
Multiple Genes with Potential Tumor Suppressive Activity Are Present on Chromosome 10q Loss in Neuroblastoma and Are Associated with Poor Prognosis
 , , , , and
, , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Primary Tumors and Cell Lines
2.2. Genomic Profile Analysis
2.3. Data Sets
2.3.1. Italian Cohort of NB Patients
2.3.2. In Silico Data Sets
2.4. Statistical Analysis
3. Results
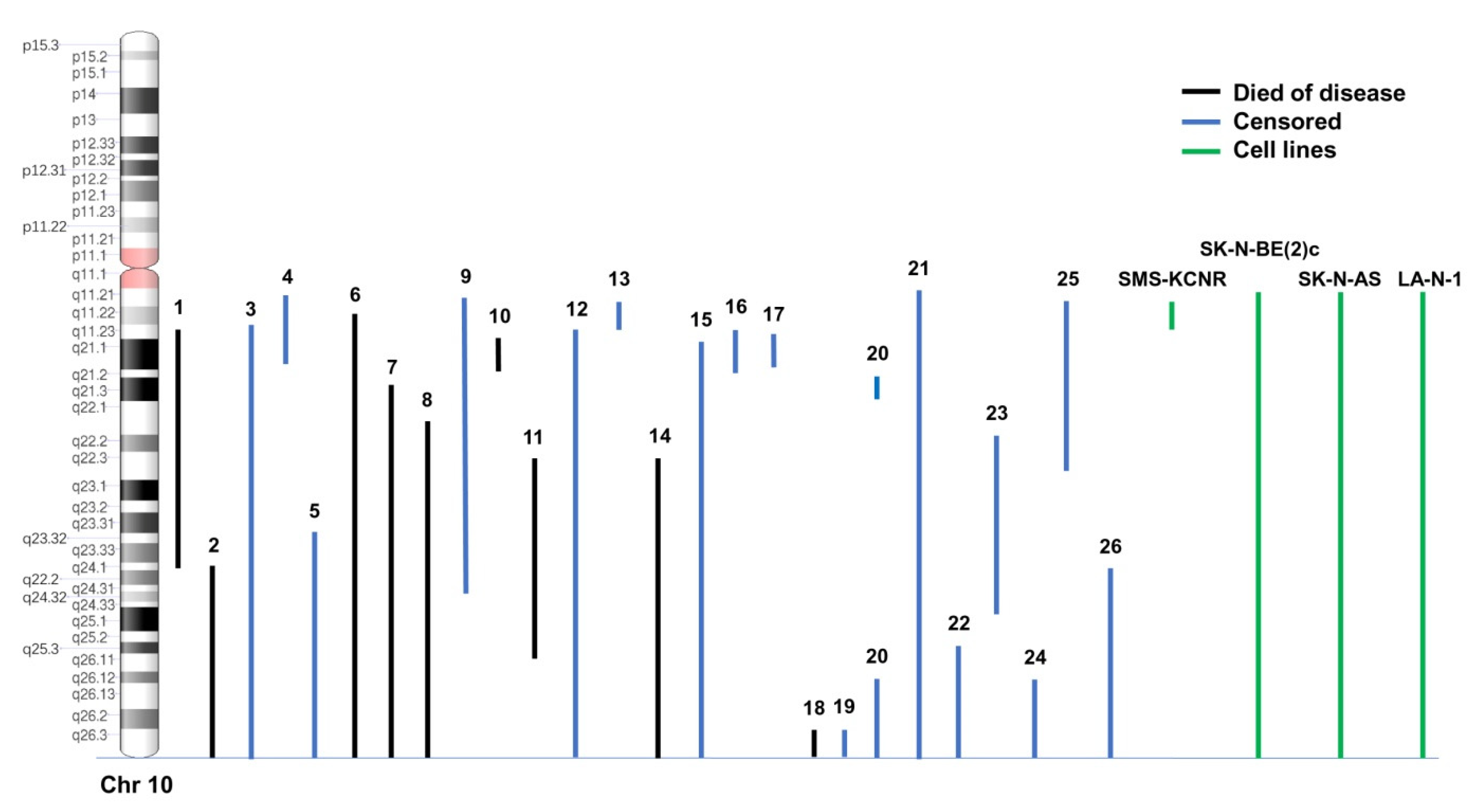
3.1. Analysis of 10q Loss in NB Samples and Cell Lines
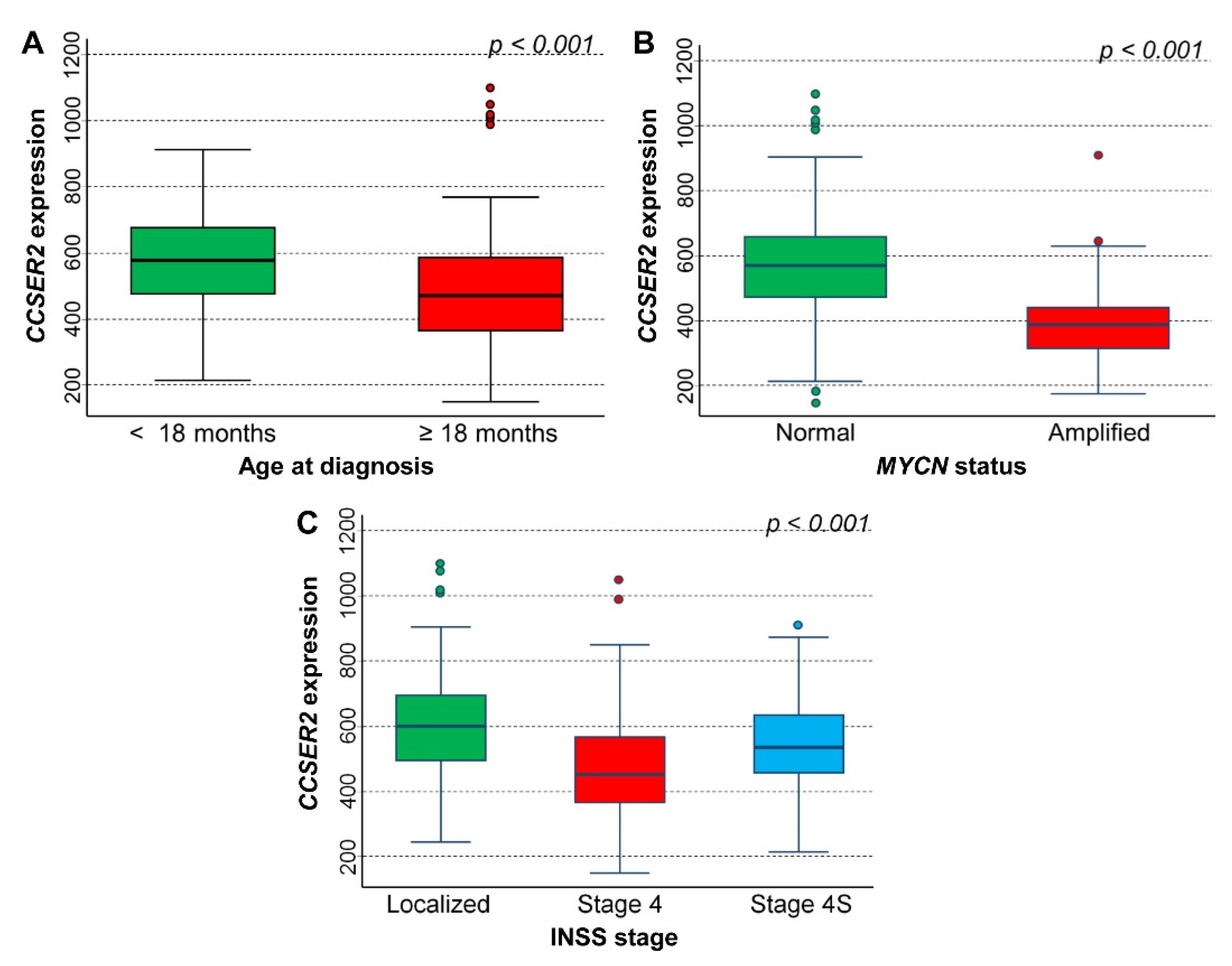
3.2. Association between NB Patients’ Characteristics and 10q Loss
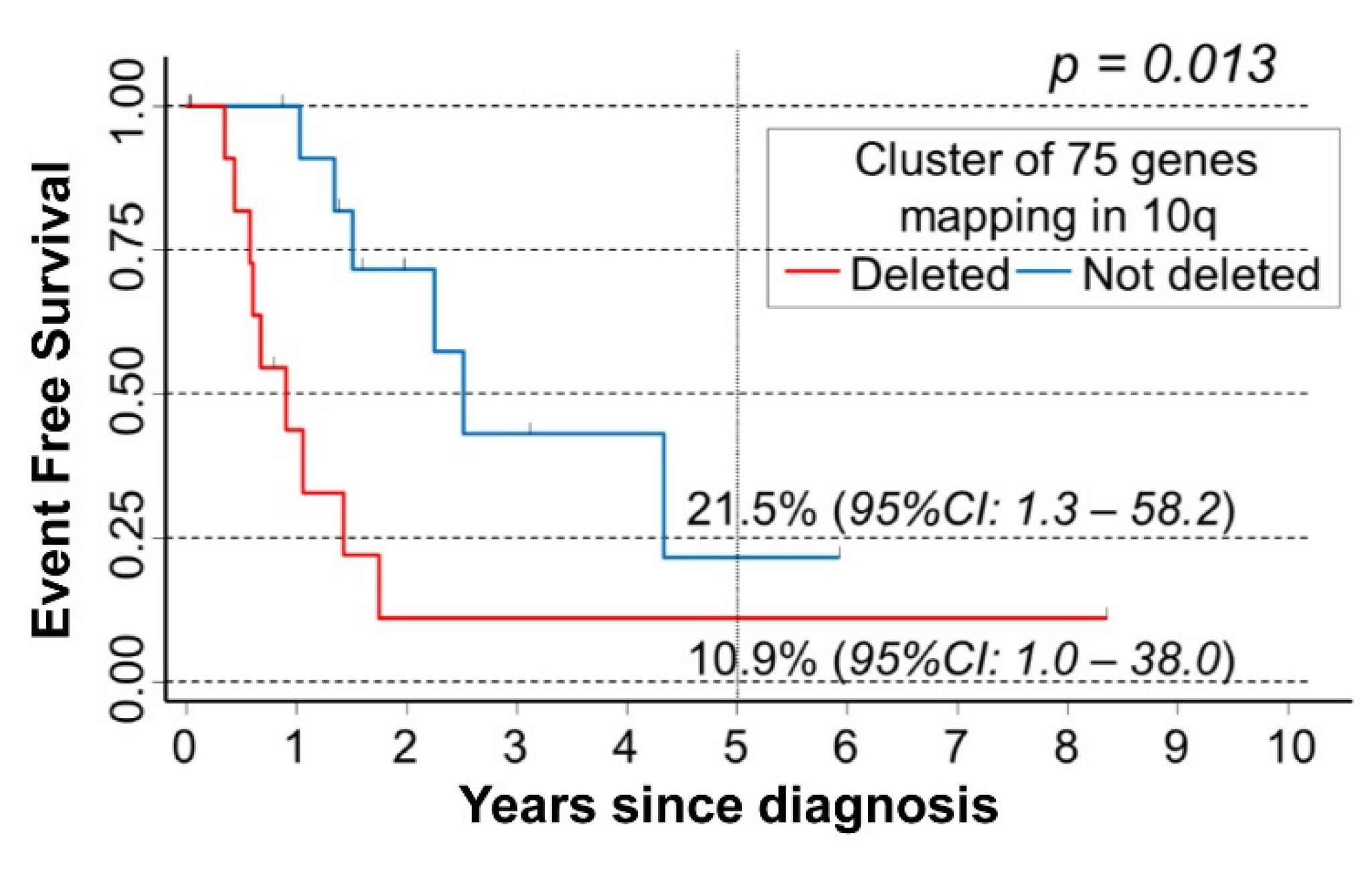
3.3. Survival Analysis of NB Patients by the Occurrence of Chromosome 10q Loss
3.4. Deletion Map of Chromosome 10q Loss Associated with NB Patients’ Survival
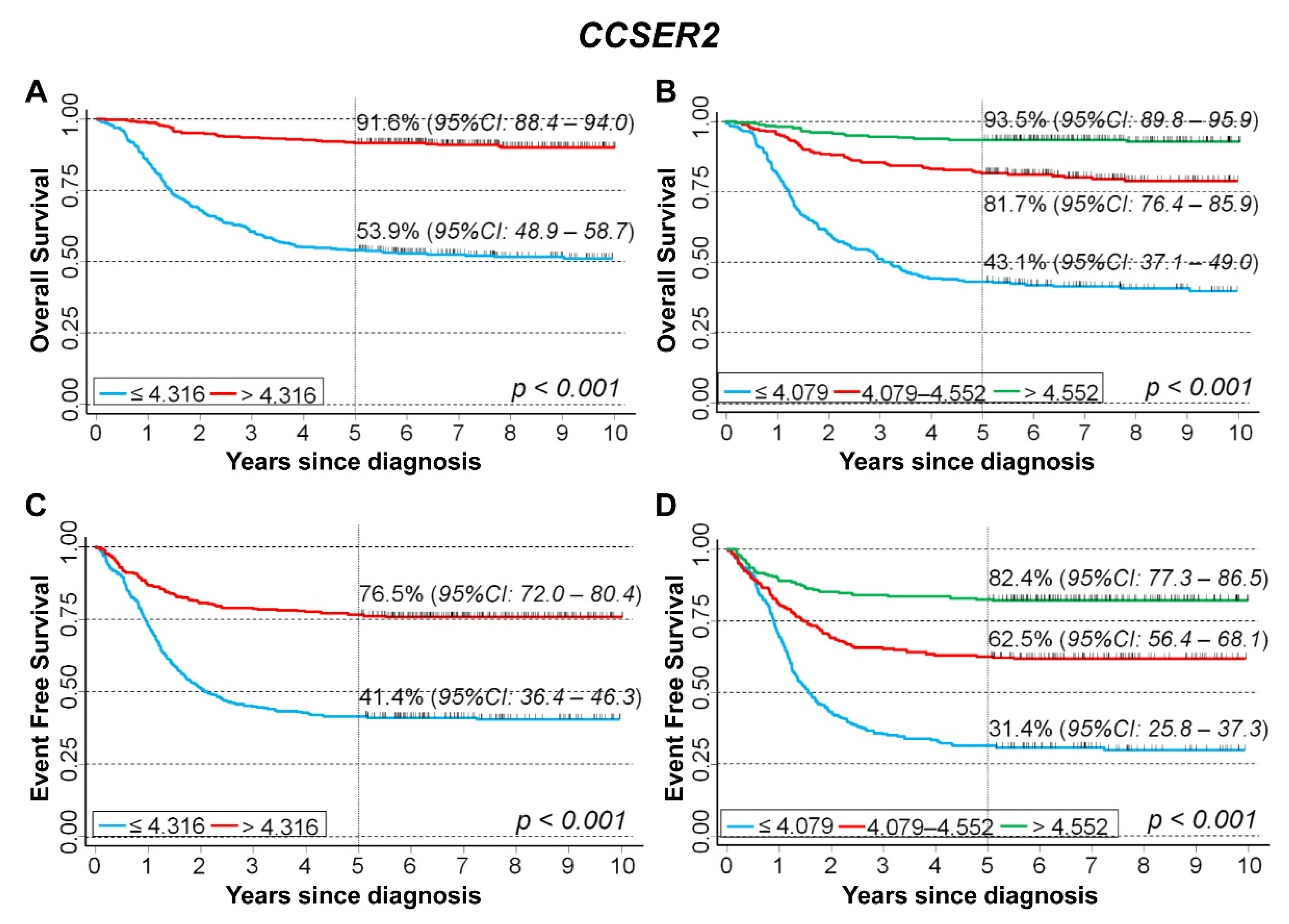
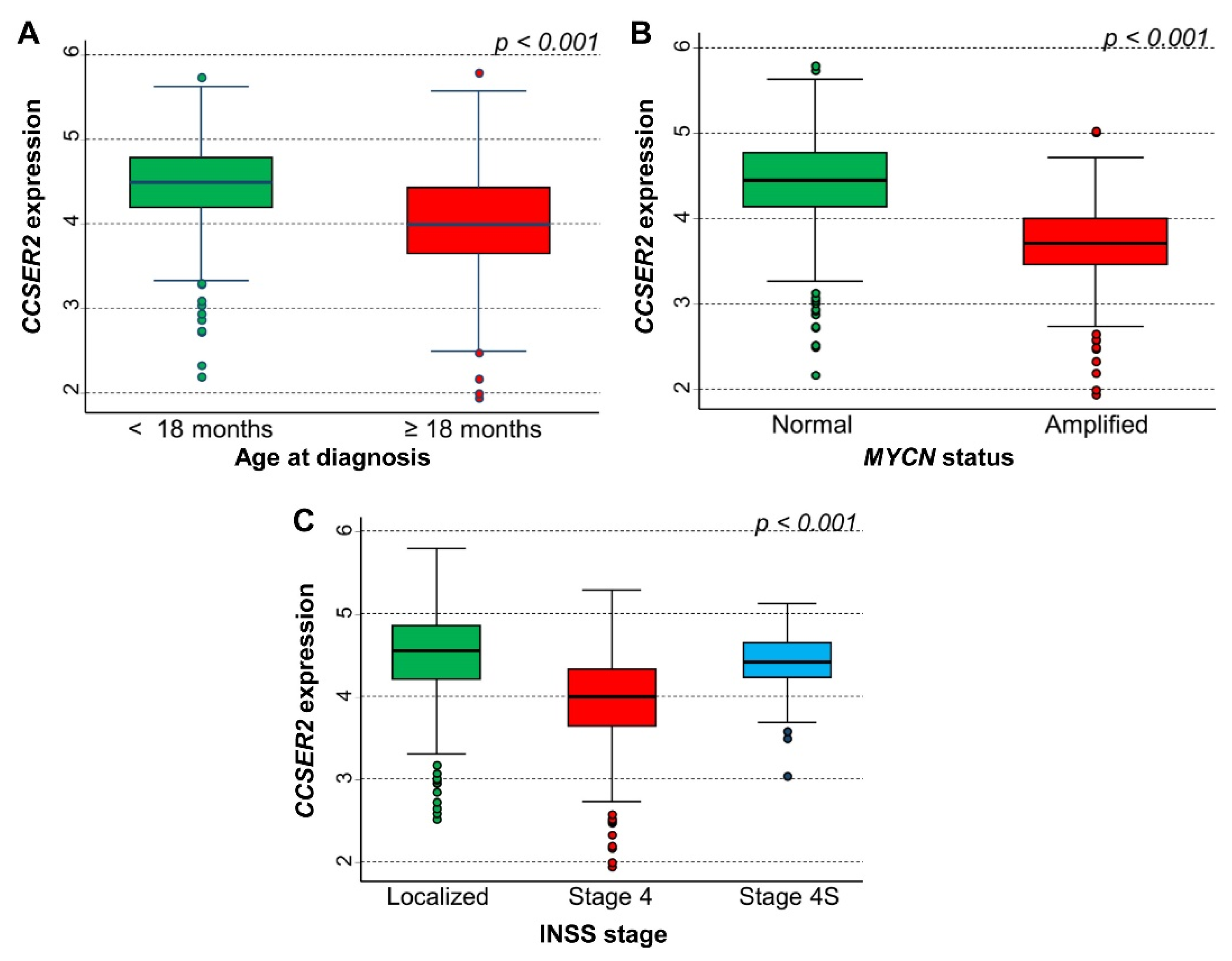
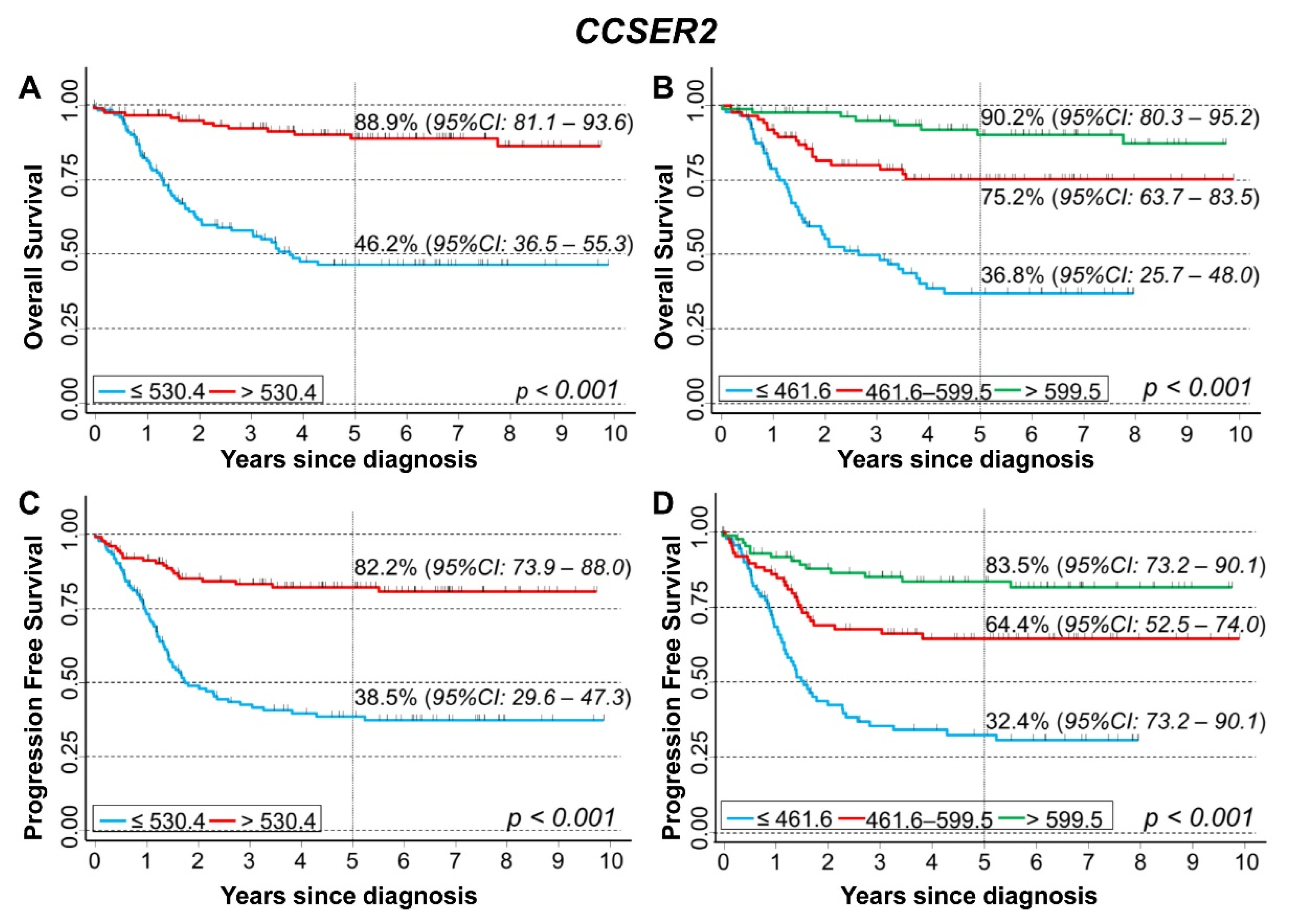
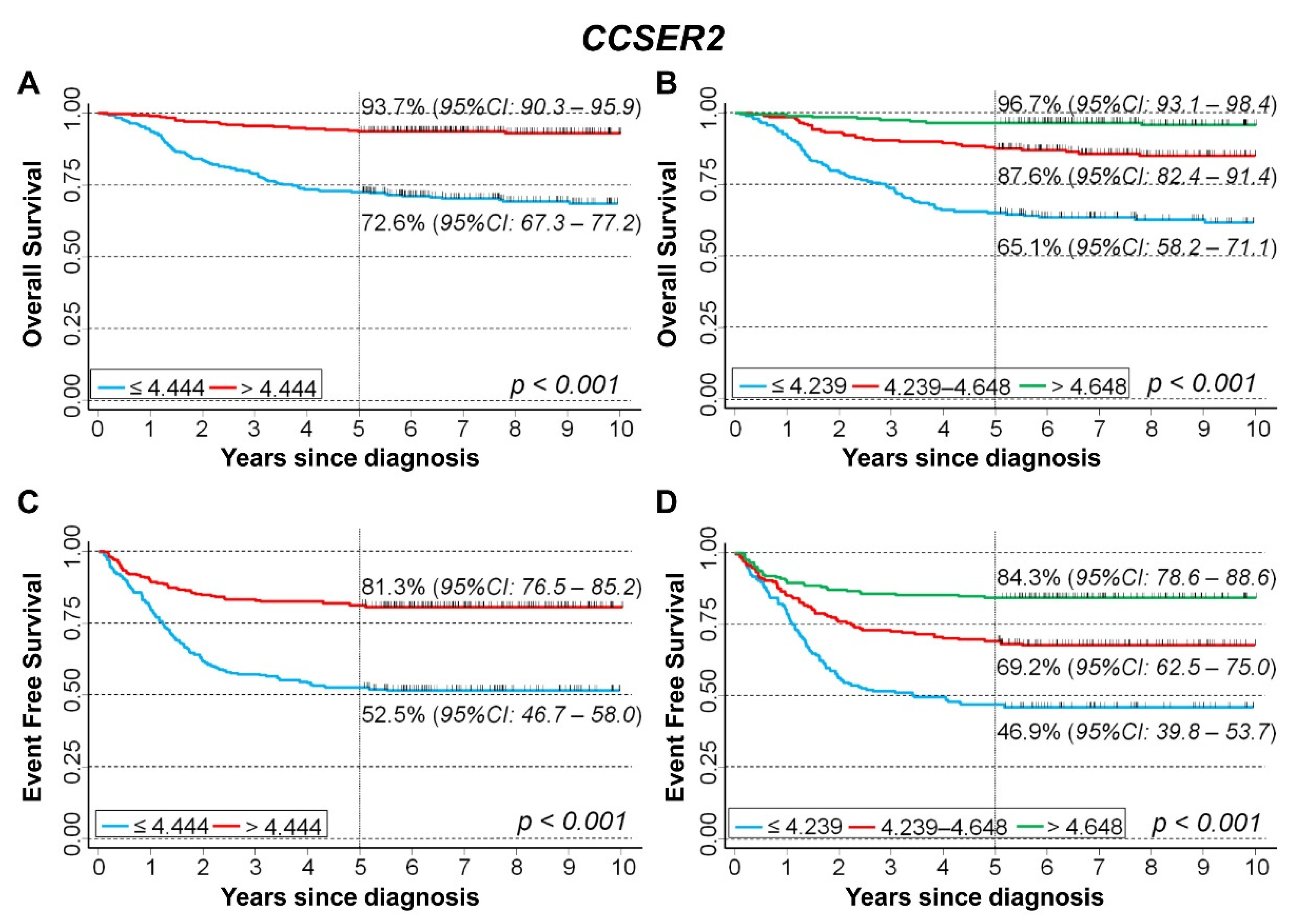
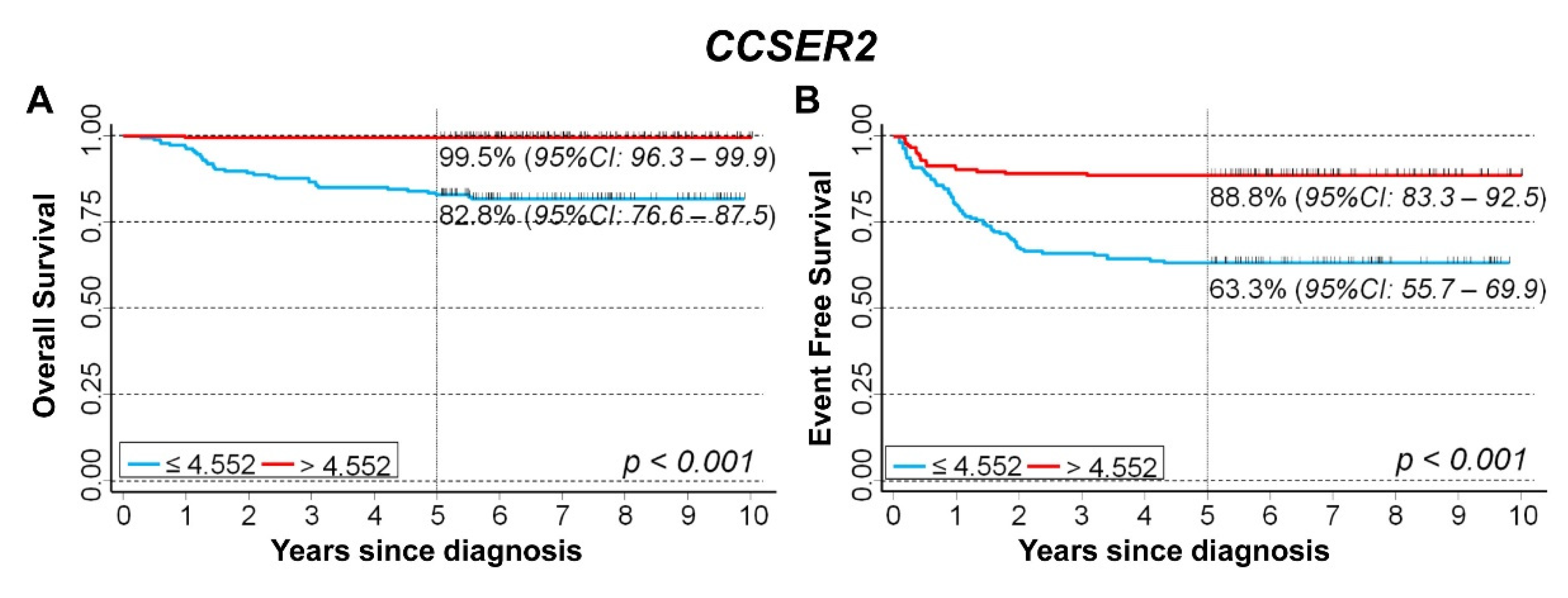
3.5. In Silico Analysis of the Association between the Expression of 75 Genes Identified in the Cluster Inside the 10q Loss Region and Survival of NB Patients
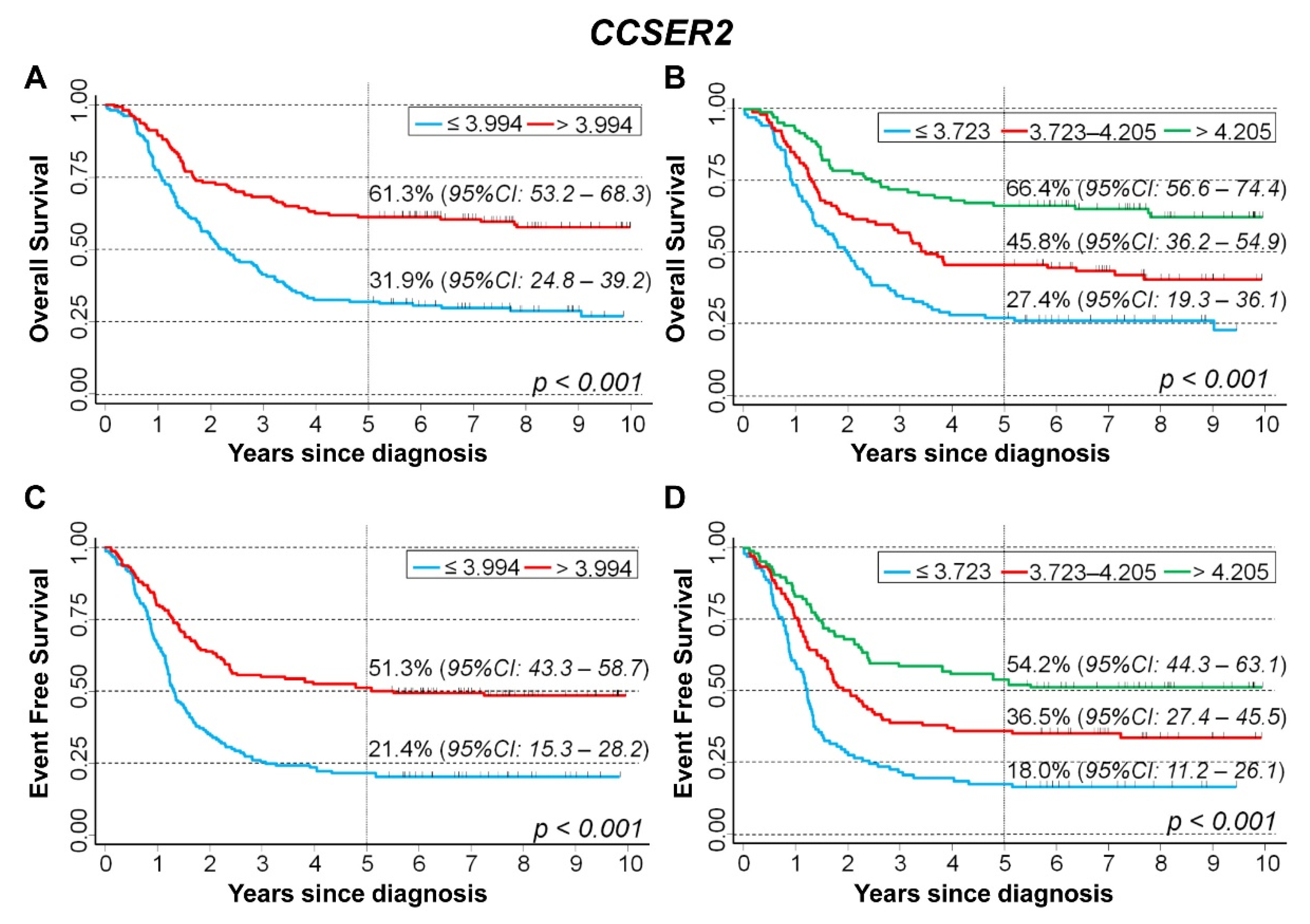
CCSER2
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Johnsen, J.I.; Dyberg, C.; Wickström, M. Neuroblastoma-A Neural Crest Derived Embryonal Malignancy. Front. Mol. Neurosci. 2019, 12, 9. [Google Scholar] [CrossRef] [PubMed]
- Newman, E.A.; Abdessalam, S.; Aldrink, J.H.; Austin, M.; Heaton, T.E.; Bruny, J.; Ehrlich, P.; Dasgupta, R.; Baertschiger, R.M.; Lautz, T.B.; et al. APSA Cancer committee. Update on neuroblastoma. J. Pediatr. Surg. 2019, 54, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Tomolonis, J.A.; Agarwal, S.; Shohet, J.M. Neuroblastoma pathogenesis: Deregulation of embryonic neural crest development. Cell Tissue Res. 2018, 372, 245–262. [Google Scholar] [CrossRef] [Green Version]
- Cheung, N.K.; Dyer, M.A. Neuroblastoma: Developmental biology, cancer genomics and immunotherapy. Nat. Rev. Cancer 2013, 13, 397–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matthay, K.K.; Maris, J.M.; Schleiermacher, G.; Nakagawara, A.; Mackall, C.L.; Diller, L.; Weiss, W.A. Neuroblastoma. Nat. Rev. Dis. Primers 2016, 2, 16078. [Google Scholar] [CrossRef]
- Louis, C.U.; Shohet, J.M. Neuroblastoma: Molecular pathogenesis and therapy. Ann. Rev. Med. 2015, 66, 49–63. [Google Scholar] [CrossRef] [Green Version]
- Brady, S.W.; Liu, Y.; Xiaotu Ma, X.; Gout, A.M.; Hagiwara, K.; Zhou, X.; Wang, J.; Macias, M.; Chen, X.; Easton, J.; et al. Pan-neuroblastoma analysis reveals age- and signature-associated driver alterations. Nat. Commun. 2020, 11, 5183. [Google Scholar] [CrossRef]
- Pugh, T.J.; Morozova, O.; Attiyeh, E.F.; Asgharzadeh, S.; Wei, J.S.; Auclair, D.; Carter, S.L.; Cibulskis, K.; Hanna, M.; Kiezun, A.; et al. The genetic landscape of high-risk neuroblastoma. Nat. Genet. 2013, 45, 279–284. [Google Scholar] [CrossRef] [Green Version]
- Newman, E.A.; Nuchtern, J.G. Recent biologic and genetic advances in neuroblastoma: Implications for diagnostic, risk stratification, and treatment strategies. Semin. Pediatr. Surg. 2016, 25, 257–264. [Google Scholar] [CrossRef]
- Bosse, K.R.; Maris, J.M. Advances in the translational genomics of neuroblastoma: From improving risk stratification and revealing novel biology to identifying actionable genomic alterations. Cancer 2016, 122, 20–33. [Google Scholar] [CrossRef] [Green Version]
- Cheung, N.K.; Zhang, J.; Lu, C.; Parker, M.; Bahrami, A.; Tickoo, S.K.; Heguy, A.; Pappo, A.S.; Federico, S.; Dalton, J.; et al. Association of age at diagnosis and genetic mutations in patients with neuroblastoma. J. Am. Med. Assoc. 2012, 307, 1062–1071. [Google Scholar] [CrossRef] [Green Version]
- Janoueix-Lerosey, I.; Schleiermacher, G.; Michels, E.; Mosseri, V.; Ribeiro, A.; Lequin, D.; Vermeulen, J.; Couturier, J.; Peuchmaur, M.; Valent, A.; et al. Overall genomic pattern is a predictor of outcome in neuroblastoma. J. Clin. Oncol. 2009, 27, 1026–1033. [Google Scholar] [CrossRef]
- Depuydt, P.; Boeva, V.; Hocking, T.D.; Cannoodt, R.; Ambros, I.M.; Ambros, P.F.; Asgharzadeh, S.; Attiyeh, E.F.; Combaret, V.; Defferrari, R.; et al. Genomic Amplifications and Distal 6q Loss: Novel Markers for Poor Survival in High-risk Neuroblastoma Patients. J. Natl. Cancer Inst. 2018, 110, 1084–1093. [Google Scholar] [CrossRef]
- Amoroso, L.; Ognibene, M.; Morini, M.; Conte, M.; Di Cataldo, A.; Tondo, A.; D’Angelo, P.; Castellano, A.; Garaventa, A.; Lasorsa, V.A.; et al. Genomic co-amplification of CDK4/MDM2/FRS2 is associated with very poor prognosis and atypical clinical features in neuroblastoma patients. Genes Chromosomes Cancer 2020, 59, 277–285. [Google Scholar] [CrossRef]
- Ognibene, M.; Amoroso, L.; Melchionda, F.; Cangelosi, D.; Zara, F.; Parodi, S.; Pezzolo, A. High Grade of Amplification of Six Regions on Chromosome 2p in a Neuroblastoma Patient with Very Poor Outcome: The Putative New Oncogene TSSC1. Cancers 2021, 13, 5792. [Google Scholar] [CrossRef]
- Ognibene, M.; Pezzolo, A. Ezrin interacts with the tumor suppressor CHL1 and promotes neuronal differentiation of human neuroblastoma. PLoS ONE 2020, 15, e0244069. [Google Scholar] [CrossRef]
- Ognibene, M.; Morini, M.; Garaventa, A.; Podestà, M.; Pezzolo, A. Identification of a minimal region of loss on chromosome 6q27 associated with poor survival of high-risk neuroblastoma patients. Cancer Biol. Ther. 2020, 21, 391–399. [Google Scholar] [CrossRef]
- Ognibene, M.; Pagnan, G.; Marimpietri, D.; Cangelosi, D.; Cilli, M.; Benedetti, M.C.; Boldrini, R.; Garaventa, A.; Frassoni, F.; Eva, A.; et al. CHL1 gene acts as a tumor suppressor in human neuroblastoma. Oncotarget 2018, 9, 25903–25921. [Google Scholar] [CrossRef] [Green Version]
- Lasorsa, V.A.; Cimmino, F.; Ognibene, M.; Mazzocco, K.; Erminio, G.; Morini, M.; Conte, M.; Iolascon, A.; Pezzolo, A.; Capasso, M. 19p loss is significantly enriched in older age neuroblastoma patients and correlates with poor prognosis. NPJ Genom Med. 2020, 5, 18. [Google Scholar] [CrossRef] [Green Version]
- Parodi, S.; Pistorio, A.; Erminio, G.; Ognibene, M.; Morini, M.; Garaventa, A.; Gigliotti, A.R.; Haupt, R.; Frassoni, F.; Pezzolo, A. Loss of whole chromosome X predicts prognosis of neuroblastoma patients with numerical genomic profile. Pediatr Blood Cancer. 2019, 66, e27635. [Google Scholar] [CrossRef]
- Brandetti, E.; Focaccetti, C.; Pezzolo, A.; Ognibene, M.; Folgiero, V.; Cotugno, N.; Benvenuto, M.; Palma, P.; Manzari, V.; Rossi, P.; et al. Enhancement of Neuroblastoma NK-Cell-Mediated Lysis through NF-kB p65 Subunit-Induced Expression of FAS and PVR, the Loss of Which Is Associated with Poor Patient Outcome. Cancers 2021, 13, 4368. [Google Scholar] [CrossRef] [PubMed]
- Altura, R.A.; Maris, J.M.; Li, H.; Boyett, J.M.; Brodeur, G.M.; Look, A.T. Novel regions of chromosomal loss in familial neuroblastoma by comparative genomic hybridization. Genes Chromosom. Cancer 1997, 19, 176–184. [Google Scholar] [CrossRef]
- Takita, J.; Hayashi, Y.; Kohno, T.; Shiseki, M.; Yamaguchi, N.; Hanada, R.; Yamamoto, K.; Yokota, J. Allelotype of neuroblastoma. Oncogene 1995, 11, 1829–1834. [Google Scholar] [PubMed]
- Kaneko, Y.; Kanda, N.; Maseki, N.; Sakurai, M.; Tsuchida, Y.; Takeda, T.; Okabe, I.; Sakurai, M. Different karyotypic patterns in early and advanced stage neuroblastomas. Cancer Res. 1987, 47, 311–318. [Google Scholar]
- Cohn, S.L.; Pearson, A.D.; London, W.B.; Monclair, T.; Ambros, P.F.; Brodeur, G.M.; Faldum, A.; Hero, B.; Iehara, T.; Machin, D.; et al. INRG Task Force. The International Neuroblastoma Risk Group (INRG) classification system: An INRG Task Force report. J. Clin. Oncol. 2009, 27, 289–297. [Google Scholar] [CrossRef] [Green Version]
- Park, J.R.; Bagatell, R.; Cohn, S.L.; Pearson, A.D.; Villablanca, J.G.; Berthold, F.; Burchill, S.; Boubaker, A.; McHugh, K.; Nuchtern, J.G.; et al. Revisions to the International neuroblastoma response criteria: A consensus statement from the National Cancer Institute clinical trials planning meeting. J. Clin. Oncol. 2017, 35, 2580–2587. [Google Scholar] [CrossRef]
- Haupt, R.; Garaventa, A.; Gambini, C.; Parodi, S.; Cangemi, G.; Casale, F.; Viscardi, E.; Bianchi, M.; Prete, A.; Jenkner, A.; et al. Improved survival of children with neuroblastoma between 1979 and 2005: A report of the Italian Neuroblastoma Registry. J. Clin. Oncol. 2010, 28, 2331–2338. [Google Scholar] [CrossRef]
- Cangelosi, D.; Morini, M.; Zanardi, N.; Sementa, A.R.; Muselli, M.; Conte, M.; Garaventa, A.; Pfeffer, U.; Bosco, M.C.; Varesio, L.; et al. Hypoxia Predicts Poor Prognosis in Neuroblastoma Patients and Associates with Biological Mechanisms Involved in Telomerase Activation and Tumor Microenvironment Reprogramming. Cancers 2020, 12, 2343. [Google Scholar] [CrossRef]
- Zhang, W.; Yu, Y.; Hertwig, F.; Thierry-Mieg, J.; Zhang, W.; Thierry-Mieg, D.; Wang, J.; Furlanello, C.; Devanarayan, V.; Cheng, J.; et al. Comparison of RNA-seq and microarray-based models for clinical endpoint prediction. Genome Biol. 2015, 16, 133. [Google Scholar] [CrossRef] [Green Version]
- Oberthuer, A.; Juraeva, D.; Hero, B.; Volland, R.; Sterz, C.; Schmidt, R.; Faldum, A.; Kahlert, Y.; Engesser, A.; Asgharzadeh, S.; et al. Revised risk estimation and treatment stratification of low- and intermediate-risk neuroblastoma patients by integrating clinical and molecular prognostic markers. Clin. Cancer Res. 2015, 21, 1904–1915. [Google Scholar] [CrossRef] [Green Version]
- Rajbhandari, P.; Lopez, G.; Capdevila, C.; Salvatori, B.; Yu, J.; Rodriguez-Barrueco, R.; Martinez, D.; Yarmarkovich, M.; Weichert-Leahey, N.; Abraham, B.J.; et al. Cross-cohort analysis identifies a TEAD4-MYCN positive feedback loop as the core regulatory element of high-risk neuroblastoma. Cancer Discov. 2018, 8, 582–599. [Google Scholar] [CrossRef] [Green Version]
- Gibbons, J.D.; Chakraborti, S. Nonparametric Statistical Inference, 5th ed.; CRC Press, Chapman & Hall: New York, NY, USA, 2010; pp. 1–650. [Google Scholar]
- Hosmer, D.W.; Lemeshow, S.; May, S. Applied Survival Analysis—Regression Modelling of Time to Event Data, 2nd ed.; John Wiley & Sons, Inc.: New York, NY, USA, 2008; pp. 1–416. [Google Scholar]
- Vandesompele, J.; Baudis, M.; De Preter, K.; Van Roy, N.; Ambros, P.; Bown, N.; Brinkschmidt, C.; Christiansen, H.; Combaret, V.; Lastowska, M.; et al. Unequivocal delineation of clinicogenetic subgroups and development of a new model for improved outcome prediction in neuroblastoma. J. Clin. Oncol. 2005, 23, 2280–2299. [Google Scholar] [CrossRef] [Green Version]
- Mikhail, F.M.; Biegel, J.A.; Cooley, L.D.; Dubuc, A.M.; Hirsch, B.; Horner, V.L.; Newman, S.; Shao, L.; Wolff, D.J.; Raca, G. Technical laboratory standards for interpretation and reporting of acquired copy-number abnormalities and copy-neutral loss of heterozygosity in neoplastic disorders: A joint consensus recommendation from the American College of Medical Genetics and Genomics (ACMG) and the Cancer Genomics Consortium (CGC). Genet Med. 2019, 21, 1903–1916. [Google Scholar]
- Lastowska, M.; Viprey, V.; Santibanez-Koref, M.; Wappler, I.; Peters, H.; Cullinane, C.; Roberts, P.; Hall, A.G.; Tweddle, D.A.; Pearson, A.D.; et al. Identification of candidate genes involved in neuroblastoma progression by combining genomic and expression microarrays with survival data. Oncogene 2007, 26, 7432–7444. [Google Scholar] [CrossRef] [Green Version]
- Brandon, N.J.; Sawa, A. Linking neurodevelopmental and synaptic theories of mental illness through DISC1. Nat. Rev. Neurosci. 2011, 12, 707–722. [Google Scholar] [CrossRef]
- Ishizuka, K.; Kamiya, A.; Oh, E.C.; Kanki, H.; Seshadri, S.; Robinson, J.F.; Murdoch, H.; Dunlop, A.J.; Kubo, K.I.; Furukori, K.; et al. DISC1-dependent switch from progenitor proliferation to migration in the developing cortex. Nature 2011, 473, 92–96. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Yan, Q.; Zhou, P.; Li, S.; Zhu, F. HERV-W env regulates calcium influx via activating TRPC3 channel together with depressing DISC1 in human neuroblastoma cells. J. Neurovirol. 2019, 25, 101–113. [Google Scholar] [CrossRef]
- You, Y.; Peng, B.; Ben, S.; Hou, W.; Sun, L.; Jiang, W. Lead Neurotoxicity on Human Neuroblastoma Cell Line SH-SY5Y is Mediated via Transcription Factor EGR1/Zif268 Induced Disrupted in Scherophernia-1 Activation. Neurochem. Res. 2018, 43, 1308–1316. [Google Scholar] [CrossRef]
- Mun, D.J.; Goo, B.S.; Suh, B.K.; Hong, J.H.; Woo, Y.; Kim, S.J.; Kim, S.; Lee, S.B.; Won, Y.; Yoo, J.Y.; et al. Gcap14 is a microtubule plus-end-tracking protein coordinating microtubule-actin crosstalk during neurodevelopment. Proc. Natl. Acad. Sci. USA 2023, 120, e2214507120. [Google Scholar] [CrossRef]
- Huang, H.; Yao, H.; Wei, Y.; Chen, M.; Sun, J. Cellular senescence-related long noncoding ribonucleic acids: Predicting prognosis in hepatocellular carcinoma. Cancer Rep. 2023, 1, e1791. [Google Scholar] [CrossRef]
- Lee, J.T. Epigenetic Regulation by Long Noncoding RNAs. Science 2012, 338, 1435–1439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Min, L.; Wang, H.; Qi, H. Astragaloside IV inhibits the progression of liver cancer by modulating macrophage polarization through the TLR4/NF-κB/STAT3 signaling pathway. Am. J. Transl. Res. 2022, 14, 1551–1566. [Google Scholar] [PubMed]
- Xu, Y.; Tian, H.; Luan, C.G.; Sun, K.; Bao, P.J.; Zhang, H.Y.; Zhang, N. Telocytes promote hepatocellular carcinoma by activating the ERK signaling pathway and miR-942-3p/MMP9 axis. Cell Death Discov. 2021, 7, 209. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Deng, Q.; Ni, T.; Liu, Y.; Lu, L.; Dai, H.; Wang, H.; Yang, W. Targeted Inhibition of LPL/FABP4/CPT1 fatty acid metabolic axis can effectively prevent the progression of nonalcoholic steatohepatitis to liver cancer. Int. J. Biol. Sci. 2021, 17, 4207–4222. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Liu, S.; Xu, F.; Kong, Y.; Wan, L.; Zhang, Y.; Zhang, Z. Inhibition of Proteasome Activity Induces Aggregation of IFIT2 in the Centrosome and Enhances IFIT2-Induced Cell Apoptosis. Int. J. Biol. Sci. 2017, 13, 383–390. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Michal, J.J.; Zhang, L.; Ding, B.; Lunney, J.K.; Liu, B.; Jiang, Z. Interferon induced IFIT family genes in host antiviral defense. Int. J. Biol. Sci. 2013, 9, 200–208. [Google Scholar] [CrossRef] [Green Version]
- Lai, K.C.; Liu, C.J.; Lin, T.J.; Mar, A.C.; Wang, H.H.; Chen, C.W.; Hong, Z.X.; Lee, T.C. Blocking TNF-α inhibits angiogenesis and growth of IFIT2-depleted metastatic oral squamous cell carcinoma cells. Cancer Lett. 2016, 370, 207–215. [Google Scholar] [CrossRef]
- Danish, H.H.; Goyal, S.; Taunk, N.K.; Wu, H.; Moran, M.S.; Haffty, B.G. Interferon-induced protein with tetratricopeptide repeats 1 (IFIT1) as a prognostic marker for local control in T1-2 N0 breast cancer treated with breast-conserving surgery and radiation therapy (BCS + RT). Breast J. 2013, 19, 231–239. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Kong, Y.; Liu, S.; Zeng, L.; Wan, L.; Zhang, Z. Curcumin induces apoptosis in human leukemic cell lines through an IFIT2-dependent pathway. Cancer Biol. Ther. 2017, 18, 43–50. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.F.; Chen, Y.; Lin, G.S.; Zhang, J.D.; Tang, W.L.; Huang, J.H.; Chen, J.S.; Wang, X.F.; Lin, Z.X. High IFIT1 expression predicts improved clinical outcome, and IFIT1 along with MGMT more accurately predicts prognosis in newly diagnosed glioblastoma. Hum. Pathol. 2016, 52, 136–144. [Google Scholar] [CrossRef]
- Yang, Y.; Zhou, Y.; Hou, J.; Bai, C.; Li, Z.; Fan, J.; Ng, I.O.L.; Zhou, W.; Sun, H.; Dong, Q.; et al. Hepatic IFIT3 predicts interferon-α therapeutic response in patients of hepatocellular carcinoma. Hepatology. 2017, 66, 152–166. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Altendorf-Hofmann, A.; Pozios, I.; Camaj, P.; Däberitz, T.; Wang, X.; Niess, H.; Seeliger, H.; Popp, F.; Betzler, C.; et al. Elevated interferon-induced protein with tetratricopeptide repeats 3 (IFIT3) is a poor prognostic marker in pancreatic ductal adenocarcinoma. J. Cancer Res. Clin. Oncol. 2017, 143, 1061–1068. [Google Scholar] [CrossRef]
- Pidugu, V.K.; Pidugu, H.B.; Wu, M.M.; Liu, C.J.; Lee, T.C. Emerging Functions of Human IFIT Proteins in Cancer. Front. Mol. Biosci. 2019, 19, 148. [Google Scholar] [CrossRef] [Green Version]
- Jung, S.H.; Lee, H.C.; Yu, D.M.; Kim, B.C.; Park, S.M.; Lee, Y.S.; Park, H.J.; Ko, Y.G.; Lee, J.S. Heparan sulfation is essential for the prevention of cellular senescence. Cell Death Differ. 2016, 23, 417–429. [Google Scholar] [CrossRef] [Green Version]
- Di Croce, L.; Helin, K. Transcriptional regulation by Polycomb group proteins. Nat. Struct. Mol. Biol. 2013, 20, 1147–1155. [Google Scholar] [CrossRef]
- Yao, M.; Zhou, X.; Zhou, J.; Gong, S.; Hu, G.; Li, J.; Huang, K.; Lai, P.; Shi, G.; Hutchins, A.P.; et al. PCGF5 is required for neural differentiation of embryonic stem cells. Nat. Commun. 2018, 9, 1463. [Google Scholar] [CrossRef] [Green Version]
- Hsu, C.L.; Chang, H.Y.; Chang, J.Y.; Hsu, W.M.; Huang, H.C.; Juan, H.F. Unveiling MYCN regulatory networks in neuroblastoma via integrative analysis of heterogeneous genomics data. Oncotarget 2016, 7, 36293–36310. [Google Scholar] [CrossRef] [Green Version]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Case N° | Age at Onset (Months) | INSS Stage | INRG Stage | MYCN Status | Cytoband and Chromosomal Coordinates of 10q Loss | Relapse | Follow-Up | Disease State |
|---|---|---|---|---|---|---|---|---|
| 1 | 47 | 4 | M | gain | 10q11.23–q21.3; chr10:51676176–69078545 | yes | dead | |
| 2 | 68 | 4 | M | gain | 10q24.1–q26.3; chr10: 98064330–135404523 | yes | dead | |
| 3 | 40 | 3 | L2 | amplified | 10q11.23–q26.3; chr10: 50955699–135372492 | no | alive | CR |
| 4 | 46 | 4 | M | single copy | 10q11.21–21.1; chr10: 42969765–57723272 | yes | alive | AD |
| 5 | 55 | 4 | M | amplified | 10q23.32–q26.3; chr10: 93204607–135404523 | no | alive | CR |
| 6 | 5 | 1 | L1 | amplified | 10q11.22–q26.3; chr10: 48334407–135404523 | yes | dead | |
| 7 | 10 | 3 | L2 | amplified | 10q21.3–q26.3; chr10: 64979620–135404523 | no | dead | |
| 8 | 14 | 1 | L1 | amplified | 10q22.1–q26.3; chr10: 73406500–135404523 | yes | dead | |
| 9 | 200 | 2A | L1 | single copy | 10q11.21–q24.32; chr10: 42969765–103988947 | yes | alive | CR |
| 10 | 45 | 4 | M | gain | 10q21.1; chr10: 55252216–56505255 | yes | dead | |
| 11 | 37 | 1 | L1 | single copy | 10q22.3–q26.11; chr10: 78015106–121458431 | yes | dead | |
| 12 | 53 | 2B | L1 | gain | CN-LOH 10q11.23–q26.3; chr10: 4541520–67615559 | yes | alive | CR |
| 13 | 52 | 4 | M | gain | 10q11.22; chr10: 46938469–48317747 | yes | alive | AD |
| 14 | 22 | 4 | M | amplified | 10q22.3–q26.3; chr10: 81234748–135404523 | yes | dead | |
| 15 | 33 | 3 | L2 | amplified | 10q21.1–q26.3; chr10: 57654752–135411735 | no | alive | AD |
| 16 | 1 | 4S | Ms | gain | CN-LOH 10q21.1–21.2; chr10: 58949883–63882016 | no | alive | CR |
| 17 | 2 | 1 | L1 | gain | 10q21.1; chr10: 56329864–58030413 | na | na | |
| 18 | 12 | 3 | L2 | gain | 10q26.3; chr10:133245712–135405996 | no | dead | |
| 19 | 10 | 4 | M | single copy | 10q26.3; chr10:133079678–134996216 | no | alive | CR |
| 20 | 31 | 4 | M | single copy | 10q21.3; 10q26.13–q26.3; chr10:68757961–69106575 chr10:123773590–135421826 | no | alive | AD |
| 21 | 30 | 4 | M | amplified | 10q11.21–q26.3; chr10:45247685–135372492 | yes | alive | AD |
| 22 | 20 | 4 | M | amplified | 10q25.3–q26.3; chr10:116405945–135434178 | no | alive | na |
| 23 | 20 | 3 | L2 | single copy | 10q22.2–q24.33; chr10:75923421–105458525 | no | alive | AD |
| 24 | 59 | 4 | M | single copy | 10q26.13–q26.3; chr10:123773590–135377532 | na | na | |
| 25 | 52 | 4 | M | single copy | 10q11.22–q22.3; chr10:49797866–77817731 | no | alive | CR |
| 26 | 46 | 4 | M | amplified | 10q24.1–q26.3; chr10: 98515392–135404523 | yes | alive | AD |
| NB Cell Line | Chromosome 10q Status | Chromosomal Coordinates of 10q Loss | MYCN (2p24.3) |
|---|---|---|---|
| SK-N-AS | 10q11.21–q26.3 loss | Chr10: 43615122–135474787 | single copy |
| SK-N-BE(2)c | 10q11.21–q26.3 loss | Chr10: 42418957–135434178 | amplification |
| LA-N-1 | 10q11.21–q26.3 loss | Chr10: 42976950–135234843 | amplification |
| SMS-KCNR | 10q11.22 loss | Chr10: 46938469–49262406 | single copy |
| SH-EP | 10q23.31–q24.32 gain | Chr10:90628315–103956178 | gain |
| GICAN | normal | - | single copy |
| ACN | normal | - | single copy |
| SH-SY5Y | normal | - | gain |
| SK-N-SH | normal | - | amplification |
| LA-N-5 | normal | - | amplification |
| IMR32 | normal | - | amplification |
| Tet-21/N | normal | - | single copy |
| Other SCA * | Loss 10q | ||||
|---|---|---|---|---|---|
| Patients’ Characteristics | N | % | N | % | p |
| Age (months) | 0.369 | ||||
| 0–17 | 91 | 38.9 | 7 | 26.9 | |
| 18–59 | 108 | 46.2 | 16 | 61.5 | |
| ≥60 | 35 | 15.0 | 3 | 11.5 | |
| Gender | 0.619 | ||||
| Male | 123 | 52.6 | 15 | 57.7 | |
| Female | 111 | 47.4 | 11 | 42.3 | |
| INSS Stage | 0.999 | ||||
| 1–2 | 55 | 23.6 | 6 | 24.0 | |
| 3 | 42 | 18.0 | 5 | 20.0 | |
| 4 | 119 | 51.1 | 13 | 52.0 | |
| 4S | 17 | 7.3 | 1 | 4.0 | |
| MYCN status | 0.345 | ||||
| Non-amplified | 165 | 70.5 | 16 | 61.5 | |
| Amplified | 69 | 29.5 | 10 | 38.5 | |
| Cangelosi et al. (N = 786) | E-MTAB-1781 (N = 709) | SEQC-498 (N = 498) | NRC-283 (N = 283) | |||||
|---|---|---|---|---|---|---|---|---|
| Patient Characteristics | N | % | N | % | N | % | N | % |
| Age at diagnosis | ||||||||
| <18 months | 449 | 57.1 | 431 | 60.8 | 305 | 61.2 | 145 | 51.2 |
| ≥18 months | 337 | 42.9 | 278 | 39.2 | 198 | 38.8 | 133 | 47.0 |
| Missing | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 5 | 1.8 |
| INSS Stage | ||||||||
| 1 | 143 | 18.2 | 159 | 22.4 | 121 | 24.3 | 50 | 17.7 |
| 2 | 125 | 15.9 | 118 | 16.6 | 78 | 15.7 | 36 | 12.7 |
| 3 | 105 | 13.4 | 93 | 13.1 | 63 | 12.7 | 43 | 15.2 |
| 4 | 320 | 40.7 | 259 | 36.5 | 183 | 36.8 | 124 | 43.8 |
| 4s | 92 | 11.7 | 80 | 11.3 | 53 | 10.6 | 27 | 9.5 |
| Missing | 1 | 0.1 | 0 | 0.0 | 0 | 0.0 | 3 | 1.1 |
| MYCN status | ||||||||
| Not amplified | 629 | 80.0 | 581 | 82.0 | 401 | 80.5 | 222 | 78.4 |
| Amplified | 153 | 19.5 | 122 | 17.2 | 92 | 18.5 | 55 | 19.4 |
| Missing | 4 | 0.5 | 6 | 0.8 | 5 | 1.0 | 6 | 2.2 |
| Events * | 320 | 40.7 1 | 256 2 | 36.1 | 183 | 36.9 | 97 3 | 34.3 |
| Deaths | 229 | 29.1 | 161 | 22.7 | 105 | 21.1 | 75 4 | 26.5 |
| Univariable Analysis | Multivariable Analysis | ||||||
|---|---|---|---|---|---|---|---|
| Gene Expression | N/O | HR | 95% CI | p | HR | 95% CI | p |
| Overall Survival | |||||||
| Median | <0.001 | <0.001 | |||||
| ≤4.316 (reference) | 393/192 | 1 | - | 1 | - | ||
| >4.316 | 393/37 | 0.14 | 0.10–0.21 | 0.37 | 0.25–0.55 | ||
| Tertiles | <0.001 | <0.001 | |||||
| ≤4.079 (reference) | 262/157 | 1 | - | 1 | - | ||
| 4.079–4.552 | 262/54 | 0.25 | 0.18–0.34 | 0.60 | 0.43–0.85 | ||
| >4.552 | 262/18 | 0.08 | 0.05–0.13 | 0.27 | 0.16–0.47 | ||
| Continuous variable | 786/229 | 0.28 | 0.23–0.33 | <0.001 | 0.56 | 0.45–0.69 | <0.001 |
| Event Free Survival | |||||||
| Median | <0.001 | <0.001 | |||||
| ≤4.316 (reference) | 377/225 | 1 | - | 1 | - | ||
| >4.316 | 392/95 | 0.32 | 0.25–0.40 | 0.55 | 0.41–0.72 | ||
| Tertiles | <0.001 | <0.001 | |||||
| ≤4.079 (reference) | 248/173 | 1 | - | 1 | - | ||
| 4.079–4.552 | 259/100 | 0.44 | 0.34–0.56 | 0.71 | 0.53–0.94 | ||
| >4.552 | 262/47 | 0.18 | 0.13–0.25 | 0.35 | 0.24–0.51 | ||
| Continuous variable | 769/320 | 0.35 | 0.30–0.41 | <0.001 | 0.55 | 0.45–0.68 | <0.001 |
| Univariable Analysis | Multivariable Analysis | ||||||
|---|---|---|---|---|---|---|---|
| Gene Expression | N/O | HR | 95% CI | p | HR | 95% CI | p |
| Overall Survival | |||||||
| Median | <0.001 | 0.003 | |||||
| ≤530.4 (reference) | 141/62 | 1 | - | 1 | - | ||
| >530.4 | 135/13 | 0.16 | 0.09–0.29 | 0.36 | 0.18–0.72 | ||
| Tertiles | <0.001 | 0.017 | |||||
| ≤461.6 (reference) | 94/48 | 1 | - | 1 | - | ||
| 461.6–599.5 | 92/19 | 0.30 | 0.18–0.52 | 0.68 | 0.37–1.3 | ||
| >599.5 | 90/8 | 0.11 | 0.05–0.23 | 0.36 | 0.15–0.85 | ||
| Continuous variable | 276/75 | 0.994 | 0.993–0.996 | <0.001 | 0.997 | 0.995–0.999 | 0.005 |
| Progression Free Survival | |||||||
| Median | <0.001 | <0.001 | |||||
| ≤530.4 (reference) | 140/75 | 1 | - | 1 | - | ||
| >530.4 | 135/22 | 0.22 | 0.14–0.37 | 0.38 | 0.22–0.66 | ||
| Tertiles | <0.001 | 0.008 | |||||
| ≤461.6 (reference) | 94/55 | 1 | - | 1 | - | ||
| 461.6–599.5 | 91/28 | 0.41 | 0.26–0.64 | 0.71 | 0.41–1.2 | ||
| >599.5 | 90/14 | 0.18 | 0.10–0.32 | 0.39 | 0.19–0.79 | ||
| Continuous variable | 265/97 | 0.996 | 0.995–0.997 | <0.001 | 0.998 | 0.997–0.999 | 0.014 |
| Univariable Analysis | Multivariable Analysis | ||||||
|---|---|---|---|---|---|---|---|
| Gene Expression | N/O | HR | 95% CI | p | HR | 95% CI | p |
| Overall Survival | |||||||
| Median | <0.001 | <0.001 | |||||
| ≤4.444 (reference) | 314/98 | 1 | - | 1 | - | ||
| >4.444 | 315/21 | 0.18 | 0.11–0.29 | 0.37 | 0.23–0.60 | ||
| Tertiles | <0.001 | <0.001 | |||||
| ≤4.239 (reference) | 209/80 | 1 | - | 1 | - | ||
| 4.239–4.648 | 210/31 | 0.32 | 0.22–0.50 | 0.66 | 0.43–1.0 | ||
| >4.648 | 210/8 | 0.08 | 0.04–0.17 | 0.20 | 0.10–0.43 | ||
| Continuous variable | 629/119 | 0.25 | 0.19–0.32 | <0.001 | 0.52 | 0.38–0.70 | <0.001 |
| Event Free Survival | |||||||
| Median | <0.001 | <0.001 | |||||
| ≤4.444 (reference) | 299/146 | 1 | - | 1 | - | ||
| >4.4444 | 315/61 | 0.33 | 0.25–0.47 | 0.44 | 0.32–0.61 | ||
| Tertiles | <0.001 | <0.001 | |||||
| ≤4.239 (reference) | 196/107 | 1 | - | 1 | - | ||
| 4.239–4.648 | 208/67 | 0.51 | 0.37–0.69 | 0.68 | 0.50–0.94 | ||
| >4.648 | 210/33 | 0.23 | 0.15–0.34 | 0.34 | 0.23–0.52 | ||
| Continuous variable | 614/207 | 0.28 | 0.22–0.37 | <0.001 | 0.44 | 0.33–0.59 | <0.001 |
| Univariable Analysis | Multivariable Analysis | ||||||
|---|---|---|---|---|---|---|---|
| Gene Expression | N/O | HR | 95% CI | p | HR | 95% CI | p |
| Overall Survival | |||||||
| Median | <0.001 | <0.001 | |||||
| ≤3.709 (reference) | 76/65 | 1 | - | 1 | - | ||
| >3.709 | 77/43 | 0.45 | 0.31–0.67 | 0.49 | 0.33–0.73 | ||
| Tertiles | <0.001 | <0.001 | |||||
| ≤3.574 (reference) | 51/44 | 1 | - | 1 | - | ||
| 3.574–3.936 | 51/39 | 0.69 | 0.45–1.1 | 0.69 | 0.45–1.1 | ||
| >3.936 | 51/25 | 0.35 | 0.21–0.57 | 0.38 | 0.23–0.63 | ||
| Continuous variable | 153/108 | 0.65 | 0.47–0.88 | 0.008 | 0.70 | 0.50–0.97 | 0.038 |
| Event Free Survival | |||||||
| Median | <0.001 | 0.001 | |||||
| ≤3.709 (reference) | 74/64 | 1 | - | 1 | - | ||
| >3.709 | 77/47 | 0.49 | 0.33–0.71 | 0.53 | 0.36–0.78 | ||
| Tertiles | <0.001 | <0.001 | |||||
| ≤3.574 (reference) | 49/43 | 1 | - | 1 | - | ||
| 3.574–3.936 | 51/40 | 0.69 | 0.45–1.1 | 0.69 | 0.45–1.1 | ||
| >3.936 | 51/28 | 0.39 | 0.24–0.63 | 0.42 | 0.26–0.68 | ||
| Continuous variable | 151/111 | 0.69 | 0.51–0.95 | 0.023 | 0.75 | 0.54–1.04 | 0.096 |
| Univariable Analysis | Multivariable Analysis | ||||||
|---|---|---|---|---|---|---|---|
| Gene Expression | N/O | HR | 95% CI | p | HR | 95% CI | p |
| Overall Survival | |||||||
| Median | <0.001 | 0.009 | |||||
| ≤3.994 (reference) | 160/116 | 1 | - | 1 | - | ||
| >3.994 | 160/67 | 0.43 | 0.32–0.58 | 0.65 | 0.47–0.90 | ||
| Tertiles | <0.001 | <0.001 | |||||
| ≤3.723 (reference) | 106/80 | 1 | - | 1 | - | ||
| 3.723–4.205 | 107/63 | 0.61 | 0.44–0.85 | 0.71 | 0.51–0.99 | ||
| >4.205 | 107/40 | 0.32 | 0.22–0.46 | 0.51 | 0.33–0.77 | ||
| Continuous variable | 320/183 | 0.52 | 0.41–0.64 | <0.001 | 0.68 | 0.53–0.87 | 0.003 |
| Event Free Survival | |||||||
| Median | <0.001 | 0.006 | |||||
| ≤3.994 (reference) | 154/123 | 1 | - | 1 | - | ||
| >3.994 | 160/83 | 0.45 | 0.34–0.60 | 0.65 | 0.48–0.89 | ||
| Tertiles | <0.001 | 0.001 | |||||
| ≤3.723 (reference) | 100/83 | 1 | - | 1 | - | ||
| 3.723–4.205 | 107/70 | 0.56 | 0.41–0.78 | 0.66 | 0.48–0.91 | ||
| >4.205 | 107/53 | 0.36 | 0.25–0.50 | 0.53 | 0.36–0.79 | ||
| Continuous variable | 314/206 | 0.51 | 0.41–0.63 | <0.001 | 0.65 | 0.51–0.85 | 0.002 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ognibene, M.; De Marco, P.; Amoroso, L.; Cangelosi, D.; Zara, F.; Parodi, S.; Pezzolo, A. Multiple Genes with Potential Tumor Suppressive Activity Are Present on Chromosome 10q Loss in Neuroblastoma and Are Associated with Poor Prognosis. Cancers 2023, 15, 2035. https://doi.org/10.3390/cancers15072035
Ognibene M, De Marco P, Amoroso L, Cangelosi D, Zara F, Parodi S, Pezzolo A. Multiple Genes with Potential Tumor Suppressive Activity Are Present on Chromosome 10q Loss in Neuroblastoma and Are Associated with Poor Prognosis. Cancers. 2023; 15(7):2035. https://doi.org/10.3390/cancers15072035
Chicago/Turabian StyleOgnibene, Marzia, Patrizia De Marco, Loredana Amoroso, Davide Cangelosi, Federico Zara, Stefano Parodi, and Annalisa Pezzolo. 2023. "Multiple Genes with Potential Tumor Suppressive Activity Are Present on Chromosome 10q Loss in Neuroblastoma and Are Associated with Poor Prognosis" Cancers 15, no. 7: 2035. https://doi.org/10.3390/cancers15072035