
Involvement of Long Non-Coding RNAs in Glucose Metabolism in Cancer


Abstract
:Simple Summary
Abstract
1. Introduction
2. Methods
3. Results
3.1. LncRNAs Regulates Pathophysiological Processes in Cancer Via the Influence of Glucose Metabolism
3.2. LncRNAs and Glucose Metabolism in Different Cancer Entities
3.2.1. Gastrointestinal Cancers
Colorectal Cancer
Hepatocellular Carcinoma
Gastric Cancer
Pancreatic Cancer
Esophageal Cancer
Gallbladder Cancer
3.2.2. Genitourinary Cancers
Kidney Cancer
Bladder Cancer
Prostate Cancer
3.2.3. Female Reproductive System Cancers
Ovarian Cancer
Cervical Cancer
3.2.4. Breast Cancer
3.2.5. Hematologic Cancer
3.2.6. Brain Cancers
3.2.7. Osteosarcoma
3.2.8. Melanoma
3.2.9. Lung Cancer
3.2.10. Head and Neck Cancer
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Martinez-Outschoorn, U.E.; Peiris-Pagés, M.; Pestell, R.G.; Sotgia, F.; Lisanti, M.P. Cancer metabolism: A therapeutic perspective. Nat. Rev. Clin. Oncol. 2017, 14, 11–31. [Google Scholar] [CrossRef]
- DeBerardinis, R.J.; Chandel, N.S. Fundamentals of cancer metabolism. Sci. Adv. 2016, 2, e1600200. [Google Scholar] [CrossRef] [Green Version]
- Fadaka, A.; Ajiboye, B.; Ojo, O.; Adewale, O.; Olayide, I.; Emuowhochere, R. Biology of glucose metabolization in cancer cells. J. Oncol. Sci. 2017, 3, 45–51. [Google Scholar] [CrossRef]
- Warburg, O. On the Origin of Cancer Cells. Science 1956, 123, 309–314. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The Next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Jiang, B. Aerobic glycolysis and high level of lactate in cancer metabolism and microenvironment. Genes Dis. 2017, 4, 25–27. [Google Scholar] [CrossRef]
- Peppicelli, S.; Bianchini, F.; Calorini, L. Extracellular acidity, a “reappreciated” trait of tumor environment driving malignancy: Perspectives in diagnosis and therapy. Cancer Metastasis Rev. 2014, 33, 823–832. [Google Scholar] [CrossRef]
- Heiden, M.G.V.; Cantley, L.C.; Thompson, C.B. Understanding the Warburg Effect: The Metabolic Requirements of Cell Proliferation. Science 2009, 324, 1029–1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ling, H.; Vincent, K.; Pichler, M.; Fodde, R.; Berindan-Neagoe, I.; Slack, F.; Calin, G. Junk DNA and the long non-coding RNA twist in cancer genetics. Oncogene 2015, 34, 5003–5011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hombach, S.; Kretz, M. Non-coding RNAs: Classification, Biology and Functioning. Adv. Exp. Med. Biol. 2016, 937, 3–17. [Google Scholar] [CrossRef] [PubMed]
- Melé, M.; Mattioli, K.; Mallard, W.; Shechner, D.M.; Gerhardinger, C.; Rinn, J.L. Chromatin environment, transcriptional regulation, and splicing distinguish lincRNAs and mRNAs. Genome Res. 2017, 27, 27–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deveson, I.W.; Hardwick, S.A.; Mercer, T.R.; Mattick, J.S. The Dimensions, Dynamics, and Relevance of the Mammalian Noncoding Transcriptome. Trends Genet. 2017, 33, 464–478. [Google Scholar] [CrossRef]
- Pichler, M.; Rodriguez-Aguayo, C.; Nam, S.Y.; Dragomir, M.P.; Bayraktar, R.; Anfossi, S.; Knutsen, E.; Ivan, C.; Fuentes-Mattei, E.; Kil Lee, S.; et al. Therapeutic potential of FLANC, a novel primate-specific long non-coding RNA in colorectal cancer. Gut 2020, 69, 1818–1831. [Google Scholar] [CrossRef] [PubMed]
- Mercer, T.R.; Dinger, M.E.; Mattick, J.S. Long non-coding RNAs: Insights into functions. Nat. Rev. Genet. 2009, 10, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, S.U.; Grote, P.; Herrmann, B.G. Mechanisms of long noncoding RNA function in development and disease. Cell. Mol. Life Sci. 2016, 73, 2491–2509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazimierczyk, M.; Kasprowicz, M.K.; Kasprzyk, M.E.; Wrzesinski, J. Human Long Noncoding RNA Interactome: Detection, Characterization and Function. Int. J. Mol. Sci. 2020, 21, 1027. [Google Scholar] [CrossRef] [Green Version]
- Laurent, G.S.; Wahlestedt, C.; Kapranov, P. The Landscape of long noncoding RNA classification. Trends Genet. 2015, 31, 239–251. [Google Scholar] [CrossRef] [Green Version]
- Carlevaro-Fita, J.; Johnson, R. Global Positioning System: Understanding Long Noncoding RNAs through Subcellular Localization. Mol. Cell 2019, 73, 869–883. [Google Scholar] [CrossRef] [Green Version]
- Fu, P.-F.; Zheng, X.; Fan, X.; Lin, A.-F. Role of cytoplasmic lncRNAs in regulating cancer signaling pathways. J. Zhejiang Univ. Sci. B 2019, 20, 1–8. [Google Scholar] [CrossRef]
- Guzel, E.; Okyay, T.M.; Yalcinkaya, B.; Karacaoglu, S.; Gocmen, M.; Akcakuyu, M.H. Tumor suppressor and oncogenic role of long non-coding RNAs in cancer. North. Clin. Istanb. 2020, 7, 81. [Google Scholar] [CrossRef]
- Barth, D.A.; Juracek, J.; Slaby, O.; Pichler, M.; Calin, G.A. lncRNA and Mechanisms of Drug Resistance in Cancers of the Genitourinary System. Cancers 2020, 12, 2148. [Google Scholar] [CrossRef]
- Gupta, S.C.; Tripathi, Y.N. Potential of long non-coding RNAs in cancer patients: From biomarkers to therapeutic targets. Int. J. Cancer 2017, 140, 1955–1967. [Google Scholar] [CrossRef] [PubMed]
- Barth, D.A.; Prinz, F.; Teppan, J.; Jonas, K.; Klec, C.; Pichler, M. Long-noncoding RNA (IncRNA) in the regulation of hypoxia-inducible factor (HIF) in cancer. Noncoding RNA 2020, 6, 27. [Google Scholar] [CrossRef]
- Eales, K.L.; Hollinshead, K.E.R.; Tennant, D.A. Hypoxia and metabolic adaptation of cancer cells. Oncogenesis 2016, 5, e190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hua, Q.; Mi, B.; Xu, F.; Wen, J.; Zhao, L.; Liu, J.; Huang, G. Hypoxia-induced lncRNA-AC020978 promotes proliferation and glycolytic metabolism of non-small cell lung cancer by regulating PKM2/HIF-1α axis. Theranostics 2020, 10, 4762–4778. [Google Scholar] [CrossRef]
- Shi, J.; Wang, H.; Feng, W.; Huang, S.; An, J.; Qiu, Y.; Wu, K. Long non-coding RNA HOTTIP promotes hypoxia-induced glycolysis through targeting miR-615-3p/HMGB3 axis in non-small cell lung cancer cells. Eur. J. Pharmacol. 2019, 862, 172615. [Google Scholar] [CrossRef]
- Liu, D.; Li, H. Long non-coding RNA GEHT1 promoted the proliferation of ovarian cancer cells via modulating the protein stability of HIF1α. Biosci. Rep. 2019, 39, 20181650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Huang, Y.; Hu, K.; Zhang, Z.; Yang, J.; Wang, Z. HIF1A activates the transcription of lncRNA RAET1K to modulate hypoxia-induced glycolysis in hepatocellular carcinoma cells via miR-100-5p. Cell Death Dis. 2020, 11, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, M.-L.; Wang, X.-Y.; Chen, W.-M. TGF-β1 upregulates the expression of lncRNA UCA1 and its downstream HXK2 to promote the growth of hepatocellular carcinoma. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 4846–4854. [Google Scholar]
- Hong, J.; Guo, F.; Lu, S.-Y.; Shen, C.; Ma, D.; Zhang, X.; Xie, Y.; Yan, T.; Yu, T.; Sun, T.; et al. F. nucleatum targets lncRNA ENO1-IT1 to promote glycolysis and oncogenesis in colorectal cancer. Gut 2020. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, H.; Chen, X. Drug resistance and combating drug resistance in cancer. Cancer Drug Resist. 2019, 2, 141–160. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Liu, Y.; Xu, X. Knockdown of LncRNA-UCA1 suppresses chemoresistance of pediatric AML by inhibiting glycolysis through the microRNA-125a/hexokinase 2 pathway. J. Cell. Biochem. 2018, 119, 6296–6308. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Hu, N.; Wang, C.; Zhao, H. HOTAIRM1 knockdown enhances cytarabine-induced cytotoxicity by suppression of glycolysis through the Wnt/β-catenin/PFKP pathway in acute myeloid leukemia cells. Arch. Biochem. Biophys. 2020, 680, 108244. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Zhou, Y.; Chen, W.; Chen, L.; Lu, J.; He, F.; Li, X.; Zhao, L. Ginsenoside 20(S)-Rg3 Prevents PKM2-Targeting miR-324-5p from H19 Sponging to Antagonize the Warburg Effect in Ovarian Cancer Cells. Cell. Physiol. Biochem. 2018, 51, 1340–1353. [Google Scholar] [CrossRef] [PubMed]
- Jing, C.; Ma, R.; Cao, H.; Wang, Z.; Liu, S.; Chen, D.; Wu, Y.; Zhang, J.; Wu, J. Long noncoding RNA and mRNA profiling in cetuximab-resistant colorectal cancer cells by RNA sequencing analysis. Cancer Med. 2019, 8, 1641–1651. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Smolle, M.; Uranitsch, S.; Gerger, A.; Pichler, M.; Haybaeck, J. Current Status of Long Non-Coding RNAs in Human Cancer with Specific Focus on Colorectal Cancer. Int. J. Mol. Sci. 2014, 15, 13993–14013. [Google Scholar] [CrossRef] [Green Version]
- Deng, H.; Wang, J.M.; Li, M.; Tang, R.; Tang, K.; Su, Y.; Hou, Y.; Zhang, J. Long non-coding RNAs: New biomarkers for prognosis and diagnosis of colon cancer. Tumor Biol. 2017, 39, 101042831770633. [Google Scholar] [CrossRef] [Green Version]
- Rigoutsos, I.; Kil Lee, S.; Nam, S.Y.; Anfossi, S.; Pasculli, B.; Pichler, M.; Jing, Y.; Rodriguez-Aguayo, C.; Telonis, A.G.; Rossi, S.; et al. N-BLR, a primate-specific non-coding transcript leads to colorectal cancer invasion and migration. Genome Biol. 2017, 18, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Sawayama, H.; Ishimoto, T.; Sugihara, H.; Miyanari, N.; Miyamoto, Y.; Baba, Y.; Yoshida, N.; Baba, H. Clinical impact of the Warburg effect in gastrointestinal cancer (Review). Int. J. Oncol. 2014, 45, 1345–1354. [Google Scholar] [CrossRef]
- Dang, C.V.; Le, A.; Gao, P. MYC-Induced Cancer Cell Energy Metabolism and Therapeutic Opportunities. Clin. Cancer Res. 2009, 15, 6479–6483. [Google Scholar] [CrossRef] [Green Version]
- Tang, J.; Yan, T.; Bao, Y.; Shen, C.; Yu, C.; Zhu, X.; Tian, X.; Guo, F.; Liang, Q.; Liu, Q.; et al. LncRNA GLCC1 promotes colorectal carcinogenesis and glucose metabolism by stabilizing c-Myc. Nat. Commun. 2019, 10, 1–15. [Google Scholar] [CrossRef]
- Feng, J.; Ma, J.; Liu, S.; Wang, J.; Chen, Y. A noncoding RNA LINC00504 interacts with c-Myc to regulate tumor metabolism in colon cancer. J. Cell. Biochem. 2019, 120, 14725–14734. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Lu, J.-H.; Wu, Q.-N.; Jin, Y.; Wang, D.-S.; Chen, Y.-X.; Liu, J.; Luo, X.-J.; Meng, Q.; Pu, H.-Y.; et al. LncRNA LINRIS stabilizes IGF2BP2 and promotes the aerobic glycolysis in colorectal cancer. Mol. Cancer 2019, 18, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Wang, P.; Du, J.; Chen, J.; Liu, W.; Ye, K. LncRNA RAD51-AS1 / miR -29b/c-3p/ NDRG2 crosstalk repressed proliferation, invasion and glycolysis of colorectal cancer. IUBMB Life 2021, 73, 286–298. [Google Scholar] [CrossRef] [PubMed]
- Qu, L.; Wu, Z.; Li, Y.; Xu, Z.; Liu, B.; Liu, F.; Bao, Y.; Wu, D.; Liu, J.; Wang, A.; et al. A feed-forward loop between lncARSR and YAP activity promotes expansion of renal tumour-initiating cells. Nat. Commun. 2016, 7, 12692. [Google Scholar] [CrossRef]
- Qu, L.; Ding, J.; Chen, C.; Wu, Z.; Liu, B.; Gao, Y.; Chen, W.; Liu, F.; Sun, W.; Li, X.-F.; et al. Exosome-transmitted lncARSR promotes sunitinib resistance in renal cancer by acting as a competing endogenous RNA. Cancer Cell 2016, 29, 653–668. [Google Scholar] [CrossRef]
- Li, S.; Zhu, K.; Liu, L.; Gu, J.; Niu, H.; Guo, J. lncARSR sponges miR-34a-5p to promote colorectal cancer invasion and metastasis via hexokinase-1-mediated glycolysis. Cancer Sci. 2020, 111, 3938–3952. [Google Scholar] [CrossRef]
- Bian, Z.; Zhang, J.; Li, M.; Feng, Y.; Wang, X.; Zhang, J.; Yao, S.; Jin, G.; Du, J.; Han, W.; et al. LncRNA–FEZF1-AS1 Promotes Tumor Proliferation and Metastasis in Colorectal Cancer by Regulating PKM2 Signaling. Clin. Cancer Res. 2018, 24, 4808–4819. [Google Scholar] [CrossRef] [Green Version]
- Ellis, B.C.; Graham, L.D.; Molloy, P.L. CRNDE, a long non-coding RNA responsive to insulin/IGF signaling, regulates genes involved in central metabolism. Biochim. Biophys. Acta (BBA) Bioenerg. 2014, 1843, 372–386. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.-L.; Li, L.; Jia, Y.-X.; Zhang, B.-Z.; Li, J.-C.; Zhu, Y.-H.; Li, M.-Q.; He, J.-Z.; Zeng, T.-T.; Ban, X.-J.; et al. LINC01554-Mediated Glucose Metabolism Reprogramming Suppresses Tumorigenicity in Hepatocellular Carcinoma via Downregulating PKM2 Expression and Inhibiting Akt/mTOR Signaling Pathway. Theranostics 2019, 9, 796–810. [Google Scholar] [CrossRef]
- Malakar, P.; Stein, I.; Saragovi, A.; Winkler, R.; Stern-Ginossar, N.; Berger, M.; Pikarsky, E.; Karni, R. Long Noncoding RNA MALAT1 Regulates Cancer Glucose Metabolism by Enhancing mTOR-Mediated Translation of TCF7L2. Cancer Res. 2019, 79, 2480–2493. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Zhao, Q.; Qi, J.; Wang, W.; Zhang, D.; Li, Z.; Qin, C. lncRNA Ftx promotes aerobic glycolysis and tumor progression through the PPARγ pathway in hepatocellular carcinoma. Int. J. Oncol. 2018, 53, 551–566. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Cheng, T.; He, Y.; Zhou, S.; Wang, Y.; Zhang, K.; Yu, P. High glucose regulates ERp29 in hepatocellular carcinoma by LncRNA MEG3-miRNA 483-3p pathway. Life Sci. 2019, 232, 116602. [Google Scholar] [CrossRef]
- Wei, S.; Fan, Q.; Yang, L.; Zhang, X.; Ma, Y.; Zong, Z.; Hua, X.; Su, N.; Sun, H.; Li, H.; et al. Promotion of glycolysis by HOTAIR through GLUT1 upregulation via mTOR signaling. Oncol. Rep. 2017, 38, 1902–1908. [Google Scholar] [CrossRef] [Green Version]
- Hu, M.; Fu, Q.; Jing, C.; Zhang, X.; Qin, T.; Pan, Y. LncRNA HOTAIR knockdown inhibits glycolysis by regulating miR-130a-3p/HIF1A in hepatocellular carcinoma under hypoxia. Biomed. Pharmacother. 2020, 125, 109703. [Google Scholar] [CrossRef] [PubMed]
- Wan, T.; Zheng, J.; Yao, R.; Yang, S.; Zheng, W.; Zhou, P. LncRNA DDX11-AS1 accelerates hepatocellular carcinoma progression via the miR-195-5p/MACC1 pathway. Ann. Hepatol. 2021, 20, 100258. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Zhang, D.; Zheng, T.; Yang, G.; Wang, J.; Meng, F.; Liu, Y.; Zhang, G.; Zhang, L.; Han, J.; et al. lncRNA-SOX2OT promotes hepatocellular carcinoma invasion and metastasis through miR-122-5p-mediated activation of PKM2. Oncogenesis 2020, 9, 54. [Google Scholar] [CrossRef]
- Qian, Y.; Song, W.; Wu, X.; Hou, G.; Wang, H.; Hang, X.; Xia, T. DLX6 Antisense RNA 1 Modulates Glucose Metabolism and Cell Growth in Gastric Cancer by Targeting microRNA-4290. Dig. Dis. Sci. 2021, 66, 460–473. [Google Scholar] [CrossRef]
- Ni, C.; Yang, P.; Guo, J.; Ye, M. Role of DiGeorge syndrome critical region gene 9, a long noncoding RNA, in gastric cancer. OncoTargets Ther. 2018, 11, 2259–2267. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Hu, M.; Zhou, L.; Ling, S.; Li, Y.; Kong, B.; Huang, P. Long non‑coding RNA HOTAIR promotes cancer cell energy metabolism in pancreatic adenocarcinoma by upregulating hexokinase‑2. Oncol. Lett. 2019, 18, 2212–2219. [Google Scholar] [CrossRef]
- Li, W.; Huang, K.; Wen, F.; Cui, G.; Guo, H.; He, Z.; Zhao, S. LINC00184 silencing inhibits glycolysis and restores mitochondrial oxidative phosphorylation in esophageal cancer through demethylation of PTEN. EBioMedicine 2019, 44, 298–310. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.-E.; Shi, H.-H.; Luo, X.-J. Upregulated Long Noncoding RNA UCA1 Enhances Warburg Effect via miR-203/HK2 Axis in Esophagal Cancer. J. Oncol. 2020, 2020, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Yu, Y.; Li, H.; Hu, Q.; Chen, X.; He, Y.; Xue, C.; Ren, F.; Ren, Z.; Li, J.; et al. Long non-coding RNA PVT1 promotes tumor progression by regulating the miR-143/HK2 axis in gallbladder cancer. Mol. Cancer 2019, 18, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.; Sun, Z.; Zhang, S.; Chen, Y.; Zhou, B.; Li, G.; Sun, Q.; Zhou, D.; Ge, Y.; Yan, S.; et al. Down-regulation of Long Non-coding RNA LINC01554 in Hepatocellular Cancer and its Clinical Significance. J. Cancer 2020, 11, 3369–3374. [Google Scholar] [CrossRef]
- Guo, J.; Chen, Z.; Jiang, H.; Yu, Z.; Peng, J.; Xie, J.; Li, Z.; Wu, W.; Cheng, Z.; Xiao, K. The lncRNA DLX6-AS1 promoted cell proliferation, invasion, migration and epithelial-to-mesenchymal transition in bladder cancer via modulating Wnt/β-catenin signaling pathway. Cancer Cell Int. 2019, 19, 312. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Li, P.; Zhao, W.; Yang, R.; Chen, S.; Bai, Y.; Dun, S.; Chen, X.; Du, Y.; Wang, Y.; et al. Expression of long non-coding RNA DLX6-AS1 in lung adenocarcinoma. Cancer Cell Int. 2015, 15, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Zhang, H.; Wu, X. Long noncoding RNA DLX6-AS1 accelerates the glioma carcinogenesis by competing endogenous sponging miR-197-5p to relieve E2F1. Gene 2019, 686, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Wang, Y.-R.; Jia, S.-H. Knockdown of long noncoding RNA DLX6-AS1 inhibits cell proliferation and invasion of cervical cancer cells by downregulating FUS. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 7307–7313. [Google Scholar]
- Yang, J.; Ye, Z.; Mei, D.; Gu, H.; Zhang, J. Long noncoding RNA DLX6-AS1 promotes tumorigenesis by modulating miR-497-5p/FZD4/ FZD6/Wnt/β-catenin pathway in pancreatic cancer. Cancer Manag. Res. 2019, 11, 4209–4221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, X.; Hu, Z.; Ke, X.; Tang, H.; Wu, B.; Wei, X.; Liu, Z. Long noncoding RNA DLX6-AS1 promotes renal cell carcinoma progression via miR-26a/PTEN axis. Cell Cycle 2017, 16, 2212–2219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; He, X.; Jin, T.; Gang, L.; Jin, Z. Long non-coding RNA DLX6-AS1 aggravates hepatocellular carcinoma carcinogenesis by modulating miR-203a/MMP-2 pathway. Biomed. Pharmacother. 2017, 96, 884–891. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.-J.; Xu, W.-R.; Chen, B.; Wang, Y.-Y.; Yang, N.; Wang, L.-J.; Zhang, Y.-L. The up-regulated lncRNA DLX6-AS1 in colorectal cancer promotes cell proliferation, invasion and migration via modulating PI3K/AKT/mTOR pathway. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 8321–8331. [Google Scholar] [PubMed]
- Liu, H.; Ye, D.; Chen, A.; Tan, D.; Zhang, W.; Jiang, W.; Wang, M.; Zhang, X. A pilot study of new promising non-coding RNA diagnostic biomarkers for early-stage colorectal cancers. Clin. Chem. Lab. Med. 2019, 57, 1073–1083. [Google Scholar] [CrossRef]
- Qiu, C.; Li, S.; Sun, D.; Yang, S. lncRNA PVT1 accelerates progression of non‑small cell lung cancer via targeting miRNA‑526b/EZH2 regulatory loop. Oncol. Lett. 2020, 19, 1267–1272. [Google Scholar] [CrossRef] [Green Version]
- Huang, F.; Chen, W.; Peng, J.; Li, Y.; Zhuang, Y.; Zhu, Z.; Shao, C.; Yang, W.; Yao, H.; Zhang, S. LncRNA PVT1 triggers Cyto-protective autophagy and promotes pancreatic ductal adenocarcinoma development via the miR-20a-5p/ULK1 Axis. Mol. Cancer 2018, 17, 1–16. [Google Scholar] [CrossRef]
- Bukowski, R.M.M. Genitourinary Oncology: Current Status and Future Challenges. Front. Oncol. 2011, 1, 32. [Google Scholar] [CrossRef] [Green Version]
- Miller, K.D.; Nogueira, L.; Mariotto, A.B.; Rowland, J.H.; Yabroff, K.R.; Alfano, C.M.; Jemal, A.; Kramer, J.L.; Siegel, R.L. Cancer treatment and survivorship statistics, 2019. CA Cancer J. Clin. 2019, 69, 363–385. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Z.-D.; Han, L.; Lee, H.; Zhuang, L.; Zhang, Y.; Baddour, J.; Nagrath, D.; Wood, C.G.; Gu, J.; Wu, X.; et al. Energy stress-induced lncRNA FILNC1 represses c-Myc-mediated energy metabolism and inhibits renal tumor development. Nat. Commun. 2017, 8, 1–13. [Google Scholar] [CrossRef]
- Hu, R.; Zhong, P.; Xiong, L.; Duan, L. Long Noncoding RNA Cancer Susceptibility Candidate 8 Suppresses the Proliferation of Bladder Cancer Cells via Regulating Glycolysis. DNA Cell Biol. 2017, 36, 767–774. [Google Scholar] [CrossRef]
- Jiang, S.; Zhang, L.-F.; Zhang, H.-W.; Hu, S.; Lu, M.-H.; Liang, S.; Li, B.; Li, Y.; Li, D.; Wang, E.-D.; et al. A novel miR-155/miR-143 cascade controls glycolysis by regulatinghexokinase 2in breast cancer cells. EMBO J. 2012, 31, 1985–1998. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Li, X.; Wu, S.; Xue, M.; Chen, W. Long non-coding RNA UCA1 promotes glycolysis by upregulating hexokinase 2 through the mTOR –STAT3/microRNA143 pathway. Cancer Sci. 2014, 105, 951–955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hung, C.-L.; Wang, L.-Y.; Yu, Y.-L.; Chen, H.-W.; Srivastava, S.; Petrovics, G.; Kung, H.-J. A long noncoding RNA connects c-Myc to tumor metabolism. Proc. Natl. Acad. Sci. USA 2014, 111, 18697–18702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rupaimoole, R.; Lee, J.; Haemmerle, M.; Ling, H.; Previs, R.A.; Pradeep, S.; Wu, S.Y.; Ivan, C.; Ferracin, M.; Dennison, J.B.; et al. Long Noncoding RNA Ceruloplasmin Promotes Cancer Growth by Altering Glycolysis. Cell Rep. 2015, 13, 2395–2402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, L.; Huang, C.; Li, J.; Gao, T.; Lin, Z.; Yao, T. Long non-coding RNA urothelial cancer associated 1 regulates radioresistance via the hexokinase 2/glycolytic pathway in cervical cancer. Int. J. Mol. Med. 2018, 42, 2247–2259. [Google Scholar] [CrossRef] [Green Version]
- Xing, Z.; Zhang, Y.; Liang, K.; Yan, L.; Xiang, Y.; Li, C.; Hu, Q.; Jin, F.; Putluri, V.; Putluri, N.; et al. Expression of Long Noncoding RNA YIYA Promotes Glycolysis in Breast Cancer. Cancer Res. 2018, 78, 4524–4532. [Google Scholar] [CrossRef] [Green Version]
- Yu, F.-X.; Zhao, B.; Guan, K.L. Hippo Pathway in Organ Size Control, Tissue Homeostasis, and Cancer. Cell 2015, 163, 811–828. [Google Scholar] [CrossRef] [Green Version]
- Zheng, X.; Han, H.; Liu, G.; Ma, Y.; Pan, R.; Sang, L.; Li, R.; Yang, L.; Marks, J.R.; Wang, W.; et al. Lnc RNA wires up Hippo and Hedgehog signaling to reprogramme glucose metabolism. EMBO J. 2017, 36, 3325–3335. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, H.; Wang, W.; Yu, X.; Xu, Q. LINC00346 regulates glycolysis by modulation of glucose transporter 1 in breast cancer cells. Mol. Cell. Probes 2020, 54, 101667. [Google Scholar] [CrossRef]
- Sun, L.-Y.; Li, X.-J.; Sun, Y.-M.; Huang, W.; Fang, K.; Han, C.; Chen, Z.-H.; Luo, X.-Q.; Chen, Y.-Q.; Wang, W.-T. LncRNA ANRIL regulates AML development through modulating the glucose metabolism pathway of AdipoR1/AMPK/SIRT1. Mol. Cancer 2018, 17, 1–6. [Google Scholar] [CrossRef]
- Congrains, A.; Kamide, K.; Ohishi, M.; Rakugi, H. ANRIL: Molecular Mechanisms and Implications in Human Health. Int. J. Mol. Sci. 2013, 14, 1278–1292. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Dai, R.; Chen, Y.; Guo, H.; Han, Y.; Zhang, Y. LncRNA LINP1 regulates acute myeloid leukemia progression via HNF4α/AMPK/WNT5A signaling pathway. Hematol. Oncol. 2019, 37, 474–482. [Google Scholar] [CrossRef]
- Zhang, Y.; He, Q.; Hu, Z.; Feng, Y.; Fan, L.; Tang, Z.; Yuan, J.; Shan, W.; Li, C.; Hu, X.; et al. Long noncoding RNA LINP1 regulates repair of DNA double-strand breaks in triple-negative breast cancer. Nat. Struct. Mol. Biol. 2016, 23, 522–530. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liu, H.; Shi, L.; Yu, X.; Gu, Y.; Sun, X. LINP1 facilitates DNA damage repair through non-homologous end joining (NHEJ) pathway and subsequently decreases the sensitivity of cervical cancer cells to ionizing radiation. Cell Cycle 2018, 17, 439–447. [Google Scholar] [CrossRef]
- Díaz-Beyá, M.; Brunet, S.; Nomdedéu, J.; Pratcorona, M.; Cordeiro, A.; Gallardo, D.; Escoda, L.; Tormo, M.; Heras, I.; Ribera, J.M.; et al. The lincRNA HOTAIRM1, located in the HOXA genomic region, is expressed in acute myeloid leukemia, impacts prognosis in patients in the intermediate-risk cytogenetic category, and is associated with a distinctive microRNA signature. Oncotarget 2015, 6, 31613–31627. [Google Scholar] [CrossRef] [Green Version]
- Hu, N.; Chen, L.; Li, Q.; Zhao, H. LncRNA HOTAIRM1 is involved in the progression of acute myeloid leukemia through targeting miR-148b. RSC Adv. 2019, 9, 10352–10359. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Ye, H.; He, M.; Zhou, X.; Sun, N.; Guo, W.; Lin, X.; Huang, H.; Lin, Y.; Yao, R.; et al. LncRNA PDIA3P interacts with c-Myc to regulate cell proliferation via induction of pentose phosphate pathway in multiple myeloma. Biochem. Biophys. Res. Commun. 2018, 498, 207–213. [Google Scholar] [CrossRef]
- Liu, N.; Feng, S.; Li, H.; Chen, X.; Bai, S.; Liu, Y. Long non-coding RNA MALAT1 facilitates the tumorigenesis, invasion and glycolysis of multiple myeloma via miR-1271-5p/SOX13 axis. J. Cancer Res. Clin. Oncol. 2020, 146, 367–379. [Google Scholar] [CrossRef] [Green Version]
- He, Z.; Ruan, X.; Liu, X.; Zheng, J.; Liu, Y.; Liu, L.; Ma, J.; Shao, L.; Wang, D.; Shen, S.; et al. FUS/circ_002136/miR-138-5p/SOX13 feedback loop regulates angiogenesis in Glioma. J. Exp. Clin. Cancer Res. 2019, 38, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.; Luo, C.; Guo, Z. LncRNA-XIST/microRNA-126 sponge mediates cell proliferation and glucose metabolism through the IRS1/PI3K/Akt pathway in glioma. J. Cell. Biochem. 2019, 121, 2170–2183. [Google Scholar] [CrossRef]
- Xu, R.; Zhu, X.; Chen, F.; Huang, C.; Ai, K.; Wu, H.; Zhang, L.; Zhao, X. LncRNA XIST/miR-200c regulates the stemness properties and tumourigenicity of human bladder cancer stem cell-like cells. Cancer Cell Int. 2018, 18, 1–10. [Google Scholar] [CrossRef]
- Zhu, H.; Zheng, T.; Yu, J.; Zhou, L.; Wang, L. LncRNA XIST accelerates cervical cancer progression via upregulating Fus through competitively binding with miR-200a. Biomed. Pharmacother. 2018, 105, 789–797. [Google Scholar] [CrossRef]
- Wei, W.; Liu, Y.; Lu, Y.; Yang, B.; Tang, L. LncRNA XIST Promotes Pancreatic Cancer Proliferation Through miR-133a/EGFR. J. Cell. Biochem. 2017, 118, 3349–3358. [Google Scholar] [CrossRef]
- Shi, J.; Zhang, Y.; Qin, B.; Wang, Y.; Zhu, X. Long non-coding RNA LINC00174 promotes glycolysis and tumor progression by regulating miR-152-3p/SLC2A1 axis in glioma. J. Exp. Clin. Cancer Res. 2019, 38, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Qin, D.; Jiang, Z.; Zhang, J. SNHG9/miR-199a-5p/Wnt2 Axis Regulates Cell Growth and Aerobic Glycolysis in Glioblastoma. J. Neuropathol. Exp. Neurol. 2019, 78, 939–948. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, L.; Luo, Y.; Cui, S.; Chen, W.; Zeng, A.; Shi, Y.; Luo, L. Long non-coding RNA SNHG5 promotes glioma progression via miR-205/E2F3 axis. Biosci. Rep. 2019, 39. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Shi, Y.; Shi, J.; Wang, H.; Sheng, Y.; Jiang, Q.; Chen, H.; Li, X.; Dong, J. The long non-coding RNA SNHG1 promotes glioma progression by competitively binding to miR-194 to regulate PHLDA1 expression. Cell Death Dis. 2019, 10, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Gao, Y.; Li, D.; Cao, Y.; Hao, B. LncRNA-TP53TG1 Participated in the Stress Response Under Glucose Deprivation in Glioma. J. Cell. Biochem. 2017, 118, 4897–4904. [Google Scholar] [CrossRef]
- Zheng, D.; Che, D.; Lin, F.; Wang, X.; Lu, L.; Chen, J.; Xu, X. LncRNA MACC1-AS1/MACC1 enhances the progression of glioma via regulating metabolic plasticity. Cell Cycle 2020, 19, 2286–2297. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Yang, Y.; Sun, Y.; Qin, L.; Yang, Y. LncRNA TUG1 affects cell viability by regulating glycolysis in osteosarcoma cells. Gene 2018, 674, 87–92. [Google Scholar] [CrossRef]
- Shen, Y.; Xu, J.; Pan, X.; Zhang, Y.; Weng, Y.; Zhou, D.; He, S. LncRNA KCNQ1OT1 sponges miR-34c-5p to promote osteosarcoma growth via ALDOA enhanced aerobic glycolysis. Cell Death Dis. 2020, 11, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Wu, X.; Liu, F.; Li, M.; Sun, Y.; Wang, Y.; Wang, C.; Zhu, K.; Jia, X.; Wang, B.; et al. Long non-coding RNA PVT1 promotes glycolysis and tumor progression by regulating miR-497/HK2 axis in osteosarcoma. Biochem. Biophys. Res. Commun. 2017, 490, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Xu, X.; Lu, S.; Hu, B. Long non‑coding RNA HAND2‑AS1 targets glucose metabolism and inhibits cancer cell proliferation in osteosarcoma. Oncol. Lett. 2019, 18, 1323–1329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, Y.; Zhu, X.; Xu, Y.; Tang, Q.; Huang, Z.; Zhao, Z.; Lu, J.; Song, G.; Xu, H.; Deng, C.; et al. Energy stress-induced lncRNA HAND2-AS1 represses HIF1α-mediated energy metabolism and inhibits osteosarcoma progression. Am. J. Cancer Res. 2018, 8, 526–537. [Google Scholar] [PubMed]
- Yang, X.; Wang, C.C.; Lee, W.Y.W.; Trovik, J.; Chung, T.K.H.; Kwong, J. Long non-coding RNA HAND2-AS1 inhibits invasion and metastasis in endometrioid endometrial carcinoma through inactivating neuromedin U. Cancer Lett. 2018, 413, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Ghafouri-Fard, S.; Esmaeili, M.; Taheri, M. H19 lncRNA: Roles in tumorigenesis. Biomed. Pharmacother. 2020, 123, 109774. [Google Scholar] [CrossRef]
- Wang, J.; Zhao, L.; Shang, K.; Liu, F.; Che, J.; Li, H.; Cao, B. Long non-coding RNA H19, a novel therapeutic target for pancreatic cancer. Mol. Med. 2020, 26, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Luan, W.; Zhou, Z.; Ni, X.; Xia, Y.; Wang, J.; Yan, Y.; Xu, B. Long non-coding RNA H19 promotes glucose metabolism and cell growth in malignant melanoma via miR-106a-5p/E2F3 axis. J. Cancer Res. Clin. Oncol. 2018, 144, 531–542. [Google Scholar] [CrossRef]
- Gong, Y.; Cao, Y.; Song, L.; Zhou, J.; Wang, C.; Wu, B. HMGB3 characterization in gastric cancer. Genet. Mol. Res. 2013, 12, 6032–6039. [Google Scholar] [CrossRef]
- Elgamal, O.A.; Park, J.-K.; Gusev, Y.; Azevedo-Pouly, A.C.P.; Jiang, J.; Roopra, A.; Schmittgen, T.D. Tumor Suppressive Function of mir-205 in Breast Cancer Is Linked to HMGB3 Regulation. PLoS ONE 2013, 8, e76402. [Google Scholar] [CrossRef]
- Zou, Z.; Gao, J.; Zhang, H.; Lin, Z.; Zhang, Y.; Luo, X.; Liu, C.; Xie, J.; Cai, C. Increased expression of HMGB3: A novel independent prognostic marker of worse outcome in patients with esophageal squamous cell carcinoma. Int. J. Clin. Exp. Pathol. 2015, 8, 345–352. [Google Scholar]
- Wang, S.; Wang, T.; Liu, D.; Kong, H. LncRNA MALAT1 Aggravates the Progression of Non-Small Cell Lung Cancer by Stimulating the Expression of COMMD8 via Targeting miR-613. Cancer Manag. Res. 2020, 12, 10735–10747. [Google Scholar] [CrossRef]
- Zhang, R.; Niu, Z.; Pei, H.; Peng, Z. Long noncoding RNA LINC00657 induced by SP1 contributes to the non-small cell lung cancer progression through targeting miR-26b-5p/COMMD8 axis. J. Cell. Physiol. 2020, 235, 3340–3349. [Google Scholar] [CrossRef] [PubMed]
- Lang, N.; Wang, C.; Zhao, J.; Shi, F.; Wu, T.; Cao, H. Long non‑coding RNA BCYRN1 promotes glycolysis and tumor progression by regulating the miR‑149/PKM2 axis in non‑small‑cell lung cancer. Mol. Med. Rep. 2020, 21, 1509–1516. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Wang, H.; Wu, B.; Zhang, C.; Yu, H.; Li, X.; Wang, Q.; Shi, X.; Fan, C.; Wang, D.; et al. Long Noncoding RNA LINC00551 Suppresses Glycolysis and Tumor Progression by Regulating c-Myc-Mediated PKM2 Expression in Lung Adenocarcinoma. OncoTargets Ther. 2020, 13, 11459–11470. [Google Scholar] [CrossRef]
- Zou, Z.W.; Ma, C.; Medoro, L.; Chen, L.; Wang, B.; Gupta, R.; Liu, T.; Yang, X.Z.; Chen, T.T.; Wang, R.Z.; et al. LncRNA ANRIL is up-regulated in nasopharyngeal carcinoma and promotes the cancer progression via increasing proliferation, reprograming cell glucose metabolism and inducing side-population stem-like cancer cells. Oncotarget 2016, 7, 61741–61754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Zhang, X.; Wang, Z.; Hu, Q.; Wu, J.; Li, Y.; Ren, X.; Wu, T.; Tao, X.; Chen, X.; et al. LncRNA-p23154 promotes the invasion-metastasis potential of oral squamous cell carcinoma by regulating Glut1-mediated glycolysis. Cancer Lett. 2018, 434, 172–183. [Google Scholar] [CrossRef]
- Shi, T.T.; Li, R.; Zhao, L. Long noncoding RNA UCA1 regulates CCR7 expression to promote tongue squamous cell carcinoma progression by sponging miR-138-5p. Neoplasma 2021, 67, 1256–1265. [Google Scholar] [CrossRef]



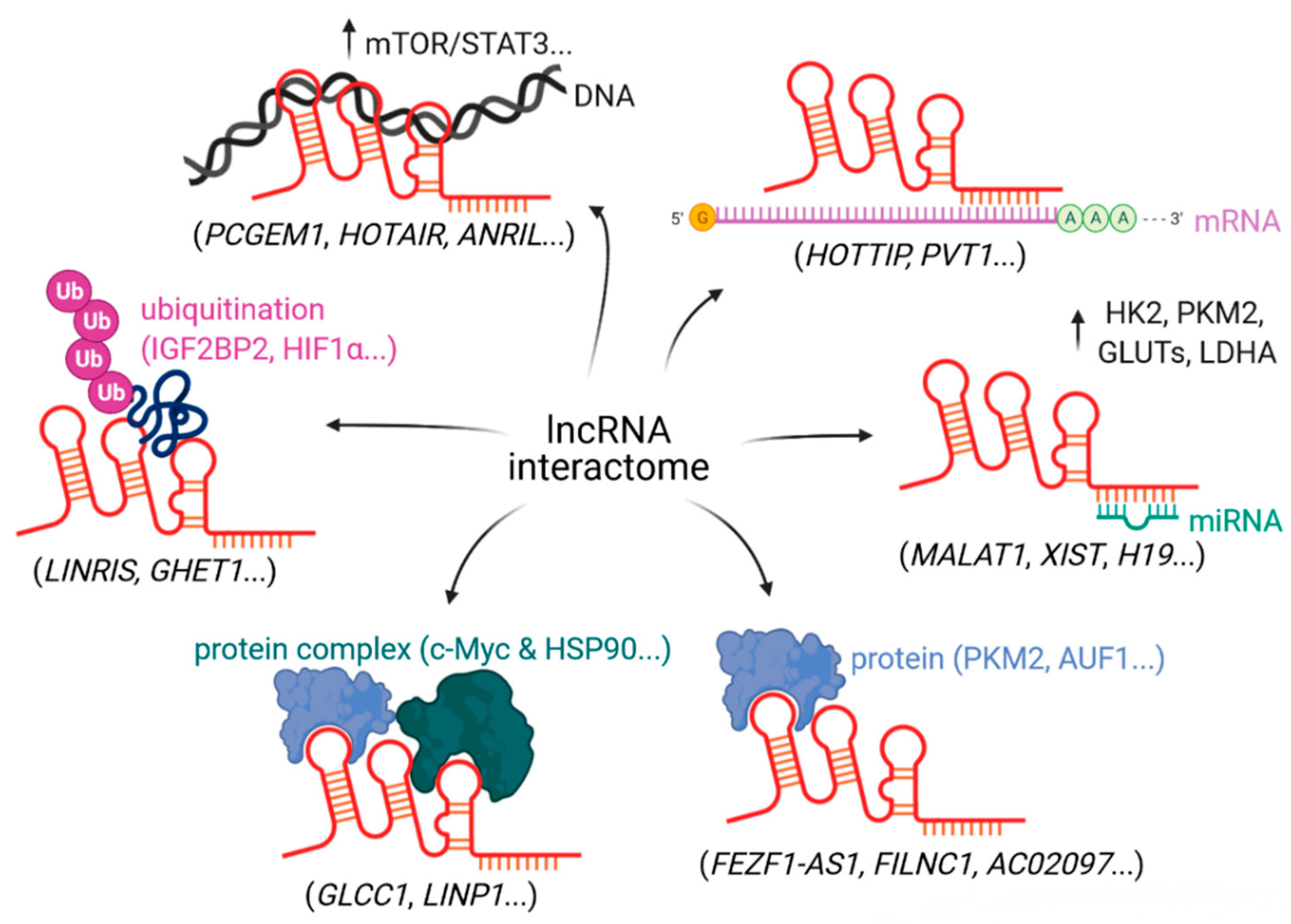{kind=link}
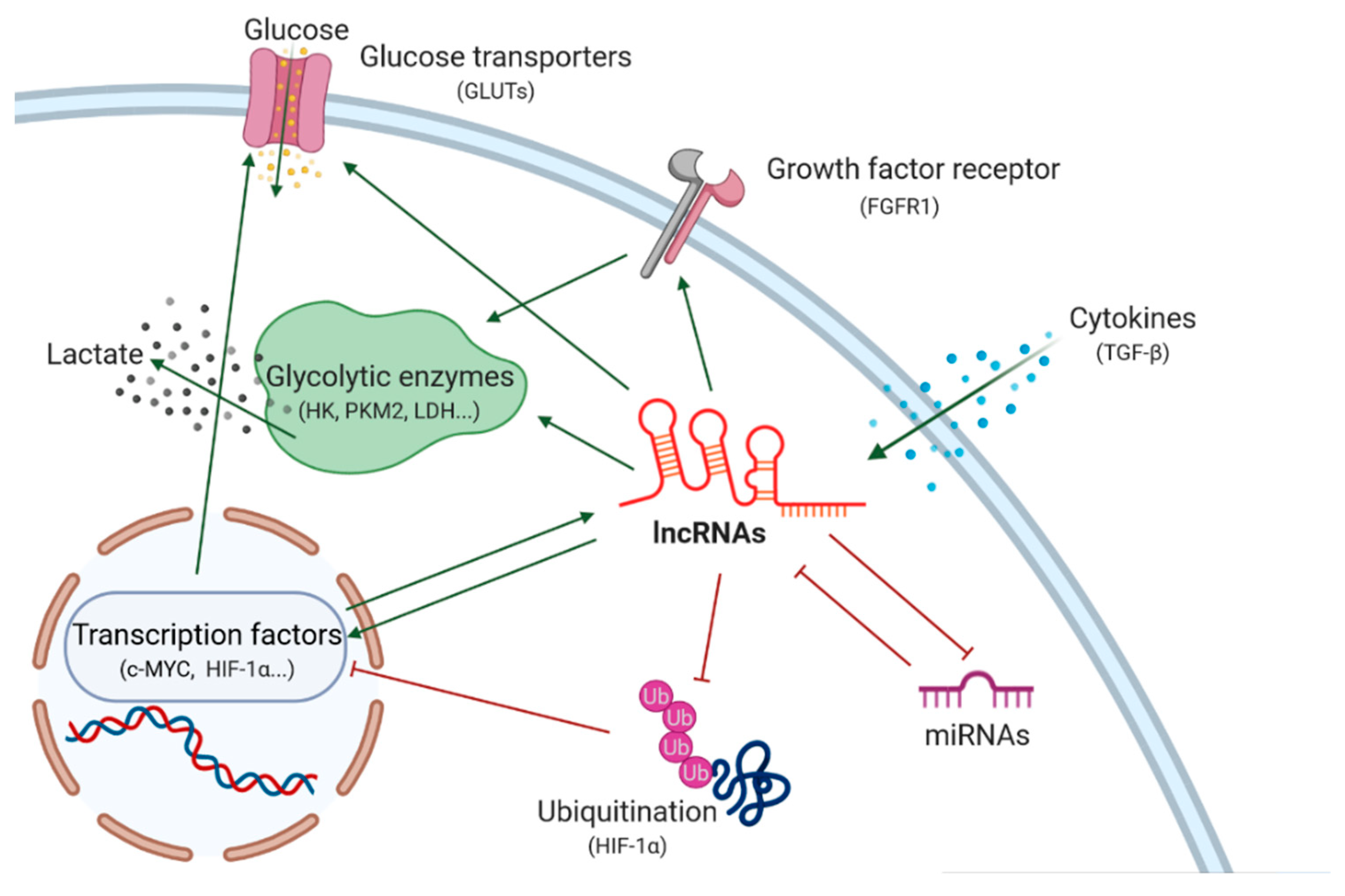{kind=link}
| lncRNA | Deregulation | Functions and Targets | Reference |
|---|---|---|---|
| AF339830 (GLCC1) | Upregulated in CRC | Stabilizes c-MYC; upregulates LDHA; increases glycolysis | [42] |
| LINRIS | Upregulated in CRC | Prevents IGF2BP2 degradation; stabilizies c-MYC; upregulates GLUT-1, PKM2 and LDHA expession | [44] |
| LINC00973 | Upregulated in cetuximab-resistant CRC cells | Increases glucose consumption and lactate production | [35] |
| LINC00504 | Upregulated in colon cancer | Increases c-MYC transcriptional activity; increases glycolysis, fatty acid synthesis, maintains redox status | [43] |
| FEZF1-AS1 | Upregulated in CRC | Stabilizes and promotes the activity of PKM2; upregulates glycolysis and STAT3 signaling | [49] |
| ENO1-IT1 | Fusobacterium nucleatum-activated lncRNA in CRC | Upregulates the alpha-enolase glycolytic enzyme | [30] |
| RAD51-AS1 | Downregulated in CRC | ceRNA for miR-29b-3p and miR-29c-3p; upregulates NDRG2; increases the levels of HK2 and GLUT-1, glucose uptake and lactate production | [45] |
| lncARSR | Upregulated in CRC | Sponges miR-34a-5p; upregulates HK1-mediated aerobic glycolysis | [48] |
| CRNDE | Upregulated in CRC | Upregulates G6PD, GLUT-4 and MLXIPL; increases glucose uptake and lactate production | [50] |
| LINC01554 | Downregulated in HCC | Promotes PKM2 degradation; inhibits Akt/mTOR signaling | [51] |
| MALAT1 | Upregulated in HCC | Upregulates TCF7L2 via the mTORC1 pathway; upregulates the expression of glycolytic enzymes | [52] |
| Ftx | Upregulated in HCC | Enhances the activity and expression of GLUT-1 and GLUT-4 via PPARγ | [53] |
| MEG3 | Downregulated in high glucose-treated HCC | Sponges miR-483-3p; suppresses ERp29 expression | [54] |
| UCA1 | Upregulated in HCC | Increases the expression of HK2; enhances glucose uptake, lactate and ATP production | [29] |
| RAET1K | Upregulated in HCC | Sponges miR-100-5p; upregulates HIF1α; suppresses hypoxia-induced glucose uptake and lactate secretion | [28] |
| HOTAIR | Upregulated in HCC | Upregulates GLUT-1; activates mTOR signaling pathway; decoy for miR-130a-3p (HIF1α target); promotes glycolysis and lactate production | [55,56] |
| DDX11-AS1 | Upregulated in HCC | Sponges miR-195-5p; increases MACC1 expression; increases glucose consumption and lactate production | [57] |
| SOX2OT | Upregulated in HCC | Sponges miR-122-5p; activates PKM2, HK2 and LDHA | [58] |
| DLX6-AS1 | Upregulated in gastric cancer | Sponges miR-4290; upregulates PDK1; elevates glycolysis rates; inhibits mitochondrial respiration | [59] |
| DGCR9 | Upregulated in gastric cancer | Increases glucose uptake | [60] |
| HOTAIR | Upregulated in pancreatic cancer | Upregulates HK2; increases glucose uptake, ATP and lactate production | [61] |
| LINC00184 | Upregulated in esophageal cancer | Silences PTEN; increases glycolysis and decreases mitochondrial oxidative phosphorylation | [62] |
| UCA1 | Upregulated in esophageal cancer | Sponges miR-203; suppresses HK2 degradation; increases glucose uptake and lactate output | [63] |
| PVT1 | Upregulated in gallbladder cancer | Sponges and repress miR-143; upregulates HK2; increases glucose uptake and lactate output | [64] |
| lncRNA | Deregulation | Functions and Targets | Reference |
|---|---|---|---|
| FILNC1 | Downregulated in renal cell cancer | Interacts with AUF1 (c-MYC mRNA-binding protein); downregulates c-MYC protein levels; decreases the glucose update, lactate production and tumor development | [79] |
| CASC8 | Downregulated in bladder cancer | Binds to the fibroblast growth factor receptor 1 (FGFR1); prevents FGFR1-mediated lactate dehydrogenase A phosphorylation; decreases glycolysis and cell proliferation | [80] |
| UCA1 | Upregulated in bladder cancer | Upregulates HK2 through mTOR and its downstream effector STAT3; upregulates HK2 by active mTOR downregulation of miR143 | [81,82] |
| PCGEM1 | Upregulated in prostate cancer | Binds to the c-MYC promotor; upregulates c-MYC; induces aerobic glycolysis, pentose phosphate shunt and NADPH | [83] |
| lncRNA | Deregulation | Functions and Targets | Reference |
|---|---|---|---|
| NRCP | Upregulated in ovarian cancers | Acts as an intermediate binder between STAT1 and RNA polymerase II; increases the expression of glucose-6-phosphate isomerase (GPI), aldolase A (ALDOA) and aldolase C (ALDOC) | [84] |
| GHET1 | Upregulated in ovarian cancers | Interacts with the E3 ubiquitin ligase von Hippel-Lindau (VHL); prevents VHL-mediated HIF1α-degradation; promotes glucose uptake and lactate production | [27] |
| H19 | Upregulated in ginsenoside 20(S)-Rg3-treated ovarian cancers | Sponges miR-324-5p; upregulates PKM2; increases glucose uptake and lactate production | [34] |
| UCA1 | Upregulated in radioresistant cervical cancer | Upregulates HK2 | [85] |
| lncRNA | Deregulation | Functions and Targets | Reference |
|---|---|---|---|
| BCAR4 | Upregulated in triple negative breast cancer | Interacts with GLI2; upregulates HK2 and PFKFB3; increases glucose uptake and lactate production | [88] |
| YIYA | Upregulated in breast cancer | Interacts with CDK6; increases phosphorylation PFKFB3; catalysis of glucose 6-phosphate to fructose bisphosphate | [86] |
| LINC00346 | Upregulated in breast cancer cells | Sponges miR-148a/b; enhances GLUT-1 expression | [89] |
| lncRNA | Deregulation | Functions and Targets | Reference |
|---|---|---|---|
| ANRIL | Upregulated in acute myeloid leukemia (AML) | Increases the expression of GLUT-1 and LDAH, upregulates AdipoR1, AMPK and SIRT1 | [90] |
| LINP1 | Upregulated in AML | Increases the expression of GLUT-1 and LDHA; upregulates HNF4α; activates the AMPK/WNT5A signaling pathway | [92] |
| UCA1 | Upregulated in AML | ceRNA of miR-125a; upregulates HK2; blocks HIF1α-dependent glycolysis | [32] |
| HOTAIRM1 | Upregulated in AML | Activates the Wnt/β-catenin pathway; enhances glucose uptake and lactate production | [33] |
| PDIA3P | Upregulated in multiple myeloma | Interacts with c-MYC; upregulates G6PD expression and PPP flux | [97] |
| MALAT1 | Upregulated in multiple myeloma | Targets miR-1271-5p; increases SOX13 expression | [98] |
| lncRNA | Deregulation | Functions and Targets | Reference |
|---|---|---|---|
| XIST | Upregulated in glioblastoma | Sponges miRNA126; upregulates the IRS1 and PI3K/Akt pathways; upregulates the GLUT-1 and GLUT-3 receptors; increases cellular glucose uptake and lactate production | [100] |
| LINC00174 | Upregulated in glioma | Sponges miR-1523-3p; upregulates SLC2A1 | [104] |
| SNHG9 | Upregulated in glioblastoma | Downregulates miR-199a-5p; upregulates the Wnt2 axis | [105] |
| SNHG1 | Upregulated in glioma | Sponges miR-194; upregulates PHLDA1 | [107] |
| SNHG5 | Upregulated in glioma | Sponges miR-205; upregulates E2F transcription factor 3 | [106] |
| TP53TG1 | Upregulated in glioma | Upregulates GRP78, LDHA and IDH1 | [108] |
| MACC1-AS1 | Upregulated in glioma | Increases the levels of GLUT-1, HK2, G6PD, MCT1, ATP, lactate and NAPDH | [109] |
| lncRNA | Deregulation | Functions and Targets | Reference |
|---|---|---|---|
| TUG1 | Upregulated in osteosarcoma | Increases HK2 expression, glucose consumption, lactate production | [110] |
| KCNQ1OT1 | Upregulated in osteosarcoma | Sponges miR-34c-5p; upregulates aldolase A (ALDOA) expression | [111] |
| PVT1 | Upregulated in osteosarcoma | Increases HK2 expression, glucose consumption, lactate production | [112] |
| HAND2-AS1 | Downregulated in osteosarcoma | Decreases GLUT-1 and glucose update | [113,114] |
| lncRNA | Deregulation | Functions and Targets | Reference |
|---|---|---|---|
| AC020978 | Upregulated in NSCLC | Interacts with and stabilizes PKM2; promotes PKM2 nuclear translocation; upregulates HIF1α, Oct-4, STAT3 and β-catenin | [25] |
| HOTTIP | Upregulated in NSCLC | Sponges miR-615-3p; increases HMGB3 expression; upregulates HK2 expression; increases glucose consumption and lactate production | [26] |
| MALAT1 | Upregulated in NSCLC | Sponges miR-613; upregulates COMMD8; increases glucose consumption and lactate production | [122] |
| BCYRN1 | Upregulated in NSCLC | Reduces miR‑149 expression; upregulates PKM2 expression; induces glycolysis | [124] |
| LINC00551 | Downregulated in lung adenocarcinoma | Decreases c-MYC expression; decreases PKM2; inhibits glycolysis | [125] |
| lncRNA | Deregulation | Functions and Targets | Reference |
|---|---|---|---|
| ANRIL | Upregulated in nasopharyngeal carcinoma | Upregulates mTOR signal pathway; increases GLUT-1 and LDHA expression | [126] |
| lnc-p23154 | Upregulated in oral squamous cell carcinoma | Sponges miR-378a-3p; upregulates GLUT-1 expression | [127] |
| UCA1 | Upregulated in tongue squamous cell carcinoma | Sponges miR-138-5p; upregulates CCR7 | [128] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balihodzic, A.; Barth, D.A.; Prinz, F.; Pichler, M. Involvement of Long Non-Coding RNAs in Glucose Metabolism in Cancer. Cancers 2021, 13, 977. https://doi.org/10.3390/cancers13050977
Balihodzic A, Barth DA, Prinz F, Pichler M. Involvement of Long Non-Coding RNAs in Glucose Metabolism in Cancer. Cancers. 2021; 13(5):977. https://doi.org/10.3390/cancers13050977
Chicago/Turabian StyleBalihodzic, Amar, Dominik A. Barth, Felix Prinz, and Martin Pichler. 2021. "Involvement of Long Non-Coding RNAs in Glucose Metabolism in Cancer" Cancers 13, no. 5: 977. https://doi.org/10.3390/cancers13050977