

Omega-3 Polyunsaturated Fatty Acids, Gut Microbiota, Microbial Metabolites, and Risk of Colorectal Adenomas
Abstract
:Simple Summary
Abstract
1. Introduction
2. Methods
2.1. Study Population
2.2. Data Collection
2.2.1. Dietary and Lifestyle Exposures
2.2.2. Biopsy Sample Collection
2.2.3. DNA Extraction and Bacterial 16S rRNA Sequencing
2.2.4. Bacterial Identification
2.2.5. Bacteria Overall Composition and Diversity Measurements: Diversity, Evenness and Richness
2.2.6. Metabolome Assays
2.3. Statistical Analysis
3. Results
3.1. Study Participant Characteristics
3.2. Association between ω-3 PUFAs and Colorectal Adenomas
3.3. Association between Gut Microbiota Abundance and Colorectal Adenomas
3.4. Interactions between ω-3 PUFA and Gut Microbiota on Colorectal Adenoma
3.5. Association between BAs Concentration and Colorectal Adenomas
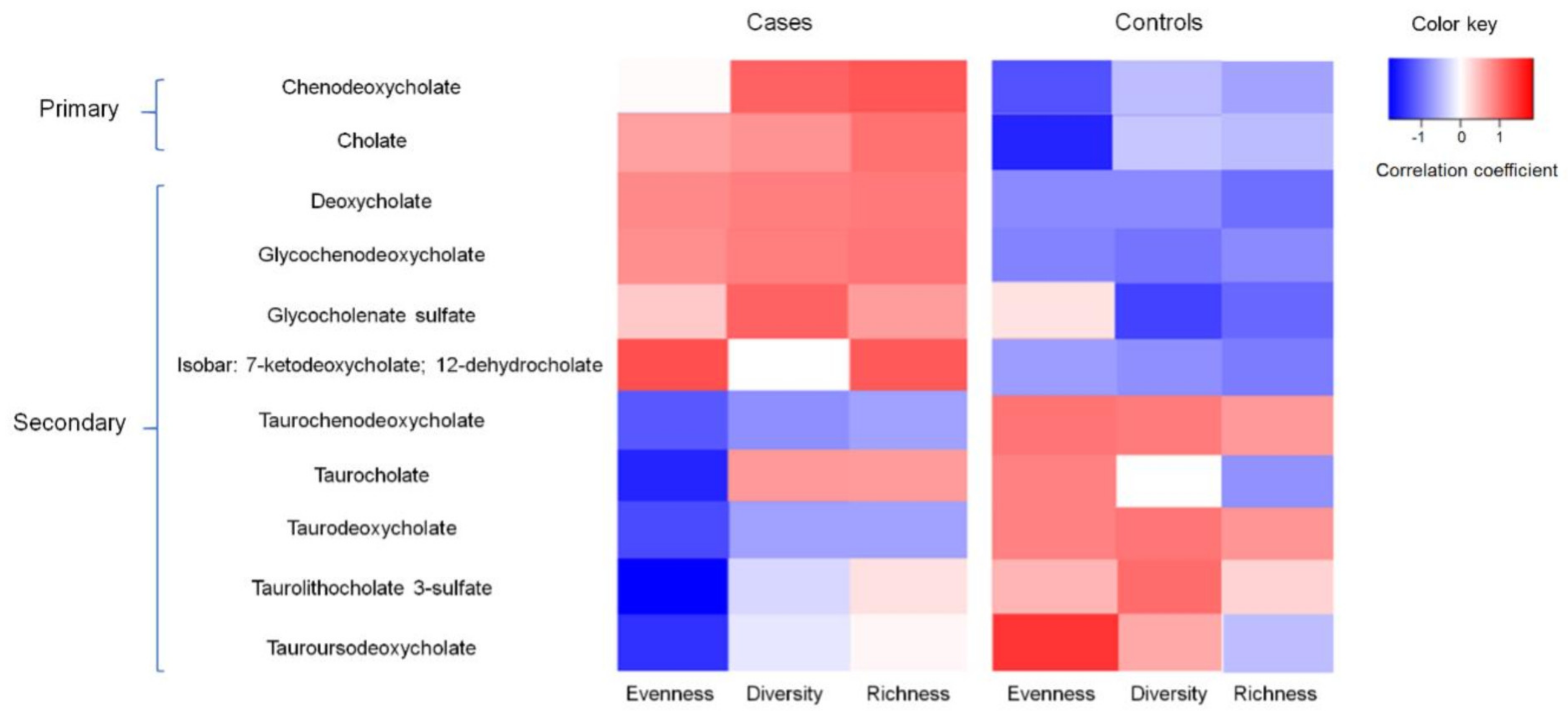
3.6. Associations between BAs, Gut Microbiota, and Adenomas in the Subset of Samples with Metabolomics Data
3.7. Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Stewart, S.L.; Wike, J.M.; Kato, I.; Lewis, D.R.; Michaud, F. A population-based study of colorectal cancer histology in the United States, 1998–2001. Cancer 2006, 107, 1128–1141. [Google Scholar] [CrossRef] [PubMed]
- Cockbain, A.J.; Toogood, G.J.; Hull, M.A. Omega-3 polyunsaturated fatty acids for the treatment and prevention of colorectal cancer. Gut 2012, 61, 135–149. [Google Scholar] [CrossRef] [PubMed]
- Larsson, S.C.; Kumlin, M.; Ingelman-Sundberg, M.; Wolk, A. Dietary long-chain n-3 fatty acids for the prevention of cancer: A review of potential mechanisms. Am. J. Clin. Nutr. 2004, 79, 935–945. [Google Scholar] [CrossRef]
- Turchini, G.M.; Nichols, P.D.; Barrow, C.; Sinclair, A.J. Jumping on the omega-3 bandwagon: Distinguishing the role of long-chain and short-chain omega-3 fatty acids. Crit. Rev. Food Sci. Nutr. 2012, 52, 795–803. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.C.; Qin, L.Q.; Lu, D.B.; Han, T.M.; Zheng, Y.; Xu, G.Z.; Wang, X.H. N-3 polyunsaturated fatty acids intake and risk of colorectal cancer: Meta-analysis of prospective studies. Cancer Causes Control 2015, 26, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Hull, M.A.; Sprange, K.; Hepburn, T.; Tan, W.; Shafayat, A.; Rees, C.J.; Clifford, G.; Logan, R.F.; Loadman, P.M.; Williams, E.A.; et al. Eicosapentaenoic acid and aspirin, alone and in combination, for the prevention of colorectal adenomas (seAFOod Polyp Prevention trial): A multicentre, randomised, double-blind, placebo-controlled, 2 × 2 factorial trial. Lancet 2018, 392, 2583–2594. [Google Scholar] [CrossRef]
- Watson, H.; Mitra, S.; Croden, F.C.; Taylor, M.; Wood, H.M.; Perry, S.L.; Spencer, J.A.; Quirke, P.; Toogood, G.J.; Lawton, C.L.; et al. A randomised trial of the effect of omega-3 polyunsaturated fatty acid supplements on the human intestinal microbiota. Gut 2018, 67, 1974–1983. [Google Scholar] [CrossRef]
- Ridlon, J.M.; Kang, D.J.; Hylemon, P.B. Bile salt biotransformations by human intestinal bacteria. J. Lipid Res. 2006, 47, 241–259. [Google Scholar] [CrossRef]
- McCoy, A.N.; Araujo-Perez, F.; Azcarate-Peril, A.; Yeh, J.J.; Sandler, R.S.; Keku, T.O. Fusobacterium is associated with colorectal adenomas. PLoS ONE 2013, 8, e53653. [Google Scholar]
- Sanapareddy, N.; Legge, R.M.; Jovov, B.; McCoy, A.; Burcal, L.; Araujo-Perez, F.; Randall, T.A.; Galanko, J.; Benson, A.; Sandler, R.S.; et al. Increased rectal microbial richness is associated with the presence of colorectal adenomas in humans. ISME J. 2012, 6, 1858–1868. [Google Scholar] [CrossRef] [Green Version]
- Dulal, S.; Keku, T.O. Gut microbiome and colorectal adenomas. Cancer J. 2014, 20, 225–231. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Chan, A.T.; Sun, J. Influence of the Gut Microbiome, Diet, and Environment on Risk of Colorectal Cancer. Gastroenterology 2020, 158, 322–340. [Google Scholar] [CrossRef] [PubMed]
- Morgan, X.C.; Huttenhower, C. Chapter 12: Human microbiome analysis. PLoS Comput. Biol. 2012, 8, e1002808. [Google Scholar] [CrossRef]
- Goedert, J.J.; Gong, Y.; Hua, X.; Zhong, H.; He, Y.; Peng, P.; Yu, G.; Wang, W.; Ravel, J.; Shi, J.; et al. Fecal Microbiota Characteristics of Patients with Colorectal Adenoma Detected by Screening: A Population-based Study. eBioMedicine 2015, 2, 597–603. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.M.; Yu, Y.N.; Wang, J.L.; Lin, Y.W.; Kong, X.; Yang, C.Q.; Yang, L.; Liu, Z.J.; Yuan, Y.Z.; Liu, F.; et al. Decreased dietary fiber intake and structural alteration of gut microbiota in patients with advanced colorectal adenoma. Am. J. Clin. Nutr. 2013, 97, 1044–1052. [Google Scholar] [CrossRef]
- Brim, H.; Yooseph, S.; Zoetendal, E.G.; Lee, E.; Torralbo, M.; Laiyemo, A.O.; Shokrani, B.; Nelson, K.; Ashktorab, H. Microbiome analysis of stool samples from African Americans with colon polyps. PLoS ONE 2013, 8, e81352. [Google Scholar] [CrossRef]
- Jonkers, I.J.; Smelt, A.H.; Princen, H.M.; Kuipers, F.; Romijn, J.A.; Boverhof, R.; Masclee, A.A.; Stellaard, F. Fish oil increases bile acid synthesis in male patients with hypertriglyceridemia. J. Nutr. 2006, 136, 987–991. [Google Scholar] [CrossRef]
- Ou, J.; Carbonero, F.; Zoetendal, E.G.; DeLany, J.P.; Wang, M.; Newton, K.; Gaskins, H.R.; O’Keefe, S.J. Diet, microbiota, and microbial metabolites in colon cancer risk in rural Africans and African Americans. Am. J. Clin. Nutr. 2013, 98, 111–120. [Google Scholar] [CrossRef]
- Hofmann, A.F.; Cravetto, C.; Molino, G.; Belforte, G.; Bona, B. Simulation of the metabolism and enterohepatic circulation of endogenous deoxycholic acid in humans using a physiologic pharmacokinetic model for bile acid metabolism. Gastroenterology 1987, 93, 693–709. [Google Scholar] [CrossRef]
- Louis, P.; Hold, G.L.; Flint, H.J. The gut microbiota, bacterial metabolites and colorectal cancer. Nat. Rev. Microbiol. 2014, 12, 661–672. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Keku, T.O.; Martin, C.; Galanko, J.; Woosley, J.T.; Schroeder, J.C.; Satia, J.A.; Halabi, S.; Sandler, R.S. Circulating levels of inflammatory cytokines and risk of colorectal adenomas. Cancer Res. 2008, 68, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Subar, A.F.; Thompson, F.E.; Kipnis, V.; Midthune, D.; Hurwitz, P.; McNutt, S.; McIntosh, A.; Rosenfeld, S. Comparative validation of the Block, Willett, and National Cancer Institute food frequency questionnaires: The Eating at America’s Table Study. Am. J. Epidemiol. 2001, 154, 1089–1099. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.J.; Rawls, J.F.; Randall, T.; Burcal, L.; Mpande, C.N.; Jenkins, N.; Jovov, B.; Abdo, Z.; Sandler, R.S.; Keku, T.O. Molecular characterization of mucosal adherent bacteria and associations with colorectal adenomas. Gut Microbes 2010, 1, 138–147. [Google Scholar] [CrossRef]
- Nugent, J.L.; McCoy, A.N.; Addamo, C.J.; Jia, W.; Sandler, R.S.; Keku, T.O. Altered tissue metabolites correlate with microbial dysbiosis in colorectal adenomas. J. Proteome Res. 2014, 13, 1921–1929. [Google Scholar] [CrossRef]
- Fadrosh, D.W.; Ma, B.; Gajer, P.; Sengamalay, N.; Ott, S.; Brotman, R.M.; Ravel, J. An improved dual-indexing approach for multiplexed 16S rRNA gene sequencing on the Illumina MiSeq platform. Microbiome 2014, 2, 6. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.G.; Rao, S.; Weir, T.L.; O’Malia, J.; Bazan, M.; Brown, R.J.; Ryan, E.P. Metabolomics and metabolic pathway networks from human colorectal cancers, adjacent mucosa, and stool. Cancer Metab. 2016, 4, 11. [Google Scholar] [CrossRef]
- Greenland, S.; Pearl, J.; Robins, J.M. Causal diagrams for epidemiologic research. Epidemiology 1999, 10, 37–48. [Google Scholar] [CrossRef]
- Dhariwal, A.; Chong, J.; Habib, S.; King, I.L.; Agellon, L.B.; Xia, J. MicrobiomeAnalyst: A web-based tool for comprehensive statistical, visual and meta-analysis of microbiome data. Nucleic Acids Res. 2017, 45, W180–W188. [Google Scholar] [CrossRef]
- Xia, J.; Wishart, D.S. Using MetaboAnalyst 3.0 for Comprehensive Metabolomics Data Analysis. Curr. Protoc. Bioinform. 2016, 55, 14.10.11–14.10.91. [Google Scholar] [CrossRef] [PubMed]
- Glickman, M.E.; Rao, S.R.; Schultz, M.R. False discovery rate control is a recommended alternative to Bonferroni-type adjustments in health studies. J. Clin. Epidemiol. 2014, 67, 850–857. [Google Scholar] [CrossRef]
- Hosmer, D.W.; Lemeshow, S. Applied Logistic Regression; John Wiley & Sons: New York, NY, USA, 2000. [Google Scholar]
- Murff, H.J.; Shrubsole, M.J.; Cai, Q.; Smalley, W.E.; Dai, Q.; Milne, G.L.; Ness, R.M.; Zheng, W. Dietary intake of PUFAs and colorectal polyp risk. Am. J. Clin. Nutr. 2012, 95, 703–712. [Google Scholar] [CrossRef] [PubMed]
- Pot, G.K.; Geelen, A.; van Heijningen, E.M.; Siezen, C.L.; van Kranen, H.J.; Kampman, E. Opposing associations of serum n-3 and n-6 polyunsaturated fatty acids with colorectal adenoma risk: An endoscopy-based case-control study. Int. J. Cancer 2008, 123, 1974–1977. [Google Scholar] [CrossRef]
- DeLuca, J.A.A.; Garcia-Villatoro, E.L.; Allred, C.D. Flaxseed Bioactive Compounds and Colorectal Cancer Prevention. Curr. Oncol. Rep. 2018, 20, 59. [Google Scholar] [CrossRef] [PubMed]
- Oikarinen, S.I.; Pajari, A.M.; Salminen, I.; Heinonen, S.M.; Adlercreutz, H.; Mutanen, M. Effects of a flaxseed mixture and plant oils rich in alpha-linolenic acid on the adenoma formation in multiple intestinal neoplasia (Min) mice. Br. J. Nutr. 2005, 94, 510–518. [Google Scholar] [CrossRef]
- Ivanov, I.I.; de Frutos, R.L.; Manel, N.; Yoshinaga, K.; Rifkin, D.B.; Sartor, R.B.; Finlay, B.B.; Littman, D.R. Specific microbiota direct the differentiation of IL-17-producing T-helper cells in the mucosa of the small intestine. Cell Host Microbe 2008, 4, 337–349. [Google Scholar] [CrossRef]
- Yang, T.; Santisteban, M.M.; Rodriguez, V.; Li, E.; Ahmari, N.; Carvajal, J.M.; Zadeh, M.; Gong, M.; Qi, Y.; Zubcevic, J.; et al. Gut dysbiosis is linked to hypertension. Hypertension 2015, 65, 1331–1340. [Google Scholar] [CrossRef]
- Kumar, M.; Pal, N.; Sharma, P.; Kumawat, M.; Sarma, D.K.; Nabi, B.; Verma, V.; Tiwari, R.R.; Shubham, S.; Arjmandi, B.; et al. Omega-3 Fatty Acids and Their Interaction with the Gut Microbiome in the Prevention and Amelioration of Type-2 Diabetes. Nutrients 2022, 14, 1723. [Google Scholar] [CrossRef]
- Menni, C.; Zierer, J.; Pallister, T.; Jackson, M.A.; Long, T.; Mohney, R.P.; Steves, C.J.; Spector, T.D.; Valdes, A.M. Omega-3 fatty acids correlate with gut microbiome diversity and production of N-carbamylglutamate in middle aged and elderly women. Sci. Rep. 2017, 7, 11079. [Google Scholar] [CrossRef]
- Raufman, J.P.; Dawson, P.A.; Rao, A.; Drachenberg, C.B.; Heath, J.; Shang, A.C.; Hu, S.; Zhan, M.; Polli, J.E.; Cheng, K. Slc10a2-null mice uncover colon cancer-promoting actions of endogenous fecal bile acids. Carcinogenesis 2015, 36, 1193–1200. [Google Scholar] [CrossRef] [Green Version]
- Dongfeng, D.; An, C.; Shujia, P.; Jikai, Y.; Tao, Y.; Rui, D.; Kai, T.; Yafeng, C.; Jianguo, L.; Xilin, D. Explanation of colon cancer pathophysiology through analyzing the disrupted homeostasis of bile acids. Afr. Health Sci. 2014, 14, 925–928. [Google Scholar] [PubMed]
- Centuori, S.M.; Martinez, J.D. Differential regulation of EGFR-MAPK signaling by deoxycholic acid (DCA) and ursodeoxycholic acid (UDCA) in colon cancer. Dig. Dis. Sci. 2014, 59, 2367–2380. [Google Scholar] [CrossRef]
- Anitha, M.; Reichardt, F.; Tabatabavakili, S.; Nezami, B.G.; Chassaing, B.; Mwangi, S.; Vijay-Kumar, M.; Gewirtz, A.; Srinivasan, S. Intestinal dysbiosis contributes to the delayed gastrointestinal transit in high-fat diet fed mice. Cell. Mol. Gastroenterol. Hepatol. 2016, 2, 328–339. [Google Scholar] [CrossRef] [PubMed]
- Ajouz, H.; Mukherji, D.; Shamseddine, A. Secondary bile acids: An underrecognized cause of colon cancer. World J. Surg. Oncol. 2014, 12, 164. [Google Scholar] [CrossRef] [PubMed]
- De Kok, T.M.; van Faassen, A.; Glinghammar, B.; Pachen, D.M.; Eng, M.; Rafter, J.J.; Baeten, C.G.; Engels, L.G.; Kleinjans, J.C. Bile acid concentrations, cytotoxicity, and pH of fecal water from patients with colorectal adenomas. Dig. Dis. Sci. 1999, 44, 2218–2225. [Google Scholar] [CrossRef]
- Bernstein, H.; Bernstein, C.; Payne, C.M.; Dvorakova, K.; Garewal, H. Bile acids as carcinogens in human gastrointestinal cancers. Mutat. Res. 2005, 589, 47–65. [Google Scholar] [CrossRef]
- Alberts, D.S.; Einspahr, J.G.; Earnest, D.L.; Krutzsch, M.F.; Lin, P.; Hess, L.M.; Heddens, D.K.; Roe, D.J.; Martinez, M.E.; Salen, G.; et al. Fecal bile acid concentrations in a subpopulation of the wheat bran fiber colon polyp trial. Cancer Epidemiol. Biomark. Prev. 2003, 12, 197–200. [Google Scholar]
- Hagio, M.; Matsumoto, M.; Fukushima, M.; Hara, H.; Ishizuka, S. Improved analysis of bile acids in tissues and intestinal contents of rats using LC/ESI-MS. J. Lipid Res. 2009, 50, 173–180. [Google Scholar] [CrossRef]
- Hosoyamada, Y.; Yamada, M. Effects of Dietary Fish Oil and Apple Polyphenol on the Concentration Serum Lipids and Excretion of Fecal Bile Acids in Rats. J. Nutr. Sci. Vitaminol. 2017, 63, 21–27. [Google Scholar] [CrossRef]
- Hale, V.L.; Chen, J.; Johnson, S.; Harrington, S.C.; Yab, T.C.; Smyrk, T.C.; Nelson, H.; Boardman, L.A.; Druliner, B.R.; Levin, T.R.; et al. Shifts in the Fecal Microbiota Associated with Adenomatous Polyps. Cancer Epidemiol. Biomark. Prev. 2017, 26, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Echarte, M.; Zulet, M.A.; Astiasaran, I. Oxidation process affecting fatty acids and cholesterol in fried and roasted salmon. J. Agric. Food Chem. 2001, 49, 5662–5667. [Google Scholar] [CrossRef] [PubMed]
- Terry, M.B.; Neugut, A.I.; Bostick, R.M.; Sandler, R.S.; Haile, R.W.; Jacobson, J.S.; Fenoglio-Preiser, C.M.; Potter, J.D. Risk factors for advanced colorectal adenomas: A pooled analysis. Cancer Epidemiol. Biomark. Prev. 2002, 11, 622–629. [Google Scholar]


{kind=link}
| Selected Characteristics | Cases (n = 217) | Controls (n = 218) | p-Value |
|---|---|---|---|
| Age (years), mean (SD) | 56.3 (6.9) | 55.2 (6.3) | 0.19 |
| Male (%) | 52.3 | 44.2 | 0.09 |
| White (%) | 81.0 | 85.3 | 0.31 |
| Family history of colorectal cancer in first-degree relative (%) | 3.4 | 4.6 | 0.54 |
| Regular (≥once/week) NSAID use (%) | 56.9 | 50.8 | 0.23 |
| Total energy intake (kcal/day), mean (SD) | 1992.4 (847.5) | 1880.8 (726.0) | 0.16 |
| Total ω-3 polyunsaturated fat intake (g/day), mean (SD) | 1.7 (1.0) | 1.6 (0.7) | 0.37 |
| Total ω-6 polyunsaturated fat intake (g/day), mean (SD) | 15.5 (9.3) | 14.8 (6.9) | 0.42 |
| Total saturated fat intake (g/day), mean (SD) | 24.1 (13.0) | 22.2 (10.1) | 0.10 |
| Total vegetables intake (servings/day), mean (SD) | 4.5 (2.6) | 4.4 (2.4) | 0.70 |
| Total fruit intake (servings/day), mean (SD) | 2.8 (1.9) | 2.8 (1.8) | 0.90 |
| Red meat (oz/day), mean (SD) | 1.5 (1.2) | 1.4 (1.2) | 0.30 |
| Dietary fiber intake (g/day), mean (SD) | 20.3 (9.8) | 20.0 (9.3) | 0.73 |
| Total calcium intake (mg/day), mean (SD) | 882.2 (421.0) | 836.3 (401.2) | 0.27 |
| Total folate intake (mcg/day), mean (SD) | 418.7 (247.0) | 425.5 (244.9) | 0.61 |
| Total vitamin E intake (mg/day), mean (SD) | 11.1 (7.4) | 10.5 (4.6) | 0.37 |
| Ever Smoked (%) | 42.5 | 45.7 | 0.52 |
| Alcohol intake, mean (SD) | 12.4 (17.1) | 11.0 (18.3) | 0.40 |
| Body mass index (kg/m2), mean (SD) | 28.1 (5.7) | 27.1 (5.7) | 0.07 |
| Waist-to-hip ratio, mean (SD) | 0.93 (0.1) | 0.91 (0.1) | 0.01 |
| Distal adenoma (%), mean (SD) | 65.2 | N.A. | N.A. |
| Adenoma size (cm), mean (SD) | 5.5 (5.0) | N.A. | N.A. |
| Bacteria diversity, mean (SD) | 8.6 (3.6) | 8.3 (3.8) | 0.42 |
| Bacteria evenness, mean (SD) | 0.7 (0.2) | 0.7 (0.3) | 0.24 |
| Bacteria richness, mean (SD) | 6.8 (2.6) | 6.6 (2.8) | 0.41 |
| Dietary Factors | Cases/Controls, n | Adjusted OR (95% CI) a |
|---|---|---|
| Total ω-3 PUFA intake, g/day | ||
| By tertiles | ||
| <1.25 | 63/64 | 1.00 (Ref) |
| 1.25– < 1.80 | 72/65 | 0.80 (0.45–1.42) |
| ≥1.80 | 68/65 | 0.49 (0.23–1.01) |
| By median | ||
| <1.47 | 94/96 | 1.00 (Ref) |
| ≥1.47 | 109/98 | 0.72 (0.41–1.24) |
| Ptrend | 0.06 | |
| Short-chain ω-3 PUFA intake, g/day | ||
| By tertiles | ||
| <1.16 | 66/64 | 1.00 (Ref) |
| 1.16– < 1.64 | 67/65 | 0.69 (0.39–1.22) |
| ≥1.64 | 70/65 | 0.45 (0.21–0.97) |
| By median | ||
| <1.37 | 100/95 | 1.00 (Ref) |
| ≥1.37 | 103/99 | 0.53 (0.30–0.92) |
| Ptrend | 0.04 | |
| Long-chain ω-3 PUFA intake, g/day | ||
| By tertiles | ||
| <0.07 | 56/52 | 1.00 (Ref) |
| 0.07– < 0.12 | 56/66 | 0.63 (0.35–1.10) |
| ≥0.12 | 91/76 | 0.89 (0.51–1.54) |
| By median | ||
| <0.09 | 81/91 | 1.00 (Ref) |
| ≥0.09 | 120/105 | 1.17 (0.74–1.87) |
| Ptrend | 0.22 |
| Characteristics | Controls/Cases, n | FC | FDR | Crude OR (95% CI) | Multivariable-Adjusted OR (95% CI) a |
|---|---|---|---|---|---|
| Bacteria taxa (Phylum; genus) | |||||
| Proteobacteria; Sphingomonas | 203/208 | 0.0642 | 2.12 × 10−13 | 2.18 (1.34–3.55) | 2.17 (1.31–3.57) |
| Proteobacteria; Marinomonas | 203/208 | 0.2053 | 3.24 × 10−6 | 1.18 (0.85–1.64) | 1.14 (0.81–1.60) |
| Proteobacteria; Sutterella | 203/208 | 1.9198 | 0.0326 | 0.78 (0.55–1.10) | 0.80 (0.56–1.14) |
| Bacteroidetes; Parabacteroides | 203/208 | 1.7654 | 0.0326 | 0.91 (0.65–1.28) | 0.94 (0.66–1.33) |
| Bacteroidetes; Bacteroides | 203/208 | 1.4240 | 0.0676 | 0.98 (0.84–1.14) | 1.02 (0.87–1.20) |
| Firmicutes; Streptococcus | 203/208 | 0.5705 | 0.0676 | 0.98 (0.79–1.22) | 0.94 (0.75–1.18) |
| Proteobacteria; Pseudoalteromonas | 203/208 | 0.5809 | 0.1577 | 0.58 (0.34–0.99) | 0.58 (0.34–1.00) |
| Firmicutes; Blautia | 203/208 | 0.7481 | 0.1874 | 1.14 (0.93–1.41) | 1.17 (0.94–1.47) |
| Firmicutes; Roseburia | 203/208 | 1.3751 | 0.2281 | 0.98 (0.75–1.27) | 0.96 (0.73–1.28) |
| Firmicutes; Phascolarctobacterium | 203/208 | 1.4854 | 0.2281 | 0.92 (0.67–1.25) | 1.02 (0.73–1.43) |
| Proteobacteria; Ralstonia | 203/208 | 0.5593 | 0.2281 | 1.20 (1.04–1.39) | 1.19 (1.02–1.38) |
| Proteobacteria; Bilophila | 203/208 | 1.2571 | 0.3822 | 1.20 (0.76–1.91) | 1.39 (0.84–2.30) |
| Actinobacteria; Collinsella | 203/208 | 1.1777 | 0.6344 | 1.02 (0.88–1.18) | 0.99 (0.85–1.16) |
| Actinobacteria; Propionibacterium | 203/208 | 1.1727 | 0.7359 | 1.04 (0.78–1.41) | 1.01 (0.73–1.38) |
| Actinobacteria; Bifidobacterium | 203/208 | 1.0527 | 0.9706 | 1.08 (0.78–1.51) | 0.99 (0.70–1.40) |
| Firmicutes; Coprococcus | 203/208 | 0.9833 | 0.9706 | 1.03 (0.84–1.26) | 1.06 (0.85–1.31) |
| Firmicutes; Ruminococcus | 203/208 | 0.9893 | 0.9706 | 1.12 (0.89–1.41) | 1.14 (0.89–1.44) |
| Firmicutes; Dorea | 203/208 | 0.9936 | 0.9706 | 1.08 (0.92–1.28) | 1.09 (0.91–1.30) |
| Bacteria Overall Composition Measurements | |||||
| Richness | 203/208 | -- | -- | 1.03 (0.96–1.11) | 1.04 (0.96–1.13) |
| Evenness | 203/208 | -- | -- | 1.61 (0.73–3.54) | 1.91 (0.83–4.39) |
| Diversity | 203/208 | -- | -- | 1.02 (0.97–1.08) | 1.04 (0.98–1.10) |
| Bacteria Characteristics | Short-Chain ω-3 PUFA Intake, g/Day | Cases (n) | Controls (n) | Single-Referenced ORs (95% CIs) | Stratified ORs (95% CIs) | p-interaction |
|---|---|---|---|---|---|---|
| Bacteria richness | ||||||
| <Median (7.38) | <Median (1.37) | 46 | 44 | 1.00 | 1.00 | |
| ≥Median (1.37) | 43 | 45 | 0.52 (0.26, 1.06) | 0.45 (0.20, 1.02) | ||
| ≥Median (7.38) | <Median (1.37) | 54 | 43 | 1.52 (0.69, 3.34) | 1.00 | |
| ≥Median (1.37) | 51 | 47 | 0.79 (0.33, 1.87) | 0.58 (0.26, 1.27) | 0.93 | |
| Bacteria evenness | ||||||
| <Median (0.80) | <Median (1.37) | 47 | 52 | 1.00 | 1.00 | |
| ≥Median (1.37) | 49 | 37 | 0.84 (0.40, 1.75) | 1.10 (0.50, 2.43) | ||
| ≥Median (0.80) | <Median (1.37) | 53 | 35 | 1.63 (0.77, 3.45) | 1.00 | |
| ≥Median (1.37) | 45 | 55 | 0.58 (0.25, 1.30) | 0.21 (0.09, 0.50) | 0.03 | |
| Bacteria diversity | ||||||
| <Median (9.33) | <Median (1.37) | 47 | 42 | 1.00 | 1.00 | |
| ≥Median (1.37) | 44 | 50 | 0.46 (0.23, 0.94) | 0.32 (0.14, 0.75) | ||
| ≥Median (9.33) | <Median (1.37) | 53 | 45 | 1.09 (0.49, 2.38) | 1.00 | |
| ≥Median (1.37) | 50 | 42 | 0.65 (0.27, 1.59) | 0.69 (0.32, 1.48) | 0.44 | |
| Sphingomonas | ||||||
| <Median (0.00) | <Median (1.37) | 65 | 49 | 1.00 | 1.00 | |
| ≥Median (1.37) | 50 | 57 | 0.40 (0.21, 0.78) | 0.40 (0.19, 0.85) | ||
| ≥ Median (0.00) | <Median (1.37) | 35 | 38 | 0.68 (0.36, 1.27) | 1.00 | |
| ≥Median (1.37) | 44 | 35 | 0.55 (0.27, 1.12) | 0.51 (0.18, 1.45) | 0.32 | |
| Marinomonas | ||||||
| <Median (0.01) | <Median (1.37) | 36 | 40 | 1.00 | 1.00 | |
| ≥Median (1.37) | 35 | 48 | 0.49 (0.23, 1.02) | 0.32 (0.13, 0.81) | ||
| ≥Median (0.01) | <Median (1.37) | 64 | 47 | 1.69 (0.87, 3.28) | 1.00 | |
| ≥Median (1.37) | 59 | 44 | 1.01 (0.47, 2.14) | 0.76 (0.35, 1.68) | 0.08 | |
| Sutterella | ||||||
| <Median (0.04) | <Median (1.37) | 53 | 43 | 1.00 | 1.00 | |
| ≥Median (1.37) | 47 | 48 | 0.43 (0.21, 0.89) | 0.55 (0.24, 1.26) | ||
| ≥Median (0.04) | <Median (1.37) | 47 | 44 | 0.81 (0.43, 1.55) | 1.00 | |
| ≥Median (1.37) | 47 | 44 | 0.51 (0.24, 1.08) | 0.43 (0.18, 1.01) | 0.53 | |
| Parabacteroides | ||||||
| <Median (0.30) | <Median (1.37) | 54 | 42 | 1.00 | 1.00 | |
| ≥Median (1.37) | 49 | 48 | 0.49 (0.25, 0.96) | 0.54 (0.24, 1.20) | ||
| ≥Median (0.30) | <Median (1.37) | 46 | 45 | 0.82 (0.43, 1.55) | 1.00 | |
| ≥Median (1.37) | 45 | 44 | 0.40 (0.22, 1.00) | 0.48 (0.20, 1.15) | 0.49 | |
| Pseudoalteromonas | ||||||
| <Median (0.04) | <Median (1.37) | 47 | 50 | 1.00 | 1.00 | |
| ≥Median (1.37) | 49 | 41 | 0.69 (0.34, 1.41) | 0.74 (0.33, 1.65) | ||
| ≥Median (0.04) | <Median (1.37) | 53 | 37 | 1.34 (0.72, 2.50) | 1.00 | |
| ≥Median (1.37) | 45 | 51 | 0.54 (0.27, 1.10) | 0.38 (0.17, 0.85) | 0.44 | |
| Ralstonia | ||||||
| <Median (0.00) | <Median (1.37) | 45 | 43 | 1.00 | 1.00 | |
| ≥Median (1.37) | 34 | 44 | 0.49 (0.23, 1.03) | 0.66 (0.28, 1.56) | ||
| ≥ Median (0.00) | <Median (1.37) | 55 | 44 | 1.17 (0.62, 2.19) | 1.00 | |
| ≥Median (1.37) | 60 | 48 | 0.64 (0.31, 1.30) | 0.38 (0.16, 0.87) | 0.49 |
| Microbial Metabolites/Bile Acids | Cases/Controls, n | FC | FDR | OR (95% CI) a |
|---|---|---|---|---|
| Cholate | 23/11 | 3.5372 | 0.0005 | 1.06 (0.51–2.18) |
| Taurolithocholate 3-sulfate | 15/3 | 5.0002 | 0.0005 | N.A. |
| Taurocholate | 20/12 | 6.5657 | 0.0007 | 2.80 (1.13–6.93) |
| Taurochenodeoxycholate | 18/13 | 6.3328 | 0.0028 | 4.45 (1.47–13.49) |
| Taurodeoxycholate | 17/11 | 8.3950 | 0.0038 | 3.68 (1.25–10.82) |
| Glycocholenate sulfate | 10/1 | 3.3605 | 0.0042 | N.A. |
| Tauroursodeoxycholate | 11/2 | 3.5215 | 0.0072 | N.A. |
| Chenodeoxycholate | 13/9 | 19.883 | 0.0113 | 5.12 (1.17–22.44) |
| Isobar: 7-ketodeoxycholate; 12-dehydrocholate | 13/5 | 2.4302 | 0.0144 | 1.34 (0.49–3.68) |
| Glycochenodeoxycholate | 12/8 | 11.6478 | 0.0312 | 3.11 (0.89–10.90) |
| Deoxycholate | 22/19 | 3.5998 | 0.0394 | 1.86 (0.93–3.73) |
| Glycocholate | 15/11 | 6.7675 | 0.0886 | 1.86 (0.76–3.54) |
| Glycoursodeoxycholate | 7/3 | 3.7757 | 0.0941 | N.A. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, T.; Brown, N.M.; McCoy, A.N.; Sandler, R.S.; Keku, T.O. Omega-3 Polyunsaturated Fatty Acids, Gut Microbiota, Microbial Metabolites, and Risk of Colorectal Adenomas. Cancers 2022, 14, 4443. https://doi.org/10.3390/cancers14184443
Wang T, Brown NM, McCoy AN, Sandler RS, Keku TO. Omega-3 Polyunsaturated Fatty Acids, Gut Microbiota, Microbial Metabolites, and Risk of Colorectal Adenomas. Cancers. 2022; 14(18):4443. https://doi.org/10.3390/cancers14184443
Chicago/Turabian StyleWang, Tengteng, Nicole M. Brown, Amber N. McCoy, Robert S. Sandler, and Temitope O. Keku. 2022. "Omega-3 Polyunsaturated Fatty Acids, Gut Microbiota, Microbial Metabolites, and Risk of Colorectal Adenomas" Cancers 14, no. 18: 4443. https://doi.org/10.3390/cancers14184443