
Low Spike Antibody Levels and Impaired BA.4/5 Neutralization in Patients with Multiple Myeloma or Waldenstrom’s Macroglobulinemia after BNT162b2 Booster Vaccination

 , , , , , and
, , , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients and Controls
2.2. SARS-CoV-2 Antibody Measurements
2.3. Statistical Analysis
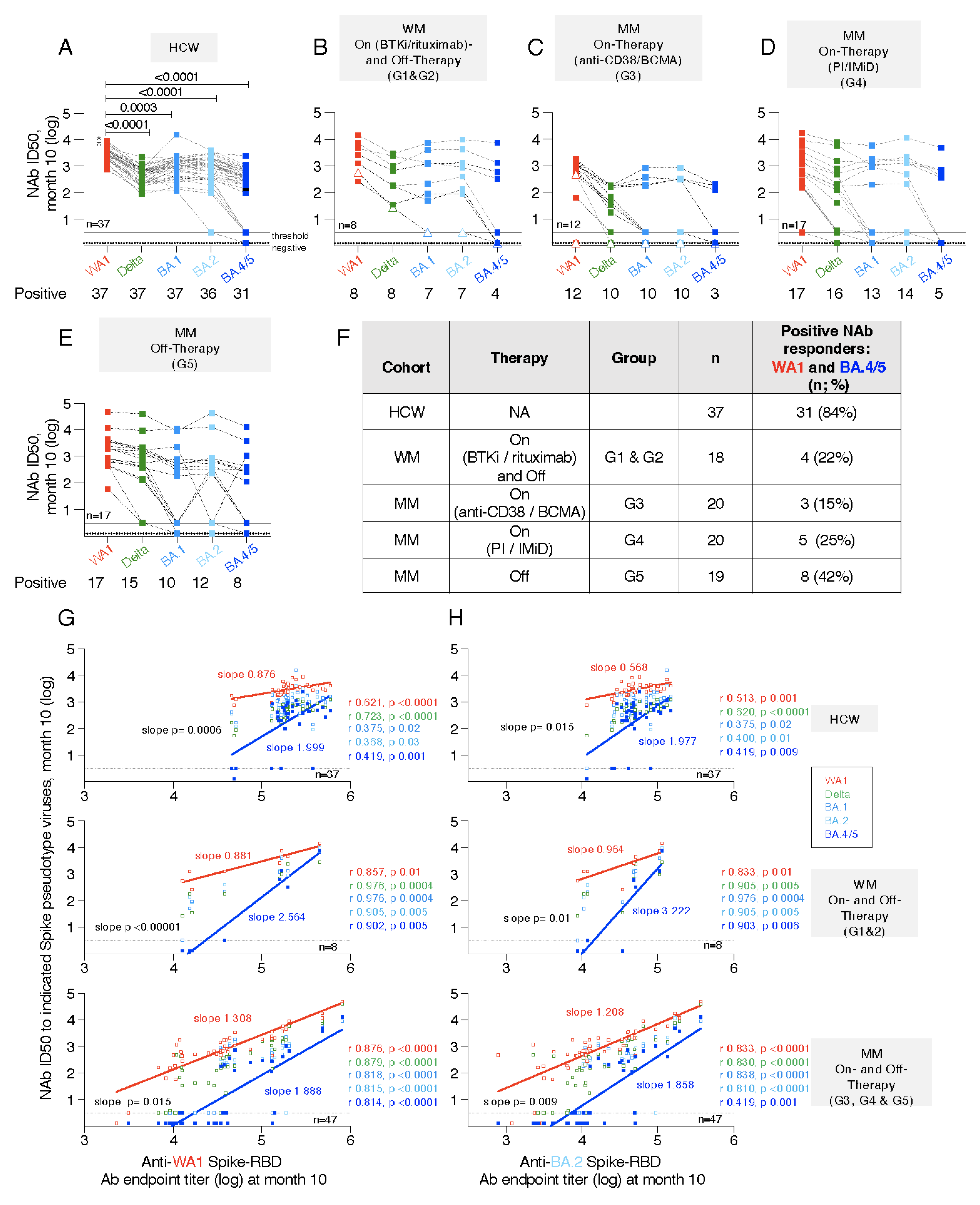
3. Results
3.1. Patients’ Characteristics
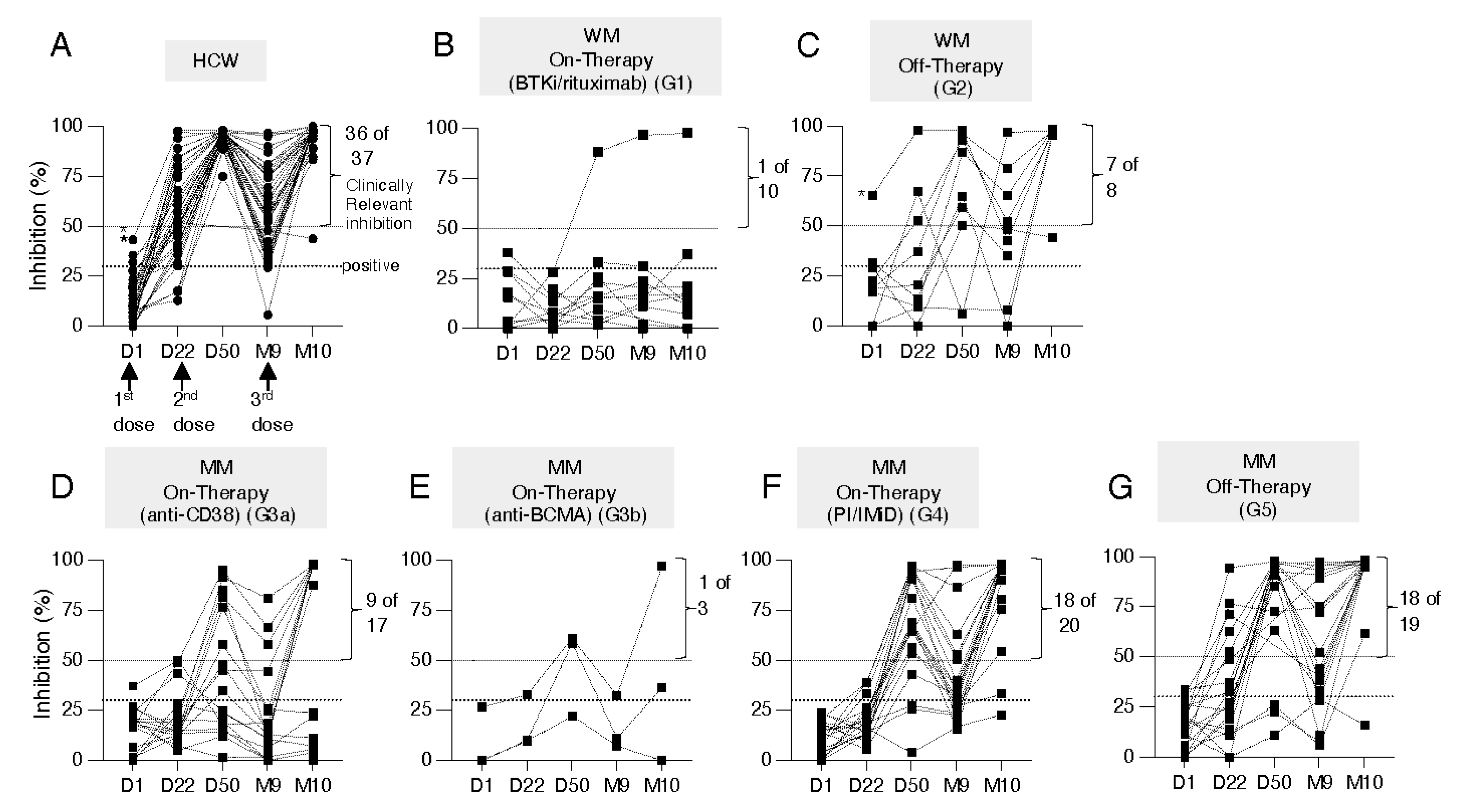
3.2. Kinetics of Development of Neutralizing Anti-WA1 Spike Antibodies
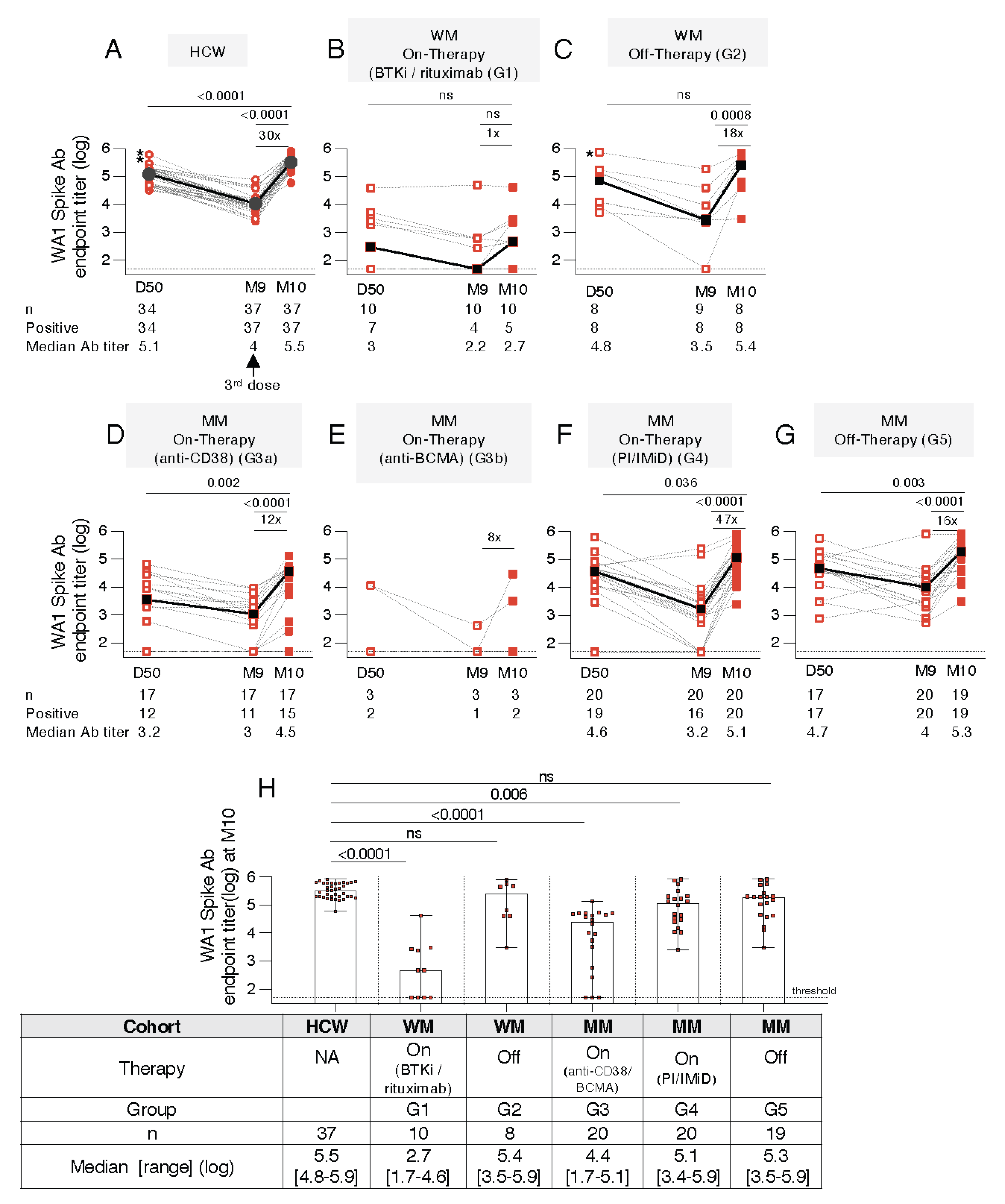
3.3. Distinct Anti-Spike Antibody Responses to the BNT162b2 mRNA Vaccine in WM and MM Patients Compared to HCW
3.4. Similar Anti-Spike Antibody Response Breadth to the BNT162b2 mRNA Vaccine in WM and MM Patients Compared to HCW
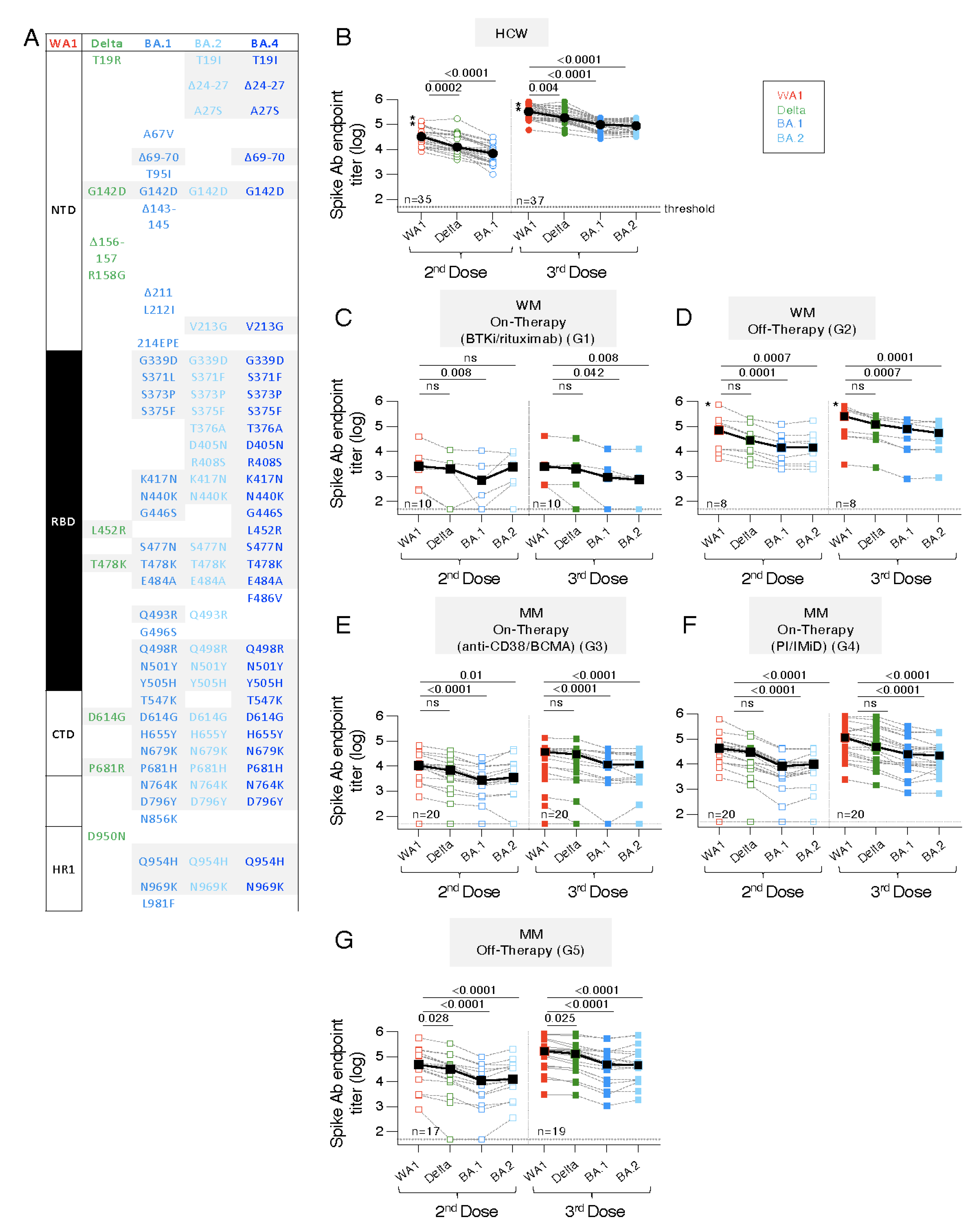
3.5. Distinct Ab Magnitude to Spike and Spike-RBD Proteins of WA1, Delta, BA.1 and BA.2
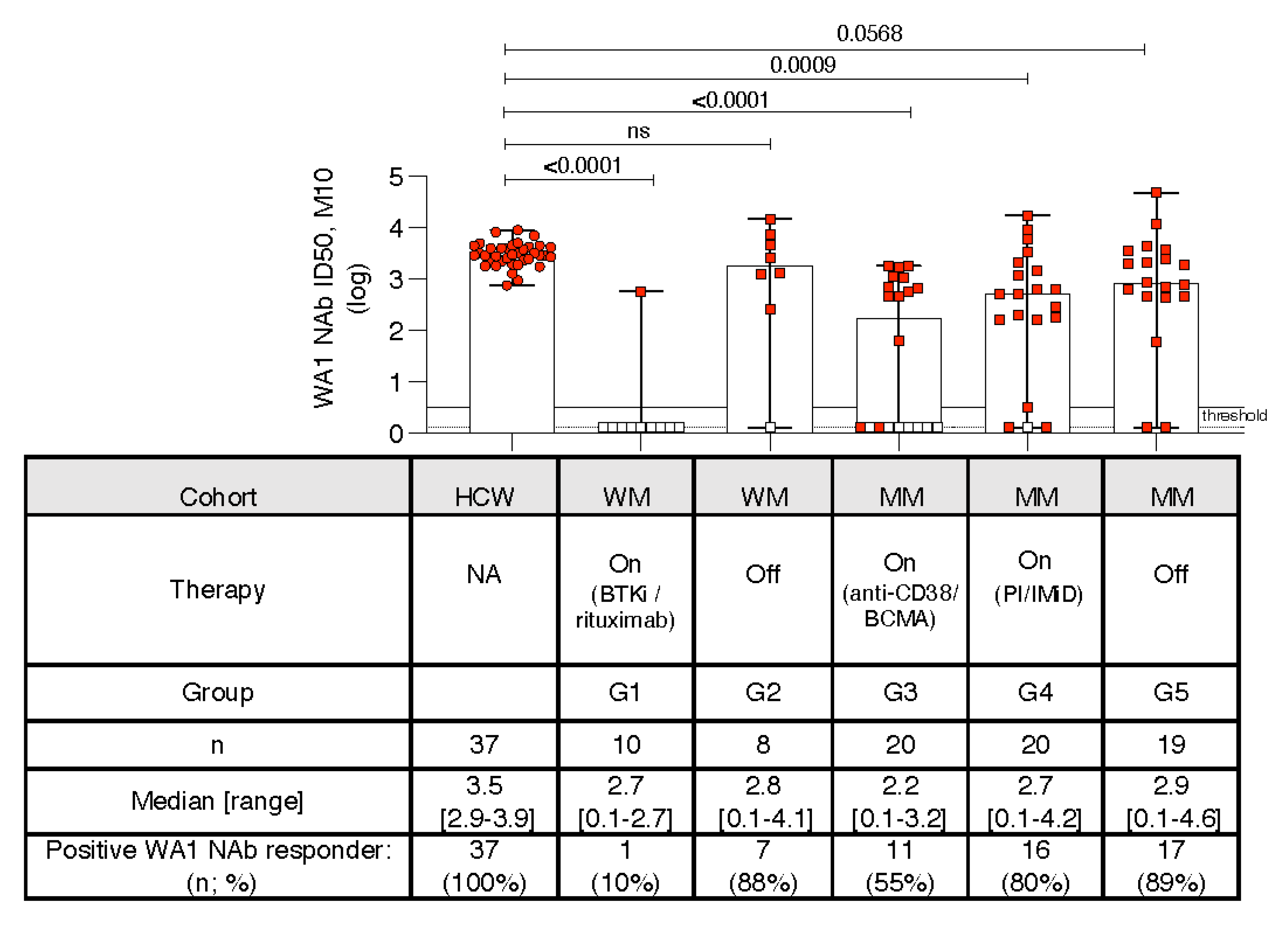
3.6. Kinetics of Development of Neutralizing Anti-WA1 Spike Ab
3.7. Neutralization Breadth with Impaired Responses to BA.4/5
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Terpos, E.; Gavriatopoulou, M.; Ntanasis-Stathopoulos, I.; Briasoulis, A.; Gumeni, S.; Malandrakis, P.; Fotiou, D.; Papanagnou, E.D.; Migkou, M.; Theodorakakou, F.; et al. The neutralizing antibody response post COVID-19 vaccination in patients with myeloma is highly dependent on the type of anti-myeloma treatment. Blood Cancer J. 2021, 11, 138. [Google Scholar] [CrossRef] [PubMed]
- Terpos, E.; Trougakos, I.P.; Gavriatopoulou, M.; Papassotiriou, I.; Sklirou, A.D.; Ntanasis-Stathopoulos, I.; Papanagnou, E.D.; Fotiou, D.; Kastritis, E.; Dimopoulos, M.A. Low neutralizing antibody responses against SARS-CoV-2 in older patients with myeloma after the first BNT162b2 vaccine dose. Blood 2021, 137, 3674–3676. [Google Scholar] [CrossRef] [PubMed]
- Aleman, A.; Upadhyaya, B.; Tuballes, K.; Kappes, K.; Gleason, C.R.; Beach, K.; Agte, S.; Srivastava, K.; Group, P.V.S.S.; van Oekelen, O.; et al. Variable cellular responses to SARS-CoV-2 in fully vaccinated patients with multiple myeloma. Cancer Cell 2021, 39, 1442–1444. [Google Scholar] [CrossRef] [PubMed]
- Terpos, E.; Gavriatopoulou, M.; Ntanasis-Stathopoulos, I.; Briasoulis, A.; Gumeni, S.; Malandrakis, P.; Papanagnou, E.D.; Migkou, M.; Kanellias, N.; Kastritis, E.; et al. Booster BNT162b2 optimizes SARS-CoV-2 humoral response in patients with myeloma: The negative effect of anti-BCMA therapy. Blood 2022, 139, 1409–1412. [Google Scholar] [CrossRef]
- Aleman, A.; Van Oekelen, O.; Upadhyaya, B.; Beach, K.; Kogan Zajdman, A.; Alshammary, H.; Serebryakova, K.; Agte, S.; Kappes, K.; Gleason, C.R.; et al. Augmentation of humoral and cellular immune responses after third-dose SARS-CoV-2 vaccination and viral neutralization in myeloma patients. Cancer Cell 2022, 40, 441–443. [Google Scholar] [CrossRef]
- Gavriatopoulou, M.; Terpos, E.; Ntanasis-Stathopoulos, I.; Briasoulis, A.; Gumeni, S.; Malandrakis, P.; Fotiou, D.; Migkou, M.; Theodorakakou, F.; Eleutherakis-Papaiakovou, E.; et al. Poor neutralizing antibody responses in 106 patients with WM after vaccination against SARS-CoV-2: A prospective study. Blood Adv. 2021, 5, 4398–4405. [Google Scholar] [CrossRef]
- Ludwig, H.; Sonneveld, P.; Facon, T.; San-Miguel, J.; Avet-Loiseau, H.; Mohty, M.; Mateos, M.V.; Moreau, P.; Cavo, M.; Pawlyn, C.; et al. COVID-19 vaccination in patients with multiple myeloma: A consensus of the European Myeloma Network. Lancet Haematol. 2021, 8, e934–e946. [Google Scholar] [CrossRef]
- Terpos, E.; Fotiou, D.; Karalis, V.; Ntanasis-Stathopoulos, I.; Sklirou, A.; Gavriatopoulou, M.; Malandrakis, P.; Iconomidou, V.; Kastritis, E.; Trougakos, I.; et al. SARS-CoV-2 humoral responses following booster BNT162b2 vaccination in patients with B-cell malignancies. Am. J. Hematol. Oncol. 2022, 10, 1002. [Google Scholar] [CrossRef]
- Enssle, J.C.; Campe, J.; Buchel, S.; Moter, A.; See, F.; Griessbaum, K.; Rieger, M.A.; Wolf, S.; Ballo, O.; Steffen, B.; et al. Enhanced but variant-dependent serological and cellular immune responses to third-dose BNT162b2 vaccination in patients with multiple myeloma. Cancer Cell 2022, 40, 587–589. [Google Scholar] [CrossRef]
- Stampfer, S.D.; Goldwater, M.S.; Bujarski, S.; Regidor, B.; Zhang, W.; Feinstein, A.J.; Swift, R.; Eshaghian, S.; Vail, E.; Berenson, J.R. Severe breakthrough COVID-19 with a heavily mutated variant in a multiple myeloma patient 10 weeks after vaccination. Clin. Infect. Pract. 2022, 13, 100130. [Google Scholar] [CrossRef]
- Chang, A.; Akhtar, A.; Linderman, S.L.; Lai, L.; Orellana-Noia, V.M.; Valanparambil, R.; Ahmed, H.; Zarnitsyna, V.I.; McCook-Veal, A.A.; Switchenko, J.M.; et al. Humoral Responses Against SARS-CoV-2 and Variants of Concern After mRNA Vaccines in Patients with Non-Hodgkin Lymphoma and Chronic Lymphocytic Leukemia. J. Clin. Oncol. 2022, 40, 3020–3031. [Google Scholar] [CrossRef]
- Bergamaschi, C.; Pagoni, M.; Rosati, M.; Angel, M.; Tzannou, I.; Vlachou, M.; Ismini Darmani, I.; Ullah, A.; Bear, J.; Devasundaram, S.; et al. Reduced antibodies and innate cytokine changes in SARS-CoV-2 BNT162b2 mRNA vaccinated transplant patients with hematological malignancies. Front. Immunol. 2022, 13, 899972. [Google Scholar] [CrossRef] [PubMed]
- Madhi, S.A.; Kwatra, G.; Myers, J.E.; Jassat, W.; Dhar, N.; Mukendi, C.K.; Nana, A.J.; Blumberg, L.; Welch, R.; Ngorima-Mabhena, N.; et al. Population Immunity and COVID-19 Severity with Omicron Variant in South Africa. N. Engl. J. Med. 2022, 386, 1314–1326. [Google Scholar] [CrossRef]
- Klompas, M.; Pandolfi, M.C.; Nisar, A.B.; Baker, M.A.; Rhee, C. Association of Omicron vs Wild-type SARS-CoV-2 Variants with Hospital-Onset SARS-CoV-2 Infections in a US Regional Hospital System. JAMA 2022, 328, 296–298. [Google Scholar] [CrossRef]
- McCallum, M.; Walls, A.C.; Sprouse, K.R.; Bowen, J.E.; Rosen, L.E.; Dang, H.V.; De Marco, A.; Franko, N.; Tilles, S.W.; Logue, J.; et al. Molecular basis of immune evasion by the Delta and Kappa SARS-CoV-2 variants. Science 2021, 374, 1621–1626. [Google Scholar] [CrossRef] [PubMed]
- Cameroni, E.; Bowen, J.E.; Rosen, L.E.; Saliba, C.; Zepeda, S.K.; Culap, K.; Pinto, D.; VanBlargan, L.A.; De Marco, A.; di Iulio, J.; et al. Broadly neutralizing antibodies overcome SARS-CoV-2 Omicron antigenic shift. Nature 2022, 602, 664–670. [Google Scholar] [CrossRef]
- Schmidt, F.; Muecksch, F.; Weisblum, Y.; Da Silva, J.; Bednarski, E.; Cho, A.; Wang, Z.; Gaebler, C.; Caskey, M.; Nussenzweig, M.C.; et al. Plasma Neutralization of the SARS-CoV-2 Omicron Variant. N. Engl. J. Med. 2022, 386, 599–601. [Google Scholar] [CrossRef] [PubMed]
- Dejnirattisai, W.; Huo, J.; Zhou, D.; Zahradnik, J.; Supasa, P.; Liu, C.; Duyvesteyn, H.M.E.; Ginn, H.M.; Mentzer, A.J.; Tuekprakhon, A.; et al. SARS-CoV-2 Omicron-B.1.1.529 leads to widespread escape from neutralizing antibody responses. Cell 2022, 185, 467–484.e15. [Google Scholar] [CrossRef]
- Meng, B.; Abdullahi, A.; Ferreira, I.; Goonawardane, N.; Saito, A.; Kimura, I.; Yamasoba, D.; Gerber, P.P.; Fatihi, S.; Rathore, S.; et al. Altered TMPRSS2 usage by SARS-CoV-2 Omicron impacts infectivity and fusogenicity. Nature 2022, 603, 706–714. [Google Scholar] [CrossRef]
- Cele, S.; Jackson, L.; Khoury, D.S.; Khan, K.; Moyo-Gwete, T.; Tegally, H.; San, J.E.; Cromer, D.; Scheepers, C.; Amoako, D.G.; et al. Omicron extensively but incompletely escapes Pfizer BNT162b2 neutralization. Nature 2022, 602, 654–656. [Google Scholar] [CrossRef]
- Andrews, N.; Stowe, J.; Kirsebom, F.; Toffa, S.; Rickeard, T.; Gallagher, E.; Gower, C.; Kall, M.; Groves, N.; O’Connell, A.M.; et al. COVID-19 Vaccine Effectiveness against the Omicron (B.1.1.529) Variant. N. Engl. J. Med. 2022, 386, 1532–1546. [Google Scholar] [CrossRef] [PubMed]
- Pajon, R.; Doria-Rose, N.A.; Shen, X.; Schmidt, S.D.; O’Dell, S.; McDanal, C.; Feng, W.; Tong, J.; Eaton, A.; Maglinao, M.; et al. SARS-CoV-2 Omicron Variant Neutralization after mRNA-1273 Booster Vaccination. N. Engl. J. Med. 2022, 386, 1088–1091. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Kainulainen, M.H.; Jiang, N.; Di, H.; Bonenfant, G.; Mills, L.; Currier, M.; Shrivastava-Ranjan, P.; Calderon, B.M.; Sheth, M.; et al. Differential neutralization and inhibition of SARS-CoV-2 variants by antibodies elicited by COVID-19 mRNA vaccines. Nat. Commun. 2022, 13, 4350. [Google Scholar] [CrossRef] [PubMed]
- Bar-On, Y.M.; Goldberg, Y.; Mandel, M.; Bodenheimer, O.; Amir, O.; Freedman, L.; Alroy-Preis, S.; Ash, N.; Huppert, A.; Milo, R. Protection by a Fourth Dose of BNT162b2 against Omicron in Israel. N. Engl. J. Med. 2022, 386, 1712–1720. [Google Scholar] [CrossRef] [PubMed]
- Hachmann, N.P.; Miller, J.; Collier, A.Y.; Ventura, J.D.; Yu, J.; Rowe, M.; Bondzie, E.A.; Powers, O.; Surve, N.; Hall, K.; et al. Neutralization Escape by SARS-CoV-2 Omicron Subvariants BA.2.12.1, BA.4, and BA.5. N. Engl. J. Med. 2022, 387, 86–88. [Google Scholar] [CrossRef] [PubMed]
- Rosati, M.; Agarwal, M.; Hu, X.; Devasundaram, S.; Stellas, D.; Chowdhury, B.; Bear, J.; Burns, R.; Donohue, D.; Pessaint, L.; et al. Control of SARS-CoV-2 infection after Spike DNA or Spike DNA + Protein co-immunization in rhesus macaques. PLoS Pathog. 2021, 17, e1009701. [Google Scholar] [CrossRef]
- Rosati, M.; Terpos, E.; Ntanasis-Stathopoulos, I.; Agarwal, M.; Bear, J.; Burns, R.; Hu, X.; Korompoki, E.; Donohue, D.; Venzon, D.J.; et al. Sequential Analysis of Binding and Neutralizing Antibody in COVID-19 Convalescent Patients at 14 months after SARS-CoV-2 infection. Front. Immunol. 2021, 12, 793953. [Google Scholar] [CrossRef]
- Walsh, E.E.; Frenck, R.W., Jr.; Falsey, A.R.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Neuzil, K.; Mulligan, M.J.; Bailey, R.; et al. Safety and Immunogenicity of Two RNA-Based COVID-19 Vaccine Candidates. N. Engl. J. Med. 2020, 383, 2439–2450. [Google Scholar] [CrossRef]
- Tan, C.W.; Chia, W.N.; Qin, X.; Liu, P.; Chen, M.I.; Tiu, C.; Hu, Z.; Chen, V.C.; Young, B.E.; Sia, W.R.; et al. A SARS-CoV-2 surrogate virus neutralization test based on antibody-mediated blockage of ACE2-spike protein-protein interaction. Nat. Biotechnol. 2020, 38, 1073–1078. [Google Scholar] [CrossRef]
- Robbiani, D.F.; Gaebler, C.; Muecksch, F.; Lorenzi, J.C.C.; Wang, Z.; Cho, A.; Agudelo, M.; Barnes, C.O.; Gazumyan, A.; Finkin, S.; et al. Convergent antibody responses to SARS-CoV-2 in convalescent individuals. Nature 2020, 584, 437–442. [Google Scholar] [CrossRef]
- Schmidt, F.; Weisblum, Y.; Muecksch, F.; Hoffmann, H.H.; Michailidis, E.; Lorenzi, J.C.C.; Mendoza, P.; Rutkowska, M.; Bednarski, E.; Gaebler, C.; et al. Measuring SARS-CoV-2 neutralizing antibody activity using pseudotyped and chimeric viruses. J. Exp. Med. 2020, 217, e20201181. [Google Scholar] [CrossRef] [PubMed]
- Bergamaschi, C.; Terpos, E.; Rosati, M.; Angel, M.; Bear, J.; Stellas, D.; Karaliota, S.; Apostolakou, F.; Bagratuni, T.; Patseas, D.; et al. Systemic IL-15, IFN-gamma, and IP-10/CXCL10 signature associated with effective immune response to SARS-CoV-2 in BNT162b2 mRNA vaccine recipients. Cell Rep. 2021, 36, 109504. [Google Scholar] [CrossRef] [PubMed]
- Bowen, J.E.; Addetia, A.; Dang, H.V.; Stewart, C.; Brown, J.T.; Sharkey, W.K.; Sprouse, K.R.; Walls, A.C.; Mazzitelli, I.G.; Logue, J.K.; et al. Omicron spike function and neutralizing activity elicited by a comprehensive panel of vaccines. Science 2022, 377, 890–894. [Google Scholar] [CrossRef] [PubMed]
- Qu, P.; Faraone, J.; Evans, J.P.; Zou, X.; Zheng, Y.M.; Carlin, C.; Bednash, J.S.; Lozanski, G.; Mallampalli, R.K.; Saif, L.J.; et al. Neutralization of the SARS-CoV-2 Omicron BA.4/5 and BA.2.12.1 Subvariants. N. Engl. J. Med. 2022, 386, 2526–2528. [Google Scholar] [CrossRef]
- Gaebler, C.; DaSilva, J.; Bednarski, E.; Muecksch, F.; Schmidt, F.; Weisblum, Y.; Millard, K.G.; Turroja, M.; Cho, A.; Wang, Z.; et al. Severe Acute Respiratory Syndrome Coronavirus 2 Neutralization After Messenger RNA Vaccination and Variant Breakthrough Infection. Open Forum Infect. Dis. 2022, 9, ofac227. [Google Scholar] [CrossRef]
- Ntanasis-Stathopoulos, I.; Karalis, V.; Gavriatopoulou, M.; Malandrakis, P.; Sklirou, A.; Eleutherakis-Papaiakovou, E.; Migkou, M.; Roussou, M.; Fotiou, D.; Alexopoulos, H.; et al. Second booster BNT162b2 restores SARS-CoV-2 humoral response in patients with multiple myeloma, excluding those under anti-BCMA therapy. Hemasphere 2022, 7, e747. [Google Scholar] [CrossRef]
- Haggenburg, S.; Hofsink, Q.; Lissenberg-Witte, B.I.; Broers, A.E.C.; van Doesum, J.A.; van Binnendijk, R.S.; den Hartog, G.; Bhoekhan, M.S.; Haverkate, N.J.E.; Burger, J.A.; et al. Antibody Response in Immunocompromised Patients with Hematologic Cancers Who Received a 3-Dose mRNA-1273 Vaccination Schedule for COVID-19. JAMA Oncol. 2022, 8, 1477–1483. [Google Scholar] [CrossRef]
- Terpos, E.; Politou, M.; Sergentanis, T.N.; Mentis, A.; Rosati, M.; Stellas, D.; Bear, J.; Hu, X.; Felber, B.K.; Pappa, V.; et al. Anti-SARS-CoV-2 Antibody Responses in Convalescent Plasma Donors Are Increased in Hospitalized Patients; Subanalyses of a Phase 2 Clinical Study. Microorganisms 2020, 8, 1885. [Google Scholar] [CrossRef]
- Terpos, E.; Stellas, D.; Rosati, M.; Sergentanis, T.N.; Hu, X.; Politou, M.; Pappa, V.; Ntanasis-Stathopoulos, I.; Karaliota, S.; Bear, J.; et al. SARS-CoV-2 Antibody Kinetics Eight Months from COVID-19 Onset: Persistence of Spike Antibodies but Loss of Neutralizing Antibodies in 24% of Convalescent Plasma Donors. Eur. J. Intern. Med. 2021, 89, 87–96. [Google Scholar] [CrossRef]
- Haggenburg, S.; Lissenberg-Witte, B.I.; van Binnendijk, R.S.; den Hartog, G.; Bhoekhan, M.S.; Haverkate, N.J.E.; de Rooij, D.M.; van Meerloo, J.; Cloos, J.; Kootstra, N.A.; et al. Quantitative analysis of mRNA-1273 COVID-19 vaccination response in immunocompromised adult hematology patients. Blood Adv. 2022, 6, 1537–1546. [Google Scholar] [CrossRef]
- Obeid, M.; Suffiotti, M.; Pellaton, C.; Bouchaab, H.; Cairoli, A.; Salvade, V.; Stevenel, C.; Hottinger, R.; Pythoud, C.; Coutechier, L.; et al. Humoral Responses Against Variants of Concern by COVID-19 mRNA Vaccines in Immunocompromised Patients. JAMA Oncol. 2022, 8, e220446. [Google Scholar] [CrossRef] [PubMed]
- Beaton, B.; Sasson, S.C.; Rankin, K.; Raedemaeker, J.; Wong, A.; Hastak, P.; Phetsouphanh, C.; Warden, A.; Klemm, V.; Munier, C.M.L.; et al. Patients with treated indolent lymphomas immunized with BNT162b2 have reduced anti-spike neutralizing IgG to SARS-CoV-2 variants, but preserved antigen-specific T cell responses. Am. J. Hematol. 2022. online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Zaleska, J.; Kwasnik, P.; Paziewska, M.; Purkot, J.; Szabelak, A.; Jurek, M.; Masny, N.; Dziatkiewicz, I.; Pronobis-Szczylik, B.; Piebiak, A.; et al. Response to anti-SARS-CoV-2 mRNA vaccines in multiple myeloma and chronic lymphocytic leukemia patients. Int. J. Cancer 2022. online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Re, D.; Seitz-Polski, B.; Brglez, V.; Carles, M.; Graca, D.; Benzaken, S.; Liguori, S.; Zahreddine, K.; Delforge, M.; Bailly-Maitre, B.; et al. Humoral and cellular responses after a third dose of SARS-CoV-2 BNT162b2 vaccine in patients with lymphoid malignancies. Nat. Commun. 2022, 13, 864. [Google Scholar] [CrossRef]
- Ramasamy, K.; Sadler, R.; Jeans, S.; Weeden, P.; Varghese, S.; Turner, A.; Larham, J.; Gray, N.; Carty, O.; Barrett, J.; et al. Immune response to COVID-19 vaccination is attenuated by poor disease control and antimyeloma therapy with vaccine driven divergent T-cell response. Br. J. Haematol. 2022, 197, 293–301. [Google Scholar] [CrossRef]
- Enssle, J.C.; Campe, J.; Schwenger, A.; Wiercinska, E.; Hellstern, H.; Durrwald, R.; Rieger, M.A.; Wolf, S.; Ballo, O.; Steffen, B.; et al. Severe impairment of T-cell responses to BNT162b2 immunization in patients with multiple myeloma. Blood 2022, 139, 137–142. [Google Scholar] [CrossRef]
- Lyke, K.E.; Atmar, R.L.; Islas, C.D.; Posavad, C.M.; Szydlo, D.; Paul Chourdhury, R.; Deming, M.E.; Eaton, A.; Jackson, L.A.; Branche, A.R.; et al. Rapid decline in vaccine-boosted neutralizing antibodies against SARS-CoV-2 Omicron variant. Cell Rep. Med. 2022, 3, 100679. [Google Scholar] [CrossRef]
- Terpos, E.; Karalis, V.; Ntanasis-Stathopoulos, I.; Evangelakou, Z.; Gavriatopoulou, M.; Manola, M.S.; Malandrakis, P.; Gianniou, D.D.; Kastritis, E.; Trougakos, I.P.; et al. Comparison of Neutralizing Antibody Responses at 6 Months Post Vaccination with BNT162b2 and AZD1222. Biomedicines 2022, 10, 338. [Google Scholar] [CrossRef]
- Suthar, M.S.; Arunachalam, P.S.; Hu, M.; Reis, N.; Trisal, M.; Raeber, O.; Chinthrajah, S.; Davis-Gardner, M.E.; Manning, K.; Mudvari, P.; et al. Durability of immune responses to the BNT162b2 mRNA vaccine. Medicina 2022, 3, 25–27. [Google Scholar] [CrossRef]
- Lassauniere, R.; Polacek, C.; Frische, A.; Boding, L.; Saekmose, S.G.; Rasmussen, M.; Fomsgaard, A. Neutralizing Antibodies Against the SARS-CoV-2 Omicron Variant (BA.1) 1 to 18 Weeks After the Second and Third Doses of the BNT162b2 mRNA Vaccine. JAMA Netw. Open 2022, 5, e2212073. [Google Scholar] [CrossRef]
- Priddy, F.H.; Williams, M.; Carson, S.; Lavender, B.; Mathieson, J.; Frampton, C.; Moreland, N.J.; McGregor, R.; Williams, G.; Brewerton, M.; et al. Immunogenicity of BNT162b2 COVID-19 vaccine in New Zealand adults. Vaccine 2022, 40, 5050–5059. [Google Scholar] [CrossRef] [PubMed]
- Stuver, R.; Shah, G.L.; Korde, N.S.; Roeker, L.E.; Mato, A.R.; Batlevi, C.L.; Chung, D.J.; Doddi, S.; Falchi, L.; Gyurkocza, B.; et al. Activity of AZD7442 (tixagevimab-cilgavimab) against Omicron SARS-CoV-2 in patients with hematologic malignancies. Cancer Cell 2022, 40, 590–591. [Google Scholar] [CrossRef] [PubMed]
- Marcacci, G.; Coppola, N.; Madonna, E.; Becchimanzi, C.; de Pascalis, S.; D’Ovidio, S.; Crisci, S.; Maiolino, P.; de Filippi, R.; Pinto, A. Post-exposure prophylaxis with sotrovimab for Omicron (B.1.1.529) SARS-CoV-2 variant during the aplastic phase of autologous stem cell transplantation. Infect. Agent Cancer 2022, 1, 41. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cancer | Patient Group (G) | Enrolled (n) | Active Therapy | Median Time Off-Therapy, Month [Range] | Gender (n) | Age | |
|---|---|---|---|---|---|---|---|
| Male | Female | Median [Range] | |||||
| WM | G1 | 11 | ON 1 | 4 | 7 | 70 | |
| [43–86] | |||||||
| WM 2 | G2 | 9 | OFF 3 | 30 | 3 | 6 | 82 |
| [10–86] | [62–88] | ||||||
| MM under anti-CD38-based regimen | G3a | 17 | ON | 8 | 9 | 69 | |
| [44–81] | |||||||
| MM under belantamab mafodotin (anti-BCMA) only regimen | G3b | 3 | ON | 2 | 1 | 67 | |
| [65–73] | |||||||
| MM under PI and/or IMiD treatments 4 | G4 | 20 | ON | 12 | 8 | 67 | |
| [48–86] | |||||||
| MM 5 | G5 | 20 | OFF 6 | 37 | 13 | 7 | 69.5 |
| [1–126] | [44–91] | ||||||
| Controls | |||||||
| HCW 7 | N/A 8 | 37 | N/A 8 | 13 | 24 | 54 | |
| [27–69] | |||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosati, M.; Terpos, E.; Bear, J.; Burns, R.; Devasundaram, S.; Ntanasis-Stathopoulos, I.; Gavriatopoulou, M.; Kastritis, E.; Dimopoulos, M.-A.; Pavlakis, G.N.; et al. Low Spike Antibody Levels and Impaired BA.4/5 Neutralization in Patients with Multiple Myeloma or Waldenstrom’s Macroglobulinemia after BNT162b2 Booster Vaccination. Cancers 2022, 14, 5816. https://doi.org/10.3390/cancers14235816
Rosati M, Terpos E, Bear J, Burns R, Devasundaram S, Ntanasis-Stathopoulos I, Gavriatopoulou M, Kastritis E, Dimopoulos M-A, Pavlakis GN, et al. Low Spike Antibody Levels and Impaired BA.4/5 Neutralization in Patients with Multiple Myeloma or Waldenstrom’s Macroglobulinemia after BNT162b2 Booster Vaccination. Cancers. 2022; 14(23):5816. https://doi.org/10.3390/cancers14235816
Chicago/Turabian StyleRosati, Margherita, Evangelos Terpos, Jenifer Bear, Robert Burns, Santhi Devasundaram, Ioannis Ntanasis-Stathopoulos, Maria Gavriatopoulou, Efstathios Kastritis, Meletios-Athanasios Dimopoulos, George N. Pavlakis, and et al. 2022. "Low Spike Antibody Levels and Impaired BA.4/5 Neutralization in Patients with Multiple Myeloma or Waldenstrom’s Macroglobulinemia after BNT162b2 Booster Vaccination" Cancers 14, no. 23: 5816. https://doi.org/10.3390/cancers14235816