipolytica. Metab. Eng. Commun. 2020, 11, e00146. [Google Scholar] [CrossRef]
- Sáez-Sáez, J.; Wang, G.; Marella, E.R.; Sudarsan, S.; Cernuda Pastor, M.; Borodina, I. Engineering the oleaginous yeast Yarrowia lipolytica for high-level resveratrol production. Metab. Eng. 2020, 62, 51–61. [Google Scholar] [CrossRef]
- Choi, K.R.; Jang, W.D.; Yang, D.; Cho, J.S.; Park, D.; Lee, S.Y. Systems metabolic engineering strategies: Integrating systems and synthetic biology with metabolic engineering. Trends Biotechnol. 2019, 37, 817–837. [Google Scholar] [CrossRef] [PubMed]
- Gu, C.; Kim, G.B.; Kim, W.J.; Kim, H.U.; Lee, S.Y. Current status and applications of genome-scale metabolic models. Genome Biol. 2019, 20, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwechheimer, S.K.; Becker, J.; Wittmann, C. Towards better understanding of industrial cell factories: Novel approaches for 13C metabolic flux analysis in complex nutrient environments. Curr. Opin. Biotechnol. 2018, 54, 128–137. [Google Scholar] [CrossRef] [PubMed]



| Strain | Carbon Source 1 | Product | Engineering 2 | Strategy | Set-Up 3 | Data from [Ref.] |
|---|---|---|---|---|---|---|
| ACA-DC 50109 | CG | 41% w/w lipids | ME | Mixtures of saturated free fatty acids, CG, and glucose | SF | [20] |
| A101 | CG | 40% w/w lipids | SE; PE | Co-overexpression of GUT1 and SCT1; pH 3.0 | SF | [36] |
| A101 | CG | 12.5% w/w lipids | SE; PE | Co-overexpression of GUT1 and SCT2; pH 3.0 | B | [36] |
| SM7 | CG | 35.80% w/w lipids; lipase activity 38 U/mL | ME | Addition of surfactants and oils in the medium; pH 6.5 | SF | [50] |
| LGAM 5(7)1 | CG | 43% w/w lipids | PE | Highly aerated and agitated continuous nitrogen-limited cultures with glycerol concentration in the feed at different dilution rates; pH 6.0 | B | [53] |
| A101 | CG | 25% w/w lipids | ME | Waste products containing various glycerol concentrations of (42–87%); pH 6.0 | SF | [54] |
| SKY7 | CG | 45.5% w/w lipids | ME | Glycerol-supplemented media with pH 6.5 and without pH control | B | [55] |
| SKY7 | CG | 29.35% w/w lipids | ME | Sludge media with different sludge solid concentrations; pH 6.8 | SF | [56] |
| DiSVA 347 | CG | 25.7% w/w lipids | SS; ME | Glycerol concentration; C/N ratio and time process; pH 6.0 | SF | [57] |
| JMY4086 | CG | 40% w/w lipids | SE; PE | Deletion of POX1–6 and TGL4 gene, overexpression of YlDGA2, YlGPD1, SUC2, HXK1; oxygenation; inoculum densities; pH 3.5 | B | [58] |
| A101 | CG | 21% w/w lipids | SE; ME | DGA1 overexpression; seawater; pH 3.0 | B | [59] |
| NCYC3825 | PG | 38% w/w lipids | SE | Genetic construct with 3 heterologous genes (glycerol dehydratase and its reactivator, wide-spectrum alcohol oxidoreductase) under the control of glycerol-induced promoter; pH 4.0 | B | [9] |
| Polh | PG | 23% w/w lipids | SE | Overexpression of ATP:citrate lyase from Mus musculus with the multi-copy integration vector pINA1292sp; pH 6.0 | SF | [60] |
| H222 | PG | 53% w/w lipids | SE | One-step double gene knock-in and site-specific gene knock-out; DGA1 and DGA2 in a POX-deleted background; deletion of the SNF1 lipid regulator; pH ≥ 2.5 | B | [61] |
| JMY3580 | PG | 53.7% w/w lipids | SE | DGA2 gene in Q4 strain (Δdga1Δdga2Δlro1Δare1); pH not regulated | SF | [52] |
| MUCL 28849 | PG | 38.1% w/w lipids | ME; PE | Addition of volatile fatty acids; two-stage fed-batch strategy; pH 5.6 | B | [62] |
| Strain | Carbon Source 1 | Product | Engineering 2 | Strategy | Set-Up 3 | Data from [Ref.] |
|---|---|---|---|---|---|---|
| LGAM S(7)1 | CG | 35 g L−1 citric acid | ME | High C/N media; pH ≥ 5.0 | SF | [65] |
| A101 | CG | 75.9 g L−1 citric/isocitric acids; 42.5 g L−1 isocitric acid | SE; ME | Co-overexpression of GUT1 and GUT2; pH 3.0 | B | [67] |
| ACA-YC 5031 | CG | 54.0 g L−1 citric acid; secondary metabolites | ME | Addition of phenolic compounds; different NaCl concentrations; pH 5.0–6.0 | SF | [68] |
| NG40/UV7 | CG | 112.0 g L−1 citric acid | ME; PE | Addition of inorganic nutrients (nitrogen, phosphorus, or sulfur); pulsed addition of glycerol; pH 5.0 | B | [69] |
| Wratislavia AWG7 | CG | 157.5 g L−1 citric acid | SE; PE | Acetate negative mutants; glycerol concentration; pulsed addition or constant feeding rate; pH 5.5 | B | [70] |
| A-101-1.22 | CG | 112.0 g L−1 citric acid | SE; PE | Acetate negative mutants; batch, repeated batch, cell recycle regimes; pH 5.5 | B | [71] |
| W29 | CG | 81.11 g L−1 citric acid; 24.90 g L−1 malic acid | ME | Optimization of the carbon and nitrogen source; pH 4.0 | B | [72] |
| A-101-B56-5 | PG | 57.15 g L−1 citric acid; invertase | SE; ME | SUC+ transformants; sucrose, mixture of glucose and fructose, glucose or glycerol; pH 6.8 | B | [73] |
| Strain | Carbon Source 1 | Product | Engineering 2 | Strategy | Set-Up 3 | Data from [Ref.] |
|---|---|---|---|---|---|---|
| Wratislavia K1 | CG | 40.7 g L−1 erythritol; 65 g L−1 citric acid | SE; ME | Acetate negative mutants; pH 3.0 for erythritol, pH 5.5/6.5 for citric acid; NaCl | B | [74] |
| Y. divulgata | CG | 59.8–62.7 g L−1 polyols from soap-derived; 76.8–79.5 g L−1 from biodiesel-derived glycerol | SS | 13 tested species of the Yarrowia clade; pH 3.0 | B | [75] |
| Wratislavia K1 | CG | 100.65 g L−1 polyols | SE; PE | Overexpression of heterologous SUC2 and native GUT1; step-wise fed-batch culture with molasses and CG; pH 3.0 | B | [97] |
| Wratislavia K1 | CG | 110 g L−1 citric acid; 81 g L−1 erythritol | SE; PE | Acetate negative mutant; fed-batch; pH 5.5 | B | [98] |
| Wratislavia K1 | CG | 170 g L−1 erythritol | SE; ME | Acetate negative mutant; effect of pH (2.5–6.5); pH 3.0 | B | [99] |
| A UV’1; A-15; Wratislavia K1 | CG | 80 g L−1 erythritol (Wratislavia K1); 27.6 g L−1 Mannitol (A UV’1, A-15) | SS; ME | Addition of NaCl; pH 3.0 | B | [100] |
| LFMB 19 | CG | 7.1 g L−1 mannitol | SS | Eukaryotic microbial strains (yeasts and Zygomycetes molds); pH 5.0–6.0 | SF | [101] |
| 2,021,417 | CG | 150 g L−1 erythritol | SE | Overexpression of GUT1, GUT2, and TKL1; Deletion of EYD1; pH 3.0 | B | [102] |
| Wratislavia K1 | PG | 132 g L−1 erythritol | ME | Concentrations of ammonium sulfate, monopotassium phosphate, and sodium chloride; C/N ratio; pH 3.0 | B | [76] |
| A101 | PG | 63.9 g L−1 citric ; 93 g L−1 citric ; 78 g L−1 erythritol | SE | Co-overexpression of GUT1 and GUT2; effect of pH (3.0 or 6.0) | B | [27] |
| MK1 | PG | 58 g L−1 erythritol; 62.5 g L−1 erythritol | SE | Overexpression of TKL1, TAL1, ZWF1, and GND1; pH 3.0 | SF; B | [103] |
| Po1d | PG | 80.6 g L−1 erythritol | SE | Overexpression of GUT1 and TKL1; disruption of EYK1; pH 3.0 | B | [104] |
| Wratislavia K1 | PG | 201.2 g L−1 erythritol | ME; PE | Addition of vitamins and nitrogen sources; batch and fed-batch; pH 3.0 | B | [105] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Theodosiou, E. Engineering Strategies for Efficient Bioconversion of Glycerol to Value-Added Products by Yarrowia lipolytica. Catalysts 2023, 13, 657. https://doi.org/10.3390/catal13040657
Theodosiou E. Engineering Strategies for Efficient Bioconversion of Glycerol to Value-Added Products by Yarrowia lipolytica. Catalysts. 2023; 13(4):657. https://doi.org/10.3390/catal13040657
Chicago/Turabian StyleTheodosiou, Eleni. 2023. "Engineering Strategies for Efficient Bioconversion of Glycerol to Value-Added Products by Yarrowia lipolytica" Catalysts 13, no. 4: 657. https://doi.org/10.3390/catal13040657