
The Pro-Tumorigenic Role of Chemotherapy-Induced Extracellular HSP70 from Breast Cancer Cells via Intratumoral Macrophages
 , , , , and
, , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Lines and Chemicals
2.2. Differentiation of THP-1 to Macrophage
2.3. Coculture Experiment
2.4. Cell Proliferation Assay, Chemoresistance Assay, and Wound Healing Assay
2.5. Collection of the Conditioned Medium and Downstream Experiments
2.6. Isolation of sEVs
2.7. Western Blotting
2.8. Fluorescence Immunocytochemistry
2.9. Small Interfering RNA Transfection
2.10. Real-Time PCR
2.11. Patients and Tissues
2.12. Immunohistochemistry
2.13. Scoring of HSP70 Immunoreactivity
2.14. Statistical Analyses
3. Results
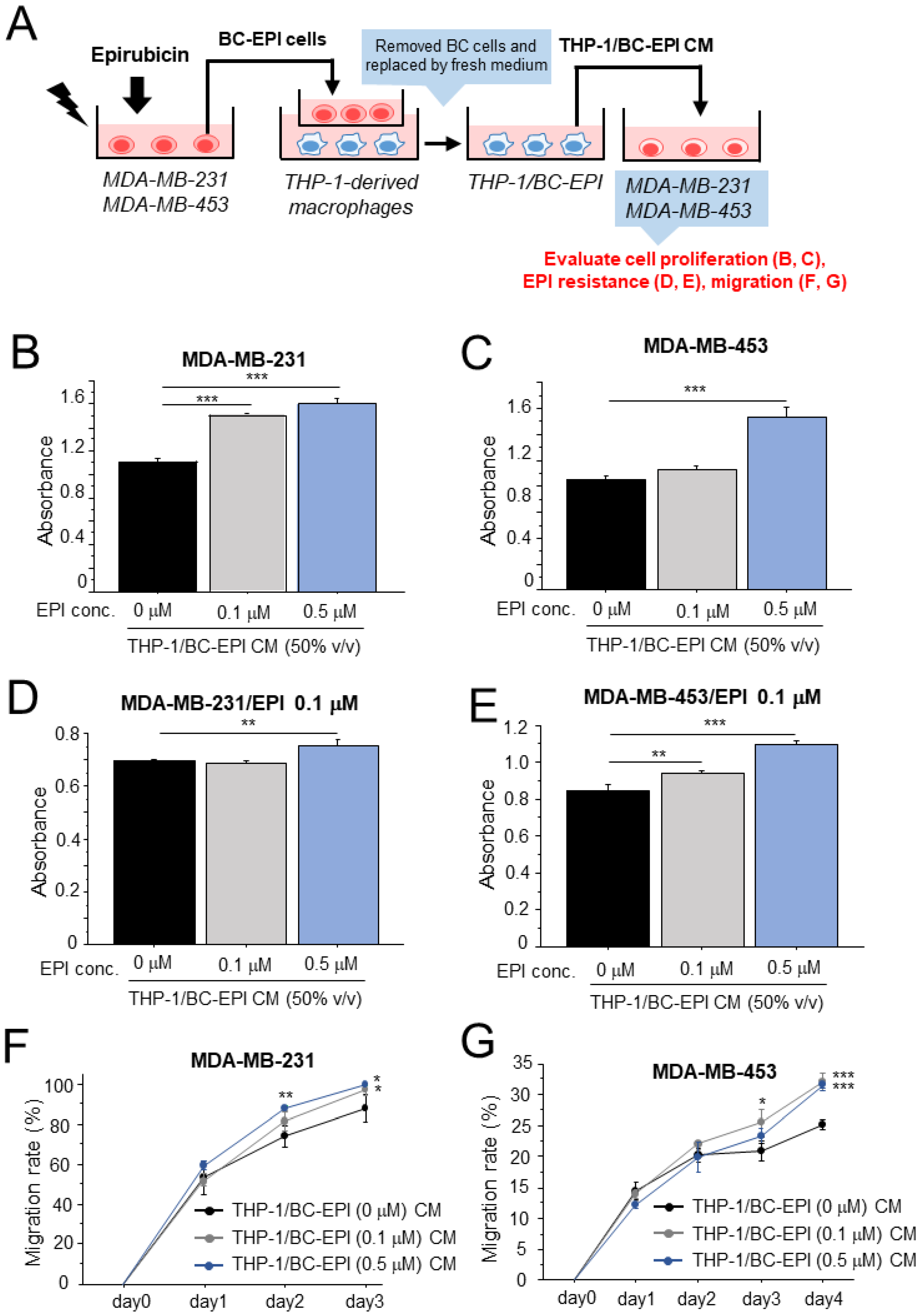
3.1. Altered Characteristics of Macrophages following Chemotherapy in Breast Cancer
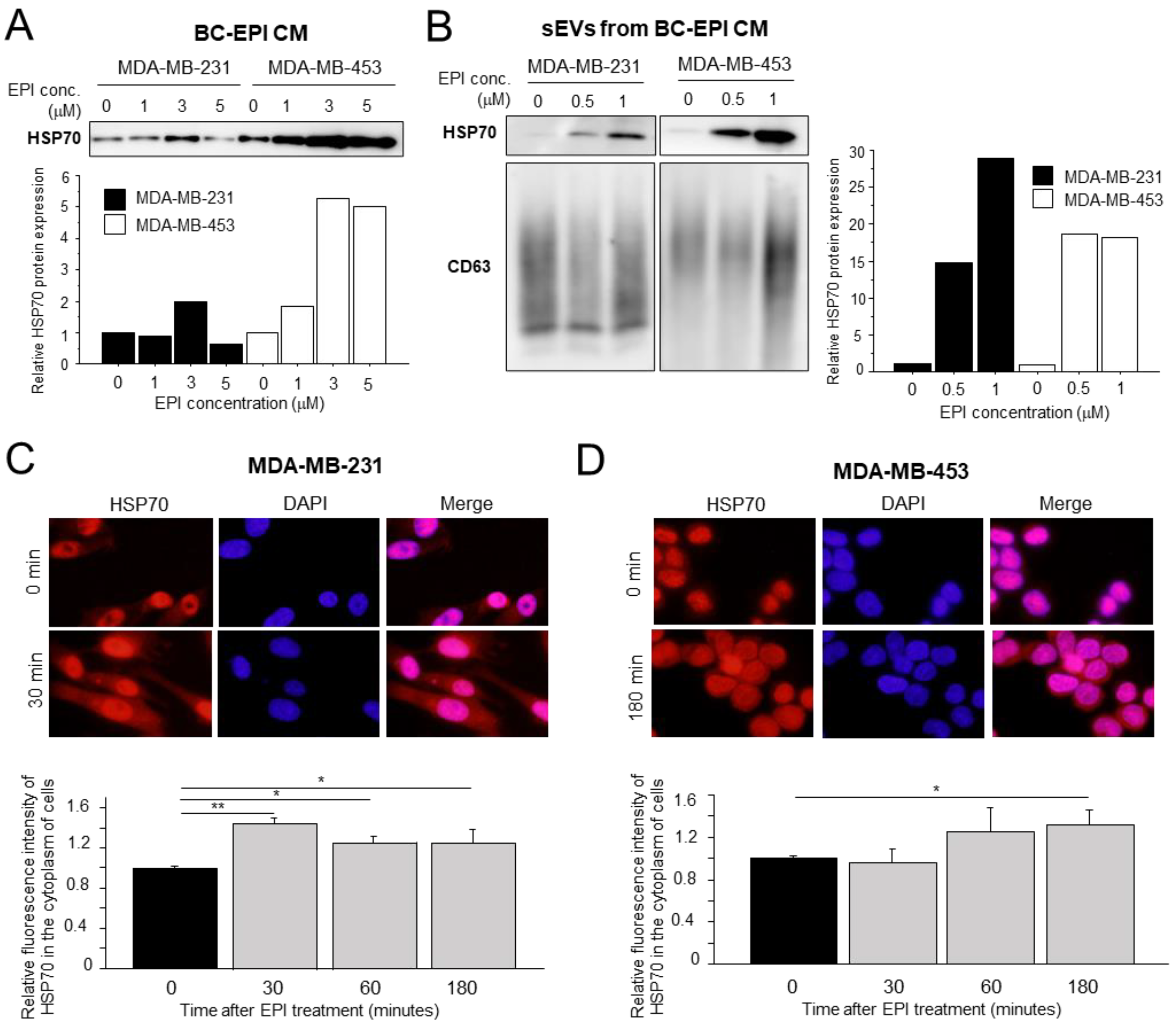
3.2. Effects of Chemotherapy on Extracellular HSP70 Secretion in Breast Cancer Cells
3.3. Direct or Indirect Effects of Extracellular HSP70 on Macrophages
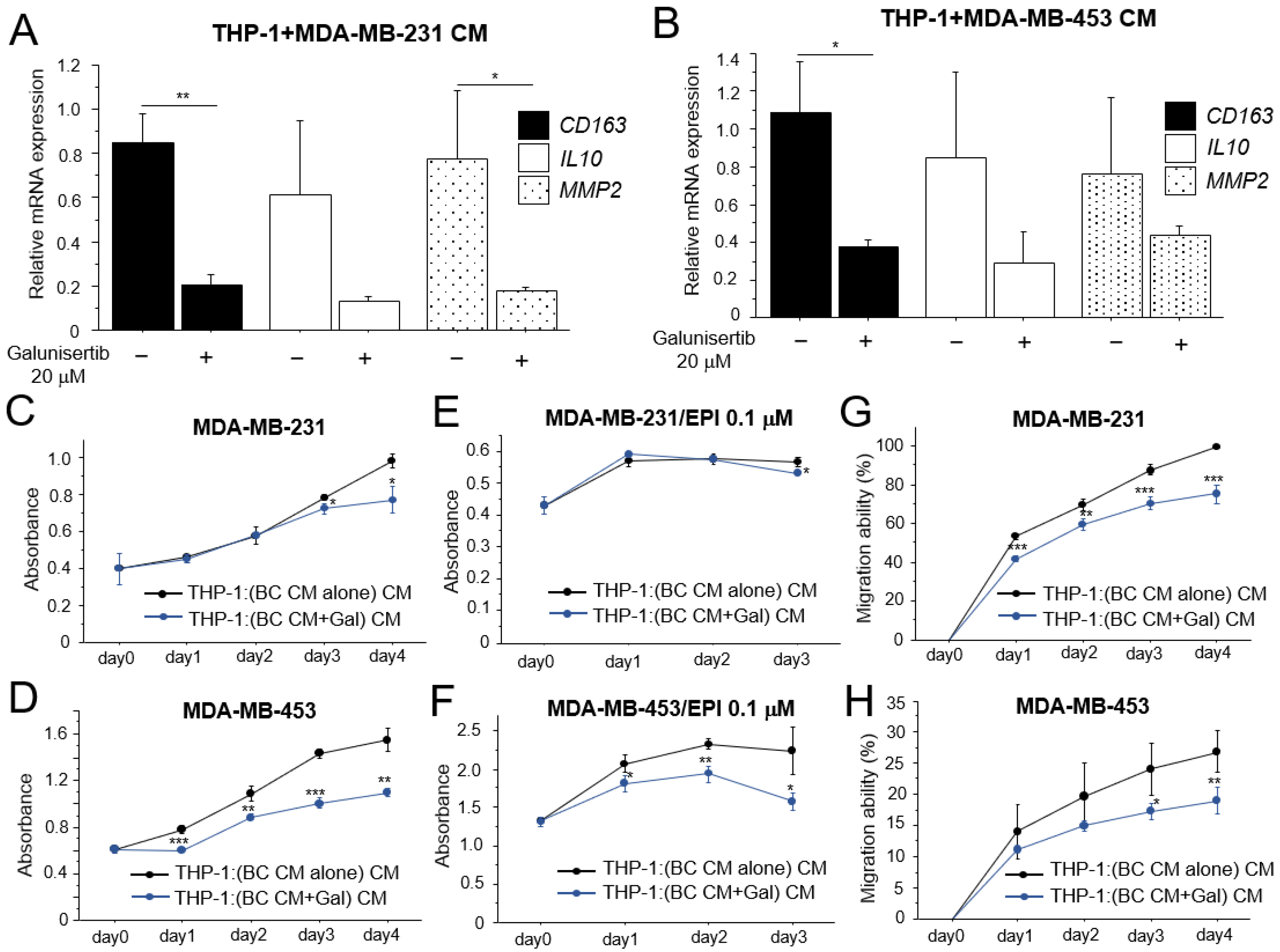
3.4. Effects of TGF-β on the Pro-Tumorigenic Effects of Macrophages
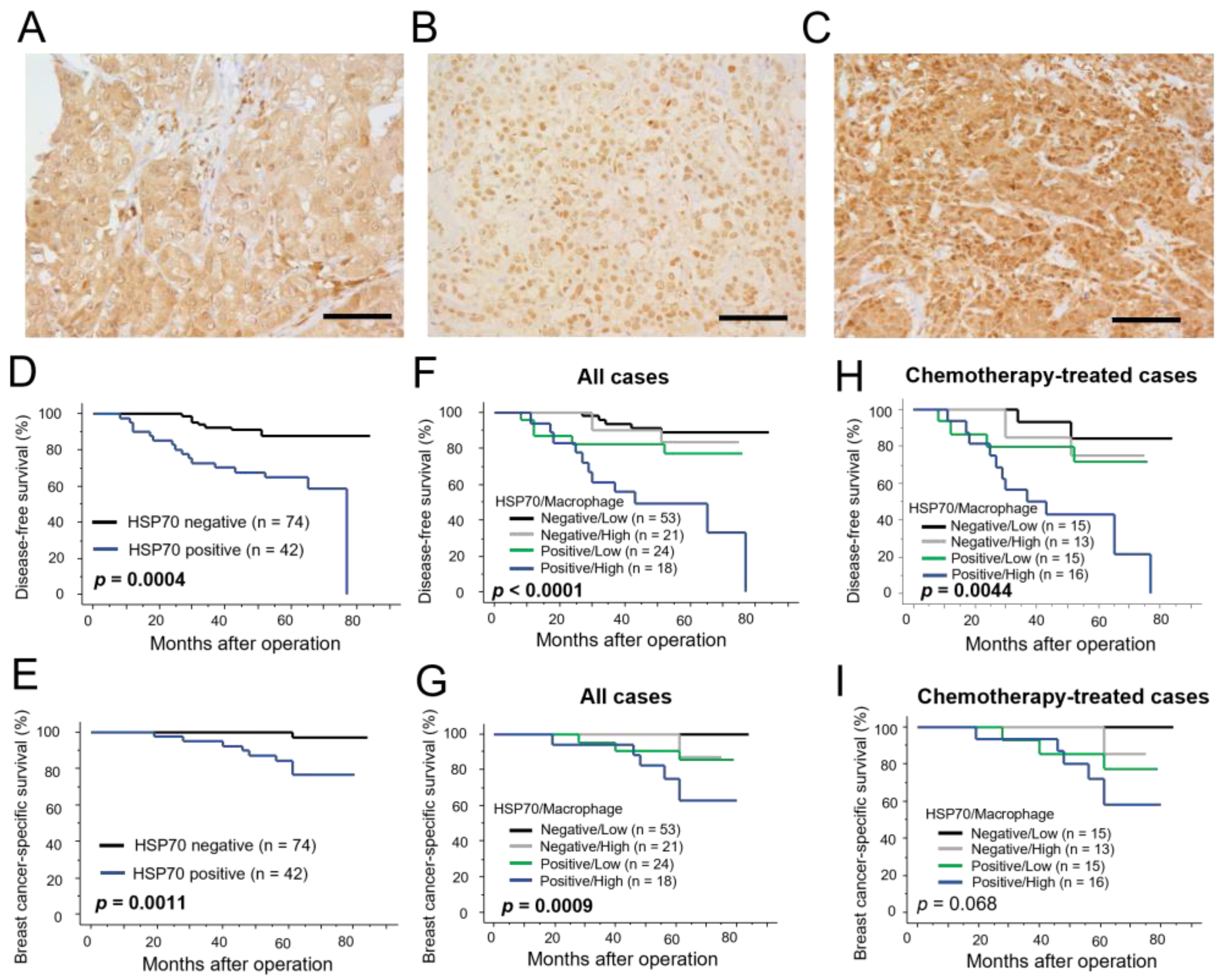
3.5. Immunolocalization of HSP70 in Human Breast Carcinoma Tissues
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tevaarwerk, A.J.; Gray, R.J.; Schneider, B.P.; Smith, M.L.; Wagner, L.I.; Fetting, J.H.; Davidson, N.; Goldstein, L.J.; Miller, K.D.; Sparano, J.A. Survival in patients with metastatic recurrent breast cancer after adjuvant chemotherapy: Little evidence of improvement over the past 30 years. Cancer 2013, 119, 1140–1148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whiteside, T.L. The tumor microenvironment and its role in promoting tumor growth. Oncogene 2008, 27, 5904–5912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, P.J.; Allen, J.E.; Biswas, S.K.; Fisher, E.A.; Gilroy, D.W.; Goerdt, S.; Gordon, S.; Hamilton, J.A.; Ivashkiv, L.B.; Lawrence, T.; et al. Macrophage activation and polarization: Nomenclature and experimental guidelines. Immunity 2014, 41, 14–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sousa, S.; Brion, R.; Lintunen, M.; Kronqvist, P.; Sandholm, J.; Mönkkönen, J.; Kellokumpu-Lehtinen, P.L.; Lauttia, S.; Tynninen, O.; Joensuu, H.; et al. Human breast cancer cells educate macrophages toward the M2 activation status. Breast Cancer Res. 2015, 17, 101. [Google Scholar] [CrossRef] [Green Version]
- Lin, E.Y.; Li, J.F.; Gnatovskiy, L.; Deng, Y.; Zhu, L.; Grzesik, D.A.; Qian, H.; Xue, X.N.; Pollard, J.W. Macrophages regulate the angiogenic switch in a mouse model of breast cancer. Cancer Res. 2006, 66, 11238–11246. [Google Scholar] [CrossRef] [Green Version]
- Kim, O.H.; Kang, G.H.; Noh, H.; Cha, J.Y.; Lee, H.J.; Yoon, J.H.; Mamura, M.; Nam, J.S.; Lee, D.H.; Kim, Y.A.; et al. Proangiogenic TIE2(+)/CD31 (+) macrophages are the predominant population of tumor-associated macrophages infiltrating metastatic lymph nodes. Mol. Cells 2013, 36, 432–438. [Google Scholar] [CrossRef] [Green Version]
- Vasiliadou, I.; Holen, I. The role of macrophages in bone metastasis. J. Bone Oncol. 2013, 2, 158–166. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Li, X.; Liu, X.; Liu, Y. The role of tumor-associated macrophages in breast carcinoma invasion and metastasis. Int. J. Clin. Exp. Pathol. 2015, 8, 6656–6664. [Google Scholar]
- Gan, L.; Qiu, Z.; Huang, J.; Li, Y.; Huang, H.; Xiang, T.; Wan, J.; Hui, T.; Lin, Y.; Li, H.; et al. Cyclooxygenase-2 in tumor-associated macrophages promotes metastatic potential of breast cancer cells through Akt pathway. Int. J. Biol. Sci. 2016, 12, 1533–1543. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Wang, J.; Zhang, M.; Xuan, Q.; Wang, Z.; Lian, X.; Zhang, Q. Jagged1 promotes aromatase inhibitor resistance by modulating tumor-associated macrophage differentiation in breast cancer patients. Breast Cancer Res. Treat. 2017, 166, 95–107. [Google Scholar] [CrossRef]
- Gelsomino, L.; Giordano, C.; Camera, G.; Sisci, D.; Marsico, S.; Campana, A.; Tarallo, R.; Rinaldi, A.; Fuqua, S.; Leggio, A.; et al. Leptin Signaling Contributes to Aromatase Inhibitor Resistant Breast Cancer Cell Growth and Activation of Macrophages. Biomolecules 2020, 10, 543. [Google Scholar] [CrossRef] [Green Version]
- García-Martínez, E.; Gil, G.L.; Benito, A.C.; González-Billalabeitia, E.; Conesa, M.A.V.; García, T.G.; García-Garre, E.; Vicente, V.; de la Peña, F.A. Tumor-infiltrating immune cell profiles and their change after neoadjuvant chemotherapy predict response and prognosis of breast cancer. Breast Cancer Res. 2014, 16, 488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Palma, M.; Lewis, C.E. Macrophage regulation of tumor responses to anticancer therapies. Cancer Cell 2013, 23, 277–286. [Google Scholar] [CrossRef] [Green Version]
- Ruffell, B.; Coussens, L.M. Macrophages and therapeutic resistance in cancer. Cancer Cell 2015, 27, 462–472. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; He, K.; Liu, P.; Xu, L.X. Iron participated in breast cancer chemoresistance by reinforcing IL-6 paracrine loop. Biochem. Biophys. Res. Commun. 2016, 475, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Mitchem, J.B.; Brennan, D.J.; Knolhoff, B.L.; Belt, B.A.; Zhu, Y.; Sanford, D.E.; Belaygorod, L.; Carpenter, D.; Collins, L.; Piwnica-Worms, D.; et al. Targeting tumor-infiltrating macrophages decreases tumor-initiating cells, relieves immunosuppression, and improves chemotherapeutic responses. Cancer Res. 2013, 73, 1128–1141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fink, A.L. Chaperone-mediated protein folding. Physiol. Rev. 1999, 79, 425–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, M.E. The HSP70 family and cancer. Carcinogenesis 2013, 34, 1181–1188. [Google Scholar] [CrossRef] [Green Version]
- Asea, A.; Kraeft, S.K.; Kurt-Jones, E.A.; Stevenson, M.A.; Chen, L.B.; Finberg, R.W.; Koo, G.C.; Calderwood, S.K. HSP70 stimulates cytokine production through a CD14-dependant pathway, demonstrating its dual role as a chaperone and cytokine. Nat. Med. 2000, 6, 435–442. [Google Scholar] [CrossRef]
- Boudesco, C.; Cause, S.; Jego, G.; Garrido, C. Hsp70: A Cancer Target Inside and Outside the Cell. Methods Mol. Biol. 2018, 1709, 371–396. [Google Scholar]
- Borges, T.J.; Wieten, L.; van Herwijnen, M.J.; Broere, F.; van der Zee, R.; Bonorino, C.; van Eden, W. The anti-inflammatory mechanisms of Hsp70. Front. Immunol. 2012, 3, 95. [Google Scholar] [CrossRef] [Green Version]
- Wachstein, J.; Tischer, S.; Figueiredo, C.; Limbourg, A.; Falk, C.; Immenschuh, S.; Blasczyk, R.; Eiz-Vesper, B. HSP70 Enhances Immunosuppressive Function of CD4+CD25+FoxP3+ T Regulatory Cells and Cytotoxicity in CD4+CD25− T Cells. PLoS ONE 2012, 7, e51747. [Google Scholar] [CrossRef] [Green Version]
- Chalmin, F.; Ladoire, S.; Mignot, G.; Vincent, J.; Bruchard, M.; Remy-Martin, J.P.; Boireau, W.; Rouleau, A.; Simon, B.; Lanneau, D.; et al. Membrane-associated Hsp72 from tumor-derived exosomes mediates STAT3-dependent immunosuppressive function of mouse and human myeloid-derived suppressor cells. J. Clin. Investig. 2010, 120, 457–471. [Google Scholar] [CrossRef] [PubMed]
- Diao, J.; Yang, X.; Song, X.; Chen, S.; He, Y.; Wang, Q.; Chen, G.; Luo, C.; Wu, X.; Zhang, Y. Exosomal Hsp70 mediates immunosuppressive activity of the myeloid-derived suppressor cells via phosphorylation of Stat3. Med. Oncol. 2015, 32, 453. [Google Scholar] [CrossRef] [PubMed]
- De Larco, J.E.; Wuertz, B.R.K.; Furcht, L.T. The Potential Role of Neutrophils in Promoting the Metastatic Phenotype of Tumors Releasing Interleukin-8. Clin. Cancer Res. 2004, 10, 4895–4900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klink, M.; Nowak, M.; Kielbik, M.; Bednarska, K.; Blus, E.; Szpakowski, M.; Szyllo, K.; Sulowska, Z. The Interaction of HspA1A with TLR2 and TLR4 in the Response of Neutrophils Induced by Ovarian Cancer Cells in Vitro. Cell Stress Chaperones 2012, 17, 661–674. [Google Scholar] [CrossRef] [Green Version]
- Linder, M.; Pogge von Strandmann, E. The Role of Extracellular HSP70 in the Function of Tumor-Associated Immune Cells. Cancers 2021, 13, 4721. [Google Scholar] [CrossRef]
- Gobbo, J.; Marcion, G.; Cordonnier, M.; Dias, A.M.M.; Pernet, N.; Hammann, A.; Richaud, S.; Mjahed, H.; Isambert, N.; Clausse, V. Restoring Anticancer Immune Response by Targeting Tumor-Derived Exosomes with a HSP70 Peptide Aptamer. J. Natl. Cancer Inst. 2015, 108, djv330. [Google Scholar] [CrossRef]
- Owji, H.; Nezafat, N.; Negahdaripour, M.; Hajiebrahimi, A.; Ghasemi, Y. A comprehensive review of signal peptides: Structure, roles, and applications. Eur. J. Cell Biol. 2018, 97, 422–441. [Google Scholar] [CrossRef]
- Mambula, S.S.; Stevenson, M.A.; Ogawa, K.; Calderwood, S.K. Mechanisms for Hsp70 secretion: Crossing membranes without a leader. Methods 2007, 43, 168–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arispe, N.; Doh, M.; Simakova, O.; Kurganov, B.; De Maio, A. Hsc70 and Hsp70 interact with phosphatidylserine on the surface of PC12 cells resulting in a decrease of viability. FASEB J. 2004, 18, 1636–1645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vega, V.L.; Rodríguez-Silva, M.; Frey, T.; Gehrmann, M.; Diaz, J.C.; Steinem, C.; Multhoff, G.; Arispe, N.; De Maio, A. Hsp70 translocates into the plasma membrane after stress and is released into the extracellular environment in a membrane-associated form that activates macrophages. J. Immunol. 2008, 180, 4299–4307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schilling, D.; Gehrmann, M.; Steinem, C.; De Maio, A.; Pockley, A.G.; Abend, M.; Molls, M.; Multhoff, G. Binding of Heat Shock Protein 70 to Extracellular Phosphatidylserine Promotes Killing of Normoxic and Hypoxic Tumor Cells. FASEB J. 2009, 23, 2467–2477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCallister, C.; Kdeiss, B.; Nikolaidis, N. Biochemical Characterization of the Interaction between HspA1A and Phospholipids. Cell Stress Chaperones 2016, 21, 41–53. [Google Scholar] [CrossRef] [Green Version]
- Broquet, A.H.; Thomas, G.; Masliah, J.; Trugnan, G.; Bachelet, M. Expression of the molecular chaperone Hsp70 in detergent-resistant microdomains correlates with its membrane delivery and release. J. Biol. Chem. 2003, 278, 21601–21606. [Google Scholar] [CrossRef] [Green Version]
- Bausero, M.A.; Gastpar, R.; Multhoff, G.; Asea, A. Alternative mechanism by which IFN-gamma enhances tumor recognition: Active release of heat shock protein 72. J. Immunol. 2005, 175, 2900–2912. [Google Scholar] [CrossRef] [Green Version]
- Gastpar, R.; Gehrmann, M.; Bausero, M.A.; Asea, A.; Gross, C.; Schroeder, J.A.; Multhoff, G. Heat shock protein 70 surface-positive tumor exosomes stimulate migratory and cytolytic activity of natural killer cells. Cancer Res. 2005, 65, 5238–5247. [Google Scholar] [CrossRef] [Green Version]
- Möller, A.; Lobb, R.J. The evolving translational potential of small extracellular vesicles in cancer. Nat. Rev. Cancer 2020, 20, 697–709. [Google Scholar] [CrossRef]
- Kanlikilicer, P.; Bayraktar, R.; Denizli, M.; Rashed, M.H.; Ivan, C.; Aslan, B.; Mitra, R.; Karagoz, K.; Bayraktar, E.; Zhang, X.; et al. Exosomal miRNA confers chemo resistance via targeting Cav1/p-gp/M2-type macrophage axis in ovarian cancer. EBioMedicine 2018, 38, 100–112. [Google Scholar] [CrossRef] [Green Version]
- Ren, J.; Ding, L.; Zhang, D.; Shi, G.; Xu, Q.; Shen, S.; Wang, Y.; Wang, T.; Hou, Y. Carcinoma-associated fibroblasts promote the stemness and chemoresistance of colorectal cancer by transferring exosomal lncRNA H19. Theragnostic 2018, 8, 3932–3948. [Google Scholar] [CrossRef]
- Zhao, S.; Mi, Y.; Guan, B.; Zheng, B.; Wei, P.; Gu, Y.; Zhang, Z.; Cai, S.; Xu, Y.; Li, X.; et al. Tumor-derived exosomal miR-934 induces macrophage M2 polarization to promote liver metastasis of colorectal cancer. J. Hematol. Oncol. 2020, 13, 156. [Google Scholar] [CrossRef]
- Hu, W.; Xu, Z.; Zhu, S.; Sun, W.; Wang, X.; Tan, C.; Zhang, Y.; Zhang, G.; Xu, Y.; Tang, J. Small extracellular vesicle-mediated Hsp70 intercellular delivery enhances breast cancer adriamycin resistance. Free Radic. Biol. Med. 2021, 164, 85–95. [Google Scholar] [CrossRef]
- Chanteloup, G.; Cordonnier, M.; Isambert, N.; Bertaut, A.; Hervieu, A.; Hennequin, A.; Luu, M.; Zanetta, S.; Coudert, B.; Bengrine, L.; et al. Monitoring HSP70 exosomes in cancer patients’ follow up: A clinical prospective pilot study. J. Extracell. Vesicles 2020, 9, 1766192. [Google Scholar] [CrossRef]
- Daigneault, M.; Preston, J.A.; Marriott, H.M.; Whyte, M.K.; Dockrell, D.H. The identification of markers of macrophage differentiation in PMA-stimulated THP-1 cells and monocyte-derived macrophages. PLoS ONE 2010, 5, e8668. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Shi, J.; Liu, W.; Zhang, K.; Zhao, H.; Zhang, H.; Zhang, Z. A simple, specific and “on-off” type MUC1 fluorescence aptasensor based on exosomes for detection of breast cancer. Sens. Actuators B Chem. 2018, 276, 552–559. [Google Scholar] [CrossRef]
- Kim, H.; Rhee, W.J. Exosome-mediated let7c-5p delivery for breast cancer therapeutic development. Biotechnol. Bioprocess Eng. 2020, 25, 513–520. [Google Scholar] [CrossRef]
- Wang, H.; He, D.; Wan, K.; Sheng, X.; Cheng, H.; Huang, J.; Zhou, X.; He, X.; Wang, K. In situ multiplex detection of serum exosomal microRNAs using an all-in-one biosensor for breast cancer diagnosis. Analyst 2020, 145, 3289–3296. [Google Scholar] [CrossRef]
- Duan, S.; Nordmeier, S.; Byrnes, A.E.; Buxton, I.L.O. Extracellular Vesicle-Mediated Purinergic Signaling Contributes to Host Microenvironment Plasticity and Metastasis in Triple Negative Breast Cancer. Int. J. Mol. Sci. 2021, 22, 597. [Google Scholar] [CrossRef]
- Minemura, H.; Takagi, K.; Sato, A.; Yamaguchi, M.; Hayashi, C.; Miki, Y.; Harada-Shoji, N.; Miyashita, M.; Sasano, H.; Suzuki, T. Isoforms of IDH in breast carcinoma: IDH2 as a potent prognostic factor associated with proliferation in estrogen-receptor positive cases. Breast Cancer 2021, 28, 915–926. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Takagi, K.; Sato, M.; Sato, A.; Miki, Y.; Onodera, Y.; Miyashita, M.; Sasano, H.; Suzuki, T. Androgens enhance the ability of intratumoral macrophages to promote breast cancer progression. Oncol. Rep. 2021, 46, 188. [Google Scholar] [CrossRef]
- Hayashi, C.; Takagi, K.; Sato, A.; Yamaguchi, M.; Minemura, H.; Miki, Y.; Harada-Shoji, N.; Miyashita, M.; Sasano, H.; Suzuki, T. D-2-hydroxyglutarate dehydrogenase in breast carcinoma as a potent prognostic marker associated with proliferation. Histol. Histopathol. 2021, 36, 1053–1062. [Google Scholar]
- Yamaguchi, M.; Takagi, K.; Sato, A.; Miki, Y.; Miyashita, M.; Sasano, H.; Suzuki, T. Rac1 activation in human breast carcinoma as a prognostic factor associated with therapeutic resistance. Breast Cancer 2020, 27, 919–928. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Takagi, K.; Narita, K.; Miki, Y.; Onodera, Y.; Miyashita, M.; Sasano, H.; Suzuki, T. Stromal CCL5 Promotes Breast Cancer Progression by Interacting with CCR3 in Tumor Cells. Int. J. Mol. Sci. 2021, 22, 1918. [Google Scholar] [CrossRef]
- Zhang, F.; Wang, H.; Wang, X.; Jiang, G.; Liu, H.; Zhang, G.; Wang, H.; Fang, R.; Bu, X.; Cai, S.; et al. TGF-β induces M2-like macrophage polarization via SNAIL-mediated suppression of a pro-inflammatory phenotype. Oncotarget 2016, 7, 52294–52306. [Google Scholar] [CrossRef] [Green Version]
- Jager, N.A.; Wallis de Vries, B.M.; Hillebrands, J.L.; Harlaar, N.J.; Tio, R.A.; Slart, R.H.; van Dam, G.M.; Boersma, H.H.; Zeebregts, C.J.; Westra, J. Distribution of Matrix Metalloproteinases in Human Atherosclerotic Carotid Plaques and Their Production by Smooth Muscle Cells and Macrophage Subsets. Mol. Imaging Biol. 2016, 18, 283–291. [Google Scholar] [CrossRef] [Green Version]
- Goldhirsch, A.; Wood, W.C.; Coates, A.S.; Gelber, R.D.; Thürlimann, B.; Senn, H.J.; Panel Members. Strategies for subtypes—Dealing with the diversity of breast cancer: Highlights of the St. Gallen International Expert Consensus on the Primary Therapy of Early Breast Cancer 2011. Ann. Oncol. 2011, 22, 1736–1747. [Google Scholar] [CrossRef]
- Ruffell, B.; Chang-Strachan, D.; Chan, V.; Rosenbusch, A.; Ho, C.M.; Pryer, N.; Daniel, D.; Hwang, E.S.; Rugo, H.S.; Coussens, L.M. Macrophage IL-10 blocks CD8+ T cell-dependent responses to chemotherapy by suppressing IL-12 expression in intratumoral dendritic cells. Cancer Cell 2014, 26, 623–637. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.H.; Wang, X.H.; Shi, J.J.; Yin, X.; Chen, C.; Chen, Y.; Wu, H.Y.; Jiong, S.; Sun, Q.; Zhang, M.; et al. Tumor-associated macrophages are associated with response to neoadjuvant chemotherapy. J. Cancer 2021, 12, 2886–2892. [Google Scholar] [CrossRef]
- Arole, V.; Nitta, H.; Wei, L.; Shen, T.; Parwani, A.V.; Li, Z. M2 tumor-associated macrophages play important role in predicting response to neoadjuvant chemotherapy in triple-negative breast carcinoma. Breast Cancer Res. Treat. 2021, 188, 37–42. [Google Scholar] [CrossRef]
- Yu, X.; Zhang, Q.; Zhang, X.; Han, Q.; Li, H.; Mao, Y.; Wang, X.; Guo, H.; Irwin, D.M.; Niu, G.; et al. Exosomes from Macrophages Exposed to Apoptotic Breast Cancer Cells Promote Breast Cancer Proliferation and Metastasis. J. Cancer 2019, 10, 2892–2906. [Google Scholar] [CrossRef] [Green Version]
- Lv, L.H.; Wan, Y.L.; Lin, Y.; Zhang, W.; Yang, M.; Li, G.L.; Lin, H.M.; Shang, C.Z.; Chen, Y.J.; Min, J. Anticancer drugs cause release of exosomes with heat shock proteins from human hepatocellular carcinoma cells that elicit effective natural killer cell antitumor responses in vitro. J. Biol. Chem. 2012, 287, 15874–15885. [Google Scholar] [CrossRef] [Green Version]
- Lopes, R.L.; Borges, T.J.; Araújo, J.F.; Pinho, N.G.; Bergamin, L.S.; Battastini, A.M.; Muraro, S.P.; Souza, A.P.; Zanin, R.F.; Bonorino, C. Extracellular mycobacterial DnaK polarizes macrophages to the M2-like phenotype. PLoS ONE 2014, 9, e113441. [Google Scholar] [CrossRef]
- Komarova, E.Y.; Marchenko, L.V.; Zhakhov, A.V.; Nikotina, A.D.; Aksenov, N.D.; Suezov, R.V.; Ischenko, A.M.; Margulis, B.A.; Guzhova, I.V. Extracellular Hsp70 Reduces the Pro-Tumor Capacity of Monocytes/Macrophages Co-Cultivated with Cancer Cells. Int. J. Mol. Sci. 2019, 21, 59. [Google Scholar] [CrossRef] [Green Version]
- Novak, M.L.; Koh, T.J. Macrophage phenotypes during tissue repair. J. Leukoc. Biol. 2013, 93, 875–881. [Google Scholar] [CrossRef] [Green Version]
- Shiraishi, D.; Fujiwara, Y.; Horlad, H.; Saito, Y.; Iriki, T.; Tsuboki, J.; Cheng, P.; Nakagata, N.; Mizuta, H.; Bekki, H.; et al. CD163 is required for protumoral activation of macrophages in human and murine sarcoma. Cancer Res. 2018, 78, 3255–3266. [Google Scholar] [CrossRef] [Green Version]
- Asea, A.; Rehli, M.; Kabingu, E.; Boch, J.A.; Bare, O.; Auron, P.E.; Stevenson, M.A.; Calderwood, S.K. Novel signal transduction pathway utilized by extracellular HSP70: Role of toll-like receptor (TLR) 2 and TLR4. J. Biol. Chem. 2002, 277, 15028–15034. [Google Scholar] [CrossRef] [Green Version]
- Somensi, N.; Brum, P.O.; de Miranda Ramos, V.; Gasparotto, J.; Zanotto-Filho, A.; Rostirolla, D.C.; da Silva Morrone, M.; Moreira, J.C.F.; Gelain, D.P. Extracellular HSP70 Activates ERK1/2, NF-kB and Pro-Inflammatory Gene Transcription Through Binding with RAGE in A549 Human Lung Cancer Cells. Cell. Physiol. Biochem. 2017, 42, 2507–2522. [Google Scholar] [CrossRef]
- Sharapova, T.N.; Romanova, E.A.; Ivanova, O.K.; Yashin, D.V.; Sashchenko, L.P. Hsp70 Interacts with the TREM-1 Receptor Expressed on Monocytes and Thereby Stimulates Generation of Cytotoxic Lymphocytes Active against MHC-Negative Tumor Cells. Int. J. Mol. Sci. 2021, 22, 6889. [Google Scholar] [CrossRef]
- Tang, L.; Han, X. The urokinase plasminogen activator system in breast cancer invasion and metastasis. Biomed. Pharmacother. 2013, 67, 179–182. [Google Scholar] [CrossRef]
- Falcone, D.J.; McCaffrey, T.A.; Haimovitz-Friedman, A.; Garcia, M. Transforming growth factor-beta 1 stimulates macrophage urokinase expression and release of matrix-bound basic fibroblast growth factor. J. Cell. Physiol. 1993, 155, 595–605. [Google Scholar] [CrossRef]
- Korc, M.; Friesel, R.E. The role of fibroblast growth factors in tumor growth. Curr. Cancer Drug Targets 2009, 9, 639–651. [Google Scholar] [CrossRef]
- Jafarzadeh, M.; Mousavizadeh, K.; Joghataei, M.T.; Bahremani, M.H.; Safa, M.; Asghari, S.M. A Fibroblast Growth Factor Antagonist Peptide Inhibits Breast Cancer in BALB/c Mice. Open Life Sci. 2018, 13, 348–354. [Google Scholar] [CrossRef] [PubMed]
- Lämmer, F.; Delbridge, C.; Würstle, S.; Neff, F.; Meyer, B.; Schlegel, J.; Kessel, K.A.; Schmid, T.E.; Schilling, D.; Combs, S.E. Correction: Cytosolic Hsp70 as a biomarker to predict clinical outcome in patients with glioblastoma. PLoS ONE 2021, 16, e0248612. [Google Scholar] [CrossRef]
- Lazaris, A.C.; Chatzigianni, E.B.; Panoussopoulos, D.; Tzimas, G.N.; Davaris, P.S.; Golematis, B.C. Proliferating cell nuclear antigen and heat shock protein 70 immunolocalization in invasive ductal breast cancer not otherwise specified. Breast Cancer Res. Treat. 1997, 43, 43–51. [Google Scholar] [CrossRef]
- Thanner, F.; Sütterlin, M.W.; Kapp, M.; Rieger, L.; Kristen, P.; Dietl, J.; Gassel, A.M.; Müller, T. Heat-shock protein 70 as a prognostic marker in node-negative breast cancer. Anticancer Res. 2003, 23, 1057–1062. [Google Scholar]
- Vargas-Roig, L.M.; Gago, F.E.; Tello, O.; Aznar, J.C.; Ciocca, D.R. Heat shock protein expression and drug resistance in breast cancer patients treated with induction chemotherapy. Int. J. Cancer 1998, 79, 468–475. [Google Scholar] [CrossRef]
- Dimas, D.T.; Perlepe, C.D.; Sergentanis, T.N.; Misitzis, I.; Kontzoglou, K.; Patsouris, E.; Kouraklis, G.; Psaltopoulou, T.; Nonni, A. The Prognostic Significance of Hsp70/Hsp90 Expression in Breast Cancer: A Systematic Review and Meta-analysis. Anticancer Res. 2018, 38, 1551–1562. [Google Scholar] [PubMed]
- Nadin, S.B.; Sottile, M.L.; Montt-Guevara, M.M.; Gauna, G.V.; Daguerre, P.; Leuzzi, M.; Gago, F.E.; Ibarra, J.; Cuello-Carrión, F.D.; Ciocca, D.R.; et al. Prognostic implication of HSPA (HSP70) in breast cancer patients treated with neoadjuvant anthracycline-based chemotherapy. Cell Stress Chaperones 2014, 19, 493–505. [Google Scholar] [CrossRef] [Green Version]
- Rashmi, R.; Santhosh Kumar, T.R.; Karunagaran, D. Human colon cancer cells differ in their sensitivity to curcumin-induced apoptosis and heat shock protects them by inhibiting the release of apoptosis-inducing factor and caspases. FEBS Lett. 2003, 538, 19–24. [Google Scholar] [CrossRef]
- Cai, M.B.; Wang, X.P.; Zhang, J.X.; Han, H.Q.; Liu, C.C.; Bei, J.X.; Peng, R.J.; Liang, Y.; Feng, Q.S.; Wang, H.Y.; et al. Expression of heat shock protein 70 in nasopharyngeal carcinomas: Different expression patterns correlate with distinct clinical prognosis. J. Transl. Med. 2012, 10, 96. [Google Scholar] [CrossRef] [Green Version]
- Rappa, F.; Unti, E.; Baiamonte, P.; Cappello, F.; Scibetta, N. Different immunohistochemical levels of Hsp60 and Hsp70 in a subset of brain tumors and putative role of Hsp60 in neuroepithelial tumorigenesis. Eur. J. Histochem. 2013, 57, e20. [Google Scholar] [CrossRef] [Green Version]
- Gao, W.W.; Xiao, R.Q.; Peng, B.L.; Xu, H.T.; Shen, H.F.; Huang, M.F.; Shi, T.T.; Yi, J.; Zhang, W.J.; Wu, X.N.; et al. Arginine methylation of HSP70 regulates retinoid acid-mediated RARβ2 gene activation. Proc. Natl. Acad. Sci. USA 2015, 112, E3327–E3336. [Google Scholar] [CrossRef] [Green Version]
- Rigg, R.A.; Healy, L.D.; Nowak, M.S.; Mallet, J.; Thierheimer, M.L.; Pang, J.; McCarty, O.J.; Aslan, J.E. Heat shock protein 70 regulates platelet integrin activation, granule secretion and aggregation. Am. J. Physiol. Cell Physiol. 2016, 310, C568–C575. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.Y.; Trevino, J.G.; Fang, B.L.; Riner, A.N.; Vudatha, V.; Zhang, G.H.; Li, Y.P. Patient-Derived Pancreatic Cancer Cells Induce C2C12 Myotube Atrophy by Releasing Hsp70 and Hsp90. Cells 2022, 11, 2756. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
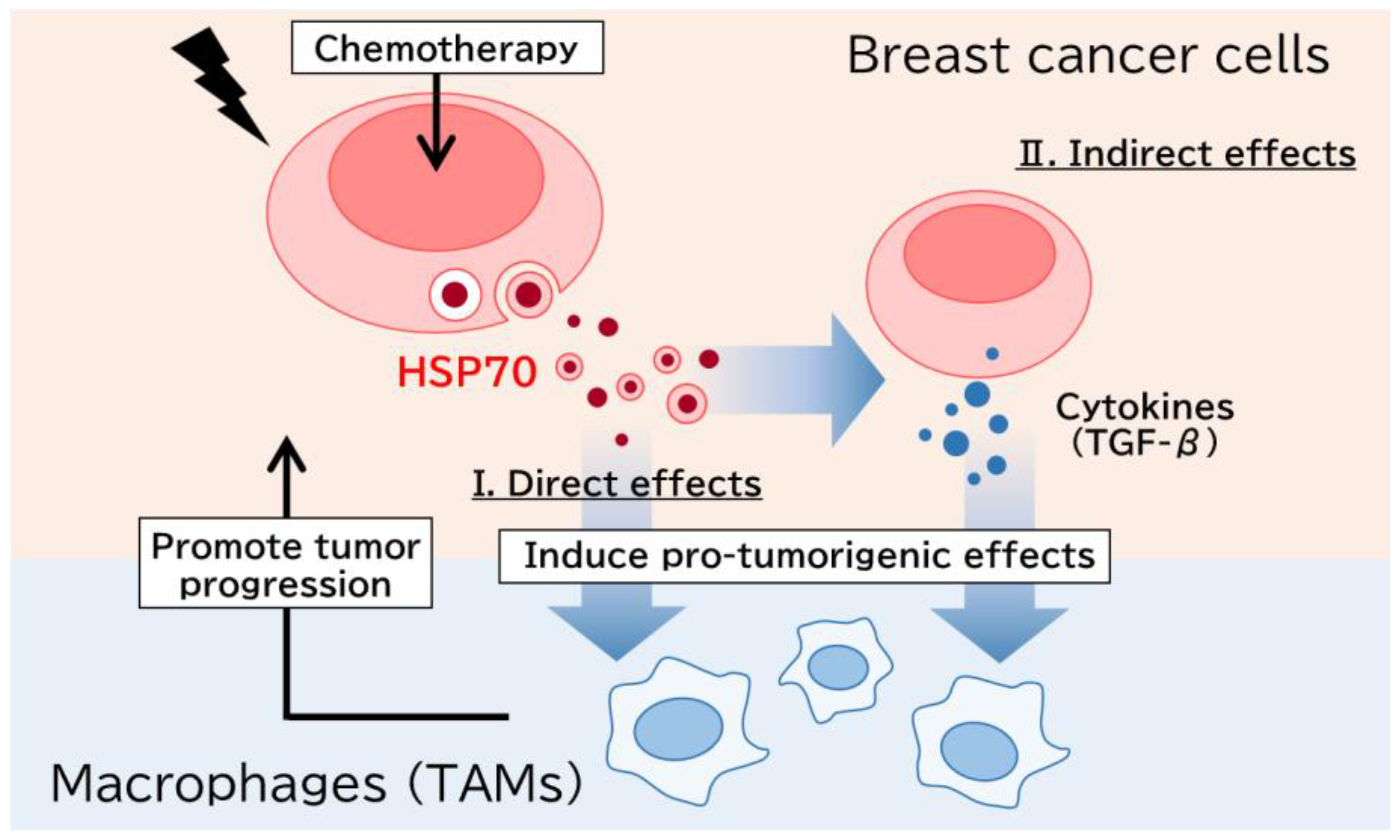{kind=link}
| Cytoplasmic HSP70 Status | p Value | |||
|---|---|---|---|---|
| Negative (n = 74) | Positive (n = 42) | |||
| Age * | 58 (29–87) | 55 (27–82) | 0.72 | |
| Menopause | ||||
| Premenopause | 26 | 15 | 0.95 | |
| Postmenopause | 48 | 27 | ||
| Stage | ||||
| I | 51 | 16 | 0.0027 | |
| II | 16 | 14 | ||
| III | 7 | 12 | ||
| Pathological T factor | ||||
| pT1 | 57 | 21 | 0.0029 | |
| pT2–4 | 17 | 21 | ||
| Lymph node metastasis | ||||
| Negative | 58 | 21 | 0.0016 | |
| Positive | 16 | 21 | ||
| Histological grade | ||||
| 1 (good) | 38 | 7 | <0.0001 | |
| 2 (intermediate) | 28 | 18 | ||
| 3 (poor) | 8 | 17 | ||
| ER | ||||
| Negative | 9 | 14 | 0.0060 | |
| Positive | 65 | 28 | ||
| PR | ||||
| Negative | 18 | 19 | 0.020 | |
| Positive | 56 | 23 | ||
| HER2 | ||||
| Negative | 62 | 37 | 0.53 | |
| Positive | 12 | 5 | ||
| Macrophage infiltration (CD163) | ||||
| Low | 53 | 24 | 0.11 | |
| High | 21 | 18 | ||
| Ki67 LI * (%) | 8.5 (1–53) | 18 (1–60) | 0.0016 | |
| Intrinsic subtype ** | ||||
| Luminal A | 44 | 14 | 0.0050 | |
| Luminal B | 21 | 14 | ||
| HER2 positive | 5 | 3 | ||
| Triple negative | 4 | 11 | ||
| Disease-Free Survival | Breast-Cancer-Specific Survival | |||||||
|---|---|---|---|---|---|---|---|---|
| Univariate | Multivariate | Univariate | Multivariate | |||||
| p * | Relative Risk ** | p | Relative Risk ** | p * | Relative Risk ** | p | Relative Risk ** | |
| Pathological T factor (pT2–4/pT1) | 0.0003 | 4.8 (2.1–11) | 0.015 | 4.3 (1.3–14) | 0.011 | 7.7 (1.6–37) | 0.47 | 2.1 (0.28–16) |
| Lymph node metastasis (Positive/Negative) | 0.0028 | 3.5 (1.5–7.8) | 0.63 | 1.3 (0.46–3.6) | 0.011 | 7.7 (1.6–37) | 0.13 | 4.3 (0.65–28) |
| Histological grade (3/1, 2) | 0.0070 | 3.1 (1.4–7.0) | 0.14 | 0.38 (0.11–1.4) | 0.0091 | 6.3 (1.6–25) | 0.68 | 0.67 (0.10–4.5) |
| ER (Negative/Positive) | 0.049 | 2.3 (1.0–5.4) | 0.20 | 0.46 (0.14–1.5) | 0.086 | 3.2 (0.85–12) | 0.46 | 0.58 (0.14–2.5) |
| PR (Negative/Positive) | 0.0030 | 3.6 (1.5–8.2) | 0.043 | 3.1 (1.0–9.1) | 0.0089 | 16.0 (2.0–128) | 0.058 | 9.4 (0.93–95) |
| HER2 (Positive/Negative) | 0.26 | 0.43 (0.10–1.8) | - | - | 0.67 | 0.63 (0.079–5.1) | - | - |
| Ki-67 LI *** | <0.0001 | 1.1 (1.0–1.1) | 0.12 | 1.0 (0.99–1.1) | 0.0012 | 1.1 (1.0–1.1) | 0.42 | 1.0 (0.97–1.1) |
| HSP70/Macrophage (++/others) | <0.0001 | 5.5 (2.5–12) | 0.012 | 4.4 (1.4–14) | 0.0020 | 8.1 (2.1–30) | 0.17 | 3.8 (0.55–27) |
| Disease-Free Survival | Breast-Cancer-Specific Survival | |||||||
|---|---|---|---|---|---|---|---|---|
| Univariate | Multivariate | Univariate | Multivariate | |||||
| p * | Relative Risk ** | p | Relative Risk ** | p * | Relative Risk ** | p | Relative Risk ** | |
| Pathological T factor (pT2–4/pT1) | 0.066 | 2.6 (0.94–7.2) | 0.0053 | 5.1 (1.6–16) | 0.22 | 2.7 (0.55–13) | - | - |
| Lymph node metastasis (Positive/Negative) | 0.24 | 1.8 (0.68–4.7) | - | - | 0.31 | 2.2 (0.47–11) | - | - |
| Histological grade (3/1, 2) | 0.29 | 1.6 (0.66–3.9) | - | - | 0.25 | 2.5 (0.56–9.0) | - | - |
| ER (Negative/Positive) | 0.40 | 1.5 (0.60–3.6) | - | - | 0.53 | 1.5 (0.41–5.7) | - | - |
| PR (Negative/Positive) | 0.059 | 2.7 (0.96–7.4) | 0.025 | 3.8 (1.2–12) | 0.077 | 6.5 (0.81–52) | 0.19 | 4.3 (0.50–38) |
| HER2 (Positive/Negative) | 0.092 | 0.28 (0.066–1.2) | 0.026 | 0.18 (0.038–0.81) | 0.35 | 0.37 (0.046–3.0) | - | |
| Ki-67 LI *** | 0.036 | 1.0 (1.0–1.1) | 0.69 | 0.99 (0.96–1.0) | 0.044 | 1.0 (1.0–1.1) | 0.41 | 1.0 (0.97–1.1) |
| HSP70/Macrophage (++/others) | 0.0012 | 4.3 (1.8–11) | 0.011 | 4.8 (1.4–16) | 0.034 | 4.2 (1.1–16) | 0.38 | 2.1 (0.41–10) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamaguchi-Tanaka, M.; Takagi, K.; Miki, Y.; Sato, A.; Iwabuchi, E.; Miyashita, M.; Suzuki, T. The Pro-Tumorigenic Role of Chemotherapy-Induced Extracellular HSP70 from Breast Cancer Cells via Intratumoral Macrophages. Cancers 2023, 15, 1903. https://doi.org/10.3390/cancers15061903
Yamaguchi-Tanaka M, Takagi K, Miki Y, Sato A, Iwabuchi E, Miyashita M, Suzuki T. The Pro-Tumorigenic Role of Chemotherapy-Induced Extracellular HSP70 from Breast Cancer Cells via Intratumoral Macrophages. Cancers. 2023; 15(6):1903. https://doi.org/10.3390/cancers15061903
Chicago/Turabian StyleYamaguchi-Tanaka, Mio, Kiyoshi Takagi, Yasuhiro Miki, Ai Sato, Erina Iwabuchi, Minoru Miyashita, and Takashi Suzuki. 2023. "The Pro-Tumorigenic Role of Chemotherapy-Induced Extracellular HSP70 from Breast Cancer Cells via Intratumoral Macrophages" Cancers 15, no. 6: 1903. https://doi.org/10.3390/cancers15061903