
A Novel NADP(H)-Dependent 7alpha-HSDH: Discovery and Construction of Substrate Selectivity Mutant by C-Terminal Truncation

Abstract
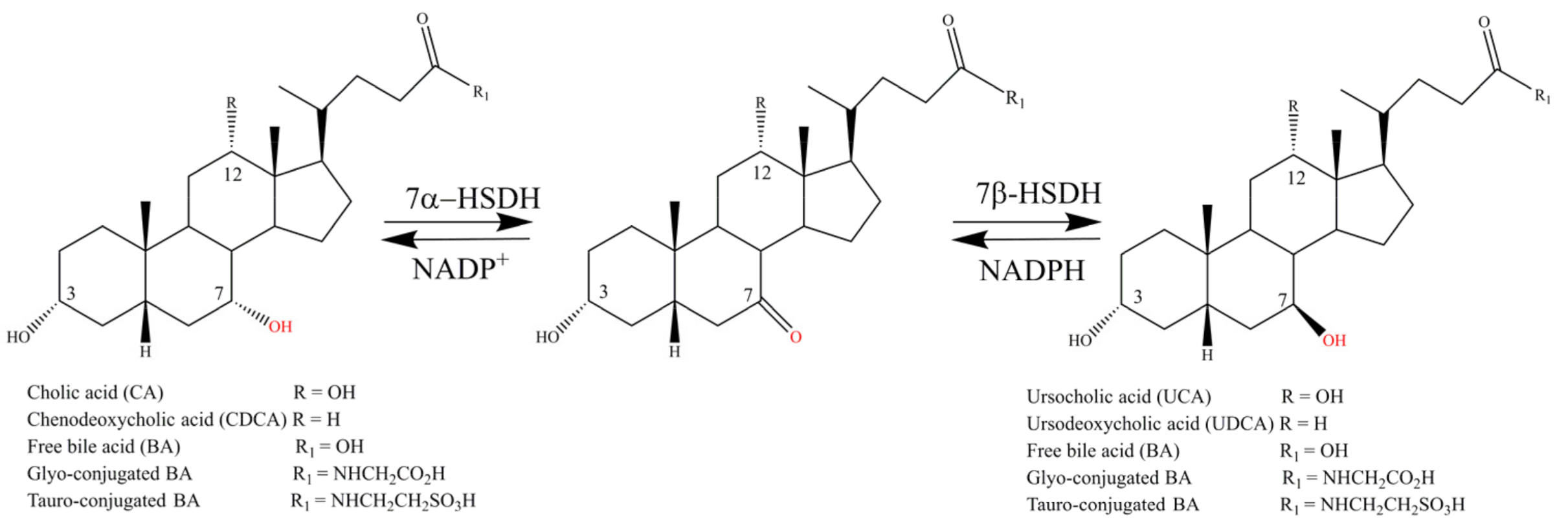
:1. Introduction
2. Results
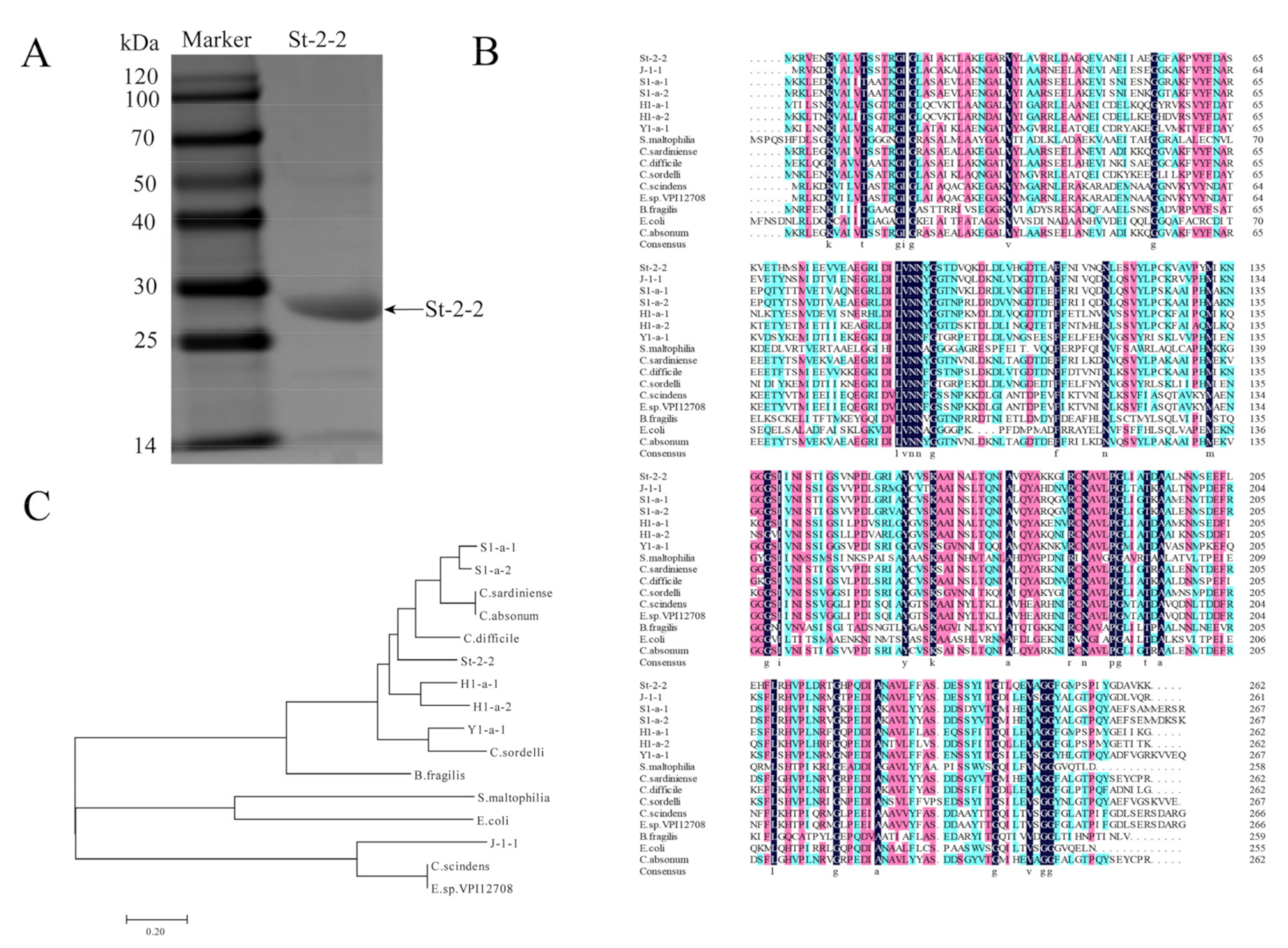
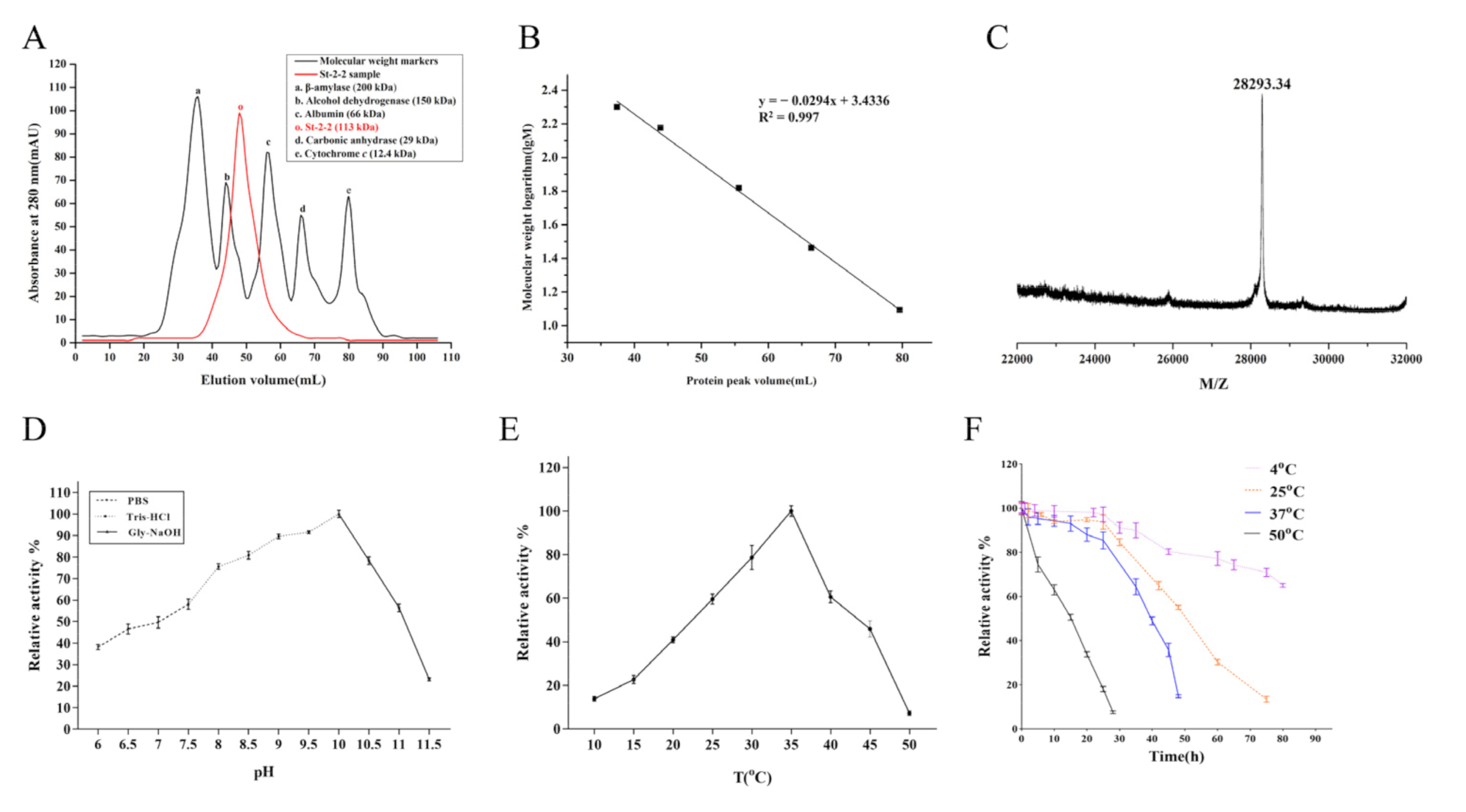
2.1. Cloning, Expression, and Purification of St-2-2
2.2. Sequence Alignments and Phylogenetic Analysis
2.3. Optimum pH, Optimum Temperature, and Thermostability of St-2-2
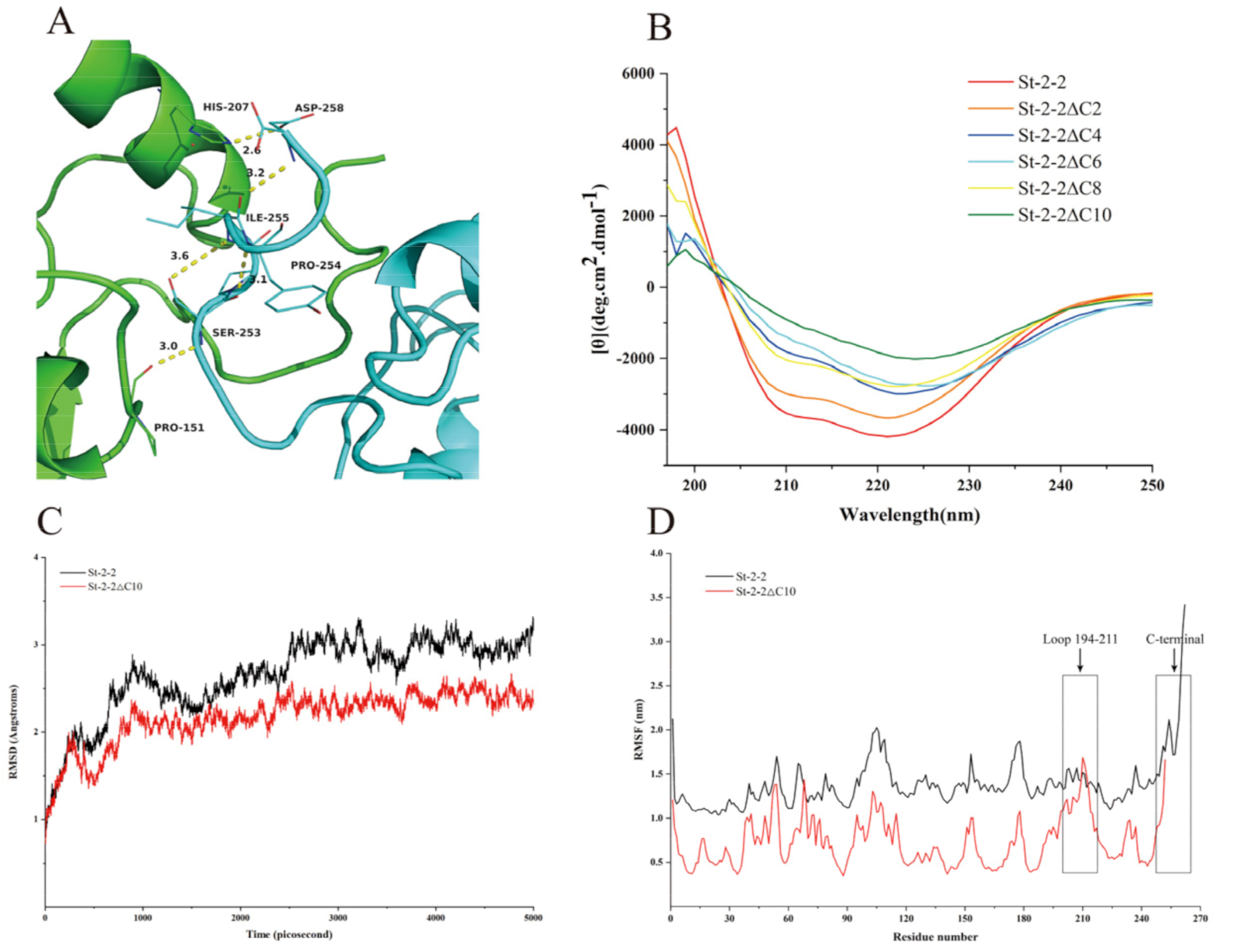
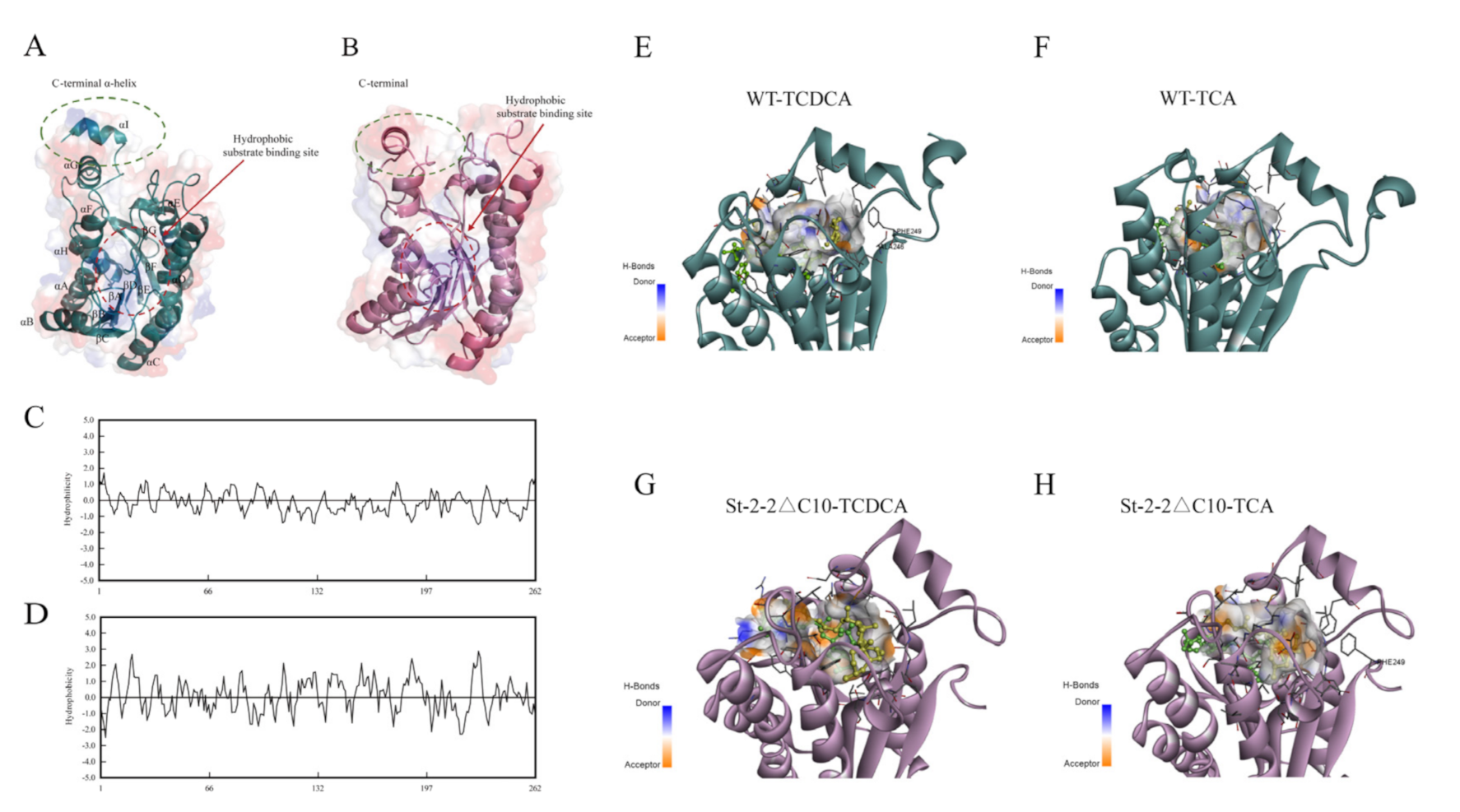
2.4. Structure Modeling and Target Mutation Identification
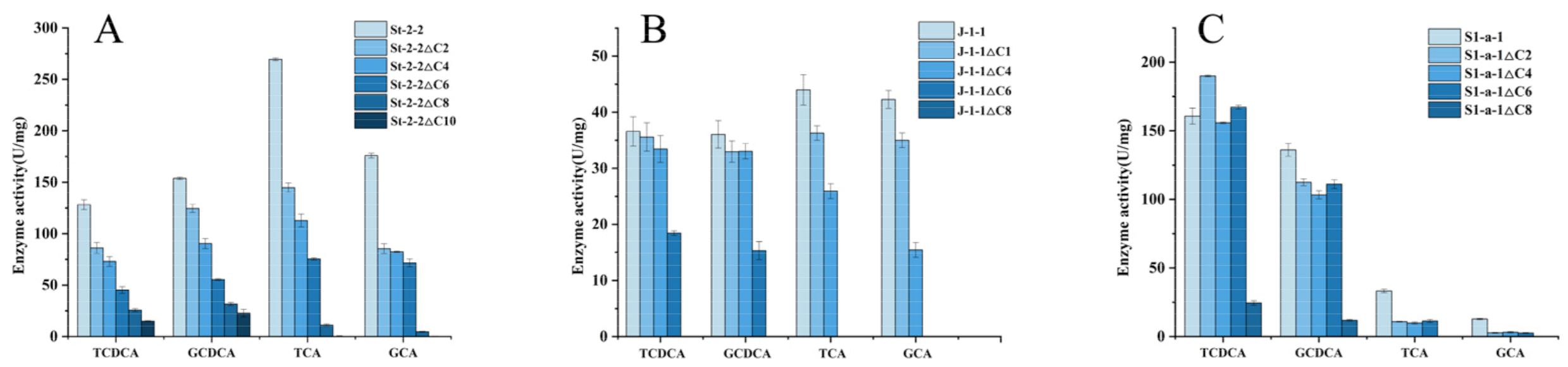
2.5. Effects of C-Terminal Domain Truncation of St-2-2 on Its Activity and Substrate Selectivity
2.6. MD Simulation and Structural Analysis
2.7. Verification of the C-Terminal Truncation Strategy on Other 7α-HSDHs
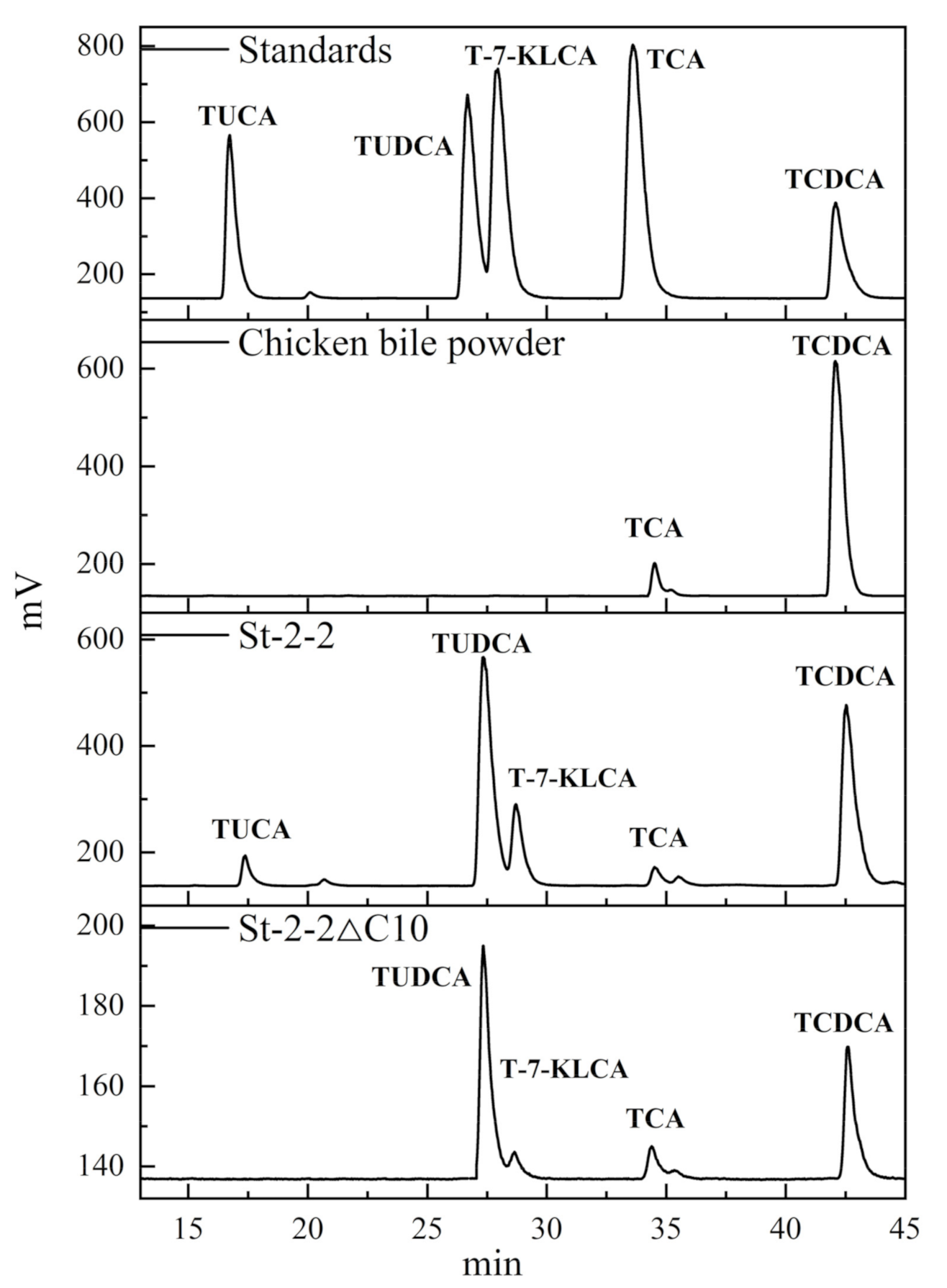
2.8. Application for Selective Production of TUDCA from Waste Chicken Bile Powder
3. Discussion
4. Materials & Methods
4.1. Chemicals and Materials
4.2. Molecular Cloning
4.3. Mutation Design and Truncated Mutagenesis
4.4. Expression and Purification of Recombinant Proteins
4.5. Phylogenetic Analysis and Sequence Alignment
4.6. Enzymatic Activity and Kinetic Analysis
4.7. Effects of pH and Temperature on Enzyme Activity and Stability
4.8. Circular Dichroism (CD) Spectroscopy Measurements
4.9. Homology Alignment, Structure Modeling, Molecular Docking, and MD Simulation
4.10. Practical Preparation of TUDCA from Waste Chicken Bile Powder Using St-2-2 and St-2-2∆C10
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| 7α-HSDH | 7α-hydroxysteroid dehydrogenase |
| GCA | glycocholic acid |
| GCDCA | glycochenodeoxycholic acid |
| TUDCA | tauroursodeoxycholic acid |
| TCA | taurocholic acid |
| TUCA | tauroursocholic acid |
| TCDCA | taurochenodeoxycholic acid |
| UDCA | ursodeoxycholic acid |
| T-7-KLCA | taurine-7-ketolithocholic acid |
References
- Tabrizian, K.; Shahramian, I.; Bazi, A.; Afshari, M.; Ghaemi, A. Alleviating Effects of Ursodeoxycholic Acid in Children with Acute Hepatitis A Infection: A Randomized Clinical Trial. Hepat. Mon. 2019, 19, e86719. [Google Scholar] [CrossRef] [Green Version]
- Suraweera, D.; Rahal, H.; Jimenez, M.; Viramontes, M.; Choi, G.; Saab, S. Treatment of primary biliary cholangitis ursodeoxycholic acid non-responders: A systematic review. Liver Int. 2017, 37, 1877–1886. [Google Scholar] [CrossRef] [PubMed]
- Crosignani, A.; Battezzati, P.M.; Setchell, K.D.R.; Invernizzi, P.; Covini, G.; Zuin, M.; Podda, M. Tauroursodeoxycholic Acid for Treatment of Primary Biliary Cirrhosis: A Dose-Response Study. Dig. Dis. Sci. 1996, 41, 809–815. [Google Scholar] [CrossRef] [PubMed]
- Tonin, F.; Arends, I.W.C.E. Latest development in the synthesis of ursodeoxycholic acid (UDCA): A critical review. Beilstein J. Org. Chem. 2018, 14, 470–483. [Google Scholar] [CrossRef] [Green Version]
- Momose, T.; Tsubaki, T.; Iida, T.; Nambara, T. An improved synthesis of taurine- and glycine-conjugated bile acids. Lipids 1997, 32, 775–778. [Google Scholar] [CrossRef]
- Chiang, J.Y.L. Bile acid metabolism and signaling in liver disease and therapy. Liver Res. 2017, 1, 3–9. [Google Scholar] [CrossRef]
- Fedorowski, T.; Salen, G.; Tint, G.S.; Mosbach, E. Transformation of Chenodeoxycholic Acid and Ursodeoxycholic Acid by Human Intestinal Bacteria. Gastroenterology 1979, 77, 1068–1073. [Google Scholar] [CrossRef]
- Lepercq, P.; Gérard, P.; Béguet, F.; Grill, J.-P.; Relano, P.; Cayuela, C.; Juste, C. Isolates from normal human intestinal flora but not lactic acid bacteria exhibit 7α- and 7β-hydroxysteroid dehydrogenase activities. Microb. Ecol. Health Dis. 2004, 16, 195–201. [Google Scholar] [CrossRef]
- Ferrandi, E.E.; Bertolesi, G.M.; Polentini, F.; Negri, A.; Riva, S.; Monti, D. In search of sustainable chemical processes: Cloning, recombinant expression, and functional characterization of the 7alpha- and 7beta-hydroxysteroid dehydrogenases from Clostridium absonum. Appl. Microbiol. Biotechnol. 2012, 95, 1221–1233. [Google Scholar] [CrossRef]
- Ridlon, J.M.; Kang, D.-J.; Hylemon, P.B. Bile salt biotransformations by human intestinal bacteria. J. Lipid Res. 2006, 47, 241–259. [Google Scholar] [CrossRef] [Green Version]
- Ji, Q.; Wang, B.; Li, C.; Hao, J.; Feng, W. Co-immobilised 7α- and 7β-HSDH as recyclable biocatalyst: High-performance production of TUDCA from waste chicken bile. RSC Adv. 2018, 8, 34192–34201. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wu, C.; Lu, D. Determination of bear bile acids in Fel Ursi and chick gall, duck gall and dog gall by capillary electrophoresis. Strait Pharm. J. 2006, 18, 61–63. [Google Scholar]
- Graff, M.; Buchholz, P.C.F.; Stockinger, P.; Bommarius, B.; Bommarius, A.S.; Pleiss, J. The Short-Chain Dehydrogenase/Reductase Engineering Database (SDRED): A classification and analysis system for a highly diverse enzyme family. Proteins 2019, 87, 443–451. [Google Scholar] [CrossRef]
- Sun, Y.; Calderini, E.; Kourist, R. A Reconstructed Common Ancestor of the Fatty Acid Photo-decarboxylase Clade Shows Photo-decarboxylation Activity and Increased Thermostability. ChemBioChem 2021, 22, 1833–1840. [Google Scholar] [CrossRef]
- Cahn, J.K.; Werlang, C.A.; Baumschlager, A.; Brinkmann-Chen, S.; Mayo, S.L.; Arnold, F.H. A General Tool for Engineering the NAD/NADP Cofactor Preference of Oxidoreductases. ACS Synth. Biol. 2017, 6, 326–333. [Google Scholar] [CrossRef] [Green Version]
- You, Z.N.; Chen, Q.; Shi, S.C.; Zheng, M.M.; Pan, J.; Qian, X.L.; Li, C.X.; Xu, J.H. Switching Cofactor Dependence of 7 beta-Hydroxysteroid Dehydrogenase for Cost-Effective Production of Ursodeoxycholic Acid. Acs Catalysis 2019, 9, 466–473. [Google Scholar] [CrossRef]
- Chen, Y.; Zhao, Y.; Zhou, X.; Liu, N.; Ming, D.; Zhu, L.; Jiang, L. Improving the thermostability of trehalose synthase from Thermomonospora curvata by covalent cyclization using peptide tags and investigation of the underlying molecular mechanism. Int. J. Biol. Macromol. 2021, 168, 13–21. [Google Scholar] [CrossRef]
- Kim, S.H.; Park, S.; Park, E.; Kim, J.H.; Ghatge, S.; Hur, H.G.; Rhee, S. Structure and substrate specificity determinants of NfnB, a dinitroaniline herbicide-catabolizing nitroreductase from Sphingopyxis sp. strain HMH. J. Biol. Chem. 2021, 297, 101143. [Google Scholar] [CrossRef]
- Kim, J.-S.; Patel, S.; Tiwari, M.; Lai, C.; Kumar, A.; Kim, Y.; Kalia, V.; Lee, J.-K. Phe-140 Determines the Catalytic Efficiency of Arylacetonitrilase from Alcaligenes faecalis. Int. J. Mol. Sci. 2020, 21, 7859. [Google Scholar] [CrossRef]
- Gao, H.; Li, J.; Sivakumar, D.; Kim, T.-S.; Patel, S.K.S.; Kalia, V.C.; Kim, I.-W.; Zhang, Y.-W.; Lee, J.-K. NADH oxidase from Lactobacillus reuteri: A versatile enzyme for oxidized cofactor regeneration. Int. J. Biol. Macromol. 2019, 123, 629–636. [Google Scholar] [CrossRef]
- Liu, Z.Y.; Zhang, R.Z.; Zhang, W.C.; Xu, Y. Ile258Met mutation of Brucella melitensis 7 alpha-hydroxysteroid dehydrogenase significantly enhances catalytic efficiency, cofactor affinity, and thermostability. Appl. Microbiol. Biotechnol. 2021, 105, 3573–3586. [Google Scholar] [CrossRef]
- Huang, B.; Zhao, Q.; Zhou, J.H.; Xu, G. Enhanced activity and substrate tolerance of 7-hydroxysteroid dehydrogenase by directed evolution for 7-ketolithocholic acid production. Appl. Microbiol. Biotechnol. 2019, 103, 2665–2674. [Google Scholar] [CrossRef]
- Dhagat, U.; Endo, S.; Mamiya, H.; Hara, A.; El-Kabbani, O. Studies on a Tyr residue critical for the binding of coenzyme and substrate in mouse 3(17)alpha-hydroxysteroid dehydrogenase (AKR1C21): Structure of the Y224D mutant enzyme. Acta Crystallogr. Sect. D-Struct. Biol. 2010, 66, 198–204. [Google Scholar] [CrossRef]
- Dhagat, U.; Endo, S.; Soda, M.; Hara, A.; El-Kabbani, O. Factorizing the role of a critical leucine residue in the binding of substrate to human 20 alpha-hydroxysteroid dehydrogenase (AKR1C1): Molecular modeling and kinetic studies of the Leu308Val mutant enzyme. Bioorg. Med. Chem. Lett. 2010, 20, 5274–5276. [Google Scholar] [CrossRef]
- Lou, D.; Wang, B.; Tan, J.; Zhu, L. Carboxyl-terminal and Arg38 are essential for activity of the 7α-hydroxysteroid dehydrogenase from Clostridium absonum. Protein Pept. Lett. 2014, 21, 894–900. [Google Scholar] [CrossRef]
- Kim, K.-H.; Lee, C.W.; Pardhe, B.D.; Hwang, J.; Do, H.; Lee, Y.M.; Lee, J.H.; Oh, T.-J. Crystal structure of an apo 7α-hydroxysteroid dehydrogenase reveals key structural changes induced by substrate and co-factor binding. J. Steroid Biochem. Mol. Biol. 2021, 212, 105945. [Google Scholar] [CrossRef]
- Pal, D.; Chakrabarti, P. Terminal residues in protein chains: Residue preference, conformation, and interaction. Biopolymers 2000, 53, 467–475. [Google Scholar] [CrossRef]
- Lou, D.; Wang, B.; Tan, J.; Zhu, L.; Cen, X.; Ji, Q.; Wang, Y.J.S.R. The three-dimensional structure of Clostridium absonum 7α-hydroxysteroid dehydrogenase: New insights into the conserved arginines for NADP(H) recognition. Sci. Rep. 2016, 6, 22885. [Google Scholar] [CrossRef] [Green Version]
- Tang, S.; Pan, Y.; Lou, D.; Ji, S.; Zhu, L.; Tan, J.; Qi, N.; Yang, Q.; Zhang, Z.; Yang, B.; et al. Structural and functional characterization of a novel acidophilic 7α-hydroxysteroid dehydrogenase. Protein Sci. 2019, 28, 910–919. [Google Scholar] [CrossRef]
- Tonin, F.; Otten, L.G.; Arends, I. NAD(+)-Dependent Enzymatic Route for the Epimerization of Hydroxysteroids. ChemSusChem. 2019, 12, 3192–3203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sebaihia, M.; Wren, B.W.; Mullany, P.; Fairweather, N.F.; Minton, N.; Stabler, R.; Thomson, N.R.; Roberts, A.P.; Cerdeño-Tárraga, A.M.; Wang, H.; et al. The multidrug-resistant human pathogen Clostridium difficile has a highly mobile, mosaic genome. Nat. Genet. 2006, 38, 779–786. [Google Scholar] [CrossRef] [PubMed]
- Coleman, J.P.; Hudson, L.L.; Adams, M.J. Characterization and regulation of the NADP-linked 7 alpha-hydroxysteroid dehydrogenase gene from Clostridium sordellii. J. Bacteriol. 1994, 176, 4865–4874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winter, J.; Morris, G.N.; O’Rourke-Locascio, S.; Bokkenheuser, V.D.; Mosbach, E.H.; Cohen, B.I.; Hylemon, P.B. Mode of action of steroid desmolase and reductases synthesized by Clostridium “scindens” (formerly Clostridium strain 19). J. Lipid Res. 1984, 25, 1124–1131. [Google Scholar] [CrossRef]
- Baron, S.F.; Franklund, C.V.; Hylemon, P.B. Cloning, sequencing, and expression of the gene coding for bile acid 7 alpha-hydroxysteroid dehydrogenase from Eubacterium sp. strain VPI 12708. J. Bacteriol. 1991, 173, 4558–4569. [Google Scholar] [CrossRef] [Green Version]
- Bennett, M.J.; McKnight, S.L.; Coleman, J.P. Cloning and characterization of the NAD-dependent 7alpha-Hydroxysteroid dehydrogenase from Bacteroides fragilis. Curr. Microbiol. 2003, 47, 475–484. [Google Scholar] [CrossRef]
- Leopold, S.R.; Magrini, V.; Holt, N.J.; Shaikh, N.; Mardis, E.R.; Cagno, J.; Ogura, Y.; Iguchi, A.; Hayashi, T.; Mellmann, A.; et al. A precise reconstruction of the emergence and constrained radiations of Escherichia coli O157 portrayed by backbone concatenomic analysis. Proc. Natl. Acad. Sci. USA 2009, 106, 8713–8718. [Google Scholar] [CrossRef] [Green Version]
- Filling, C.; Berndt, K.D.; Benach, J.; Knapp, S.; Prozorovski, T.; Nordling, E.; Ladenstein, R.; Jornvall, H.; Oppermann, U. Critical residues for structure and catalysis in short-chain dehydrogenases/reductases. J. Biol. Chem. 2002, 277, 25677–25684. [Google Scholar] [CrossRef] [Green Version]
- Song, C.; Wang, B.; Tan, J.; Zhu, L.; Lou, D. Discovery of tauroursodeoxycholic acid biotransformation enzymes from the gut microbiome of black bears using metagenomics. Sci. Rep. 2017, 7, 45495. [Google Scholar] [CrossRef]
- Lou, D.; Wang, Y.; Tan, J.; Zhu, L.; Ji, S.; Wang, B. Functional contribution of coenzyme specificity-determining sites of 7α-hydroxysteroid dehydrogenase from Clostridium absonum. Comput. Biol. Chem. 2017, 70, 89–95. [Google Scholar] [CrossRef]
- Martínez, L. Automatic identification of mobile and rigid substructures in molecular dynamics simulations and fractional structural fluctuation analysis. PLoS ONE 2015, 10, e0119264. [Google Scholar] [CrossRef] [Green Version]
- Ji, S.; Pan, Y.; Zhu, L.; Tan, J.; Tang, S.; Yang, Q.; Zhang, Z.; Lou, D.; Wang, B. A novel 7 alpha-hydroxysteroid dehydrogenase: Magnesium ion significantly enhances its activity and thermostability. Int. J. Biol. Macromol. 2021, 177, 111–118. [Google Scholar] [CrossRef]
- An, J.; Nie, Y.; Xu, Y. Structural insights into alcohol dehydrogenases catalyzing asymmetric reductions. Crit. Rev. Biotechnol. 2019, 39, 366–379. [Google Scholar] [CrossRef]
- Bakonyi, D.; Hummel, W. Cloning, expression, and biochemical characterization of a novel NADP+-dependent 7α-hydroxysteroid dehydrogenase from Clostridium difficile and its application for the oxidation of bile acids. Enzyme Microb. Technol. 2017, 99, 16–24. [Google Scholar] [CrossRef]
- Yoshimoto, T.; Higashi, H.; Kanatani, A.; Lin, X.S.; Nagai, H.; Oyama, H.; Kurazono, K.; Tsuru, D. Cloning and sequencing of the 7 alpha-hydroxysteroid dehydrogenase gene from Escherichia coli HB101 and characterization of the expressed enzyme. J. Bacteriol. 1991, 173, 2173–2179. [Google Scholar] [CrossRef] [Green Version]
- Pedrini, P.; Andreotti, E.; Guerrini, A.; Dean, M.; Fantin, G.; Giovannini, P.P. Xanthomonas maltophilia CBS 897.97 as a source of new 7β- and 7α-hydroxysteroid dehydrogenases and cholylglycine hydrolase: Improved biotransformations of bile acids. Steroids 2006, 71, 189–198. [Google Scholar] [CrossRef]
- Lee, J.; Goodey, N.M. Catalytic Contributions from Remote Regions of Enzyme Structure. Chem. Rev. 2011, 111, 7595–7624. [Google Scholar] [CrossRef]
- Yang, X.; Wu, L.; Li, A.; Ye, L.; Zhou, J.; Yu, H. The engineering of decameric d-fructose-6-phosphate aldolase a by combinatorial modulation of inter- and intra-subunit interactions. Chem. Commun. 2020, 56, 7561–7564. [Google Scholar] [CrossRef]
- Martínez, D.; Cutiño-Avila, B.; Pérez, E.R.; Menéndez, C.; Hernández, L.; del Monte-Martínez, A. A thermostable exo-β-fructosidase immobilised through rational design. Food Chem. 2014, 145, 826–831. [Google Scholar] [CrossRef]
- Ban, X.; Liu, Y.; Zhang, Y.; Gu, Z.; Li, C.; Cheng, L.; Hong, Y.; Dhoble, A.S.; Li, Z. Thermostabilization of a thermophilic 1,4-alpha-glucan branching enzyme through C-terminal truncation. Int. J. Biol. Macromol. 2018, 107, 1510–1518. [Google Scholar] [CrossRef]
- Gribenko, A.V.; Patel, M.M.; Liu, J.; McCallum, S.A.; Wang, C.; Makhatadze, G.I. Rational stabilization of enzymes by computational redesign of surface charge-charge interactions. Proc. Natl. Acad. Sci. USA 2009, 106, 2601–2606. [Google Scholar] [CrossRef] [Green Version]
- Arabnejad, H.; Dal Lago, M.; Jekel, P.A.; Floor, R.J.; Thunnissen, A.W.H.; Terwisscha van Scheltinga, A.C.; Wijma, H.J.; Janssen, D.B. A robust cosolvent-compatible halohydrin dehalogenase by computational library design. Protein Eng. Des. Sel. 2017, 30, 173–187. [Google Scholar] [CrossRef]
- Li, G.Y.; Yao, P.Y.; Gong, R.; Li, J.L.; Liu, P.; Lonsdale, R.; Wu, Q.Q.; Lin, J.P.; Zhu, D.M.; Reetz, M.T. Simultaneous engineering of an enzyme’s entrance tunnel and active site: The case of monoamine oxidase MAO-N. Chem. Sci. 2017, 8, 4093–4099. [Google Scholar] [CrossRef] [Green Version]
- Lou, D.S.; Tan, J.; Zhu, L.C.; Ji, S.L.; Tang, S.J.; Yao, K.Y.; Han, J.X.; Wang, B.C. Engineering Clostridium absonum 7 alpha-hydroxysteroid Dehydrogenase for Enhancing Thermostability Based on Flexible Site and Delta Delta G Prediction. Protein Pept. Lett. 2018, 25, 230–235. [Google Scholar] [CrossRef]
- Zheng, M.M.; Chen, K.C.; Wang, R.F.; Li, H.; Li, C.X.; Xu, J.H. Engineering 7 beta-Hydroxysteroid Dehydrogenase for Enhanced Ursodeoxycholic Acid Production by Multiobjective Directed Evolution. J. Agric. Food Chem. 2017, 65, 1178–1185. [Google Scholar] [CrossRef]
- Tanaka, N.; Nonaka, T.; Tanabe, T.; Yoshimoto, T.; Tsuru, D.; Mitsui, Y. Crystal Structures of the Binary and Ternary Complexes of 7α-Hydroxysteroid Dehydrogenase from Escherichia coli. Biochemistry 1996, 35, 7715–7730. [Google Scholar] [CrossRef]
- Chaudhari, S.N.; Luo, J.N.; Harris, D.A.; Aliakbarian, H.; Yao, L.; Paik, D.; Subramaniam, R.; Adhikari, A.A.; Vernon, A.H.; Kiliç, A.; et al. A microbial metabolite remodels the gut-liver axis following bariatric surgery. Cell Host Microbe. 2021, 29, 408–424. [Google Scholar] [CrossRef]
- Bernstein, C.; Holubec, H.; Bhattacharyya, A.K.; Nguyen, H.; Payne, C.M.; Zaitlin, B.; Bernstein, H. Carcinogenicity of deoxycholate, a secondary bile acid. Arch. Toxicol. 2011, 85, 863–871. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | Substrate | Vmax (U/mg) | Km (mM) | Kcat (S−1) | kcat/Km |
|---|---|---|---|---|---|
| St-2-2 | TCDCA | 152.85 ± 7.34 | 0.2 ± 0.01 | 102.34 ± 3.62 | 511.70 |
| St-2-2∆C10 | TCDCA | 22.17 ± 2.12 | 3.97 ± 0.15 | 10.42 ± 0.09 | 2.62 |
| J-1-1 | TCDCA | 387.14 ± 6.51 | 0.10 ± 0.01 | 192.86 ± 0.93 | 1872.40 |
| J-1-1∆C6 | TCDCA | 56.18 ± 1.91 | 2.59 ± 0.03 | 27.08 ± 0.21 | 10.46 |
| S1-a-1 | TCDCA | 369.23 ± 3.87 | 0.13 ± 0.01 | 236.54 ± 5.15 | 1791.95 |
| S1-a-1∆C8 | TCDCA | 51.81 ± 3.13 | 2.04 ± 0.11 | 24.97 ± 3.11 | 12.23 |
| TCDCA Conversionb (%) | TUDCA Yield (%) | The Proportion of TUDCA in Total Bile Acid (%) | |
|---|---|---|---|
| St-2-2 | 79.93 | 31.25 | 25.08 |
| St-2-2∆C10 | 84.23 | 35.34 | 26.15 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pan, Y.; Tang, S.; Zhou, M.; Ao, F.; Tang, Z.; Zhu, L.; Lou, D.; Tan, J.; Wang, B. A Novel NADP(H)-Dependent 7alpha-HSDH: Discovery and Construction of Substrate Selectivity Mutant by C-Terminal Truncation. Catalysts 2022, 12, 781. https://doi.org/10.3390/catal12070781
Pan Y, Tang S, Zhou M, Ao F, Tang Z, Zhu L, Lou D, Tan J, Wang B. A Novel NADP(H)-Dependent 7alpha-HSDH: Discovery and Construction of Substrate Selectivity Mutant by C-Terminal Truncation. Catalysts. 2022; 12(7):781. https://doi.org/10.3390/catal12070781
Chicago/Turabian StylePan, Yinping, Shijin Tang, Minghai Zhou, Fanglin Ao, Zhuozhou Tang, Liancai Zhu, Deshuai Lou, Jun Tan, and Bochu Wang. 2022. "A Novel NADP(H)-Dependent 7alpha-HSDH: Discovery and Construction of Substrate Selectivity Mutant by C-Terminal Truncation" Catalysts 12, no. 7: 781. https://doi.org/10.3390/catal12070781