
Upregulation of Cell Surface Glycoproteins in Correlation with KSHV LANA in the Kaposi Sarcoma Tumor Microenvironment
 , ,
, ,
Abstract
:Simple Summary
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Patient Sample Collection
2.2. Compliance
2.3. Cell Culture
2.4. L1T2 Murine Xenograft Generation
2.5. Immunohistochemistry (IHC)
2.6. Dual-Immunofluorescence (IF)
2.7. Imaging
2.8. Statistical Analysis
3. Results
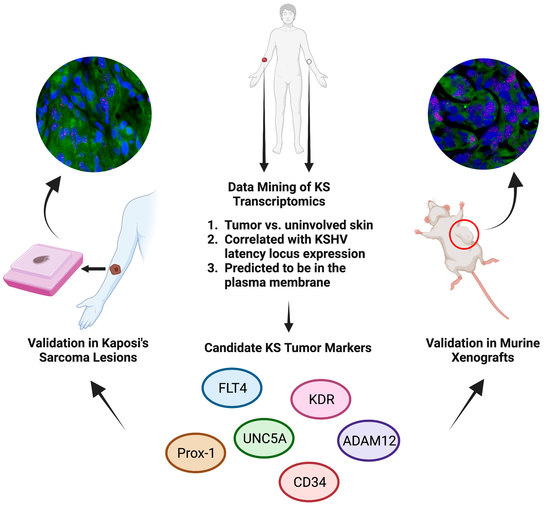
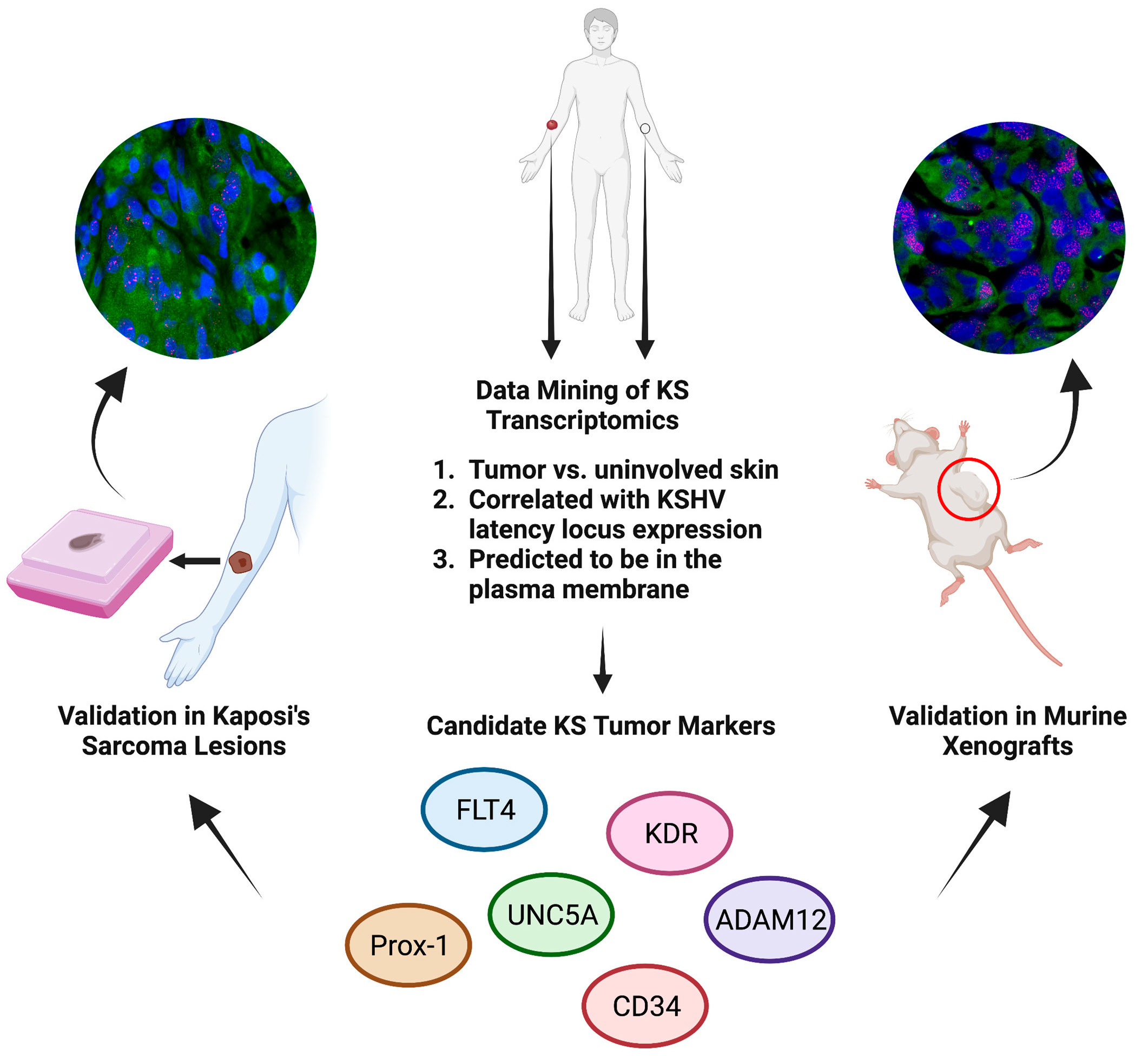
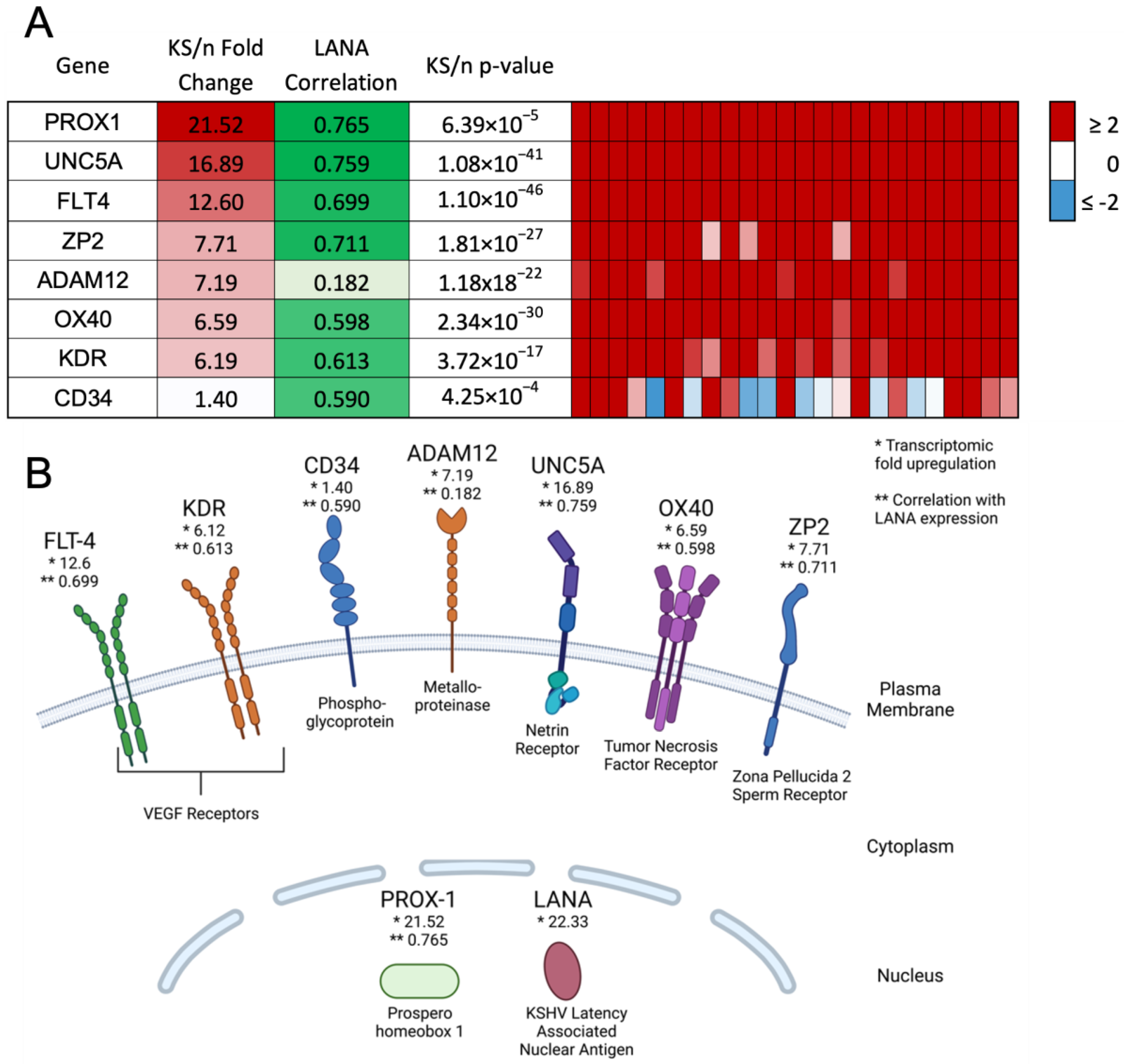
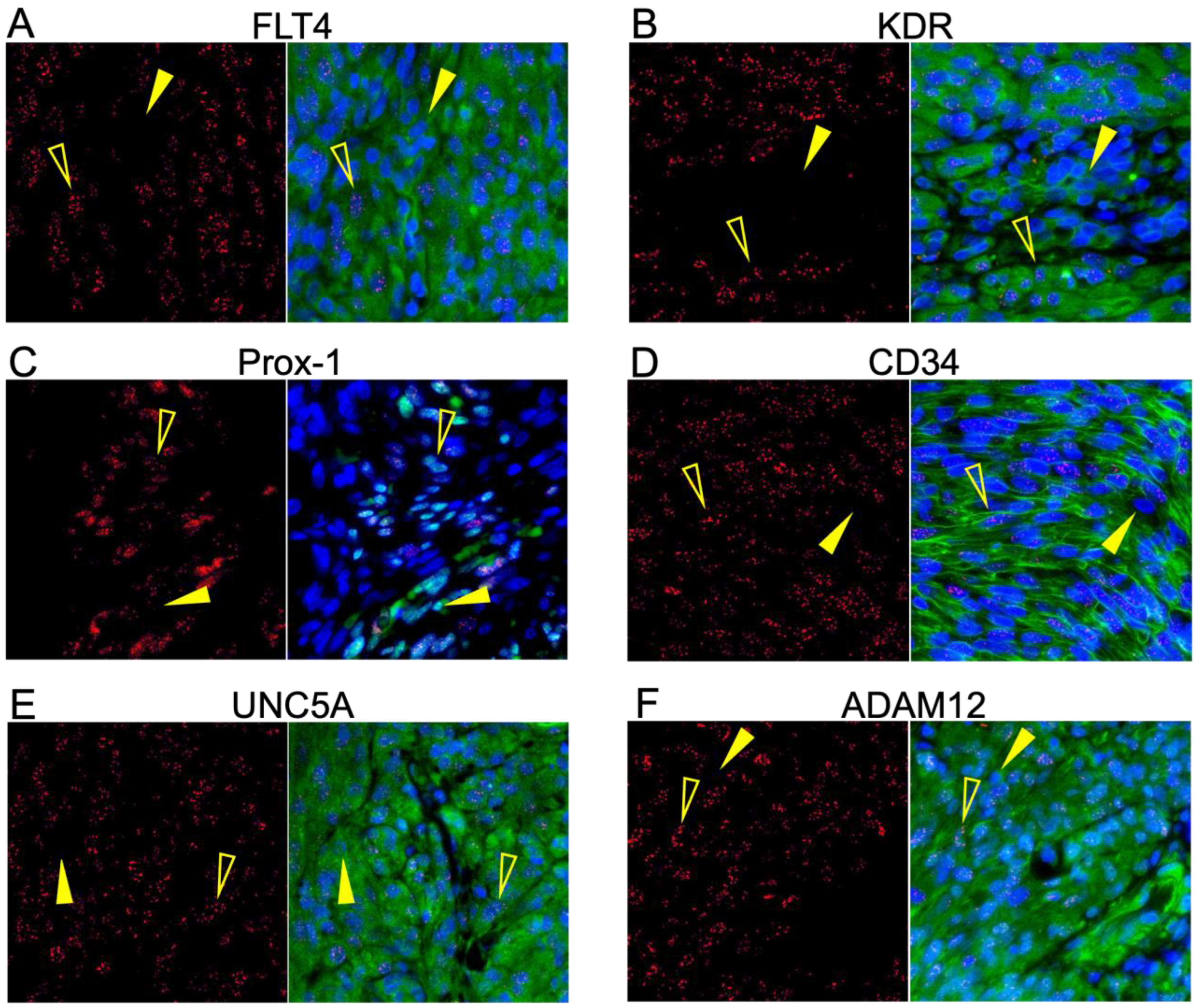
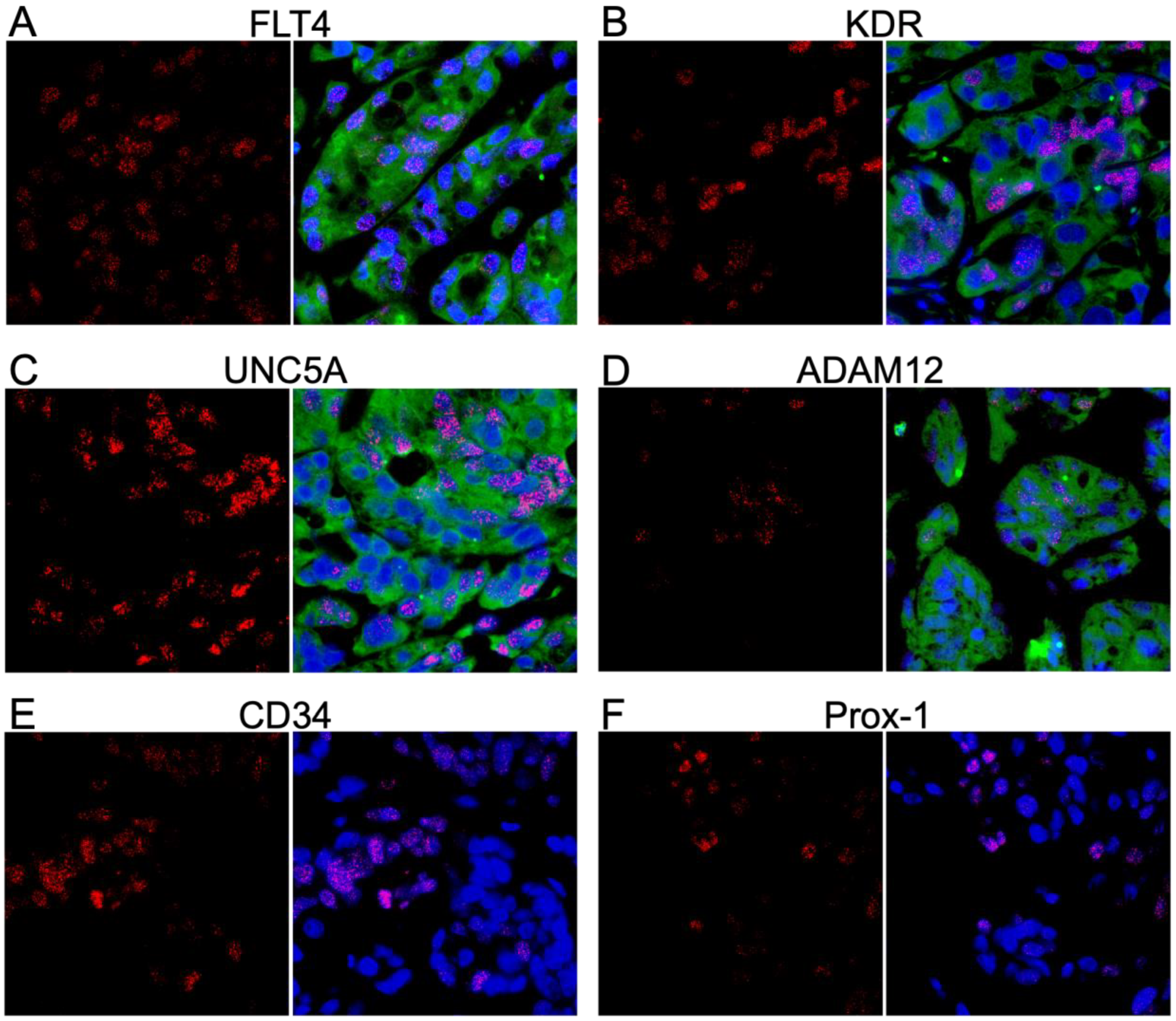
3.1. Identification of Glycoprotein Transcripts That Correlate with KSHV LANA
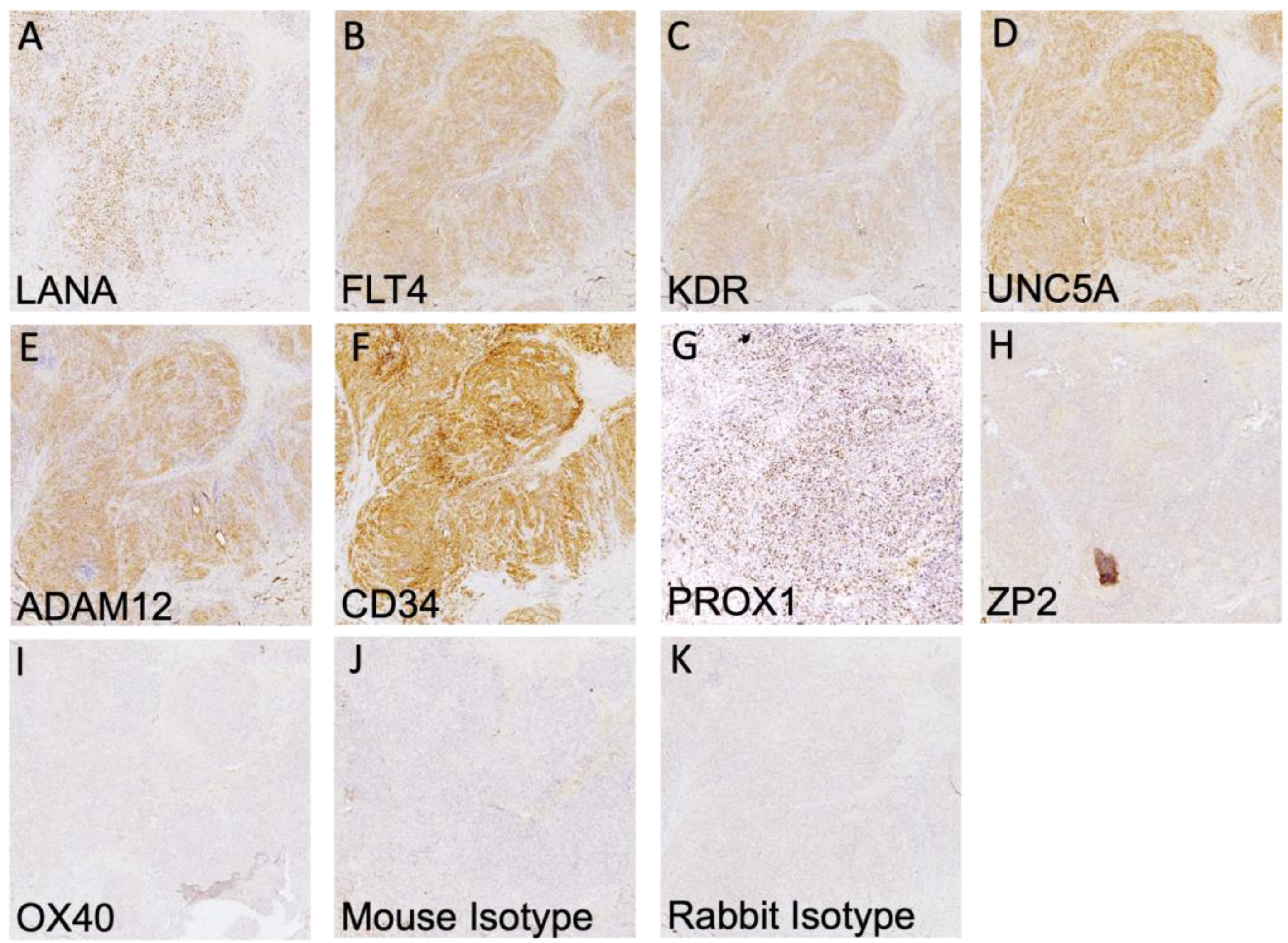
3.2. Validation of Protein Expression in KS Tissues
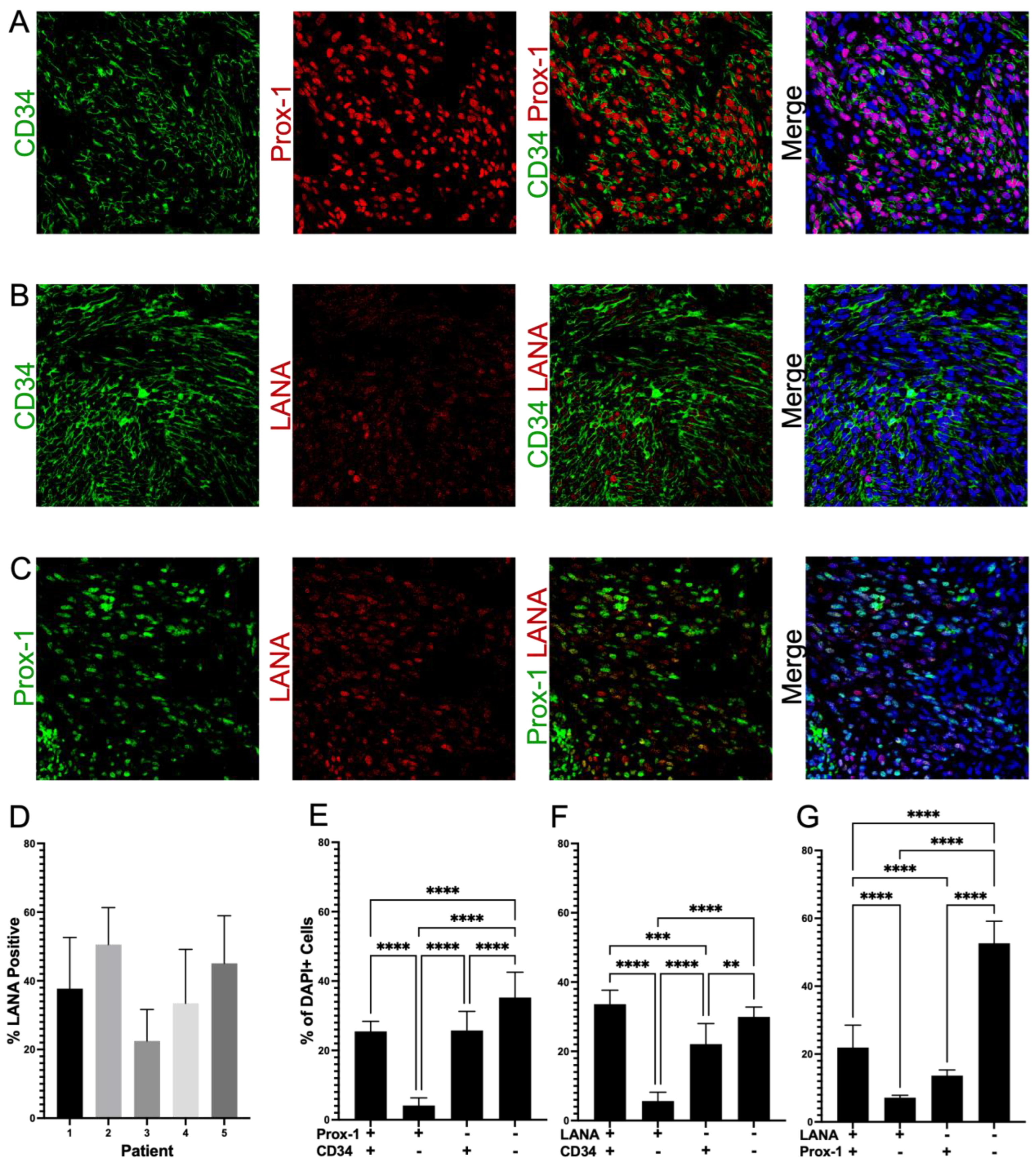
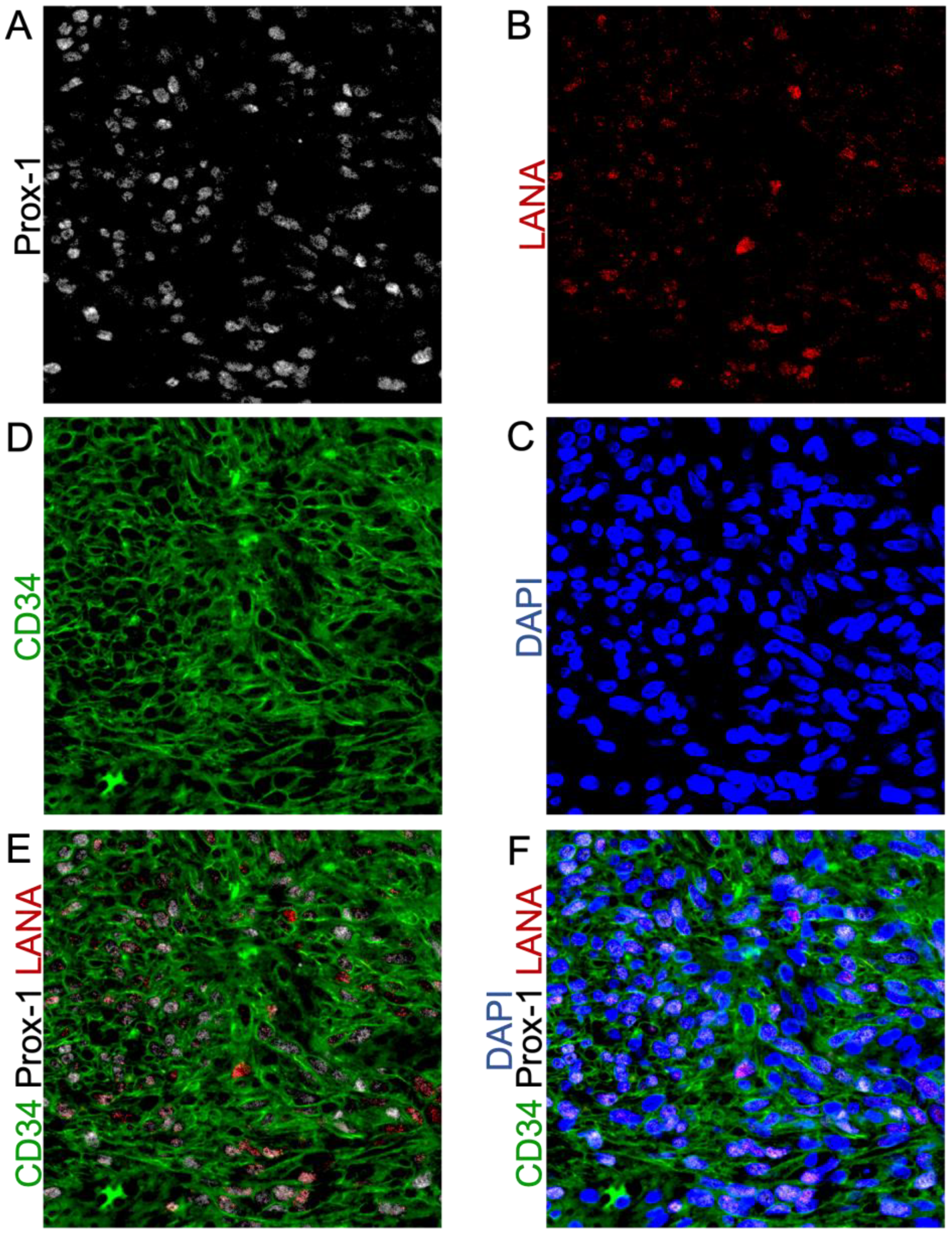
3.3. Highly Expressed Proteins Are Not Exclusive to KSHV-Infected Tumor Cells
3.4. Cells in KS Lesions Co-Express Prox-1 and CD34
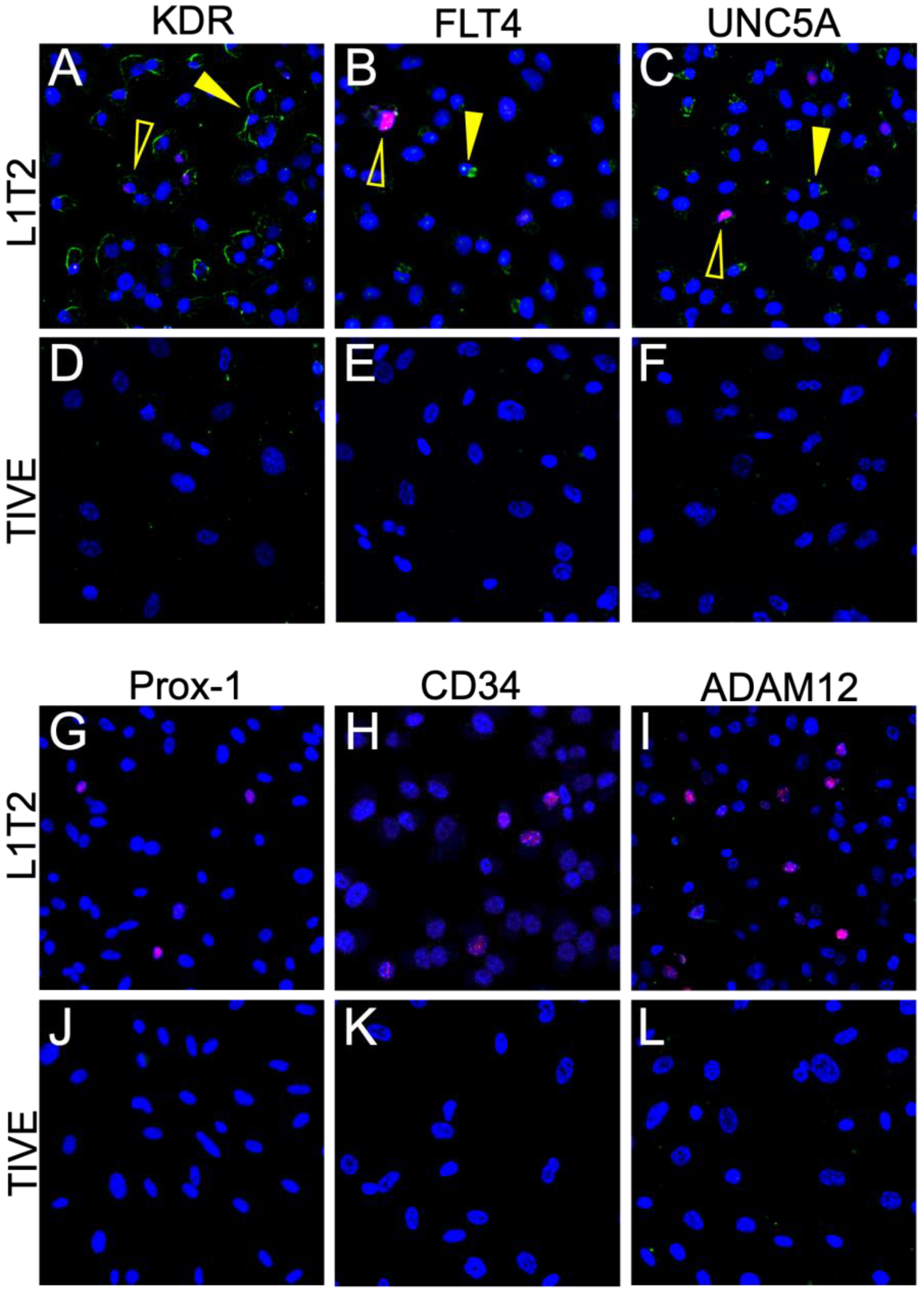
3.5. Limited Protein Expression in KSHV-Infected Endothelial Cell Lines
3.6. L1T2-Derived Xenografts Reflect the Protein Expression Observed in Human KS Tissues
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chang, Y.; Cesarman, E.; Pessin, M.S.; Lee, F.; Culpepper, J.; Knowles, D.M.; Moore, P.S. Identification of herpesvirus-like DNA sequences in AIDS-associated Kaposi’s sarcoma. Science 1994, 266, 1865–1869. [Google Scholar] [CrossRef] [Green Version]
- Moore, P.S.; Chang, Y. Detection of herpesvirus-like DNA sequences in Kaposi’s sarcoma in patients with and those without HIV infection. N. Engl. J. Med. 1995, 332, 1181–1185. [Google Scholar] [CrossRef] [Green Version]
- Cesarman, E.; Damania, B.; Krown, S.E.; Martin, J.; Bower, M.; Whitby, D. Kaposi sarcoma. Nat. Rev. Dis. Prim. 2019, 5, 9. [Google Scholar] [CrossRef] [PubMed]
- Ragi, S.D.; Moseley, I.; Ouellette, S.; Rao, B. Epidemiology and Survival of Kaposi’s Sarcoma by Race in the United States: A Surveillance, Epidemiology, and End Results Database Analysis. Clin. Cosmet. Investig. Dermatol. 2022, 15, 1681–1685. [Google Scholar] [CrossRef]
- Grabar, S.; Costagliola, D. Epidemiology of kaposi’s sarcoma. Cancers 2021, 13, 5692. [Google Scholar] [CrossRef]
- Motlhale, M.; Sitas, F.; Bradshaw, D.; Chen, W.C.; Singini, M.G.; de Villiers, C.B.; Lewis, C.M.; Muchengeti, M.; Waterboer, T.; Mathew, C.G.; et al. Epidemiology of Kaposi’s sarcoma in sub-Saharan Africa. Cancer Epidemiol. 2022, 78, 102167. [Google Scholar] [CrossRef] [PubMed]
- Grulich, A.E.; Vajdic, C.M. The epidemiology of cancers in human immunodeficiency virus infection and after organ transplantation. Semin. Oncol. 2015, 42, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Friedman-Kien, A.E. Disseminated Kaposi’s sarcoma syndrome in young homosexual men. J. Am. Acad. Dermatol. 1981, 5, 468–471. [Google Scholar] [CrossRef]
- Ngalamika, O.; Munsaka, S.; Lidenge, S.J.; West, J.T.; Wood, C. Antiretroviral Therapy for HIV-Associated Cutaneous Kaposi’s Sarcoma: Clinical, HIV-Related, and Sociodemographic Predictors of Outcome. AIDS Res. Hum. Retrovir. 2021, 37, 368–372. [Google Scholar] [CrossRef]
- Pyakurel, P.; Pak, F.; Mwakigonja, A.R.; Kaaya, E.; Heiden, T.; Biberfeld, P. Lymphatic and vascular origin of Kaposi’s sarcoma spindle cells during tumor development. Int. J. Cancer 2006, 119, 1262–1267. [Google Scholar] [CrossRef] [PubMed]
- Kerr, D.A.; Busarla, S.V.P.; Gimbel, D.C.; Sohani, A.R.; Nazarian, R.M. mTOR, VEGF, PDGFR, and c-kit signaling pathway activation in Kaposi sarcoma. Hum. Pathol. 2017, 65, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Lidenge, S.J.; Kossenkov, A.V.; Tso, F.Y.; Wickramasinghe, J.; Privatt, S.R.; Ngalamika, O.; Ngowi, J.R.; Mwaiselage, J.; Lieberman, P.M.; West, J.T.; et al. Comparative transcriptome analysis of endemic and epidemic Kaposi’s sarcoma (KS) lesions and the secondary role of HIV-1 in KS pathogenesis. PLoS Pathog. 2020, 16, e1008681. [Google Scholar] [CrossRef] [PubMed]
- Tso, F.Y.; Kossenkov, A.V.; Lidenge, S.J.; Ngalamika, O.; Ngowi, J.R.; Mwaiselage, J.; Wickramasinghe, J.; Kwon, E.H.; West, J.T.; Lieberman, P.M.; et al. RNA-Seq of Kaposi’s sarcoma reveals alterations in glucose and lipid metabolism. PLoS Pathog. 2018, 14, e1006844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirakawa, S.; Hong, Y.K.; Harvey, N.; Schacht, V.; Matsuda, K.; Libermann, T.; Detmar, M. Identification of vascular lineage-specific genes by transcriptional profiling of isolated blood vascular and lymphatic endothelial cells. Am. J. Pathol. 2003, 162, 575–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, D.; Park, E.; Kim, K.E.; Jung, E.; Seong, Y.J.; Zhao, L.; Madhavan, S.; Daghlian, G.; Lee, H.H.; Daghlian, P.T.; et al. The Lymphatic Cell Environment Promotes Kaposi Sarcoma Development by Prox1-Enhanced Productive Lytic Replication of Kaposi Sarcoma Herpes Virus. Cancer Res. 2020, 80, 3130–3144. [Google Scholar] [CrossRef]
- Ntikoudi, E.; Pergaris, A.; Kykalos, S.; Politi, E.; Theocharis, S. The Role of PROX1 in Neoplasia: A Key Player Often Overlooked. Diagnostics 2022, 12, 1624. [Google Scholar] [CrossRef]
- Kanitakis, J.; Narvaez, D.; Claudy, A. Expression of the CD34 antigen distinguishes Kaposi’s sarcoma from pseudo-Kaposi’s sarcoma (acroangiodermatitis). Br. J. Dermatol. 1996, 134, 44–46. [Google Scholar] [CrossRef] [PubMed]
- Russell Jones, R.; Orchard, G.; Zelger, B.; Wilson Jones, E. Immunostaining for CD31 and CD34 in Kaposi sarcoma. J. Clin. Pathol. 1995, 48, 1011–1016. [Google Scholar] [CrossRef] [Green Version]
- An, F.Q.; Folarin, H.M.; Compitello, N.; Roth, J.; Gerson, S.L.; McCrae, K.R.; Fakhari, F.D.; Dittmer, D.P.; Renne, R. Long-term-infected telomerase-immortalized endothelial cells: A model for Kaposi’s sarcoma-associated herpesvirus latency in vitro and in vivo. J. Virol. 2006, 80, 4833–4846. [Google Scholar] [CrossRef] [Green Version]
- Roy, D.; Sin, S.H.; Lucas, A.; Venkataramanan, R.; Wang, L.; Eason, A.; Chavakula, V.; Hilton, I.B.; Tamburro, K.M.; Damania, B.; et al. mTOR inhibitors block Kaposi sarcoma growth by inhibiting essential autocrine growth factors and tumor angiogenesis. Cancer Res. 2013, 73, 2235–2246. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.X.; Kang, G.; Kumar, P.; Lu, W.; Li, Y.; Zhou, Y.; Li, Q.; Wood, C. Humanized-BLT mouse model of Kaposi’s sarcoma-associated herpesvirus infection. Proc. Natl. Acad. Sci. USA 2014, 111, 3146–3151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tso, F.Y.; Sawyer, A.; Kwon, E.H.; Mudenda, V.; Langford, D.; Zhou, Y.; West, J.; Wood, C. Kaposi’s Sarcoma-Associated Herpesvirus Infection of Neurons in HIV-Positive Patients. J. Infect. Dis. 2017, 215, 1898–1907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- AbuSamra, D.B.; Aleisa, F.A.; Al-Amoodi, A.S.; Jalal Ahmed, H.M.; Chin, C.J.; Abuelela, A.F.; Bergam, P.; Sougrat, R.; Merzaban, J.S. Not just a marker: CD34 on human hematopoietic stem/progenitor cells dominates vascular selectin binding along with CD44. Blood Adv. 2017, 1, 2799–2816. [Google Scholar] [CrossRef] [Green Version]
- Melincovici, C.S.; Bosca, A.B.; Susman, S.; Marginean, M.; Mihu, C.; Istrate, M.; Moldovan, I.M.; Roman, A.L.; Mihu, C.M. Vascular endothelial growth factor (VEGF)–key factor in normal and pathological angiogenesis. Rom. J. Morphol. Embryol. 2018, 59, 455–467. [Google Scholar] [PubMed]
- Liu, Y.; Beyer, A.; Aebersold, R. On the Dependency of Cellular Protein Levels on mRNA Abundance. Cell 2016, 165, 535–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koussounadis, A.; Langdon, S.P.; Um, I.H.; Harrison, D.J.; Smith, V.A. Relationship between differentially expressed mRNA and mRNA-protein correlations in a xenograft model system. Sci. Rep. 2015, 5, 10775. [Google Scholar] [CrossRef] [Green Version]
- Lidenge, S.J.; Tso, F.Y.; Ngalamika, O.; Kolape, J.; Ngowi, J.R.; Mwaiselage, J.; Wood, C.; West, J.T. Lack of CD8+ T-cell co-localization with Kaposi’s sarcoma-associated herpesvirus infected cells in Kaposi’s sarcoma tumors. Oncotarget 2020, 11, 1556–1572. [Google Scholar] [CrossRef]
- Litscher, E.S.; Wassarman, P.M. Zona Pellucida Proteins, Fibrils, and Matrix. Annu. Rev. Biochem. 2020, 89, 695–715. [Google Scholar] [CrossRef]
- Wegler, C.; Olander, M.; Wisniewski, J.R.; Lundquist, P.; Zettl, K.; Asberg, A.; Hjelmesaeth, J.; Andersson, T.B.; Artursson, P. Global variability analysis of mRNA and protein concentrations across and within human tissues. NAR Genom. Bioinform. 2020, 2, lqz010. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Bockermann, R.; Hadi, M.; Safari, I.; Carrion, B.; Kveiborg, M.; Issazadeh-Navikas, S. ADAM12 is a costimulatory molecule that determines Th1 cell fate and mediates tissue inflammation. Cell. Mol. Immunol. 2021, 18, 1904–1919. [Google Scholar] [CrossRef]
- Wang, R.; Godet, I.; Yang, Y.; Salman, S.; Lu, H.; Lyu, Y.; Zuo, Q.; Wang, Y.; Zhu, Y.; Chen, C.; et al. Hypoxia-inducible factor-dependent ADAM12 expression mediates breast cancer invasion and metastasis. Proc. Natl. Acad. Sci. USA 2021, 118, e2020490118. [Google Scholar] [CrossRef] [PubMed]
- Atfi, A.; Dumont, E.; Colland, F.; Bonnier, D.; L’Helgoualc’h, A.; Prunier, C.; Ferrand, N.; Clement, B.; Wewer, U.M.; Theret, N. The disintegrin and metalloproteinase ADAM12 contributes to TGF-beta signaling through interaction with the type II receptor. J. Cell Biol. 2007, 178, 201–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, Y.; Chen, W.; Lu, Z.; Wang, Y.; Yuan, Y. Kaposi’s sarcoma-associated herpesvirus promotes mesenchymal-to-endothelial transition by resolving the bivalent chromatin of PROX1 gene. PLoS Pathog. 2021, 17, e1009847. [Google Scholar] [CrossRef]
- Li, Y.; Zhong, C.; Liu, D.; Yu, W.; Chen, W.; Wang, Y.; Shi, S.; Yuan, Y. Evidence for Kaposi Sarcoma Originating from Mesenchymal Stem Cell through KSHV-induced Mesenchymal-to-Endothelial Transition. Cancer Res. 2018, 78, 230–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruce, A.G.; Barcy, S.; DiMaio, T.; Gan, E.; Garrigues, H.J.; Lagunoff, M.; Rose, T.M. Quantitative Analysis of the KSHV Transcriptome Following Primary Infection of Blood and Lymphatic Endothelial Cells. Pathogens 2017, 6, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiedler, U.; Christian, S.; Koidl, S.; Kerjaschki, D.; Emmett, M.S.; Bates, D.O.; Christofori, G.; Augustin, H.G. The sialomucin CD34 is a marker of lymphatic endothelial cells in human tumors. Am. J. Pathol. 2006, 168, 1045–1053. [Google Scholar] [CrossRef] [Green Version]
- Simons, M.; Gordon, E.; Claesson-Welsh, L. Mechanisms and regulation of endothelial VEGF receptor signalling. Nat. Rev. Mol. Cell Biol. 2016, 17, 611–625. [Google Scholar] [CrossRef]
- Maslowska, K.; Halik, P.K.; Tymecka, D.; Misicka, A.; Gniazdowska, E. The Role of VEGF Receptors as Molecular Target in Nuclear Medicine for Cancer Diagnosis and Combination Therapy. Cancers 2021, 13, 1072. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Privatt, S.R.; Ngalamika, O.; Zhang, J.; Li, Q.; Wood, C.; West, J.T. Upregulation of Cell Surface Glycoproteins in Correlation with KSHV LANA in the Kaposi Sarcoma Tumor Microenvironment. Cancers 2023, 15, 2171. https://doi.org/10.3390/cancers15072171
Privatt SR, Ngalamika O, Zhang J, Li Q, Wood C, West JT. Upregulation of Cell Surface Glycoproteins in Correlation with KSHV LANA in the Kaposi Sarcoma Tumor Microenvironment. Cancers. 2023; 15(7):2171. https://doi.org/10.3390/cancers15072171
Chicago/Turabian StylePrivatt, Sara R., Owen Ngalamika, Jianshui Zhang, Qinsheng Li, Charles Wood, and John T. West. 2023. "Upregulation of Cell Surface Glycoproteins in Correlation with KSHV LANA in the Kaposi Sarcoma Tumor Microenvironment" Cancers 15, no. 7: 2171. https://doi.org/10.3390/cancers15072171