
The Prognostic Role and Significance of Dll4 and Toll-like Receptors in Cancer Development

 , ,
, ,  , ,
, ,  , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
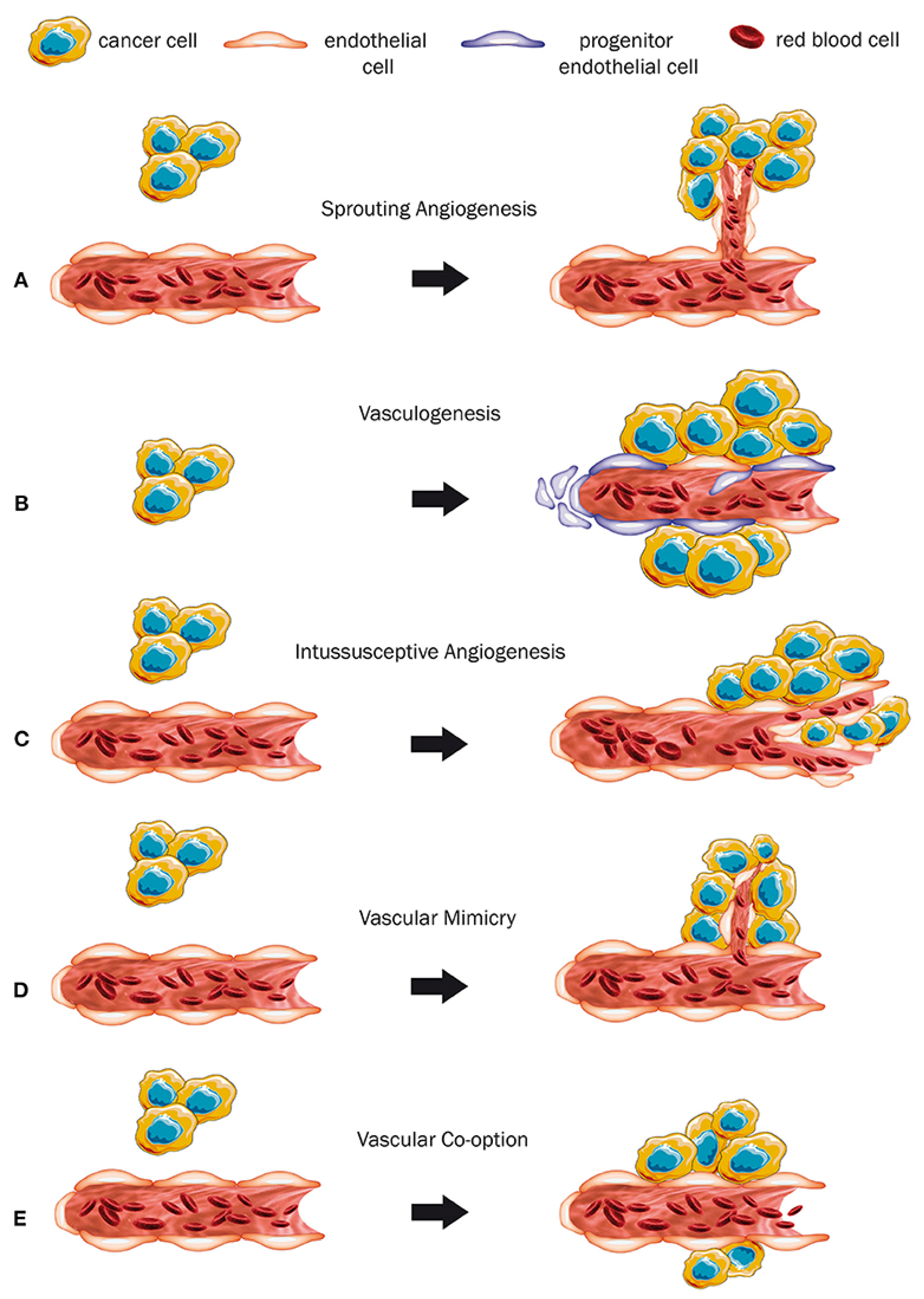
2. Angiogenesis in Cancer
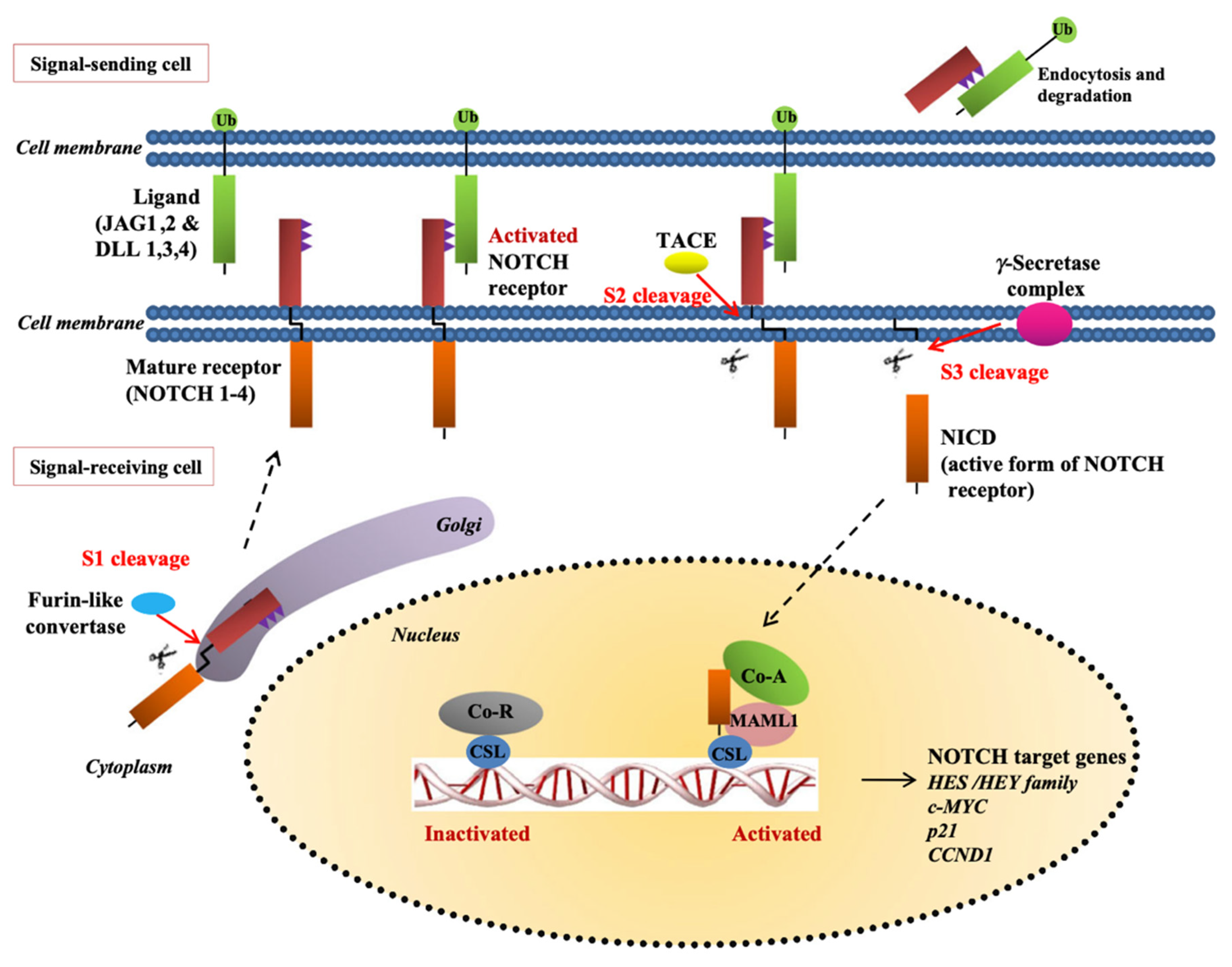
2.1. Angiogenesis via Notch Stimulation
2.2. Notch Ligands’ Role in Angiogenesis
3. Dll4 in Cancer Development
4. Cancer Stem Cells and Dll4 Expression in Endometrial Cancer
5. Immunotherapy and Tumor-Draining Lymph Nodes
6. Toll-like Receptors in Endometrial Cancer
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Folkman, J.; Judah, F. Tumor angiogenesis: Therapeutic implications. N. Engl. J. Med. 1971, 285, 1182–1186. [Google Scholar] [CrossRef] [PubMed]
- Folkman, J. Tumor angiogenesis factor. Cancer Res. 1974, 34, 2109–2113. [Google Scholar] [PubMed]
- Muthukkaruppan, V.R.; Kubai, L.; Auerbach, R. Tumor-induced neovascularization in the mouse eye. J. Natl. Cancer Inst. 1982, 69, 699–708. [Google Scholar] [CrossRef] [PubMed]
- Holmgren, L.; O’Reilly, M.S.; Folkman, J. Dormancy of micrometastases: Balanced proliferation and apoptosis in the presence of angiogenesis suppression. Nat. Med. 1995, 1, 149–153. [Google Scholar] [CrossRef]
- Denekamp, J. Angiogenesis, neovascular proliferation and vascular pathophysiology as targets for cancer therapy. Br. J. Radiol. 1993, 66, 181–196. [Google Scholar] [CrossRef] [PubMed]
- Dameron, K.M.; Volpert, O.V.; Tainsky, M.A.; Bouck, N. Control of Angiogenesis in Fibroblasts by p53 Regulation of Thrombospondin-1. Science 1994, 265, 1582–1584. [Google Scholar] [CrossRef] [PubMed]
- Dufraine, J.; Funahashi, Y.; Kitajewski, J. Notch signaling regulates tumor angiogenesis by diverse mechanisms. Oncogene 2008, 27, 5132–5137. [Google Scholar] [CrossRef] [Green Version]
- Hoey, T.; Yen, W.-C.; Axelrod, F.; Basi, J.; Donigian, L.; Dylla, S.; Fitch-Bruhns, M.; Lazetic, S.; Park, I.-K.; Sato, A.; et al. DLL4 Blockade Inhibits Tumor Growth and Reduces Tumor-Initiating Cell Frequency. Cell Stem Cell 2009, 5, 168–177. [Google Scholar] [CrossRef] [Green Version]
- Gurney, A.; Hoey, T. Anti-DLL4, a cancer therapeutic with multiple mechanisms of action. Vasc. Cell 2011, 3, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribatti, D.; Crivellato, E. “Sprouting angiogenesis”, a reappraisal. Dev. Biol. 2012, 372, 157–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, L.; Rizzo, P.; Osipo, C.; Pannuti, A.; Wyatt, D.; Cheung, L.W.-K.; Sonenshein, G.; Osborne, A.B.; Miele, L. Notch-1 activates estrogen receptor-α-dependent transcription via IKKα in breast cancer cells. Oncogene 2009, 29, 201–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klinakis, A.; Szabolcs, M.; Politi, K.; Kiaris, H.; Artavanis-Tsakonas, S.; Efstratiadis, A. Myc is a Notch1 transcriptional target and a requisite for Notch1-induced mammary tumorigenesis in mice. Proc. Natl. Acad. Sci. USA 2006, 103, 9262–9267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schepers, G.E.; Teasdale, R.D.; Koopman, P. Twenty Pairs of Sox: Extent, Homology, and Nomenclature of the Mouse and Human Sox Transcription Factor Gene Families. Dev. Cell 2002, 3, 167–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karamboulas, C.; Ailles, L. Developmental signaling pathways in cancer stem cells of solid tumors. Biochim. Biophys. Acta (BBA) Gen. Subj. 2013, 1830, 2481–2495. [Google Scholar] [CrossRef]
- Wang, K.; Zhang, Q.; Li, D.; Ching, K.; Zhang, C.; Zheng, X.; Ozeck, M.; Shi, S.; Li, X.; Wang, H.; et al. PEST Domain Mutations in Notch Receptors Comprise an Oncogenic Driver Segment in Triple-Negative Breast Cancer Sensitive to a γ-Secretase Inhibitor. Clin. Cancer Res. 2015, 21, 1487–1496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shutter, J.R.; Scully, S.; Fan, W.; Richards, W.G.; Kitajewski, J.; Deblandre, G.A.; Kintner, C.R.; Stark, K.L. Dll4, a novel Notch ligand expressed in arterial endothelium. Genes Dev. 2000, 14, 1313–1318. [Google Scholar] [CrossRef] [PubMed]
- Hellström, M.; Phng, L.-K.; Gerhardt, H. VEGF and Notch Signaling. Cell Adh. Migr. 2007, 1, 133–136. [Google Scholar] [CrossRef] [PubMed]
- Benedito, R.; Hellström, M. Notch as a hub for signaling in angiogenesis. Exp. Cell Res. 2013, 319, 1281–1288. [Google Scholar] [CrossRef] [PubMed]
- Kuhnert, F.; Chen, G.; Coetzee, S.; Thambi, N.; Hickey, C.; Shan, J.; Kovalenko, P.; Noguera-Troise, I.; Smith, E.; Fairhurst, J.L.; et al. Dll4 Blockade in Stromal Cells Mediates Antitumor Effects in Preclinical Models of Ovarian Cancer. Cancer Res. 2015, 75, 4086–4096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, J.; Ye, X.; Fan, F.; Xia, L.; Bhattacharya, R.; Bellister, S.; Tozzi, F.; Sceusi, E.; Zhou, Y.; Tachibana, I.; et al. Endothelial Cells Promote the Colorectal Cancer Stem Cell Phenotype through a Soluble Form of Jagged-1. Cancer Cell 2013, 23, 171–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, T.S.; Costello, M.A.; Talsma, C.E.; Flack, C.G.; Crowley, J.G.; Hamm, L.L.; He, X.; Umper, S.L.H.-J.; Heth, J.A.; Muraszko, K.M.; et al. Endothelial Cells Create a Stem Cell Niche in Glioblastoma by Providing NOTCH Ligands That Nurture Self-Renewal of Cancer Stem-Like Cells. Cancer Res. 2011, 71, 6061–6072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Indraccolo, S.; Minuzzo, S.A.; Masiero, M.; Amador, A. Ligand-driven activation of the Notch pathway in T-all and solid tumors: Why Not(ch)? Cell Cycle 2010, 9, 80–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonoshita, M.; Aoki, M.; Fuwa, H.; Aoki, K.; Hosogi, H.; Sakai, Y.; Hashida, H.; Takabayashi, A.; Sasaki, M.; Robine, S.; et al. Suppression of Colon Cancer Metastasis by Aes through Inhibition of Notch Signaling. Cancer Cell 2011, 19, 125–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patenaude, A.; Woerher, S.; Umlandt, P.; Wong, F.; Ibrahim, R.; Kyle, A.; Unger, S.; Fuller, M.; Parker, J.; Minchinton, A.; et al. A novel population of local pericyte precursor cells in tumor stroma that require Notch signaling for differentiation. Microvasc. Res. 2015, 101, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Benedito, R.; Roca, C.; Sörensen, I.; Adams, S.; Gossler, A.; Fruttiger, M.; Adams, R.H. The Notch Ligands Dll4 and Jagged1 Have Opposing Effects on Angiogenesis. Cell 2009, 137, 1124–1135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boareto, M.; Jolly, M.K.; Lu, M.; Onuchic, J.N.; Clementi, C.; Ben-Jacob, E. Jagged–Delta asymmetry in Notch signaling can give rise to a Sender/Receiver hybrid phenotype. Proc. Natl. Acad. Sci. USA 2015, 112, E402–E409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedrosa, A.-R.; Graça, J.L.; Carvalho, S.; Peleteiro, M.C.; Duarte, A.; Trindade, A. Notch signaling dynamics in the adult healthy prostate and in prostatic tumor development. Prostate 2015, 76, 80–96. [Google Scholar] [CrossRef] [PubMed]
- Ghajar, C.M.; Peinado, H.; Mori, H.; Matei, I.R.; Evason, K.J.; Brazier, H.; Almeida, D.; Koller, A.; Hajjar, K.A.; Stainier, D.Y.; et al. The perivascular niche regulates breast tumour dormancy. Nat. Cell Biol. 2013, 15, 807–817. [Google Scholar] [CrossRef]
- Wang, H.; Huang, X.; Zhang, J.; Shao, N.; Chen, L.O.; Ma, D.; Ji, C. The expression of VEGF and Dll4/Notch pathway molecules in ovarian cancer. Clin. Chim. Acta 2014, 436, 243–248. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Fan, F.; Wang, A.; Zheng, S.; Lu, Y. Dll4-Notch signaling in regulation of tumor angiogenesis. J. Cancer Res. Clin. Oncol. 2013, 140, 525–536. [Google Scholar] [CrossRef] [PubMed]
- Miao, Z.-F.; Xu, H.; Xu, H.-M.; Wang, Z.; Zhao, T.-T.; Song, Y.-X.; Xu, Y.-Y. DLL4 overexpression increases gastric cancer stem/progenitor cell self-renewal ability and correlates with poor clinical outcome via Notch-1 signaling pathway activation. Cancer Med. 2016, 6, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Lu, C.; Dong, H.H.; Huang, J.; Shen, D.-Y.; Stone, R.L.; Nick, A.M.; Shahzad, M.M.K.; Mora, E.; Jennings, N.B.; et al. Biological Roles of the Delta Family Notch Ligand Dll4 in Tumor and Endothelial Cells in Ovarian Cancer. Cancer Res. 2011, 71, 6030–6039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yen, W.-C.; Fischer, M.M.; Shah, J.; Wei, J.; Cain, J.; Yeung, P.; Beviglia, L.; Cancilla, B.; Kapoun, A.; Lewicki, J.; et al. Anti-DLL4 (demcizumab) inhibits tumor growth and reduces cancer stem cell frequency in patient-derived ovarian cancer xenografts. Cancer Res. 2013, 73, 3725. [Google Scholar] [CrossRef]
- Bao, H.; Chen, X.; Thomas, S.; Shi, X.; Strout, P.; Hurt, E.; Cho, S.; Burke, P.A.; Dar, M.M. Evaluation of anti-cancer stem cell activity of the anti-DLL4 antibody MEDI0639 in a phase I clinical trial of SCLC. J. Clin. Oncol. 2016, 34, e20093. [Google Scholar] [CrossRef]
- Mendonça, L.; Trindade, A.; Carvalho, C.; Correia, J.; Badenes, M.; Gigante, J.; Duarte, A. Metastasis is impaired by endothelial-specific Dll4 loss-of-function through inhibition of epithelial-to-mesenchymal transition and reduction of cancer stem cells and circulating tumor cells. Clin. Exp. Metastasis 2019, 36, 365–380. [Google Scholar] [CrossRef] [PubMed]
- Yen, W.-C.; Fischer, M.M.; Hynes, M.; Wu, J.; Kim, E.; Beviglia, L.; Yeung, V.P.; Song, X.; Kapoun, A.M.; Lewicki, J.; et al. Anti-DLL4 Has Broad Spectrum Activity in Pancreatic Cancer Dependent on Targeting DLL4-Notch Signaling in Both Tumor and Vasculature Cells. Clin. Cancer Res. 2012, 18, 5374–5386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zohny, S.F.; Zamzami, M.A.; Al-Malki, A.L.; Trabulsi, N.H. Highly Expressed DLL4 and JAG1: Their Role in Incidence of Breast Cancer Metastasis. Arch. Med. Res. 2020, 51, 145–152. [Google Scholar] [CrossRef]
- Kato, K. Endometrial cancer stem cells: A new target for cancer therapy. Anticancer Res. 2012, 32, 2283–2293. [Google Scholar] [PubMed]
- Fasoulakis, Z.; Galazios, G.; Koukourakis, M.; Nikolettos, N.; Giatromanolaki, A.; Koutlaki, N.; NKontomanolis, E. Expression of Delta Like Ligand 4 (DLL4) in endometrial carcinomas and tumor vasculature. J. Buon. 2021, 26, 1327–1332. [Google Scholar] [PubMed]
- Mazella, J.; Liang, S.; Tseng, L. Expression of Delta-Like Protein 4 in the Human Endometrium. Endocrinology 2007, 149, 15–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badenes, M.; Trindade, A.; Pissarra, H.; Lopes-Da-Costa, L.; Duarte, A. Delta-like 4/Notch signaling promotes Apc Min/+ tumor initiation through angiogenic and non-angiogenic related mechanisms. BMC Cancer 2017, 17, 50. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Duan, Y.; Ye, X.; Hu, L.; Xu, T.; Tong, L.; Yu, M. Stable silencing of dll4 gene suppresses the growth and metastasis of esophagus cancer cells by attenuating Akt phosphorylation. Oncol. Rep. 2018, 40, 495–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ElSahwi, K.S.; Bellone, S.; Cocco, E.; Cargnelutti, M.; Casagrande, F.; Bellone, M.; Abukhalaf, M.; Buza, N.; Tavassoli, F.A.; Hui, P.; et al. In vitro activity of pertuzumab in combination with trastuzumab in uterine serous papillary adenocarcinoma. Br. J. Cancer 2009, 102, 134–143. [Google Scholar] [CrossRef]
- Jenkins, D.W.; Ross, S.; Veldman-Jones, M.; Foltz, I.N.; Clavette, B.C.; Manchulenko, K.; Eberlein, C.; Kendrew, J.; Petteruti, P.; Cho, S.; et al. MEDI0639: A Novel Therapeutic Antibody Targeting Dll4 Modulates Endothelial Cell Function and Angiogenesis In Vivo. Mol. Cancer Ther. 2012, 11, 1650–1660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fatima, I.; Barman, S.; Rai, R.; Thiel, K.W.; Chandra, V. Targeting Wnt Signaling in Endometrial Cancer. Cancers 2021, 13, 2351. [Google Scholar] [CrossRef] [PubMed]
- Yeom, D.-H.; Lee, Y.-S.; Ryu, I.; Lee, S.; Sung, B.; Lee, H.-B.; Kim, D.; Ahn, J.-H.; Ha, E.; Choi, Y.-S.; et al. ABL001, a Bispecific Antibody Targeting VEGF and DLL4, with Chemotherapy, Synergistically Inhibits Tumor Progression in Xenograft Models. Int. J. Mol. Sci. 2020, 22, 241. [Google Scholar] [CrossRef]
- Chiorean, E.G.; LoRusso, P.; Strother, R.M.; Diamond, J.R.; Younger, A.; Messersmith, W.A.; Adriaens, L.; Liu, L.; Kao, R.J.; DoCioccio, A.T.; et al. A phase I first-in-human study of enoticumab (REGN421), a fully human delta-like ligand 4 (Dll4) monoclonal antibody in patients with advanced solid tumors. Clin. Cancer Res. 2015, 21, 2695–2703. [Google Scholar]
- Frimer, M.; Hou, J.Y.; McAndrew, T.C.; Goldberg, G.L.; Shahabi, S. The Clinical Relevance of Rising CA-125 Levels within the Normal Range in Patients with Uterine Papillary Serous Cancer. Reprod. Sci. 2012, 20, 449–455. [Google Scholar] [CrossRef] [Green Version]
- Makker, V.; Taylor, M.H.; Aghajanian, C.; Oaknin, A.; Mier, J.; Cohn, A.L.; Romeo, M.; Bratos, R.; Brose, M.S.; DiSimone, C.; et al. Lenvatinib Plus Pembrolizumab in Patients with Advanced Endometrial. Cancer J. Clin. Oncol. 2020, 38, 2981–2992. [Google Scholar]
- Makker, V.; Rasco, D.; Vogelzang, N.J.; Brose, M.S.; Cohn, A.L.; Mier, J.; Di Simone, C.; Hyman, D.M.; Stepan, D.E.; Dutcus, C.E.; et al. Lenvatinib plus pembrolizumab in patients with advanced endometrial cancer: An interim analysis of a multicentre, open-label, single-arm, phase 2 trial. Lancet Oncol. 2019, 20, 711–718. [Google Scholar] [CrossRef]
- Arora, S.; Balasubramaniam, S.; Zhang, W.; Zhang, L.; Sridhara, R.; Spillman, D.; Mathai, J.P.; Scott, B.; Golding, S.J.; Coory, M.; et al. FDA Approval Summary: Pembrolizumab plus Lenvatinib for Endometrial Carcinoma, a Collaborative International Review under Project Orbis. Clin. Cancer Res. 2020, 26, 5062–5067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- NIHUS National Library of Medicine. Available online: https://www.clinicaltrials.gov/ct2/results?cond=Endometrial+Cancer&term=serous&cntry=&state=&city=&dist (accessed on 31 December 2020).
- Sheng, X.; Yan, X.; Wang, L.; Shi, Y.-X.; Yao, X.; Luo, H.; Shi, B.; Liu, J.-Y.; He, Z.; Yu, G.; et al. Open-label, Multicenter, Phase II Study of RC48-ADC, a HER2-Targeting Antibody–Drug Conjugate, in Patients with Locally Advanced or Metastatic Urothelial Carcinoma. Clin. Cancer Res. 2020, 27, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Banerji, U.; van Herpen, C.M.L.; Saura, C.; Thistlethwaite, F.; Lord, S.; Moreno, V.; Macpherson, I.R.; Boni, V.; Rolfo, C.; de Vries, E.G.E.; et al. Trastuzumab duocarmazine in locally advanced and metastatic solid tumours and HER2-expressing breast cancer: A phase 1 dose-escalation and dose-expansion study. Lancet Oncol. 2019, 20, 1124–1135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanova, E.; Kuraguchi, M.; Xu, M.; Portell, A.; Taus, L.; Diala, I.; Lalani, A.S.; Choi, J.; Chambers, E.S.; Li, S.; et al. Use of ExVivo Patient-Derived Tumor Organotypic Spheroids to Identify Combination Therapies for HER2 Mutant Non–Small Cell Lung Cancer. Clin. Cancer Res. 2020, 26, 2393–2403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santin, A.D.; Filiaci, V.; Bellone, S.; Ratner, E.S.; Mathews, C.A.; Cantuaria, G.; Gunderson, C.C.; Rutledge, T.; Buttin, B.M.; Lankes, H.A.; et al. Phase II evaluation of copanlisib, a selective inhibitor of Pi3kca, in patients with persistent or recurrent endometrial carcinoma harboring PIK3CA hotspot mutations: An NRG Oncology study (NRG-GY008). Gynecol. Oncol. Rep. 2020, 31, 100532. [Google Scholar]
- Bonazzoli, E.; Cocco, E.; Lopez, S.; Bellone, S.; Zammataro, L.; Bianchi, A.; Manzano, A.; Yadav, G.; Manara, P.; Perrone, E.; et al. PI3K oncogenic mutations mediate resistance to afatinib in HER2/neu overexpressing gynecological cancers. Gynecol. Oncol. 2019, 153, 158–164. [Google Scholar] [PubMed]
- Lopez, S.; Cocco, E.; Black, J.; Bellone, S.; Bonazzoli, E.; Predolini, F.; Ferrari, F.; Schwab, C.L.; English, D.P.; Ratner, E.; et al. Dual HER2/PIK3CA Targeting Overcomes Single-Agent Acquired Resistance in HER2-Amplified Uterine Serous Carcinoma Cell Lines In Vitro and In Vivo. Mol. Cancer Ther. 2015, 14, 2519–2526. [Google Scholar] [PubMed] [Green Version]
- Zaman, G.J.; De Roos, J.A.; Libouban, M.A.; Prinsen, M.B.; De Man, J.; Buijsman, R.C.; Uitdehaag, J.C. TTK Inhibitors as a Targeted Therapy for CTNNB1 (β-catenin) Mutant Cancers. Mol. Cancer Ther. 2017, 16, 2609–2617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webster, E.M.; Zeybek, B.; Tymon-Rosario, J.; Santin, A.D. Sacituzumab govitecan: A promising antibody-drug conjugate for the treatment of poorly differentiated endometrial cancer. Oncoscience 2020, 7, 68–69. [Google Scholar] [CrossRef] [PubMed]
- Romero, I.; Rubio, M.J.; Medina, M.; Matias-Guiu, X.; Santacana, M.; Schoenenberger, J.-A.; Guerra, E.M.; Cortés, A.; Minig, L.; Coronado, P.; et al. An olaparib window-of-opportunity trial in patients with early-stage endometrial carcinoma: POLEN study. Gynecol. Oncol. 2020, 159, 721–731. [Google Scholar] [CrossRef] [PubMed]
- Molenkamp, B.G.; Sluijter, B.J.; van Leeuwen, P.A.; Santegoets, S.J.; Meijer, S.; Wijnands, P.G.; Haanen, J.B.; van den Eertwegh, A.J.; Scheper, R.J.; de Gruijl, T.D. Cancer therapy: Clinical local administration of PF-3512676 CpG-B Instigates T CD8 T-cell reactivity in melanoma patients. Clin. Cancer Res. 2008, 14, 4532–4543. [Google Scholar] [PubMed] [Green Version]
- Chow, M.T.; Ozga, A.J.; Servis, R.L.; Frederick, D.T.; Lo, J.A.; Fisher, D.E.; Freeman, G.J.; Boland, G.M.; Luster, A.D. Intratumoral Activity of the CXCR3 Chemokine System Is Required for the Efficacy of Anti-PD-1 Therapy. Immunity 2019, 50, 1498–1512.e5. [Google Scholar] [CrossRef] [PubMed]
- Ellmark, P.; Mangsbo, S.M.; Furebring, C.; Norlén, P.; Tötterman, T.H. Tumor-directed immunotherapy can generate tumor-specific T cell responses through localized co-stimulation. Cancer Immunol. Immunother. 2016, 66, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Paulete, A.R.; Teijeira, Á.; Quetglas, J.I.; Rodríguez-Ruiz, M.E.; Sánchez-Arráez, Á.; Labiano, S.; Etxeberria, I.; Az-pilikueta, A.; Bolaños, E.; Ballesteros-Briones, M.C.; et al. Intratumoral Immunotherapy with XCL1 and sFlt3L Encoded in Recombinant Semliki Forest Virus–Derived Vectors Fosters Dendritic Cell–Mediated T-cell Cross-Priming. Cancer Res. 2018, 78, 6643–6654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandin, L.C.; Orlova, A.; Gustafsson, E.; Ellmark, P.; Tolmachev, V.; Tötterman, T.H.; Mangsbo, S.M. Locally Delivered CD40 Agonist Antibody Accumulates in Secondary Lymphoid Organs and Eradicates Experimental Disseminated Bladder Cancer. Cancer Immunol. Res. 2013, 2, 80–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandin, L.C.; Tötterman, T.H.; Mangsbo, S.M. Local immunotherapy based on agonistic CD40 antibodies effectively inhibits experimental bladder cancer. OncoImmunology 2014, 3, e27400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Hooren, L.; Sandin, L.C.; Moskalev, I.; Ellmark, P.; Dimberg, A.; Black, P.; Tötterman, T.H.; Mangsbo, S.M. Local checkpoint inhibition of CTLA-4 as a monotherapy or in combination with anti-PD1 prevents the growth of murine bladder cancer. Eur. J. Immunol. 2017, 47, 385–393. [Google Scholar] [PubMed]
- Fransen, M.F.; van der Sluis, T.; Ossendorp, F.; Arens, R.; Melief, C.J. Controlled Local Delivery of CTLA-4 Blocking Antibody Induces CD8+ T-Cell–Dependent Tumor Eradication and Decreases Risk of Toxic Side Effects. Clin. Cancer Res. 2013, 19, 5381–5389. [Google Scholar] [CrossRef] [Green Version]
- Francis, D.M.; Manspeaker, M.P.; Schudel, A.; Sestito, L.F.; O’Melia, M.J.; Kissick, H.T.; Pollack, B.P.; Waller, E.K.; Thomas, S.N. Blockade of immune checkpoints in lymph nodes through locoregional delivery augments cancer immunotherapy. Sci. Transl. Med. 2020, 12, eaay3575. [Google Scholar] [CrossRef]
- Koster, B.D.; van den Hout, M.F.C.M.; Sluijter, B.J.R.; Molenkamp, B.G.; Vuylsteke, R.J.C.L.M.; Baars, A.; van Leeuwen, P.A.M.; Scheper, R.J.; Petrousjka van den Tol, M. Local adjuvant treatment with low-dose CpG-B offers durable protection against disease recurrence in clinical stage I-II melanoma: Data from two randomized phase II trials. Clin. Cancer Res. 2017, 23, 5679–5686. [Google Scholar] [PubMed] [Green Version]
- Notohardjo, J.; Van Pul, K.; Stam, A.; Chondronasiou, D.; Lougheed, S.; Tol, P.V.D.; Jooss, K.; Vuylsteke, R.; Eertwegh, A.V.D.; De Gruijl, T. The role of tumor-draining lymph nodes in the tuning of systemic T cell immunity by CTLA-4 blockade is revealed by local delivery of tremelimumab in early-stage melanoma: Data from a Phase-I trial. J. Immunother. Cancer 2020, 8, A336. [Google Scholar] [CrossRef]
- Faries, M.B.; Thompson, J.F.; Cochran, A.J.; Andtbacka, R.H.; Mozzillo, N.; Zager, J.S.; Jahkola, T.; Bowles, T.L.; Testori, A.; Beitsch, P.D.; et al. Completion Dissection or Observation for Sentinel-Node Metastasis in Melanoma. N. Engl. J. Med. 2017, 376, 2211–2222. [Google Scholar] [CrossRef] [PubMed]
- Donker, M.; van Tienhoven, G.; Straver, M.E.; Meijnen, P.; van de Velde, C.J.H.; Mansel, R.E.; Cataliotti, L.; Westenberg, A.H.; Klinkenbijl, J.H.G.; Orzalesi, L.; et al. Radiotherapy or surgery of the axilla after a positive sentinel node in breast cancer (EORTC 10981-22023 AMAROS): A randomised, multicentre, open-label, phase 3 non-inferiority trial. Lancet Oncol. 2014, 15, 1303–1310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Campos, J.; Gallotta, M.; Gong, M.; Crain, C.; Naik, E.; Coffman, R.L.; Guiducci, C. Intratumoral injection of a CpG oligonucleotide reverts resistance to PD-1 blockade by expanding multifunctional CD8 + T cells. Proc. Natl. Acad. Sci. USA 2016, 113, E7240–E7249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Middleton, M.R.; Hoeller, C.; Michielin, O.; Robert, C.; Caramella, C.; Öhrling, K.; Hauschild, A. Intratumoural immunotherapies for unresectable and metastatic melanoma: Current status and future perspectives. Br. J. Cancer 2020, 123, 885–897. [Google Scholar] [CrossRef] [PubMed]
- Andtbacka, R.H.I.; Ross, M.; Puzanov, I.; Milhem, M.; Collichio, F.; Delman, K.A.; Amatruda, T.; Zager, J.S.; Cranmer, L.; Hsueh, E.; et al. Patterns of Clinical Response with Talimogene Laherparepvec (T-VEC) in Patients with Melanoma Treated in the OPTiM Phase III Clinical Trial. Ann. Surg. Oncol. 2016, 23, 4169–4177. [Google Scholar] [CrossRef] [Green Version]
- Kaufman, H.L.; Amatruda, T.; Reid, T.; Gonzalez, R.; Glaspy, J.; Whitman, E.; Harrington, K.; Nemunaitis, J.; Zloza, A.; Wolf, M.; et al. Systemic versus local responses in melanoma patients treated with talimogene laherparepvec from a multi-institutional phase II study. J. Immunother. Cancer 2016, 4, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gogas, H.; Samoylenko, I.; Schadendorf, D.; Gutzmer, R.; Grob, J.; Sacco, J.; Gorski, K.; Anderson, A.; Liu, C.; Malvehy, J. Talimogene laherparepvec (T-VEC) treatment increases intratumoral effector T-cell and natural killer (NK) cell density in noninjected tumors in patients (pts) with stage IIIB–IVM1c melanoma: Evidence for systemic effects in a phase II, single-arm study. Ann. Oncol. 2018, 29, viii443. [Google Scholar] [CrossRef]
- Ribas, A.; Dummer, R.; Puzanov, I.; VanderWalde, A.; Andtbacka, R.H.I.; Michielin, O.; Olszanski, A.J.; Malvehy, J.; Cebon, J.; Fernandez, E.; et al. Oncolytic virotherapy promotes intratumoral T cell infiltration and improves anti-PD-1 immunotherapy. Cell 2017, 170, 1109–1119.e10. [Google Scholar]
- Marabelle, A.; Andtbacka, R.; Harrington, K.; Melero, I.; Leidner, R.; de Baere, T.; Robert, C.; Ascierto, P.A.; Baurain, J.-F.; Imperiale, M.; et al. Starting the fight in the tumor: Expert recommendations for the development of human intratumoral immunotherapy (HIT-IT). Ann. Oncol. 2018, 29, 2163–2174. [Google Scholar] [CrossRef] [PubMed]
- Takeda, K.; Kaisho, T.; Akira, S. Toll-like receptors. Annu. Rev. Immunol. 2003, 21, 335–376. [Google Scholar] [PubMed]
- Takeda, K.; Akira, S. Toll-like receptors in innate immunity. Int. Immunol. 2005, 17, 135–145. [Google Scholar]
- Kawai, T.; Akira, S. The role of pattern-recognition receptors in innate immunity: Update on Toll-like receptors. Nat. Immunol. 2010, 11, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Janeway, C.A., Jr.; Medzhitov, R. Innate immune recognition. Annu. Rev. Immunol. 2002, 20, 197–216. [Google Scholar] [PubMed] [Green Version]
- Behzadi, P.; García-Perdomo, H.A.; Karpiński, T.M. Toll-Like Receptors: General Molecular and Structural Biology. J. Immunol. Res. 2021, 2021, 9914854. [Google Scholar] [CrossRef]
- Kanzler, H.; Barrat, F.J.; Hessel, E.M.; Coffman, R.L. Therapeutic targeting of innate immunity with Toll-like receptor agonists and antagonists. Nat. Med. 2007, 13, 552–559. [Google Scholar] [CrossRef] [PubMed]
- Zou, X.; Guo, B.; Ling, Q.; Mo, Z. Toll-Like Receptors Serve as Biomarkers for Early Diagnosis and Prognosis Assessment of Kidney Renal Clear Cell Carcinoma by Influencing the Immune Microenvironment: Comprehensive Bioinformatics Analysis Combined with Experimental Validation. Front. Mol. Biosci. 2022, 9, 832238. [Google Scholar] [CrossRef]
- Verstrepen, L.; Bekaert, T.; Chau, T.L.; Tavernier, J.; Chariot, A.; Beyaert, R. TLR-4, IL-1R and TNF-R signaling to NF-kappaB: Variations on a common theme. Cell Mol. Life Sci. 2008, 65, 2964–2978. [Google Scholar]
- Aggarwal, B.B. Signalling pathways of the TNF superfamily: A double-edged sword. Nat. Rev. Immunol. 2003, 3, 745–756. [Google Scholar] [CrossRef] [PubMed]
- Basith, S.; Manavalan, B.; Yoo, T.H.; Kim, S.G.; Choi, S. Roles of toll-like receptors in Cancer: A double-edged sword for defense and offense. Arch. Pharmacal Res. 2012, 35, 1297–1316. [Google Scholar] [CrossRef]
- Rakoff-Nahoum, S.; Medzhitov, R. Toll-like receptors and cancer. Nat. Cancer 2008, 9, 57–63. [Google Scholar] [CrossRef]
- Carmeliet, P. VEGF as a Key Mediator of Angiogenesis in Cancer. Oncology 2005, 69, 4–10. [Google Scholar] [CrossRef] [PubMed]
- Ridnour, L.A.; Cheng, R.; Switzer, C.H.; Heinecke, J.L.; Ambs, S.; Glynn, S.; Young, H.A.; Trinchieri, G.; Wink, D.A. Molecular Pathways: Toll-like Receptors in the Tumor Microenvironment—Poor Prognosis or New Therapeutic Opportunity. Clin. Cancer Res. 2012, 19, 1340–1346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, Y.; Goto, Y.; Narita, N.; Hoon, D.S. Cancer Cells Expressing Toll-like Receptors and the Tumor Microenvironment. Cancer Microenviron. 2009, 2, 205–214. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Role | Target Gene |
|---|---|
| Apoptosis | NFKB1, CDKN1A, CFLAR, IL2RA |
| Cell cycle regulators | CCND1, P21, P27, IL2RA |
| Cell proliferation | P21, P27, ERBB2, FOSL1, IL2RA |
| Cell differentiation | DTX1, HES6, PPARG |
| Neurogenesis | HES1, HEY1, HEY2 |
| Transcription | NFKB1, NR4A2, PPARG, STAT6, DTX1, HES1, HES6, HEY1, HEY2, FOS, FOSL1 |
| Unspecified | CD44, CHUK, PTCRA, LOR, MAP2K7, PDPK1, MGC61598, HES5, IFNG, IL 17B, IVL, KRT1, KRT10, KRT14, KRT5, LOR |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fasoulakis, Z.; Koutras, A.; Ntounis, T.; Pergialiotis, V.; Chionis, A.; Katrachouras, A.; Palios, V.-C.; Symeonidis, P.; Valsamaki, A.; Syllaios, A.; et al. The Prognostic Role and Significance of Dll4 and Toll-like Receptors in Cancer Development. Cancers 2022, 14, 1649. https://doi.org/10.3390/cancers14071649
Fasoulakis Z, Koutras A, Ntounis T, Pergialiotis V, Chionis A, Katrachouras A, Palios V-C, Symeonidis P, Valsamaki A, Syllaios A, et al. The Prognostic Role and Significance of Dll4 and Toll-like Receptors in Cancer Development. Cancers. 2022; 14(7):1649. https://doi.org/10.3390/cancers14071649
Chicago/Turabian StyleFasoulakis, Zacharias, Antonios Koutras, Thomas Ntounis, Vasilios Pergialiotis, Athanasios Chionis, Alexandros Katrachouras, Vasileios-Chrysovalantis Palios, Panagiotis Symeonidis, Asimina Valsamaki, Athanasios Syllaios, and et al. 2022. "The Prognostic Role and Significance of Dll4 and Toll-like Receptors in Cancer Development" Cancers 14, no. 7: 1649. https://doi.org/10.3390/cancers14071649