
Combination Therapy with a Bispecific Antibody Targeting the hERG1/β1 Integrin Complex and Gemcitabine in Pancreatic Ductal Adenocarcinoma
 , , , and
, , , and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. In Silico Analysis
2.2. Sample Collection and Clinicopathological Characteristics
2.3. Immunohistochemistry (IHC)
2.4. Western Blot (WB) and Co-Immunoprecipitation (Co-IP)
2.5. Cell Culture
2.6. Cell Viability Assay
2.7. scDb-hERG1-β1 Antibody Production and Purification
2.8. Labelling of the scDb-hERG1/β1 with Alexa 488 and Indiocyanine Green (ICG)
2.9. Immunofluorescence (IF)
2.10. Mouse In Vivo Model
2.11. Ultrasound (US) and Photoacoustic (PA) Imaging
2.12. Statistics
3. Results
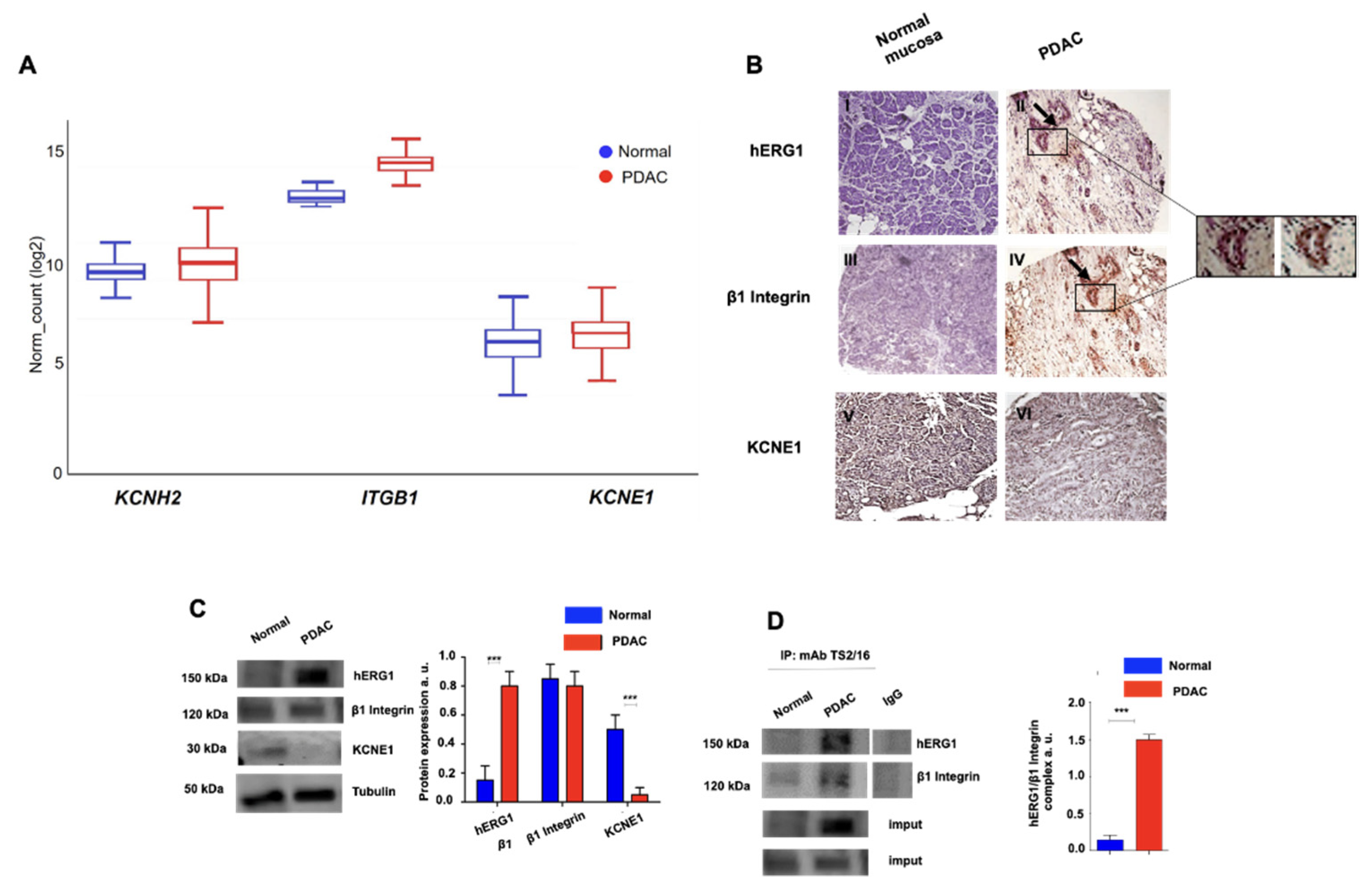
3.1. Expression of the hERG1/β1 Integrin Complex in PDAC
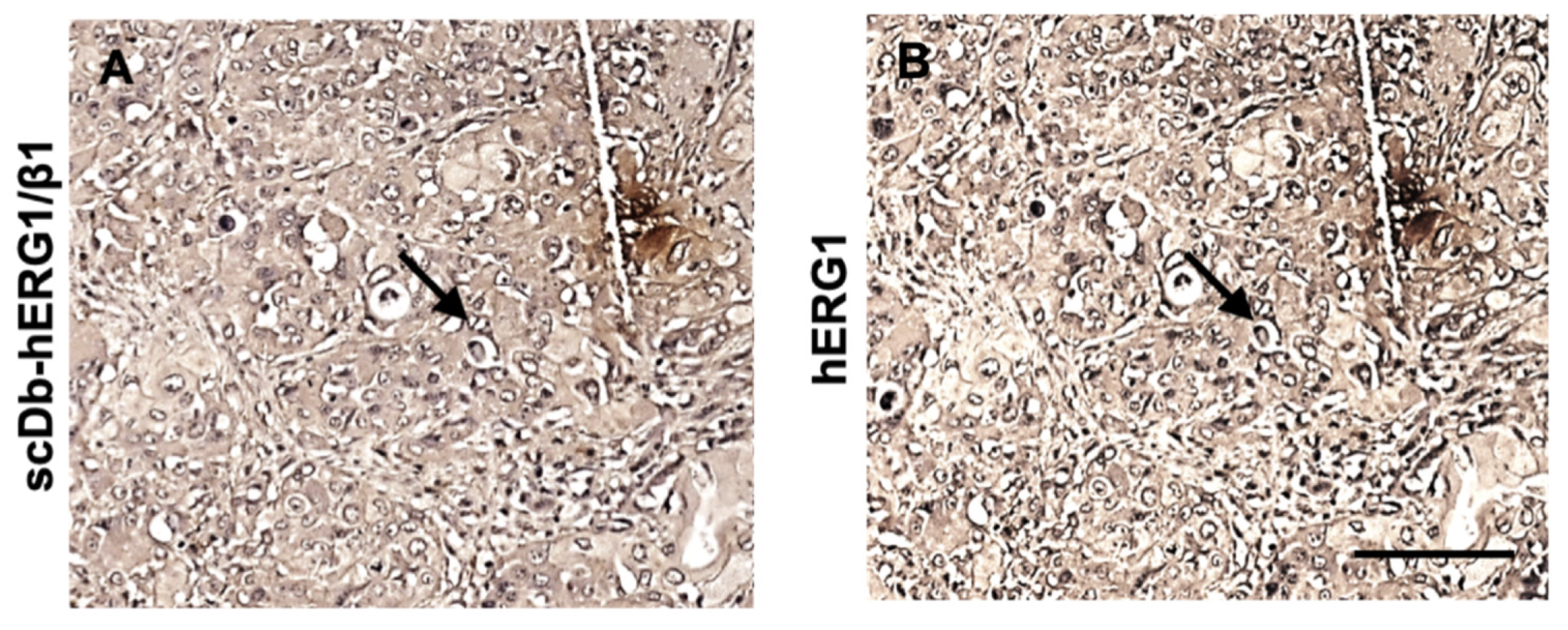
3.1.1. Primary Human PDAC Samples
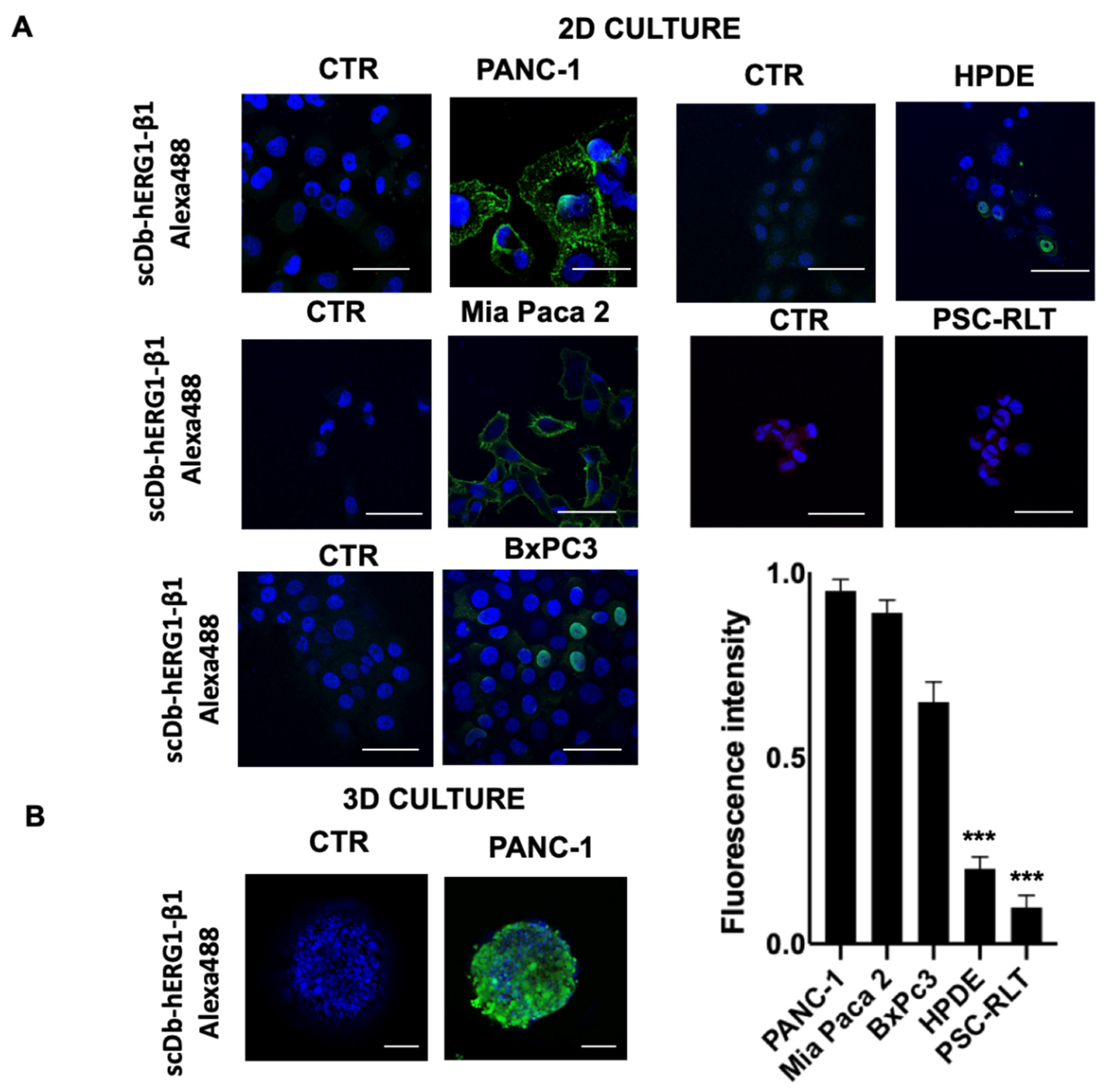
3.1.2. PDAC Cell Lines
3.2. Combination of scDb hERG1/β1 Integrin with Gemcitabine: In Vitro Data
3.3. Combination of the scDb hERG1/β1 Integrin with Gemcitabine: In Vivo Data
3.3.1. Penetration of the scDb hERG1/β1 into the Tumour Masses of an Orthotopic Xenograft PDAC Mouse Model: Evidence with Photo Acoustic (PA) Imaging
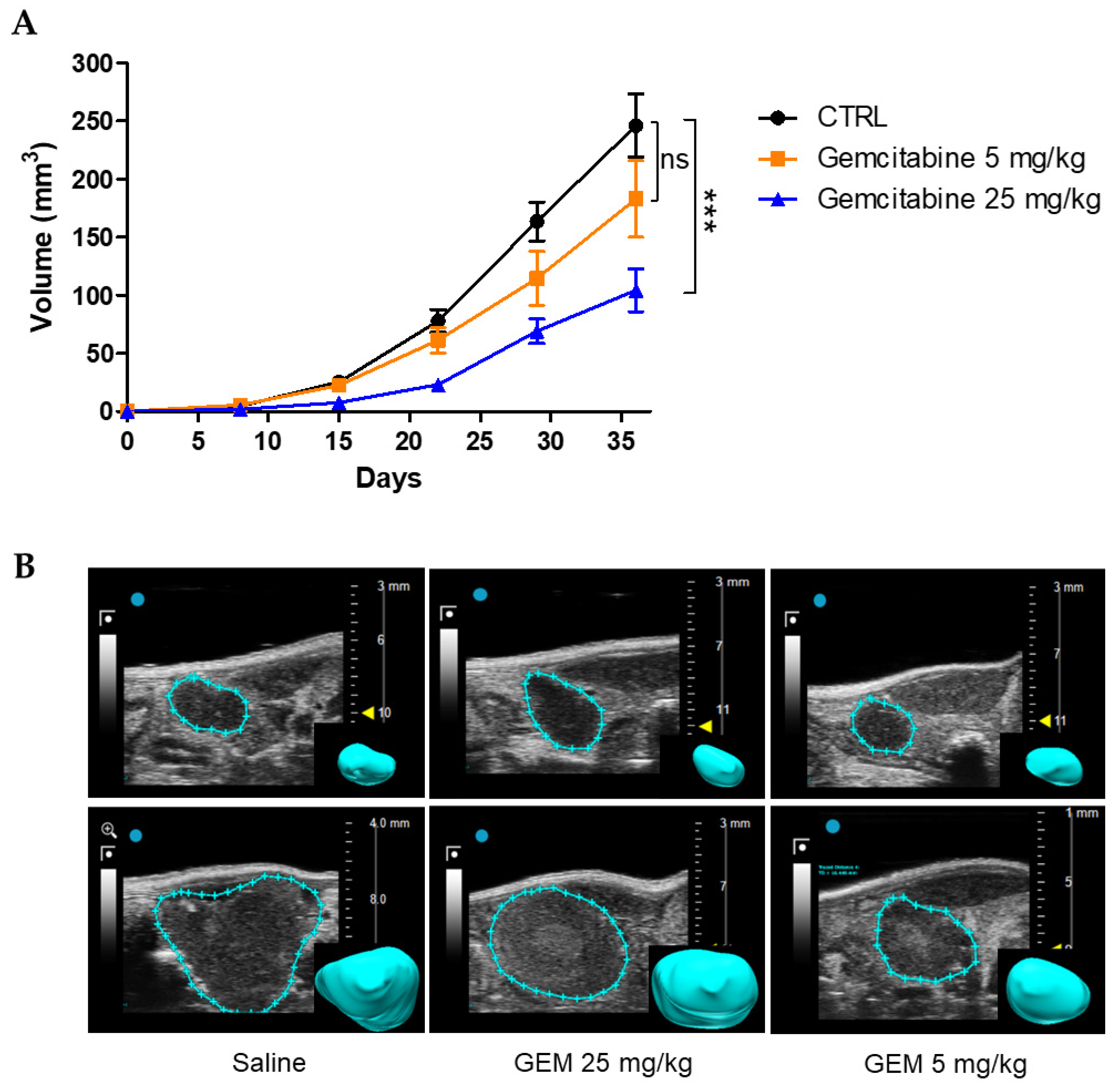
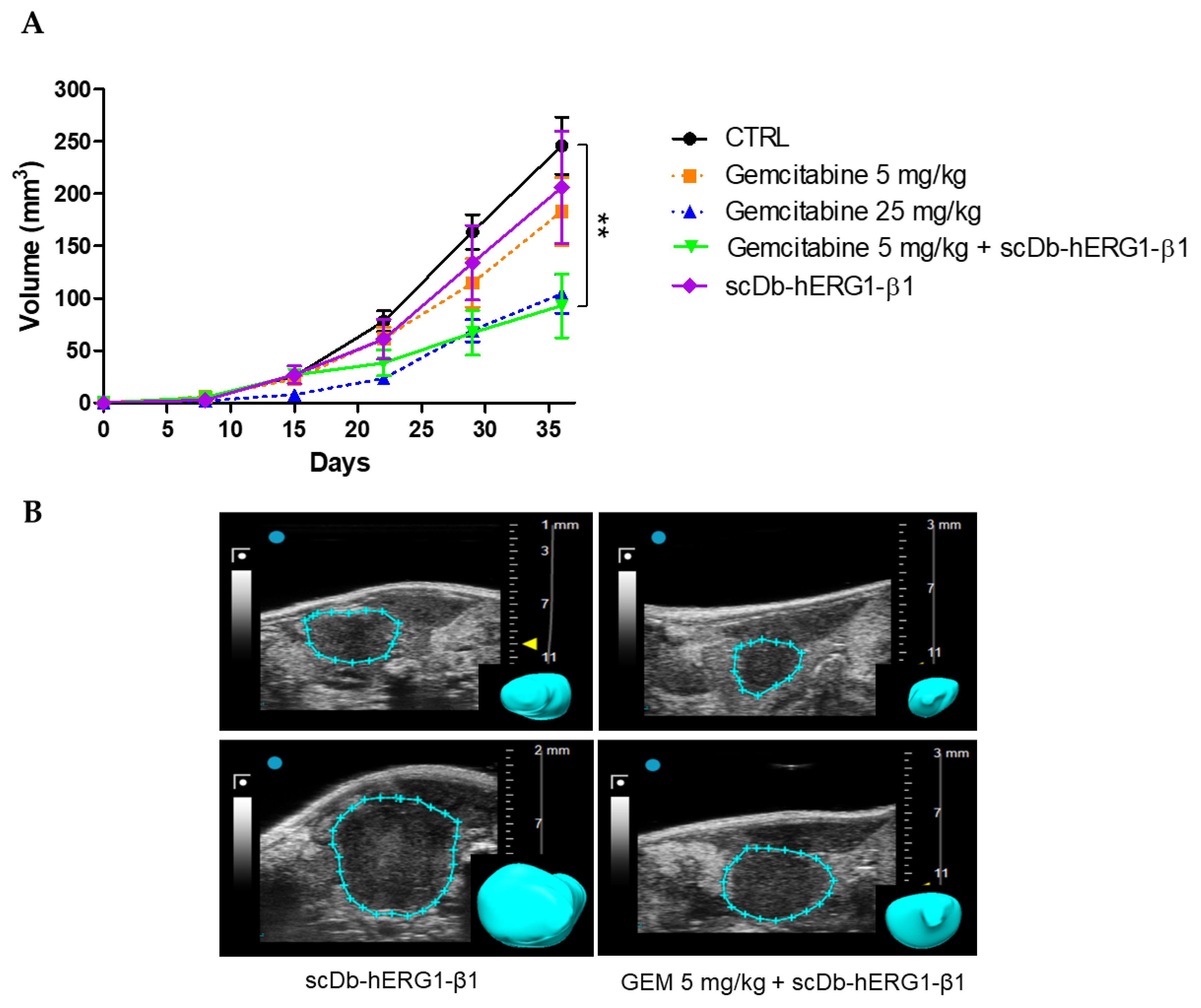
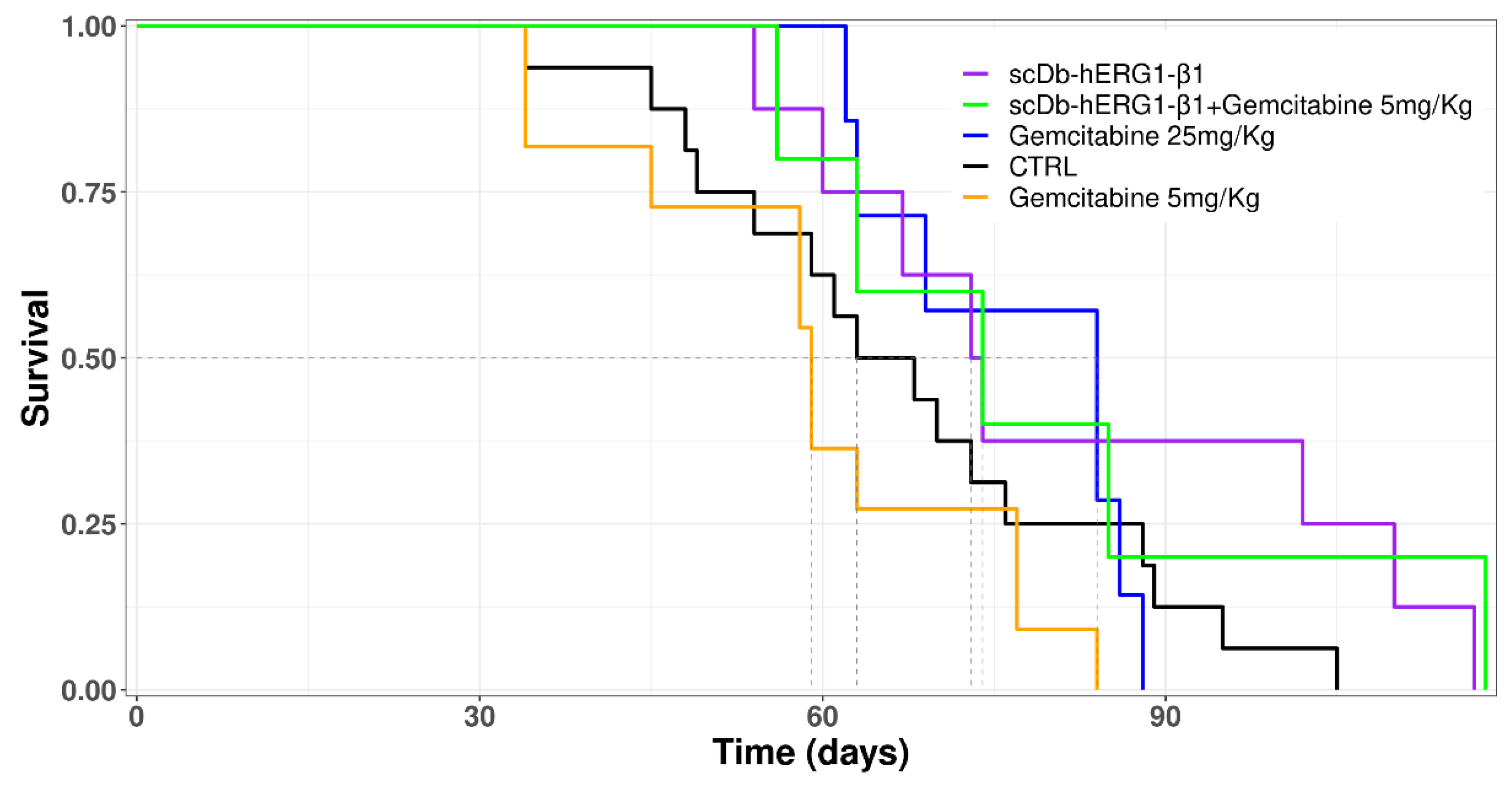
3.3.2. Effects of Gemcitabine in the Orthotopic Xenograft PDAC Mouse Model
3.3.3. Effects of Gemcitabine in Combination with scDb-hERG1-β1
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer Incidence and Mortality Worldwide: Sources, Methods and Major Patterns in GLOBOCAN 2012. Int. J. Cancer 2015, 136, E359–E386. [Google Scholar] [CrossRef] [PubMed]
- Partensky, C. Toward a Better Understanding of Pancreatic Ductal Adenocarcinoma. Pancreas 2013, 42, 729–739. [Google Scholar] [CrossRef] [PubMed]
- Amanam, I.; Chung, V. Targeted Therapies for Pancreatic Cancer. Cancers 2018, 10, 36. [Google Scholar] [CrossRef] [PubMed]
- Baudino, T. Targeted Cancer Therapy: The Next Generation of Cancer Treatment. Curr. Drug. Discov. Technol. 2015, 12, 3–20. [Google Scholar] [CrossRef]
- Mokhtari, R.B.; Homayouni, T.S.; Baluch, N.; Morgatskaya, E.; Kumar, S.; Das, B.; Yeger, H. Combination Therapy in Combating Cancer. Oncotarget 2017, 8, 38022–38043. [Google Scholar] [CrossRef]
- Lastraioli, E.; Iorio, J.; Arcangeli, A. Ion Channel Expression as Promising Cancer Biomarker. Biochim. Biophys. Acta BBA Biomembr. 2015, 1848, 2685–2702. [Google Scholar] [CrossRef]
- Huang, X.; Jan, L.Y. Targeting Potassium Channels in Cancer. J. Cell Biol. 2014, 206, 151–162. [Google Scholar] [CrossRef]
- Bianchi, L.; Wible, B.; Arcangeli, A.; Taglialatela, M.; Morra, F.; Castaldo, P.; Crociani, O.; Rosati, B.; Faravelli, L.; Olivotto, M.; et al. Herg Encodes a K+ Current Highly Conserved in Tumors of Different Histogenesis: A Selective Advantage for Cancer Cells? Cancer Res. 1998, 58, 815–822. [Google Scholar]
- Cherubini, A.; Taddei, G.L.; Crociani, O.; Paglierani, M.; Buccoliero, A.M.; Fontana, L.; Noci, I.; Borri, P.; Borrani, E.; Giachi, M.; et al. HERG Potassium Channels Are More Frequently Expressed in Human Endometrial Cancer as Compared to Non-Cancerous Endometrium. Br. J. Cancer 2000, 83, 1722–1729. [Google Scholar] [CrossRef]
- Crociani, O.; Guasti, L.; Balzi, M.; Becchetti, A.; Wanke, E.; Olivotto, M.; Wymore, R.S.; Arcangeli, A. Cell Cycle-Dependent Expression of HERG1 and HERG1B Isoforms in Tumor Cells. J. Biol. Chem. 2003, 278, 2947–2955. [Google Scholar] [CrossRef]
- Lastraioli, E.; Guasti, L.; Crociani, O.; Polvani, S.; Hofmann, G.; Witchel, H.; Bencini, L.; Calistri, M.; Messerini, L.; Scatizzi, M.; et al. Herg1 Gene and HERG1 Protein Are Overexpressed in Colorectal Cancers and Regulate Cell Invasion of Tumor Cells. Cancer Res. 2004, 64, 606–611. [Google Scholar] [CrossRef] [PubMed]
- Masi, A.; Becchetti, A.; Restano-Cassulini, R.; Polvani, S.; Hofmann, G.; Buccoliero, A.M.; Paglierani, M.; Pollo, B.; Taddei, G.L.; Gallina, P.; et al. HERG1 Channels Are Overexpressed in Glioblastoma Multiforme and Modulate VEGF Secretion in Glioblastoma Cell Lines. Br. J. Cancer 2005, 93, 781–792. [Google Scholar] [CrossRef]
- Shao, X.-D.; Wu, K.-C.; Guo, X.-Z.; Xie, M.-J.; Zhang, J.; Fan, D.-M. Expression and Significance of HERG Protein in Gastric Cancer. Cancer Biol. 2008, 7, 45–50. [Google Scholar] [CrossRef]
- Lastraioli, E.; Perrone, G.; Sette, A.; Fiore, A.; Crociani, O.; Manoli, S.; D’Amico, M.; Masselli, M.; Iorio, J.; Callea, M.; et al. HERG1 Channels Drive Tumour Malignancy and May Serve as Prognostic Factor in Pancreatic Ductal Adenocarcinoma. Br. J. Cancer 2015, 112, 1076–1087. [Google Scholar] [CrossRef]
- Iorio, J.; Meattini, I.; Bianchi, S.; Bernini, M.; Maragna, V.; Dominici, L.; Casella, D.; Vezzosi, V.; Orzalesi, L.; Nori, J.; et al. HERG1 Channel Expression Associates with Molecular Subtypes and Prognosis in Breast Cancer. Cancer Cell Int. 2018, 18, 93. [Google Scholar] [CrossRef] [PubMed]
- Lastraioli, E.; Pillozzi, S.; Mari, A.; Tellini, R.; Duranti, C.; Baldazzi, V.; Venturini, S.; Minervini, A.; Lapini, A.; Nesi, G.; et al. HERG1 and CA IX Expression Are Associated with Disease Recurrence in Surgically Resected Clear Cell Renal Carcinoma. Eur. J. Surg. Oncol. 2020, 46, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Sanguinetti, M.C.; Jiang, C.; Curran, M.E.; Keating, M.T. A Mechanistic Link between an Inherited and an Acquird Cardiac Arrthytmia: HERG Encodes the IKr Potassium Channel. Cell 1995, 81, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Bauer, C.K.; Schwarz, J.R. Ether-à-Go-Go K + Channels: Effective Modulators of Neuronal Excitability. J. Physiol. 2018, 596, 769–783. [Google Scholar] [CrossRef] [PubMed]
- Parr, E.; Pozo, M.J.; Horowitz, B.; Nelson, M.T.; Mawe, G.M. ERG K + Channels Modulate the Electrical and Contractile Activities of Gallbladder Smooth Muscle. Am. J. Physiol. Gastrointest. Liver Physiol. 2003, 284, G392–G398. [Google Scholar] [CrossRef]
- Bauer, C.K.; Schäfer, R.; Schiemann, D.; Reid, G.; Hanganu, I.; Schwarz, J.R. A Functional Role of the Erg-like Inward-Rectifying K+ Current in Prolactin Secretion from Rat Lactotrophs. Mol. Cell. Endocrinol. 1999, 148, 37–45. [Google Scholar] [CrossRef]
- Gullo, F.; Ales, E.; Rosati, B.; Lecchi, M.; Masi, A.; Guasti, L.; Cano-Abad, M.F.; Arcangeli, A.; Lopez, M.G.; Wanke, E. ERG K + Channel Blockade Enhances Firing and Epinephrine Secretion in Rat Chromaffin Cells: The Missing Link to LQT2-related Sudden Death? FASEB J. 2003, 17, 330–332. [Google Scholar] [CrossRef] [PubMed]
- Binggeli, R.; Weinstein, R.C. Membrane Potentials and Sodium Channels: Hypotheses for Growth Regulation and Cancer Formation Based on Changes in Sodium Channels and Gap Junctions. J. Biol. 1986, 123, 377–401. [Google Scholar] [CrossRef] [PubMed]
- Becchetti, A.; Duranti, C.; Arcangeli, A. Dynamics and Physiological Meaning of Complexes between Ion Channels and Integrin Receptors: The Case of Kv11.1. Am. J. Physiol. Cell Physiol. 2022, 322, C1138–C1150. [Google Scholar] [CrossRef]
- Becchetti, A.; Crescioli, S.; Zanieri, F.; Petroni, G.; Mercatelli, R.; Coppola, S.; Gasparoli, L.; D’Amico, M.; Pillozzi, S.; Crociani, O.; et al. The Conformational State of HERG1 Channels Determines Integrin Association, Downstream Signaling, and Cancer Progression. Sci. Signal. 2017, 10, eaaf3236. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.-W.; Luo, H.-S.; Luo, B.; Xu, D.-Q.; Gao, S. Overexpression of HERG1 in Resected Esophageal Squamous Cell Carcinomas: A Marker for Poor Prognosis. J. Surg. Oncol. 2008, 97, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Iorio, J.; Lastraioli, E.; Tofani, L.; Petroni, G.; Antonuzzo, L.; Messerini, L.; Perrone, G.; Caputo, D.; Francesconi, M.; Amato, M.M.; et al. HERG1 and HIF-2α Behave as Biomarkers of Positive Response to Bevacizumab in Metastatic Colorectal Cancer Patients. Transl. Oncol. 2020, 13, 100740. [Google Scholar] [CrossRef] [PubMed]
- Anderson, K.J.; Cormier, R.T.; Scott, P.M. Role of Ion Channels in Gastrointestinal Cancer. World J. Gastroenterol. 2019, 25, 5732–5772. [Google Scholar] [CrossRef]
- Arcangeli, A.; Duranti, C.; Iorio, J.; Lastraioli, E. The Role of Potassium Channels in Tumours of the Gastrointestinal Tract: A Focus on the Human Ether-à-go-go Related Gene 1 Channels. J. Physiol. 2022. [Google Scholar] [CrossRef]
- Hofschröer, V.; Najder, K.; Rugi, M.; Bouazzi, R.; Cozzolino, M.; Arcangeli, A.; Panyi, G.; Schwab, A. Ion Channels Orchestrate Pancreatic Ductal Adenocarcinoma Progression and Therapy. Front. Pharm. 2021, 11, 586599. [Google Scholar] [CrossRef]
- Arcangeli, A.; Becchetti, A. HERG Channels: From Antitargets to Novel Targets for Cancer Therapy. Clin. Cancer Res. 2017, 23, 3–5. [Google Scholar] [CrossRef]
- Duranti, C.; Iorio, J.; Lottini, T.; Lastraioli, E.; Crescioli, S.; Bagni, G.; Lulli, M.; Capitani, C.; Bouazzi, R.; Stefanini, M.; et al. Harnessing the HERG1/Β1 Integrin Complex via a Novel Bispecific Single-Chain Antibody: An Effective Strategy against Solid Cancers. Mol. Cancer Ther. 2021, 20, 1338–1349. [Google Scholar] [CrossRef]
- James, M.L.; Gambhir, S.S. A Molecular Imaging Primer: Modalities, Imaging Agents, and Applications. Physiol. Rev. 2012, 92, 897–965. [Google Scholar] [CrossRef] [PubMed]
- Goldman, M.J.; Craft, B.; Hastie, M.; Repečka, K.; McDade, F.; Kamath, A.; Banerjee, A.; Luo, Y.; Rogers, D.; Brooks, A.N.; et al. Visualizing and Interpreting Cancer Genomics Data via the Xena Platform. Nat. Biotechnol. 2020, 38, 675–678. [Google Scholar] [CrossRef] [PubMed]
- Aguet, F.; Anand, S.; Ardlie, K.G.; Gabriel, S.; Getz, G.A.; Graubert, A.; Hadley, K.; Handsaker, R.E.; Huang, K.H.; Kashin, S.; et al. The GTEx Consortium Atlas of Genetic Regulatory Effects across Human Tissues. Science 2020, 369, 1318–1330. [Google Scholar] [CrossRef]
- Iorio, J.; Antonuzzo, L.; Scarpi, E.; D’Amico, M.; Duranti, C.; Messerini, L.; Sparano, C.; Caputo, D.; Lavacchi, D.; Borzomati, D.; et al. Prognostic Role of HERG1 Potassium Channels in Neuroendocrine Tumours of the Ileum and Pancreas. Int. J. Mol. Sci. 2022, 23, 10623. [Google Scholar] [CrossRef] [PubMed]
- Pillozzi, S.; D’Amico, M.; Bartoli, G.; Gasparoli, L.; Petroni, G.; Crociani, O.; Marzo, T.; Guerriero, A.; Messori, L.; Severi, M.; et al. The Combined Activation of KCa3.1 and Inhibition of Kv11.1/HERG1 Currents Contribute to Overcome Cisplatin Resistance in Colorectal Cancer Cells. Br. J. Cancer 2018, 118, 200–212. [Google Scholar] [CrossRef]
- Duranti, C.; Lastraioli, E.; Iorio, J.; Capitani, C.; Carraresi, L.; Gonnelli, L.; Arcangeli, A. Expression and Purification of a Novel Single-Chain Diabody (ScDb-HERG1/Β1) from Pichia Pastoris Transformants. Protein Expr. Purif. 2021, 184, 105879. [Google Scholar] [CrossRef]
- Lottini, T.; Buonamici, M.; Duranti, C.; Arcangeli, A. Generation of an Orthotopic Xenograft of Pancreatic Cancer Cells by Ultrasound-Guided Injection. J. Vis. Exp. 2021, e63123. [Google Scholar] [CrossRef]
- Pramanik, A.; Xu, Z.; Shamsuddin, S.H.; Khaled, Y.S.; Ingram, N.; Maisey, T.; Tomlinson, D.; Coletta, P.L.; Jayne, D.; Hughes, T.A.; et al. Affimer Tagged Cubosomes: Targeting of Carcinoembryonic Antigen Expressing Colorectal Cancer Cells Using In Vitro and In Vivo Models. ACS Appl. Mater. Interfaces 2022, 14, 11078–11091. [Google Scholar] [CrossRef]
- Ben-Aharon, O.; Magnezi, R.; Leshno, M.; Goldstein, D.A. Median Survival or Mean Survival: Which Measure Is the Most Appropriate for Patients, Physicians, and Policymakers? Oncologist 2019, 24, 1469–1478. [Google Scholar] [CrossRef]
- Vandenberg, J.I.; Perry, M.D.; Perrin, M.J.; Mann, S.A.; Ke, Y.; Hill, A.P. HERG K+ Channels: Structure, Function, and Clinical Significance. Physiol. Rev. 2012, 92, 1393–1478. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, H.; Mou, L.; Luk, C.; Liu, N.; Karaskova, J.; Squire, J.; Tsao, M.-S. Immortal Human Pancreatic Duct Epithelial Cell Lines with Near Normal Genotype and Phenotype. Am. J. Pathol. 2000, 157, 1623–1631. [Google Scholar] [CrossRef] [PubMed]
- Jesnowski, R.; Fürst, D.; Ringel, J.; Chen, Y.; Schrödel, A.; Kleeff, J.; Kolb, A.; Schareck, W.D.; Löhr, M. Immortalization of Pancreatic Stellate Cells as an in Vitro Model of Pancreatic Fibrosis: Deactivation Is Induced by Matrigel and N-Acetylcysteine. Lab. Investig. 2005, 85, 1276–1291. [Google Scholar] [CrossRef] [PubMed]
- Korb, M.L.; Hartman, Y.E.; Kovar, J.; Zinn, K.R.; Bland, K.I.; Rosenthal, E.L. Use of Monoclonal Antibody-IRDye800CW Bioconjugates in the Resection of Breast Cancer. J. Surg. Res. 2014, 188, 119–128. [Google Scholar] [CrossRef]
- Day, K.E.; Sweeny, L.; Kulbersh, B.; Zinn, K.R.; Rosenthal, E.L. Preclinical Comparison of Near-Infrared-Labeled Cetuximab and Panitumumab for Optical Imaging of Head and Neck Squamous Cell Carcinoma. Mol. Imaging Biol. 2013, 15, 722–729. [Google Scholar] [CrossRef]
- Duranti, C.; Arcangeli, A. Ion Channel Targeting with Antibodies and Antibody Fragments for Cancer Diagnosis. Antibodies 2019, 8, 33. [Google Scholar] [CrossRef] [PubMed]
- Duranti, C.; Carraresi, L.; Sette, A.; Stefanini, M.; Lottini, T.; Crescioli, S.; Crociani, O.; Iamele, L.; de Jonge, H.; Gherardi, E.; et al. Generation and Characterization of Novel Recombinant Anti-HERG1 ScFv Antibodies for Cancer Molecular Imaging. Oncotarget 2018, 9, 34972–34989. [Google Scholar] [CrossRef]
- Santini, L.; Duranti, C.; Palandri, C.; Giammarino, L.; Carlucci, L.; Capitani, C.; Colasurdo, R.; Recchia, F.; Cerbai, E.; Coppini, R.; et al. Functional Characterization of a Novel Recombinant Bispecific Antibody Targeting the Ether-à-Go-Go Related Gene 1 (HERG1)-B1 Integrin Macromolecular Complex in Human Cardiomyocytes. 2023; manuscript in preparation; to be submitted. [Google Scholar]
- Gargiulo, S.; Albanese, S.; Mancini, M. State-of-the-Art Preclinical Photoacoustic Imaging in Oncology: Recent Advances in Cancer Theranostics. Contrast Media Mol. Imaging 2019, 2019, 5080267. [Google Scholar] [CrossRef]
- Yu, J.; Nguyen, H.N.Y.; Steenbergen, W.; Kim, K. Recent Development of Technology and Application of Photoacoustic Molecular Imaging Toward Clinical Translation. J. Nucl. Med. 2018, 59, 1202–1207. [Google Scholar] [CrossRef]
- Nishio, N.; van den Berg, N.S.; Martin, B.A.; van Keulen, S.; Fakurnejad, S.; Rosenthal, E.L.; Wilson, K.E. Photoacoustic Molecular Imaging for the Identification of Lymph Node Metastasis in Head and Neck Cancer Using an Anti-EGFR Antibody–Dye Conjugate. J. Nucl. Med. 2021, 62, 648–655. [Google Scholar] [CrossRef]
- Wang, R.; Song, C.; Gao, A.; Liu, Q.; Guan, W.; Mei, J.; Ma, L.; Cui, D. Antibody-Conjugated Liposomes Loaded with Indocyanine Green for Oral Targeted Photoacoustic Imaging-Guided Sonodynamic Therapy of Helicobacter Pylori Infection. Acta Biomater. 2022, 143, 418–427. [Google Scholar] [CrossRef] [PubMed]
- Amrutkar, M.; Gladhaug, I. Pancreatic Cancer Chemoresistance to Gemcitabine. Cancers 2017, 9, 157. [Google Scholar] [CrossRef] [PubMed]
- Djamgoz, M.B.A. Combinatorial Therapy of Cancer: Possible Advantages of Involving Modulators of Ionic Mechanisms. Cancers 2022, 14, 2703. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Normal Mucosa | Ductal Adenocarcinoma | ||||||
|---|---|---|---|---|---|---|---|
| hERG1 − | hERG1 + | Pearson Correlation | hERG1 − | hERG1 + | Pearson Correlation | ||
| β1 Integrin − | 23/40 (57.5%) | 0/40 (0%) | R: −0.1594 p: 0.33 | β1 Integrin − | 5/40 (12.5%) | 2/40 (5%) | R: 0.4835 p: 0.001 |
| β1 Integrin + | 17/40 (42.5%) | 0/40 (0%) | β1 Integrin + | 6/40 (15%) | 27/40 (67.5%) | ||
| hERG1 − | hERG1 + | Pearson Correlation | hERG1 − | hERG1 + | Pearson Correlation | ||
| KCNE1 − | 21/40 (52.5%) | 0/40 (0%) | R: −0.0918 p: 0.58 | KCNE1 − | 9/40 (22.5%) | 26/40 (65%) | R: −0.0954 p: 0.96 |
| KCNE1 + | 19/40 (47.5%) | 0/40 (0%) | KCNE1 + | 2/40 (5%) | 3/40 (7.5%) | ||
| β1 Integrin − | β1 Integrin + | Pearson Correlation | β1 Integrin − | β1 Integrin + | Pearson Correlation | ||
| KCNE1 − | 21/40 (52.5%) | 0/40 (0%) | R: 1.0000 p < 0.0001 | KCNE1 − | 7/40 (17.5%) | 28/40 (70%) | R: 0.2212 p: 0.19 |
| KCNE1 + | 2/40 (5%) | 17/40 (42.5%) | KCNE1 + | 0/40 (0%) | 5/40 (12.5%) | ||
| Cell Line | IC50 Gemcitabine (μM) | IC50 scDb-hERG1-β1 (μg/mL) | Combination Index (CI) |
|---|---|---|---|
| PANC-1 | 40.66 | 18.13 | 0.97 |
| MiaPaca-2 | 47.90 | 20.40 | 0.93 |
| BxPC3 | 156.23 | 551.78 | 0.92 |
| PSC-RLT | 4.17 | 166.21 | 1.34 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lottini, T.; Duranti, C.; Iorio, J.; Martinelli, M.; Colasurdo, R.; D’Alessandro, F.N.; Buonamici, M.; Coppola, S.; Devescovi, V.; La Vaccara, V.; et al. Combination Therapy with a Bispecific Antibody Targeting the hERG1/β1 Integrin Complex and Gemcitabine in Pancreatic Ductal Adenocarcinoma. Cancers 2023, 15, 2013. https://doi.org/10.3390/cancers15072013
Lottini T, Duranti C, Iorio J, Martinelli M, Colasurdo R, D’Alessandro FN, Buonamici M, Coppola S, Devescovi V, La Vaccara V, et al. Combination Therapy with a Bispecific Antibody Targeting the hERG1/β1 Integrin Complex and Gemcitabine in Pancreatic Ductal Adenocarcinoma. Cancers. 2023; 15(7):2013. https://doi.org/10.3390/cancers15072013
Chicago/Turabian StyleLottini, Tiziano, Claudia Duranti, Jessica Iorio, Michele Martinelli, Rossella Colasurdo, Franco Nicolás D’Alessandro, Matteo Buonamici, Stefano Coppola, Valentina Devescovi, Vincenzo La Vaccara, and et al. 2023. "Combination Therapy with a Bispecific Antibody Targeting the hERG1/β1 Integrin Complex and Gemcitabine in Pancreatic Ductal Adenocarcinoma" Cancers 15, no. 7: 2013. https://doi.org/10.3390/cancers15072013