
The Role of Purinergic P2X7 Receptor in Inflammation and Cancer: Novel Molecular Insights and Clinical Applications

 ,
, 
 ,
,
Abstract
:Simple Summary
Abstract
1. Background
2. P2X7 Receptor Genetics, Characteristics, Function, and Tissue Distribution
3. The Role of P2X7 Receptor in Cancer
4. P2X7 Receptor Activation in Inflammation
4.1. P2X7R in Pathogenic Infection-Driven Inflammation
4.2. P2X7 Role in Tumour-Associated Inflammation
5. P2X7R as a Therapeutic Target in Cancer and Inflammation
5.1. P2X7R Agonists
5.2. P2X7R Antagonists
6. Conclusions and Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Di Virgilio, F.; Adinolfi, E. Extracellular purines, purinergic receptors and tumor growth. Oncogene 2017, 36, 293–303. [Google Scholar] [CrossRef]
- Giuliani, A.L.; Sarti, A.C.; Di Virgilio, F. Extracellular nucleotides and nucleosides as signalling molecules. Immunol. Lett. 2019, 205, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Greiner, J.V.; Glonek, T. Intracellular ATP Concentration and Implication for Cellular Evolution. Biology 2021, 10, 1166. [Google Scholar] [CrossRef]
- De Marchi, E.; Orioli, E.; Pegoraro, A.; Sangaletti, S.; Portararo, P.; Curti, A.; Colombo, M.P.; Di Virgilio, F.; Adinolfi, E. The P2X7 receptor modulates immune cells infiltration, ectonucleotidases expression and extracellular ATP levels in the tumor microenvironment. Oncogene 2019, 38, 3636–3650. [Google Scholar] [CrossRef]
- Joyce, J.A.; Fearon, D.T. T cell exclusion, immune privilege, and the tumor microenvironment. Science 2015, 348, 74–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inami, Y.; Fukushima, M.; Kume, T.; Uta, D. Histamine enhances ATP-induced itching and responsiveness to ATP in keratinocytes. J. Pharmacol. Sci. 2022, 148, 255–261. [Google Scholar] [CrossRef] [PubMed]
- North, R.A. P2X receptors. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016, 37, 20150427. [Google Scholar] [CrossRef]
- Lara, R.; Adinolfi, E.; Harwood, C.A.; Philpott, M.; Barden, J.A.; Di Virgilio, F.; McNulty, S. P2X7 in Cancer: From Molecular Mechanisms to Therapeutics. Front. Pharmacol. 2020, 11, 793. [Google Scholar] [CrossRef] [PubMed]
- Burnstock, G.; Kennedy, C. P2X Receptors in Health and Disease. Adv. Pharmacol. 2011, 61, 333–372. [Google Scholar]
- Xu, X.Y.; He, X.T.; Wang, J.; Li, X.; Xia, Y.; Tan, Y.Z.; Chen, F.M. Role of the P2X7 receptor in inflammation-mediated changes in the osteogenesis of periodontal ligament stem cells. Cell Death Dis. 2019, 10, 20. [Google Scholar] [CrossRef]
- Yang, C.; Shi, S.; Su, Y.; Tong, J.S.; Li, L. P2X7R promotes angiogenesis and tumour-associated macrophage recruitment by regulating the NF-κB signalling pathway in colorectal cancer cells. J. Cell. Mol. Med. 2020, 24, 10830–10841. [Google Scholar] [CrossRef]
- Adinolfi, E.; De Marchi, E.; Orioli, E.; Pegoraro, A.; Di Virgilio, F. Role of the P2X7 receptor in tumor-associated inflammation. Curr. Opin. Pharmacol. 2019, 47, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Popper, L.D.; Batra, S. Calcium mobilization and cell proliferation activated by extracellular ATP in human ovarian tumour cells. Cell Calcium 1993, 14, 209–218. [Google Scholar] [CrossRef]
- Hope, J.M.; Greenlee, J.D.; King, M.R. Mechanosensitive Ion Channels: TRPV4 and P2X7 in Disseminating Cancer Cells. Cancer J. 2018, 24, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Notomi, S.; Hisatomi, T.; Kanemaru, T.; Takeda, A.; Ikeda, Y.; Enaida, H.; Kroemer, G.; Ishibashi, T. Critical Involvement of Extracellular ATP Acting on P2RX7 Purinergic Receptors in Photoreceptor Cell Death. Am. J. Pathol. 2011, 179, 2798–2809. [Google Scholar] [CrossRef]
- Hou, Z.; Cao, J. Comparative study of the P2X gene family in animals and plants. Purinergic Signal. 2016, 12, 269–281. [Google Scholar] [CrossRef] [Green Version]
- Burnstock, G.; Knight, G.E. Cellular distribution and functions of P2 receptor subtypes in different systems. Int. Rev. Cytol. 2004, 240, 301–304. [Google Scholar]
- Benzaquen, J.; Heeke, S.; Janho dit Hreich, S.; Douguet, L.; Marquette, C.H.; Hofman, P.; Vouret-Craviari, V. Alternative splicing of P2RX7 pre-messenger RNA in health and diseases: Myth or reality? Biomed. J. 2019, 42, 141–151. [Google Scholar] [CrossRef]
- Jimenez-Mateos, E.M.; Smith, J.; Nicke, A.; Engel, T. Regulation of P2X7 receptor expression and function in the brain. Brain Res. Bull. 2019, 151, 153–163. [Google Scholar] [CrossRef]
- Zhou, L.; Qi, X.; Potashkin, J.A.; Abdul-Karim, F.W.; Gorodeski, G.I. MicroRNAs miR-186 and miR-150 Down-regulate Expression of the Pro-apoptotic Purinergic P2X7 Receptor by Activation of Instability Sites at the 3′-Untranslated Region of the Gene That Decrease Steady-state Levels of the Transcript. J. Biol. Chem. 2008, 283, 28274–28286. [Google Scholar] [CrossRef] [Green Version]
- Oton-Gonzalez, L.; Rotondo, J.C.; Cerritelli, L.; Malagutti, N.; Lanzillotti, C.; Bononi, I.; Ciorba, A.; Bianchini, C.; Mazziotta, C.; De Mattei, M.; et al. Association between oncogenic human papillomavirus type 16 and Killian polyp. Infect. Agent. Cancer 2021, 16, 3. [Google Scholar] [CrossRef]
- Rotondo, J.C.; Lanzillotti, C.; Mazziotta, C.; Tognon, M.; Martini, F. Epigenetics of male infertility: The role of DNA methylation. Front. Cell Dev. Biol. 2021, 9, 689624. [Google Scholar] [CrossRef] [PubMed]
- Rotondo, J.C.; Mazziotta, C.; Lanzillotti, C.; Tognon, M.; Martini, F. Epigenetic Dysregulations in Merkel Cell Polyomavirus-Driven Merkel Cell Carcinoma. Int. J. Mol. Sci. 2021, 22, 11464. [Google Scholar] [CrossRef]
- Lanzillotti, C.; De Mattei, M.; Mazziotta, C.; Taraballi, F.; Rotondo, J.C.; Tognon, M.; Martini, F. Long Non-coding RNAs and MicroRNAs Interplay in Osteogenic Differentiation of Mesenchymal Stem Cells. Front. Cell Dev. Biol. 2021, 9, 646032. [Google Scholar] [CrossRef] [PubMed]
- Rotondo, J.C.; Giari, L.; Guerranti, C.; Tognon, M.; Castaldelli, G.; Fano, E.A.; Martini, F. Environmental doses of perfluorooctanoic acid change the expression of genes in target tissues of common carp. Environ. Toxicol. Chem. 2018, 37, 942–948. [Google Scholar] [CrossRef]
- Roger, S.; Jelassi, B.; Couillin, I.; Pelegrin, P.; Besson, P.; Jiang, L.H. Understanding the roles of the P2X7 receptor in solid tumour progression and therapeutic perspectives. Biochim. Biophys. Acta—Biomembr. 2015, 1848, 2584–2602. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Luo, S.; Zhang, Y.; Jia, L.; Yang, C.; Peng, X.; Zhao, R. Production, characterization, and application of a monoclonal antibody specific for the extracellular domain of human P2X7R. Appl. Microbiol. Biotechnol. 2020, 104, 2017–2028. [Google Scholar] [CrossRef]
- Dalgarno, R.; Leduc-Pessah, H.; Pilapil, A.; Kwok, C.H.T.; Trang, T. Intrathecal delivery of a palmitoylated peptide targeting Y382-384 within the P2X7 receptor alleviates neuropathic pain. Mol. Pain 2018, 14, 1744806918795793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karasawa, A.; Michalski, K.; Mikhelzon, P.; Kawate, T. The P2X7 receptor forms a dye-permeable pore independent of its intracellular domain but dependent on membrane lipid composition. eLife 2017, 6, e31186. [Google Scholar] [CrossRef]
- Gonnord, P.; Delarasse, C.; Auger, R.; Benihoud, K.; Prigent, M.; Cuif, M.H.; Lamaze, C.; Kanellopoulos, J.M. Palmitoylation of the P2X7 receptor, an ATP-gated channel, controls its expression and association with lipid rafts. FASEB J. 2009, 23, 795–805. [Google Scholar] [CrossRef]
- Wickert, L.E.; Blanchette, J.B.; Waldschmidt, N.V.; Bertics, P.J.; Denu, J.M.; Denlinger, L.C.; Lenertz, L.Y. The C-Terminus of Human Nucleotide Receptor P2X7 Is Critical for Receptor Oligomerization and N-Linked Glycosylation. PLoS ONE 2013, 8, e63789. [Google Scholar] [CrossRef]
- Martínez-cuesta, M.Á.; Blanch-ruiz, M.A.; Ortega-luna, R.; Sánchez-lópez, A.; Álvarez, Á.; Karasawa, A.; Kawate, T. Structural and Functional Basis for Understanding the Biological Significance of P2X7 Receptor. Int. J. Mol. Sci. 2020, 21, 8454. [Google Scholar] [CrossRef] [PubMed]
- Wiley, J.S.; Dao-Ung, L.P.; Li, C.; Shemon, A.N.; Gu, B.J.; Smart, M.L.; Fuller, S.J.; Barden, J.A.; Petrou, S.; Sluyter, R. An Ile-568 to Asn Polymorphism Prevents Normal Trafficking and Function of the Human P2X7 Receptor. J. Biol. Chem. 2003, 278, 17108–17113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradley, H.J.; Liu, X.; Collins, V.; Owide, J.; Goli, G.R.; Smith, M.; Surprenant, A.; White, S.J.; Jiang, L.H. Identification of an intracellular microdomain of the P2X7 receptor that is crucial in basolateral membrane targeting in epithelial cells. FEBS Lett. 2010, 584, 4740–4744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smart, M.L.; Gu, B.; Panchal, R.G.; Wiley, J.; Cromer, B.; Williams, D.A.; Petrou, S. P2X7 Receptor Cell Surface Expression and Cytolytic Pore Formation Are Regulated by a Distal C-terminal Region. J. Biol. Chem. 2003, 278, 8853–8860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa-Junior, H.M.; Vieira, F.S.; Coutinho-Silva, R. C terminus of the P2X7 receptor: Treasure hunting. Purinergic Signal. 2011, 7, 7–19. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Xiao, Y.; Li, Z. P2X7 receptor positively regulates MyD88-dependent NF-κB activation. Cytokine 2011, 55, 229–236. [Google Scholar] [CrossRef]
- Jiang, L.H.; Caseley, E.A.; Muench, S.P.; Roger, S. Structural basis for the functional properties of the P2X7 receptor for extracellular ATP. Purinergic Signal. 2021, 17, 331–344. [Google Scholar] [CrossRef]
- Orioli, E.; De Marchi, E.; Giuliani, A.L.; Adinolfi, E. P2X7 Receptor Orchestrates Multiple Signalling Pathways Triggering Inflammation, Autophagy and Metabolic/Trophic Responses. Curr. Med. Chem. 2017, 24, 2261–2275. [Google Scholar] [CrossRef]
- Pelegrin, P.; Surprenant, A. Pannexin-1 Couples to Maitotoxin- and Nigericin-induced Interleukin-1β Release through a Dye Uptake-independent Pathway. J. Biol. Chem. 2007, 282, 2386–2394. [Google Scholar] [CrossRef] [Green Version]
- Baroja-Mazo, A.; Barberà-Cremades, M.; Pelegrín, P. The participation of plasma membrane hemichannels to purinergic signaling. Biochim. Biophys. Acta—Biomembr. 2013, 1828, 79–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Virgilio, F.; Giuliani, A.L.; Vultaggio-Poma, V.; Falzoni, S.; Sarti, A.C. Non-nucleotide agonists triggering P2X7 receptor activation and pore formation. Front. Pharmacol. 2018, 9, 39. [Google Scholar] [CrossRef] [PubMed]
- North, R.A. Molecular physiology of P2X receptors. Physiol. Rev. 2002, 82, 1013–1067. [Google Scholar] [CrossRef]
- Bergamin, L.S.; Capece, M.; Salaro, E.; Sarti, A.C.; Falzoni, S.; Pereira, M.S.L.; De Bastiani, M.A.; Scholl, J.N.; Battastini, A.M.O.; Di Virgilio, F. Role of the P2X7 receptor in in vitro and in vivo glioma tumor growth. Oncotarget 2019, 10, 4840–4856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roger, S.; Mei, Z.Z.; Baldwin, J.M.; Dong, L.; Bradley, H.; Baldwin, S.A.; Surprenant, A.; Jiang, L.H. Single nucleotide polymorphisms that were identified in affective mood disorders affect ATP-activated P2X7 receptor functions. J. Psychiatr. Res. 2010, 44, 347–355. [Google Scholar] [CrossRef]
- Young, C.N.J.; Górecki, D.C. P2RX7 purinoceptor as a therapeutic target-The second coming? Front. Chem. 2018, 6, 248. [Google Scholar] [CrossRef] [Green Version]
- Young, C.N.J.; Chira, N.; Róg, J.; Al-Khalidi, R.; Benard, M.; Galas, L.; Chan, P.; Vaudry, D.; Zabłocki, K.; Górecki, D.C. Sustained activation of P2X7 induces MMP-2-evoked cleavage and functional purinoceptor inhibition. J. Mol. Cell Biol. 2018, 10, 229–242. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Ren, W.; Zhao, F.; Han, Y.; Liu, C.; Jia, K. Curcumin amends Ca2+ dysregulation in microglia by suppressing the activation of P2X7 receptor. Mol. Cell. Biochem. 2020, 465, 65–73. [Google Scholar] [CrossRef]
- Furini, F.; Giuliani, A.L.; Parlati, M.E.; Govoni, M.; Di Virgilio, F.; Bortoluzzi, A. P2X7 receptor expression in patients with serositis related to systemic lupus erythematosus. Front. Pharmacol. 2019, 10, 435. [Google Scholar] [CrossRef]
- Burnstock, G.; Knight, G.E. The potential of P2X7 receptors as a therapeutic target, including inflammation and tumour progression. Purinergic Signal. 2018, 14, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Petrou, S.; Ugur, M.; Drummond, R.M.; Singer, J.J.; Walsh, J.V. P2X7 purinoceptor expression in Xenopus oocytes is not sufficient to produce a pore-forming P2Z-like phenotype. FEBS Lett. 1997, 411, 339–345. [Google Scholar] [CrossRef] [Green Version]
- Browne, L.E.; Compan, V.; Bragg, L.; North, R.A. P2X7 Receptor Channels Allow Direct Permeation of Nanometer-Sized Dyes. J. Neurosci. 2013, 33, 3557–3566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Virgilio, F.; Jiang, L.H.; Roger, S.; Falzoni, S.; Sarti, A.C.; Vultaggio-Poma, V.; Chiozzi, P.; Adinolfi, E. Structure, function and techniques of investigation of the P2X7 receptor (P2X7R) in mammalian cells. Methods Enzymol. 2019, 629, 115–150. [Google Scholar] [PubMed]
- Rossi, L.; Salvestrini, V.; Ferrari, D.; Di Virgilio, F.; Lemoli, R.M. The sixth sense: Hematopoietic stem cells detect danger through purinergic signaling. Blood 2012, 120, 2365–2375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, L.H.; Hao, Y.; Mousawi, F.; Peng, H.; Yang, X. Expression of P2 Purinergic Receptors in Mesenchymal Stem Cells and Their Roles in Extracellular Nucleotide Regulation of Cell Functions. J. Cell. Physiol. 2017, 232, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, A.; Gartland, A. P2x7 receptors: Role in bone cell formation and function. J. Mol. Endocrinol. 2015, 54, R75–R88. [Google Scholar] [CrossRef] [Green Version]
- Fan, X.; Ma, W.; Zhang, Y.; Zhang, L. P2X7 Receptor (P2X7R) of microglia mediates neuroinflammation by regulating (NOD)-like receptor protein 3 (NLRP3) inflammasome-dependent inflammation after spinal cord injury. Med. Sci. Monit. 2020, 21, e925491. [Google Scholar] [CrossRef]
- Kopp, R.; Krautloher, A.; Ramírez-Fernández, A.; Nicke, A. P2X7 Interactions and Signaling—Making Head or Tail of It. Front. Mol. Neurosci. 2019, 12, 183. [Google Scholar] [CrossRef]
- Giannuzzo, A.; Pedersen, S.F.; Novak, I. The P2X7 receptor regulates cell survival, migration and invasion of pancreatic ductal adenocarcinoma cells. Mol. Cancer 2015, 14, 203. [Google Scholar] [CrossRef] [Green Version]
- Di Virgilio, F.; Ferrari, D.; Adinolfi, E. P2X7: A growth-promoting receptor—Implications for cancer. Purinergic Signal. 2009, 5, 251–256. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, S.; Oliphant, C.; Hassan, S.; Peille, A.; Bronsert, P.; Falzoni, S.; Di Virgilio, F.; McNulty, S.; Lara, R. ATP in the tumour microenvironment drives expression of nfP2X 7, a key mediator of cancer cell survival. Oncogene 2019, 38, 194–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Ding, J.; Wang, L. The role of P2X7 receptor in prognosis and metastasis of colorectal cancer. Adv. Med. Sci. 2019, 64, 388–394. [Google Scholar] [CrossRef]
- Calik, I.; Calik, M.; Turken, G.; Ozercan, I.H. A promising independent prognostic biomarker in colorectal cancer: P2X7 receptor. Int. J. Clin. Exp. Pathol. 2020, 13, 107–1214. [Google Scholar]
- Li, Q.; Zhu, X.; Song, W.; Peng, X.; Zhao, R. The P2X7 purinergic receptor: A potential therapeutic target for lung cancer. J. Cancer Res. Clin. Oncol. 2020, 146, 2731–2741. [Google Scholar] [CrossRef]
- Maxová, H.; Bacáková, L.; Lisá, V.; Novotná, J.; Tomásová, H.; Vízek, M.; Herget, J. Production of proteolytic enzymes in mast cells, fibroblasts, vascular smooth muscle and endothelial cells cultivated under normoxic or hypoxic conditions. Physiol. Res. 2010, 59, 711–719. [Google Scholar] [CrossRef] [PubMed]
- Adinolfi, E.; Raffaghello, L.; Giuliani, A.L.; Cavazzini, L.; Capece, M.; Chiozzi, P.; Bianchi, G.; Kroemer, G.; Pistoia, V.; Di Virgilio, F. Expression of P2X7 receptor increases in vivo tumor growth. Cancer Res. 2012, 72, 2957–2969. [Google Scholar] [CrossRef] [Green Version]
- Tafani, M.; Schito, L.; Pellegrini, L.; Villanova, L.; Marfe, G.; Anwar, T.; Rosa, R.; Indelicato, M.; Fini, M.; Pucci, B.; et al. Hypoxia-increased RAGE and P2X7R expression regulates tumor cell invasion through phosphorylation of Erk1/2 and Akt and nuclear translocation of NF-κB. Carcinogenesis 2011, 32, 1167–1175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Jiang, M.; Cui, B.W.; Jin, C.H.; Wu, Y.L.; Shang, Y.; Yang, H.X.; Wu, M.; Liu, J.; Qiao, C.Y.; et al. P2X7 receptor-targeted regulation by tetrahydroxystilbene glucoside in alcoholic hepatosteatosis: A new strategy towards macrophage–hepatocyte crosstalk. Br. J. Pharmacol. 2020, 177, 2793–2811. [Google Scholar] [CrossRef]
- Souza, C.O.; Santoro, G.F.; Figliuolo, V.R.; Nanini, H.F.; De Souza, H.S.P.; Castelo-Branco, M.T.L.; Abalo, A.A.; Paiva, M.M.; Coutinho, C.M.L.M.; Coutinho-Silva, R. Extracellular ATP induces cell death in human intestinal epithelial cells. Biochim. Biophys. Acta—Gen. Subj. 2012, 1820, 1867–1878. [Google Scholar] [CrossRef]
- Hattori, F.; Ohshima, Y.; Seki, S.; Tsukimoto, M.; Sato, M.; Takenouchi, T.; Suzuki, A.; Takai, E.; Kitani, H.; Harada, H.; et al. Feasibility study of B16 melanoma therapy using oxidized ATP to target purinergic receptor P2X7. Eur. J. Pharmacol. 2012, 695, 20–26. [Google Scholar] [CrossRef]
- Amoroso, F.; Capece, M.; Rotondo, A.; Cangelosi, D.; Ferracin, M.; Franceschini, A.; Raffaghello, L.; Pistoia, V.; Varesio, L.; Adinolfi, E. The P2X7 receptor is a key modulator of the PI3K/GSK3β/VEGF signaling network: Evidence in experimental neuroblastoma. Oncogene 2015, 34, 5240–5251. [Google Scholar] [CrossRef]
- Gu, B.J.; Wiley, J.S. Rapid ATP-induced release of matrix metalloproteinase 9 is mediated by the P2X7 receptor. Blood 2006, 107, 4946–4953. [Google Scholar] [CrossRef] [Green Version]
- Xia, J.; Yu, X.; Tang, L.; Li, G.; He, T. P2X7 receptor stimulates breast cancer cell invasion and migration via the AKT pathway. Oncol. Rep. 2015, 34, 103–110. [Google Scholar] [CrossRef] [Green Version]
- Jelassi, B.; Chantme, A.; Alcaraz-Pérez, F.; Baroja-Mazo, A.; Cayuela, M.L.; Pelegrin, P.; Surprenant, A.; Roger, S. P2X 7 receptor activation enhances SK3 channels- and cystein cathepsin-dependent cancer cells invasiveness. Oncogene 2011, 30, 2108–2122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bekes, E.M.; Schweighofer, B.; Kupriyanova, T.A.; Zajac, E.; Ardi, V.C.; Quigley, J.P.; Deryugina, E.I. Tumor-recruited neutrophils and neutrophil TIMP-free MMP-9 regulate coordinately the levels of tumor angiogenesis and efficiency of malignant cell intravasation. Am. J. Pathol. 2011, 179, 1455–1470. [Google Scholar] [CrossRef] [PubMed]
- Murphy, N.; Lynch, M.A. Activation of the P2X7 receptor induces migration of glial cells by inducing cathepsin B degradation of tissue inhibitor of metalloproteinase 1. J. Neurochem. 2012, 123, 761–770. [Google Scholar] [CrossRef]
- Arnaud-Sampaio, V.F.; Rabelo, I.L.A.; Ulrich, H.; Lameu, C. The P2X7 Receptor in the Maintenance of Cancer Stem Cells, Chemoresistance and Metastasis. Stem Cell Rev. Rep. 2020, 16, 288–300. [Google Scholar] [CrossRef]
- Schneider, G.; Glaser, T.; Lameu, C.; Abdelbaset-Ismail, A.; Sellers, Z.P.; Moniuszko, M.; Ulrich, H.; Ratajczak, M.Z. Extracellular nucleotides as novel, underappreciated pro-metastatic factors that stimulate purinergic signaling in human lung cancer cells. Mol. Cancer 2015, 14, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takai, E.; Tsukimoto, M.; Harada, H.; Kojima, S. Autocrine signaling via release of ATP and activation of P2X7 receptor influences motile activity of human lung cancer cells. Purinergic Signal. 2014, 10, 487–497. [Google Scholar] [CrossRef] [Green Version]
- Jelassi, B.; Anchelin, M.; Chamouton, J.; Cayuela, M.L.; Clarysse, L.; Li, J.; Goré, J.; Jiang, L.H.; Roger, S. Anthraquinone emodin inhibits human cancer cell invasiveness by antagonizing P2X7 receptors. Carcinogenesis 2013, 34, 1487–1496. [Google Scholar] [CrossRef] [Green Version]
- Amoroso, F.; Salaro, E.; Falzoni, S.; Chiozzi, P.; Giuliani, A.L.; Cavallesco, G.; Maniscalco, P.; Puozzo, A.; Bononi, I.; Martini, F.; et al. P2X7 targeting inhibits growth of human mesothelioma. Oncotarget 2016, 7, 49664–49676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-Villafuertes, R.; García-Huerta, P.; Díaz-Hernández, J.I.; Miras-Portugal, M.T. PI3K/Akt signaling pathway triggers P2X7 receptor expression as a pro-survival factor of neuroblastoma cells under limiting growth conditions. Sci. Rep. 2015, 5, 18417. [Google Scholar] [CrossRef] [Green Version]
- Amoroso, F.; Falzoni, S.; Adinolfi, E.; Ferrari, D.; Di Virgilio, F. The P2X7 receptor is a key modulator of aerobic glycolysis. Cell Death Dis. 2012, 3, e370. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Huang, X.; Tian, Y.; Yan, X.; Wang, F.; Chen, J.; Zhang, Q.; Zhang, Q.; Yuan, X. Phosphorylated GSK-3β protects stress-induced apoptosis of myoblasts via the PI3K/Akt signaling pathway. Mol. Med. Rep. 2020, 22, 317–327. [Google Scholar] [CrossRef]
- Kunnimalaiyaan, S.; Schwartz, V.K.; Jackson, I.A.; Clark Gamblin, T.; Kunnimalaiyaan, M. Antiproliferative and apoptotic effect of LY2090314, a GSK-3 inhibitor, in neuroblastoma in vitro. BMC Cancer 2018, 18, 560. [Google Scholar] [CrossRef] [Green Version]
- Qin, L.X. Inflammatory Immune Responses in Tumor Microenvironment and Metastasis of Hepatocellular Carcinoma. Cancer Microenviron. 2012, 5, 203–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pellegatti, P.; Raffaghello, L.; Bianchi, G.; Piccardi, F.; Pistoia, V.; Di Virgilio, F. Increased Level of Extracellular ATP at Tumor Sites: In Vivo Imaging with Plasma Membrane Luciferase. PLoS ONE 2008, 3, e2599. [Google Scholar] [CrossRef] [PubMed]
- Savio, L.E.B.; Mello, P.d.A.; da Silva, C.G.; Coutinho-Silva, R. The P2X7 receptor in inflammatory diseases: Angel or demon? Front. Pharmacol. 2018, 9, 52. [Google Scholar] [CrossRef] [Green Version]
- Burnstock, G. P2X ion channel receptors and inflammation. Purinergic Signal. 2016, 12, 59–67. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Castejon, G.; Brough, D. Understanding the mechanism of IL-1β secretion. Cytokine Growth Factor Rev. 2011, 22, 189–195. [Google Scholar] [CrossRef]
- Carta, S.; Rubartelli, R.L. Different Members of the IL-1 Family Come Out in Different Ways: DAMPs vs. Cytokines? Front. Immunol. 2013, 4, 123. [Google Scholar] [CrossRef] [Green Version]
- Scarpellino, G.; Genova, T.; Munaron, L. Purinergic P2X7 Receptor: A Cation Channel Sensitive to Tumor Microenvironment. Recent Pat. Anticancer. Drug Discov. 2019, 14, 32–38. [Google Scholar] [CrossRef]
- Di Virgilio, F.; Dal Ben, D.; Sarti, A.C.; Giuliani, A.L.; Falzoni, S. The P2X7 Receptor in Infection and Inflammation. Immunity 2017, 47, 15–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Killeen, M.E.; Ferris, L.; Kupetsky, E.A.; Falo, L.; Mathers, A.R. Signaling through Purinergic Receptors for ATP Induces Human Cutaneous Innate and Adaptive Th17 Responses: Implications in the Pathogenesis of Psoriasis. J. Immunol. 2013, 190, 4324–4336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eberhardt, N.; Bergero, G.; Mazzocco Mariotta, Y.L.; Aoki, M.P. Purinergic modulation of the immune response to infections. Purinergic Signal. 2022, 1, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Soare, A.Y.; Freeman, T.L.; Min, A.K.; Malik, H.S.; Osota, E.O.; Swartz, T.H. P2RX7 at the Host-Pathogen Interface of Infectious Diseases. Microbiol. Mol. Biol. Rev. 2021, 85, e00055-20. [Google Scholar] [CrossRef]
- Zhang, C.; He, H.; Wang, L.; Zhang, N.; Huang, H.; Xiong, Q.; Yan, Y.; Wu, N.; Ren, H.; Han, H.; et al. Virus-Triggered ATP Release Limits Viral Replication through Facilitating IFN-β Production in a P2X7-Dependent Manner. J. Immunol. 2017, 199, 1372–1381. [Google Scholar] [CrossRef] [PubMed]
- Corrêa, G.; Marques da Silva, C.; de Abreu Moreira-Souza, A.C.; Vommaro, R.C.; Coutinho-Silva, R. Activation of the P2X(7) receptor triggers the elimination of Toxoplasma gondii tachyzoites from infected macrophages. Microbes Infect. 2010, 12, 497–504. [Google Scholar] [CrossRef]
- Lee, B.H.; Hwang, D.M.; Palaniyar, N.; Grinstein, S.; Philpott, D.J.; Hu, J. Activation of P2X7 Receptor by ATP Plays an Important Role in Regulating Inflammatory Responses during Acute Viral Infection. PLoS ONE 2012, 7, e35812. [Google Scholar] [CrossRef] [Green Version]
- Paoletti, A.; Raza, S.Q.; Voisin, L.; Law, F.; Pipoli da Fonseca, J.; Caillet, M.; Kroemer, G.; Perfettini, J.L. Multifaceted roles of purinergic receptors in viral infection. Microbes Infect. 2012, 14, 1278–1283. [Google Scholar] [CrossRef]
- Pacheco, P.A.F.; Faria, R.X.; Ferreira, L.G.B.; Paixão, I.C.N.P. Putative roles of purinergic signaling in human immunodeficiency virus-1 infection. Biol. Direct 2014, 9, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barat, C.; Gilbert, C.; Imbeault, M.; Tremblay, M.J. Extracellular ATP reduces HIV-1 transfer from immature dendritic cells to CD4+ T lymphocytes. Retrovirology 2008, 5, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, J.M.; Han, Z. Purinergic Receptor Functionality Is Necessary for Infection of Human Hepatocytes by Hepatitis Delta Virus and Hepatitis B Virus. PLoS ONE 2010, 5, e15784. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, D.E.; Oliveira-Giacomelli, Á.; Glaser, T.; Arnaud-Sampaio, V.F.; Andrejew, R.; Dieckmann, L.; Baranova, J.; Lameu, C.; Ratajczak, M.Z.; Ulrich, H. Hyperactivation of P2X7 receptors as a culprit of COVID-19 neuropathology. Mol. Psychiatry 2020, 26, 1044–1059. [Google Scholar] [CrossRef] [PubMed]
- Franciosi, M.L.M.; Lima, M.D.M.; Schetinger, M.R.C.; Cardoso, A.M. Possible role of purinergic signaling in COVID-19. Mol. Cell. Biochem. 2021, 476, 2891–2898. [Google Scholar] [CrossRef]
- Rotondo, J.C.; Martini, F.; Maritati, M.; Mazziotta, C.; Di Mauro, G.; Lanzillotti, C.; Barp, N.; Gallerani, A.; Tognon, M.; Contini, C. SARS-CoV-2 Infection: New Molecular, Phylogenetic, and Pathogenetic Insights. Efficacy of Current Vaccines and the Potential Risk of Variants. Viruses 2021, 13, 1687. [Google Scholar] [CrossRef] [PubMed]
- Contini, C.; Rotondo, J.C.; Magagnoli, F.; Maritati, M.; Seraceni, S.; Graziano, A.; Poggi, A.; Capucci, R.; Vesce, F.; Tognon, M.; et al. Investigation on silent bacterial infections in specimens from pregnant women affected by spontaneous miscarriage. J. Cell. Physiol. 2018, 234, 100–107. [Google Scholar] [CrossRef] [Green Version]
- Coutinho-Silva, R.; Perfettini, J.L.; Persechini, P.M.; Dautry-Varsat, A.; Ojcius, D.M. Modulation of P2Z/P2X7 receptor activity in macrophages infected with Chlamydia psittaci. Am. J. Physiol.—Cell Physiol. 2001, 280, C81–C89. [Google Scholar] [CrossRef] [Green Version]
- Abdul-Sater, A.A.; Saïd-Sadier, N.; Padilla, E.V.; Ojcius, D.M. Chlamydial infection of monocytes stimulates IL-1β secretion through activation of the NLRP3 inflammasome. Microbes Infect. 2010, 12, 652–661. [Google Scholar] [CrossRef] [Green Version]
- Santiago-Carvalho, I.; de Almeida-Santos, G.; Bomfim, C.C.B.; Souza, P.C.D.; Melo, B.M.S.D.; Amaral, E.P.; Cione, M.V.P.; Lasunskaia, E.; Hirata, M.H.; Alves-Filho, J.C.F.; et al. P2x7 Receptor Signaling Blockade Reduces Lung Inflammation and Necrosis During Severe Experimental Tuberculosis. Front. Cell. Infect. Microbiol. 2021, 11, 385. [Google Scholar] [CrossRef]
- Csóka, B.; Németh, Z.H.; Töro, G.; Idzko, M.; Zech, A.; Koscsó, B.; Spolarics, Z.; Antonioli, L.; Cseri, K.; Erdélyi, K.; et al. Extracellular ATP protects against sepsis through macrophage P2X7 purinergic receptors by enhancing intracellular bacterial killing. FASEB J. 2015, 29, 3626–3637. [Google Scholar] [CrossRef] [Green Version]
- Savio, L.E.B.; de Andrade Mello, P.; Figliuolo, V.R.; de Avelar Almeida, T.F.; Santana, P.T.; Oliveira, S.D.S.; Silva, C.L.M.; Feldbrügge, L.; Csizmadia, E.; Minshall, R.D.; et al. CD39 limits P2X7 receptor inflammatory signaling and attenuates sepsis-induced liver injury. J. Hepatol. 2017, 67, 716–726. [Google Scholar] [CrossRef] [Green Version]
- Van De Veerdonk, F.L.; Joosten, L.A.B.; Devesa, I.; Héctor, M.M.M.; Kanneganti, T.D.; Dinarello, C.A.; Van Der Meer, J.W.M.; Gow, N.A.R.; Kullberg, B.J.; Netea, M.G. Bypassing Pathogen-Induced Inflammasome Activation for the Regulation of Interleukin-1β Production by the Fungal Pathogen Candida albicans. J. Infect. Dis. 2009, 199, 1087–1096. [Google Scholar] [CrossRef] [Green Version]
- Feriotti, C.; de Araújo, E.F.; Loures, F.V.; da Costa, T.A.; Galdino, N.A.D.L.; Zamboni, D.S.; Calich, V.L.G. NOD-like receptor P3 inflammasome controls protective Th1/Th17 immunity against pulmonary paracoccidioidomycosis. Front. Immunol. 2017, 8, 786. [Google Scholar] [CrossRef]
- Snelgrove, R.J.; Gregory, L.G.; Peiró, T.; Akthar, S.; Campbell, G.A.; Walker, S.A.; Lloyd, C.M. Alternaria-derived serine protease activity drives IL-33–mediated asthma exacerbations. J. Allergy Clin. Immunol. 2014, 134, 583–592.e6. [Google Scholar] [CrossRef] [Green Version]
- Quan, J.H.; Huang, R.; Wang, Z.; Huang, S.; Choi, I.W.; Zhou, Y.; Lee, Y.H.; Chu, J.Q. P2X7 receptor mediates NLRP3-dependent IL-1β secretion and parasite proliferation in Toxoplasma gondii-infected human small intestinal epithelial cells. Parasites Vectors 2018, 11, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaves, S.P.; Torres-Santos, E.C.; Marques, C.; Figliuolo, V.R.; Persechini, P.M.; Coutinho-Silva, R.; Rossi-Bergmann, B. Modulation of P2X7 purinergic receptor in macrophages by Leishmania amazonensis and its role in parasite elimination. Microbes Infect. 2009, 11, 842–849. [Google Scholar] [CrossRef]
- Oliveira, S.D.A.d.S.; Coutinho-Silva, R.; Silva, C.L.M. Endothelial P2X7 receptors’ expression is reduced by schistosomiasis. Purinergic Signal. 2012, 9, 81–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, S.D.A.S.; Nanini, H.F.; Savio, L.E.B.; Waghabi, M.C.; Silva, C.L.M.; Coutinho-Silva, R. Macrophage P2X7 Receptor Function is Reduced during Schistosomiasis: Putative Role of TGF-β 1. Mediators Inflamm. 2014, 2014, 134974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruz, L.N.; Wu, Y.; Ulrich, H.; Craig, A.G.; Garcia, C.R.S. Tumor necrosis factor reduces Plasmodium falciparum growth and activates calcium signaling in human malaria parasites. Biochim. Biophys. Acta—Gen. Subj. 2016, 1860, 1489–1497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levano-Garcia, J.; Dluzewski, A.R.; Markus, R.P.; Garcia, C.R.S. Purinergic signalling is involved in the malaria parasite Plasmodium falciparum invasion to red blood cells. Purinergic Signal. 2010, 6, 365–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamarsheh, S.; Zeiser, R. NLRP3 Inflammasome Activation in Cancer: A Double-Edged Sword. Front. Immunol. 2020, 11, 1444. [Google Scholar] [CrossRef]
- Makarenkova, H.P.; Shah, S.B.; Shestopalov, V.I. The two faces of pannexins: New roles in inflammation and repair. J. Inflamm. Res. 2018, 11, 273–288. [Google Scholar] [CrossRef] [Green Version]
- Hung, S.C.; Choi, C.H.; Said-Sadier, N.; Johnson, L.; Atanasova, K.R.; Sellami, H.; Yilmaz, Ö.; Ojcius, D.M. P2X4 Assembles with P2X7 and Pannexin-1 in Gingival Epithelial Cells and Modulates ATP-induced Reactive Oxygen Species Production and Inflammasome Activation. PLoS ONE 2013, 8, e70210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Draganov, D.; Gopalakrishna-Pillai, S.; Chen, Y.R.; Zuckerman, N.; Moeller, S.; Wang, C.; Ann, D.; Lee, P.P. Modulation of P2X4/P2X7/Pannexin-1 sensitivity to extracellular ATP via Ivermectin induces a non-apoptotic and inflammatory form of cancer cell death. Sci. Rep. 2015, 5, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Miao, E.A.; Rajan, J.V.; Aderem, A. Caspase-1-induced pyroptotic cell death. Immunol. Rev. 2011, 243, 206–214. [Google Scholar] [CrossRef]
- Lin, L.; Huang, S.; Zhu, Z.; Han, J.; Wang, Z.; Huang, W.; Huang, Z. P2X7 receptor regulates EMMPRIN and MMP-9 expression through AMPK/MAPK signaling in PMA-induced macrophages. Mol. Med. Rep. 2018, 18, 3027–3033. [Google Scholar] [CrossRef] [Green Version]
- Corazza, M.; Oton-Gonzalez, L.; Scuderi, V.; Rotondo, J.C.; Lanzillotti, C.; Di Mauro, G.; Tognon, M.; Martini, F.; Borghi, A. Tissue cytokine/chemokine profile in vulvar lichen sclerosus: An observational study on keratinocyte and fibroblast cultures. J. Dermatol. Sci. 2020, 100, 223–226. [Google Scholar] [CrossRef] [PubMed]
- Siebers, K.; Fink, B.; Zakrzewicz, A.; Agné, A.; Richter, K.; Konzok, S.; Hecker, A.; Zukunft, S.; Küllmar, M.; Klein, J.; et al. Alpha-1 antitrypsin inhibits ATP-mediated release of interleukin-1β via CD36 and nicotinic acetylcholine receptors. Front. Immunol. 2018, 9, 877. [Google Scholar] [CrossRef] [Green Version]
- Rotondo, J.C.; Oton-Gonzalez, L.; Selvatici, R.; Rizzo, P.; Pavasini, R.; Campo, G.C.; Lanzillotti, C.; Mazziotta, C.; De Mattei, M.; Tognon, M.; et al. SERPINA1 Gene Promoter Is Differentially Methylated in Peripheral Blood Mononuclear Cells of Pregnant Women. Front. Cell Dev. Biol. 2020, 8, 5505. [Google Scholar] [CrossRef]
- Rotondo, J.C.; Aquila, G.; Oton-Gonzalez, L.; Selvatici, R.; Rizzo, P.; De Mattei, M.; Pavasini, R.; Tognon, M.; Campo, G.C.; Martini, F. Methylation of SERPINA1 gene promoter may predict chronic obstructive pulmonary disease in patients affected by acute coronary syndrome. Clin. Epigenet. 2021, 13, 79. [Google Scholar] [CrossRef] [PubMed]
- Lotfi, R.; Steppe, L.; Hang, R.; Rojewski, M.; Massold, M.; Jahrsdörfer, B.; Schrezenmeier, H. ATP promotes immunosuppressive capacities of mesenchymal stromal cells by enhancing the expression of indoleamine dioxygenase. Immun. Inflamm. Dis. 2018, 6, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Di Virgilio, F.; Sarti, A.C.; Falzoni, S.; De Marchi, E.; Adinolfi, E. Extracellular ATP and P2 purinergic signalling in the tumour microenvironment. Nat. Rev. Cancer 2018, 18, 601–618. [Google Scholar] [CrossRef]
- Zhang, Y.; Cheng, H.; Li, W.; Wu, H.; Yang, Y. Highly-expressed P2X7 receptor promotes growth and metastasis of human HOS/MNNG osteosarcoma cells via PI3K/Akt/GSK3β/β-catenin and mTOR/HIF1α/VEGF signaling. Int. J. Cancer 2019, 145, 1068–1082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, Z.; Xie, Y.; Guan, Y.; Zhang, Y.; Cho, K.S.; Ji, M.; You, Y. Involvement of P2X7 Receptor in Proliferation and Migration of Human Glioma Cells. Biomed Res. Int. 2018, 9, 8591397. [Google Scholar] [CrossRef] [Green Version]
- Vijayan, D.; Young, A.; Teng, M.W.L.; Smyth, M.J. Targeting immunosuppressive adenosine in cancer. Nat. Rev. Cancer 2017, 17, 709–724. [Google Scholar] [CrossRef]
- Antonioli, L.; Blandizzi, C.; Pacher, P.; Haskó, G. Immunity, inflammation and cancer: A leading role for adenosine. Nat. Rev. Cancer 2013, 13, 842–857. [Google Scholar] [CrossRef]
- Li, M.; Toombes, G.E.S.; Silberberg, S.D.; Swartz, K.J. Physical basis of apparent pore dilation of ATP-activated P2X receptor channels. Nat. Neurosci. 2015, 18, 1577–1583. [Google Scholar] [CrossRef]
- Acuna, C.; Capelli, C.; Coddou, C.; Escobar, A.; Imarai, M.; Yohana, L.; Lopex, X.; Rios, M.; Lopez, M. WO2013023319A1—In Vitro Method for Modifying the Depletion Profile of Treg Cells Present in a Total Splenocyte Population of a Biological Sample by Means of the Isolation, Culturing and Exposure Thereof to an Atp and Polymixin b Medium. U.S. Patent 14/181,810, 18 December 2014. [Google Scholar]
- Feng, L.L.; Cai, Y.Q.; Zhu, M.C.; Xing, L.J.; Wang, X. The yin and yang functions of extracellular ATP and adenosine in tumor immunity. Cancer Cell Int. 2020, 20, 110. [Google Scholar] [CrossRef] [PubMed]
- Avanzato, D.; Genova, T.; Fiorio Pla, A.; Bernardini, M.; Bianco, S.; Bussolati, B.; Mancardi, D.; Giraudo, E.; Maione, F.; Cassoni, P.; et al. Activation of P2X7 and P2Y11 purinergic receptors inhibits migration and normalizes tumor-derived endothelial cells via cAMP signaling. Sci. Rep. 2016, 6, 1–16. [Google Scholar]
- Barden, J.A. Non-Functional P2X7: A Novel and Ubiquitous Target in Human Cancer. J. Clin. Cell. Immunol. 2014, 5, 4. [Google Scholar] [CrossRef] [Green Version]
- Stokes, L.; Bidula, S.; Bibič, L.; Allum, E. To Inhibit or Enhance? Is There a Benefit to Positive Allosteric Modulation of P2X Receptors? Front. Pharmacol. 2020, 11, 627. [Google Scholar] [CrossRef]
- Donnelly-Roberts, D.L.; Namovic, M.T.; Han, P.; Jarvis, M.F. Mammalian P2X7 receptor pharmacology: Comparison of recombinant mouse, rat and human P2X7 receptors. Br. J. Pharmacol. 2009, 157, 1203–1214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solini, A.; Santini, E.; Chimenti, D.; Chiozzi, P.; Pratesi, F.; Cuccato, S.; Falzoni, S.; Lupi, R.; Ferrannini, E.; Pugliese, G.; et al. Multiple P2X receptors are involved in the modulation of apoptosis in human mesangial cells: Evidence for a role of P2X4. Am. J. Physiol.—Ren. Physiol. 2007, 292, 1537–1547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adriouch, S.; Hubert, S.; Pechberty, S.; Koch-Nolte, F.; Haag, F.; Seman, M. NAD + Released during Inflammation Participates in T Cell Homeostasis by Inducing ART2-Mediated Death of Naive T Cells In Vivo. J. Immunol. 2007, 179, 186–194. [Google Scholar] [CrossRef] [Green Version]
- Sanz, J.M.; Chiozzi, P.; Ferrari, D.; Colaianna, M.; Idzko, M.; Falzoni, S.; Fellin, R.; Trabace, L.; Di Virgilio, F. Activation of Microglia by Amyloid β Requires P2X 7 Receptor Expression. J. Immunol. 2009, 182, 4378–4385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elssner, A.; Duncan, M.; Gavrilin, M.; Wewers, M.D. A Novel P2X 7 Receptor Activator, the Human Cathelicidin-Derived Peptide LL37, Induces IL-1β Processing and Release. J. Immunol. 2004, 172, 4987–4994. [Google Scholar] [CrossRef] [Green Version]
- Ferrari, D.; Pizzirani, C.; Gulinelli, S.; Callegari, G.; Chiozzi, P.; Idzko, M.; Panther, E.; Di Virgilio, F. Modulation of P2X 7 receptor functions by polymyxin B: Crucial role of the hydrophobic tail of the antibiotic molecule. Br. J. Pharmacol. 2007, 150, 445–454. [Google Scholar] [CrossRef] [Green Version]
- Greve, A.S.; Skals, M.; Fagerberg, S.K.; Tonnus, W.; Ellermann-Eriksen, S.; Evans, R.J.; Linkermann, A.; Praetorius, H.A. P2X1, P2X4, and P2X7 Receptor Knock Out Mice Expose Differential Outcome of Sepsis Induced by α-Haemolysin Producing Escherichia coli. Front. Cell. Infect. Microbiol. 2017, 7, 113. [Google Scholar] [CrossRef] [Green Version]
- Allsopp, R.C.; Dayl, S.; Schmid, R.; Evans, R.J. Unique residues in the ATP gated human P2X7 receptor define a novel allosteric binding pocket for the selective antagonist AZ10606120. Sci. Rep. 2017, 7, 725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allsopp, R.C.; Dayl, S.; Dayel, A.B.; Schmid, R.; Evans, R.J. Mapping the allosteric action of antagonists A740003 and A438079 reveals a role for the left flipper in ligand sensitivity at P2X7 receptorss. Mol. Pharmacol. 2018, 93, 553–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartlett, R.; Stokes, L.; Sluyter, R. The p2x7 receptor channel: Recent developments and the use of p2x7 antagonists in models of disease. Pharmacol. Rev. 2014, 66, 638–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karoutzou, O.; Kwak, S.H.; Lee, S.D.; Martínez-Falguera, D.; Sureda, F.X.; Vázquez, S.; Kim, Y.C.; Barniol-Xicota, M. Towards a Novel Class of Multitarget-Directed Ligands: Dual P2X7–NMDA Receptor Antagonists. Molecules 2018, 23, 230. [Google Scholar] [CrossRef] [Green Version]
- Chhabra, P.; Linden, J.; Lobo, P.; Douglas Okusa, M.; Lewis Brayman, K. The Immunosuppressive Role of Adenosine A2A Receptors in Ischemia Reperfusion Injury and Islet Transplantation. Curr. Diabetes Rev. 2012, 8, 419–433. [Google Scholar] [CrossRef] [Green Version]
- Kan, L.K.; Seneviratne, S.; Drummond, K.J.; Williams, D.A.; O’Brien, T.J.; Monif, M. P2X7 receptor antagonism inhibits tumour growth in human high-grade gliomas. Purinergic Signal. 2020, 16, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Tognon, M.; Tagliapietra, A.; Magagnoli, F.; Mazziotta, C.; Oton-Gonzalez, L.; Lanzillotti, C.; Vesce, F.; Contini, C.; Rotondo, J.C.; Martini, F.; et al. Investigation on Spontaneous Abortion and Human Papillomavirus Infection. Vaccines 2020, 8, 473. [Google Scholar] [CrossRef]
- Tognon, M.; Luppi, M.; Corallini, A.; Taronna, A.; Barozzi, P.; Rotondo, J.C.; Comar, M.; Casali, M.V.; Bovenzi, M.; D’Agostino, A.; et al. Immunologic evidence of a strong association between non-Hodgkin lymphoma and simian virus 40. Cancer 2015, 121, 2618–2626. [Google Scholar] [CrossRef]
- Mazziotta, C.; Pellielo, G.; Tognon, M.; Martini, F.; Rotondo, J.C. Significantly low levels of IgG antibodies against oncogenic Merkel cell polyomavirus in sera from females affected by spontaneous abortion. Front. Microbiol. 2021, 12, 789991. [Google Scholar] [CrossRef] [PubMed]
- Rotondo, J.C.; Mazzoni, E.; Bononi, I.; Tognon, M.G.; Martini, F. Association Between Simian Virus 40 and Human Tumors. Front. Oncol. 2019, 9, 670. [Google Scholar] [CrossRef]
- Mazziotta, C.; Rotondo, J.C.; Lanzillotti, C.; Campione, G.; Martini, F.; Tognon, M. Cancer biology and molecular genetics of A3 adenosine receptor. Oncogene 2021, 41, 301–308. [Google Scholar] [CrossRef]
- Vincenzi, F.; Rotondo, J.C.; Pasquini, S.; Di Virgilio, F.; Varani, K.; Tognon, M. A3 Adenosine and P2X7 Purinergic Receptors as New Targets for an Innovative Pharmacological Therapy of Malignant Pleural Mesothelioma. Front. Oncol. 2021, 11, 3918. [Google Scholar] [CrossRef] [PubMed]
- Drill, M.; Powell, K.L.; Kan, L.K.; Jones, N.C.; O’Brien, T.J.; Hamilton, J.A.; Monif, M. Inhibition of purinergic P2X receptor 7 (P2X7R) decreases granulocyte-macrophage colony-stimulating factor (GM-CSF) expression in U251 glioblastoma cells. Sci. Rep. 2020, 10, 14844. [Google Scholar] [CrossRef]
- Giannuzzo, A.; Saccomano, M.; Napp, J.; Ellegaard, M.; Alves, F.; Novak, I. Targeting of the P2X7 receptor in pancreatic cancer and stellate cells. Int. J. Cancer 2016, 139, 2540–2552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.J.; Luo, C.; Huang, C.; Pu, F.Q.; Zhu, J.F.; Zhu, Z. ming PI3K/Akt/GSK-3β signal pathway is involved in P2X7 receptor-induced proliferation and EMT of colorectal cancer cells. Eur. J. Pharmacol. 2021, 899, 174041. [Google Scholar] [CrossRef] [PubMed]
- Talreja, O.; Kerndt, C.C.; Cassagnol, M. Simvastatin. xPharm Compr. Pharmacol. Ref. 2021, 1–4. [Google Scholar]
- Stock, T.C.; Bloom, B.J.; Wei, N.; Ishaq, S.; Park, W.; Wang, X.; Gupta, P.; Mebus, C.A. Efficacy and Safety of CE-224,535, an Antagonist of P2X7 Receptor, in Treatment of Patients with Rheumatoid Arthritis Inadequately Controlled by Methotrexate. J. Rheumatol. 2012, 39, 720–727. [Google Scholar] [CrossRef]
- McInnes, I.B.; Schett, G. Cytokines in the pathogenesis of rheumatoid arthritis. Nat. Rev. Immunol. 2007, 7, 429–442. [Google Scholar] [CrossRef]
- Drill, M.; Jones, N.C.; Hunn, M.; O’Brien, T.J.; Monif, M. Antagonism of the ATP-gated P2X7 receptor: A potential therapeutic strategy for cancer. Purinergic Signal. 2021, 17, 215–227. [Google Scholar] [CrossRef]
- Deng, Y.; Zhou, M.; Zhao, X.; Xue, X.; Liao, L.; Wang, J.; Li, Y. Immune response studies based on P2X7 receptors: A Mini-Review. Curr. Pharm. Des. 2022, 28, 35100953. [Google Scholar] [CrossRef]
- Sheng, D.; Hattori, M. Recent progress in the structural biology of P2X receptors. Proteins Struct. Funct. Bioinform. 2022. [Google Scholar] [CrossRef]


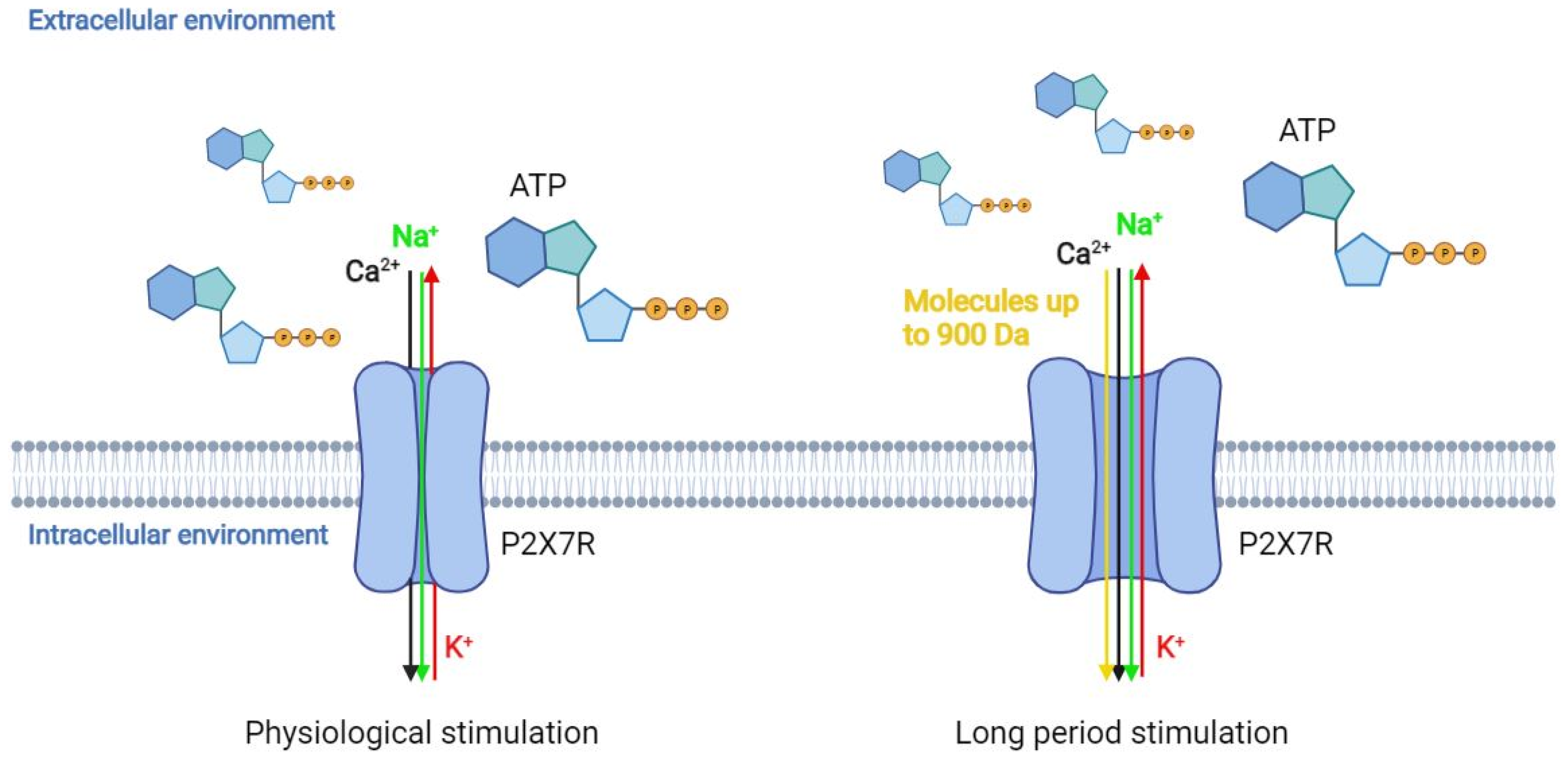{kind=link}
| dbSNP ID | Base | Amino Acid | Effect on | Implicated Conditions |
|---|---|---|---|---|
| Substitution | Substitution | P2X7 Function | ||
| rsl7525809 | 370T > C | A76V | Loss | Multiple sclerosis |
| rs28360447 | 474G > A | G150R | Loss | Osteoporosis |
| rs208294 | 489C > T | H155Y | Gain | Multiple sclerosis, chronic pain, severe sepsis, children’s febrile seizures |
| rs7958311 | 835G > A | R270H | Loss | Chronic pain |
| rs28360457 | 946G > A | R307Q | Loss | Osteoporosis |
| rs1718119 | 1068G > A | A348T | Gain | Osteoporosis, anxiety disorder, toxoplasmosis |
| rs2230911 | 1096C > G | T357S | Loss | Osteoporosis |
| rs2230912 | 1405A > G | Q460R | Loss | Osteoporosis, severe sepsis, bipolar disorders, major depressive disorders |
| rs3751143 | 1513A > C | E496A | Loss | Osteoporosis, tuberculosis, cardiovascular risks |
| rs2230913 | 1563C > G | H521Q | Neutral | - |
| rs1653624 | 1729T > A | I568N | Loss | Osteoporosis |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rotondo, J.C.; Mazziotta, C.; Lanzillotti, C.; Stefani, C.; Badiale, G.; Campione, G.; Martini, F.; Tognon, M. The Role of Purinergic P2X7 Receptor in Inflammation and Cancer: Novel Molecular Insights and Clinical Applications. Cancers 2022, 14, 1116. https://doi.org/10.3390/cancers14051116
Rotondo JC, Mazziotta C, Lanzillotti C, Stefani C, Badiale G, Campione G, Martini F, Tognon M. The Role of Purinergic P2X7 Receptor in Inflammation and Cancer: Novel Molecular Insights and Clinical Applications. Cancers. 2022; 14(5):1116. https://doi.org/10.3390/cancers14051116
Chicago/Turabian StyleRotondo, John Charles, Chiara Mazziotta, Carmen Lanzillotti, Chiara Stefani, Giada Badiale, Giulia Campione, Fernanda Martini, and Mauro Tognon. 2022. "The Role of Purinergic P2X7 Receptor in Inflammation and Cancer: Novel Molecular Insights and Clinical Applications" Cancers 14, no. 5: 1116. https://doi.org/10.3390/cancers14051116