
Molecular Classification of Breast Cancer Utilizing Long Non-Coding RNA (lncRNA) Transcriptomes Identifies Novel Diagnostic lncRNA Panel for Triple-Negative Breast Cancer


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. RNA-Seq Data Analysis
2.2. ROC and Binary Regression Analysis
2.3. Survival and Statistical Analysis
3. Results
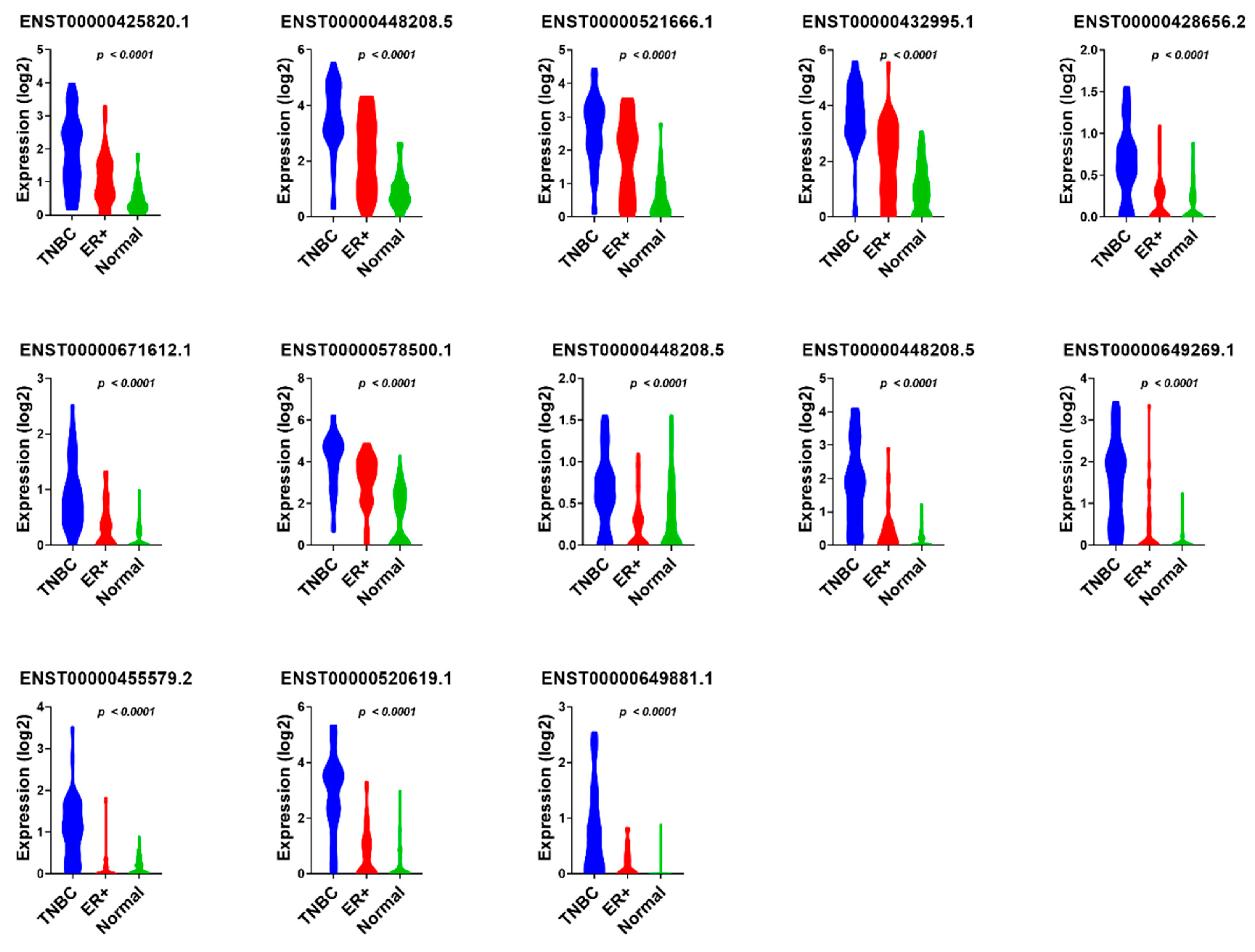
3.1. Identification of lncRNA-Based Biomarkers Associated with TNBC, ER+, and Normal Breast Tissue
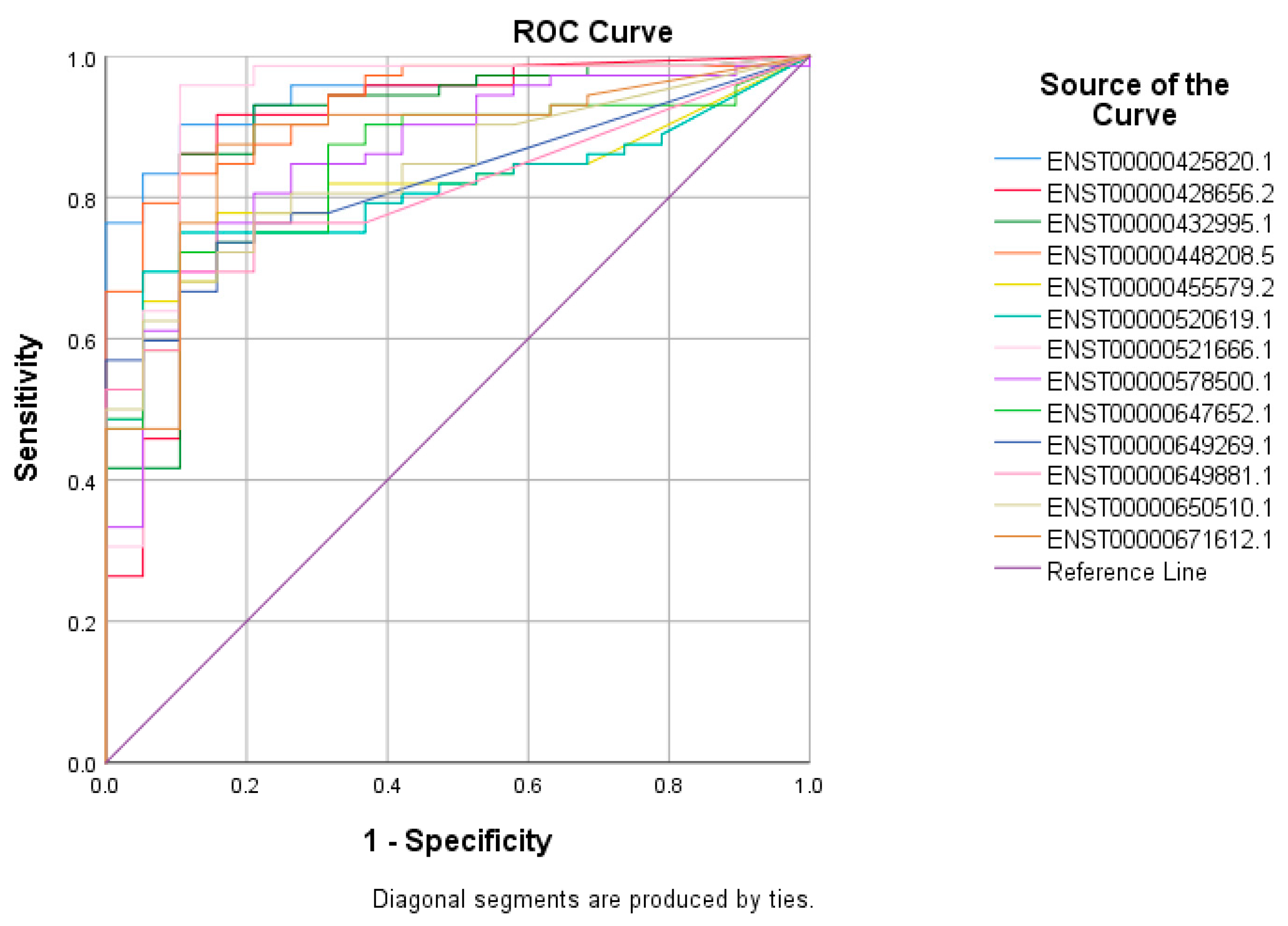
3.2. Receiver Operating Characteristic (ROC) Curves for Putative lncRNA Markers Associated with TNBC, ER+, and Normal Breast Tissue
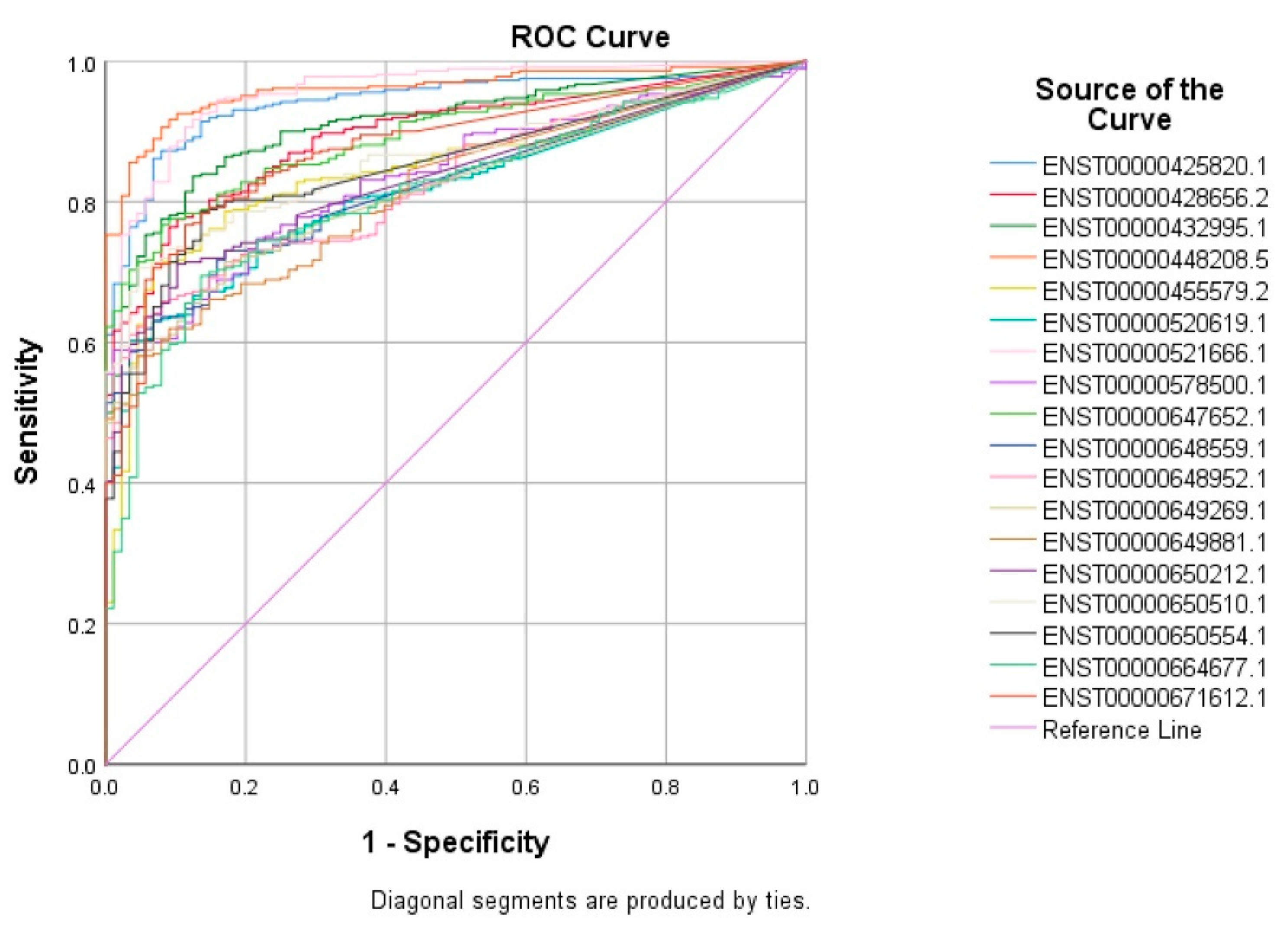
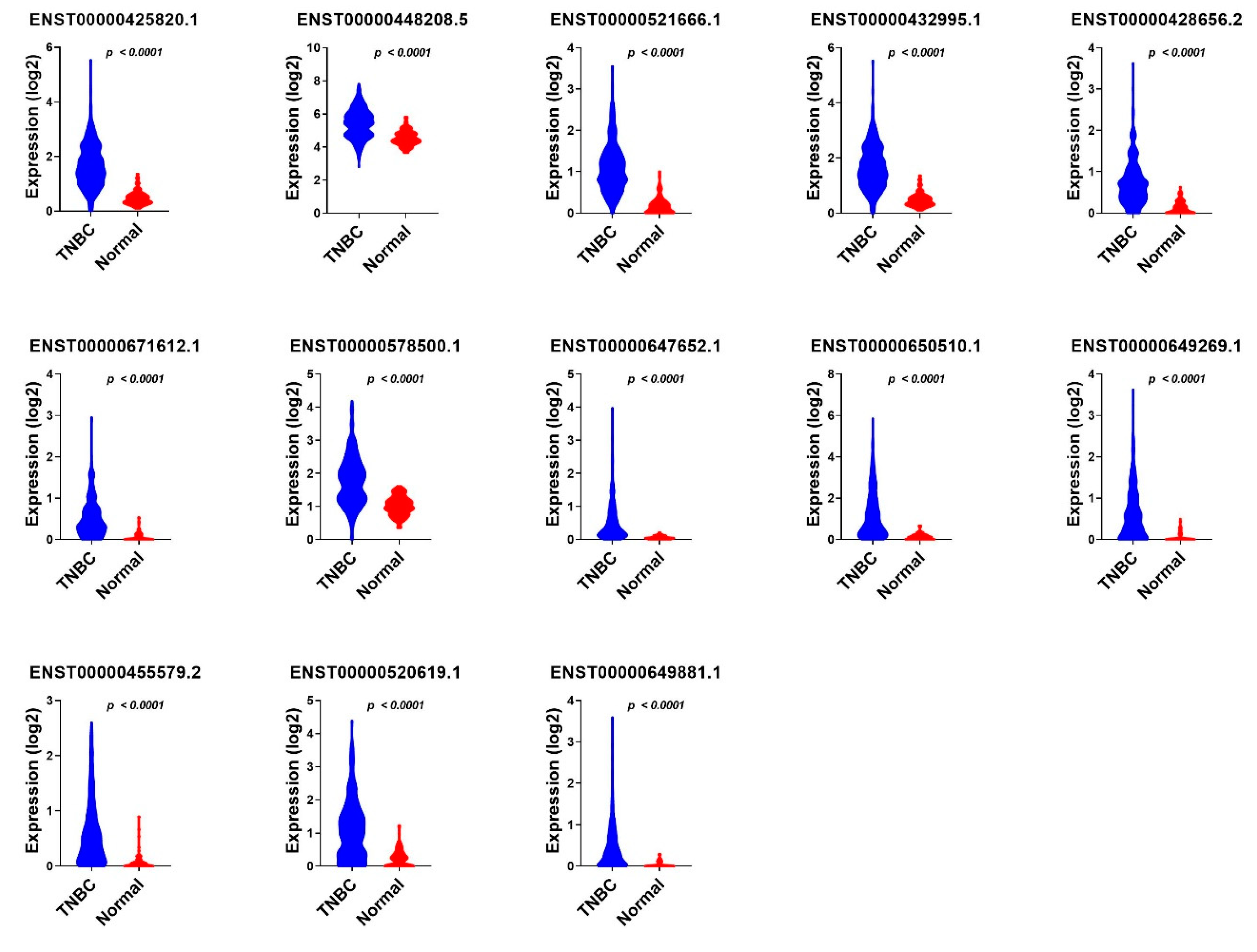
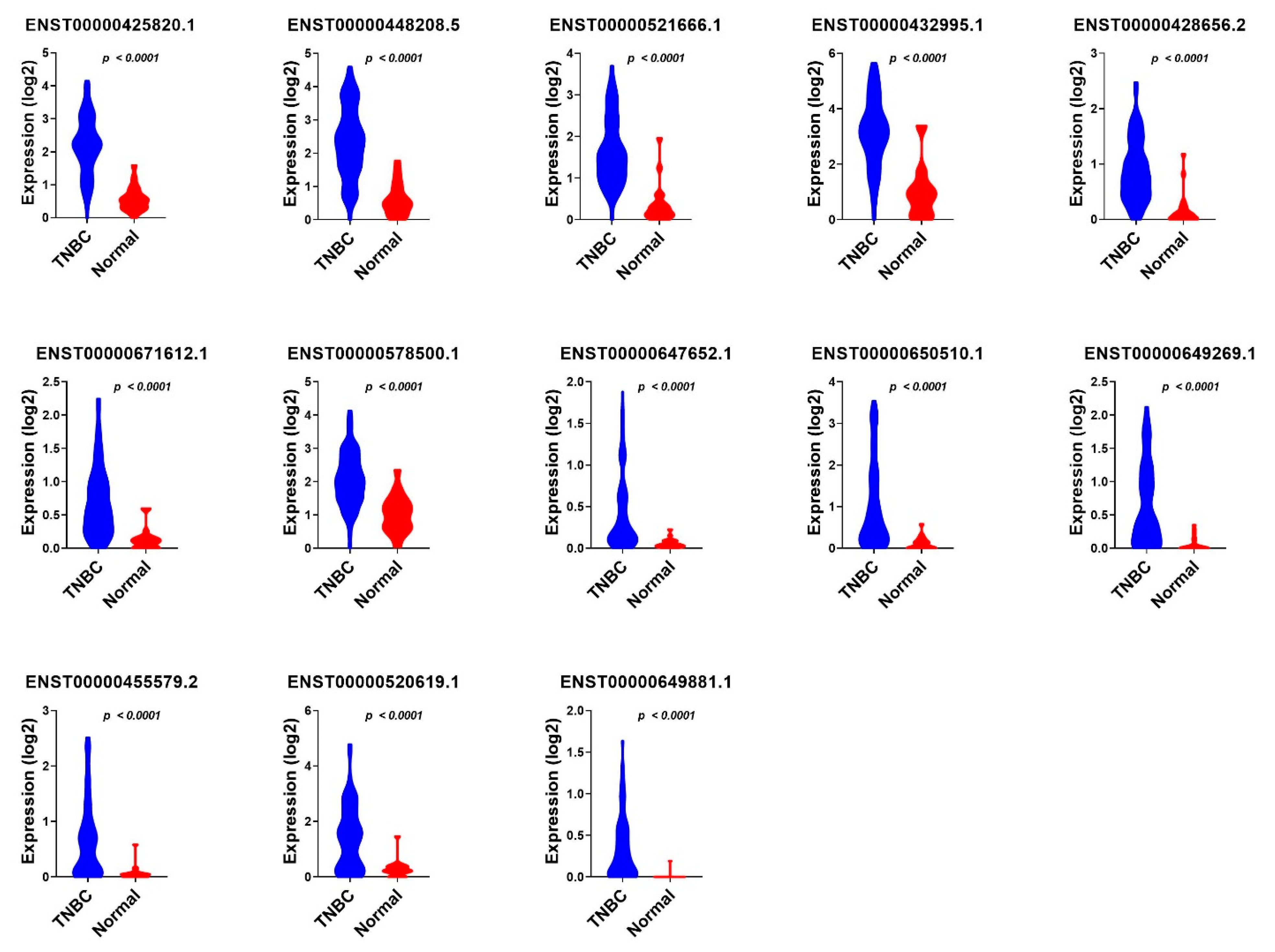
3.3. Validation of Common lncRNA Markers in the Second Cohort of TNBC and Normal Breast Tissue
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2021. CA Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef]
- World Health Organisation. Breast Cancer. (Online) Who.int. 2021. Available online: https://www.who.int/news-room/fact-sheets/detail/breast-cancer (accessed on 25 October 2021).
- Ramamoorthy, P.; Dandawate, P.; Jensen, R.A.; Anant, S. Celastrol and Triptolide Suppress Stemness in Triple Negative Breast Cancer: Notch as a Therapeutic Target for Stem Cells. Biomedicines 2021, 9, 482. [Google Scholar] [CrossRef] [PubMed]
- Grizzi, G.; Ghidini, M.; Botticelli, A.; Tomasello, G.; Ghidini, A.; Grossi, F.; Fusco, N.; Cabiddu, M.; Savio, T.; Petrelli, F. Strategies for Increasing the Effectiveness of Aromatase Inhibitors in Locally Advanced Breast Cancer: An Evidence-Based Review on Current Options. Cancer Manag. Res. 2020, 12, 675–686. [Google Scholar] [CrossRef] [Green Version]
- Llombart-Cussac, A.; Cortes, J.; Pare, L.; Galvan, P.; Bermejo, B.; Martinez, N.; Vidal, M.; Pernas, S.; Lopez, R.; Munoz, M.; et al. HER2-enriched subtype as a predictor of pathological complete response following trastuzumab and lapatinib without chemotherapy in early-stage HER2-positive breast cancer (PAMELA): An open-label, single-group, multicentre, phase 2 trial. Lancet Oncol. 2017, 18, 545–554. [Google Scholar] [CrossRef]
- Kagihara, J.A.; Shagisultanova, E.; Afghahi, A.; Diamond, J.R. Moving Towards Targeted Therapies for Triple-Negative Breast Cancer. Curr. Breast Cancer Rep. 2021, 13, 216–226. [Google Scholar] [CrossRef] [PubMed]
- O’Halloran, N.; Lowery, A.; Curran, C.; McLaughlin, R.; Malone, C.; Sweeney, K.; Keane, M.; Kerin, M. A Review of the Impact of Neoadjuvant Chemotherapy on Breast Surgery Practice and Outcomes. Clin. Breast Cancer 2019, 19, 377–382. [Google Scholar] [CrossRef] [PubMed]
- Boughey, J.C.; McCall, L.M.; Ballman, K.V.; Mittendorf, E.A.; Ahrendt, G.M.; Wilke, L.G.; Taback, B.; Leitch, A.M.; Flippo-Morton, T.; Hunt, K.K. Tumor biology correlates with rates of breast-conserving surgery and pathologic complete response after neoadjuvant chemotherapy for breast cancer: Findings from the ACOSOG Z1071 (Alliance) Prospective Multicenter Clinical Trial. Ann. Surg. 2014, 260, 608–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esserman, L.J.; Berry, D.A.; DeMichele, A.; Carey, L.; Davis, S.E.; Buxton, M.; Hudis, C.; Gray, J.W.; Perou, C.; Yau, C.; et al. Pathologic complete response predicts recurrence-free survival more effectively by cancer subset: Results from the I-SPY 1 TRIAL—CALGB 150007/150012, ACRIN 6657. J. Clin. Oncol. 2012, 30, 3242–3249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- von Minckwitz, G.; Untch, M.; Blohmer, J.U.; Costa, S.D.; Eidtmann, H.; Fasching, P.A.; Gerber, B.; Eiermann, W.; Hilfrich, J.; Huober, J.; et al. Definition and impact of pathologic complete response on prognosis after neoadjuvant chemotherapy in various intrinsic breast cancer subtypes. J. Clin. Oncol. 2012, 30, 1796–1804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asaoka, M.; Narui, K.; Suganuma, N.; Chishima, T.; Yamada, A.; Sugae, S.; Kawai, S.; Uenaka, N.; Teraoka, S.; Miyahara, K.; et al. Clinical and pathological predictors of recurrence in breast cancer patients achieving pathological complete response to neoadjuvant chemotherapy. Eur. J. Surg. Oncol. 2019, 45, 2289–2294. [Google Scholar] [CrossRef] [PubMed]
- Liedtke, C.; Mazouni, C.; Hess, K.R.; Andre, F.; Tordai, A.; Mejia, J.A.; Symmans, W.F.; Gonzalez-Angulo, A.M.; Hennessy, B.; Green, M.; et al. Response to neoadjuvant therapy and long-term survival in patients with triple-negative breast cancer. J. Clin. Oncol. 2008, 26, 1275–1281. [Google Scholar] [CrossRef] [PubMed]
- Vishnubalaji, R.; Shaath, H.; Elkord, E.; Alajez, N.M. Long non-coding RNA (lncRNA) transcriptional landscape in breast cancer identifies LINC01614 as non-favorable prognostic biomarker regulated by TGFbeta and focal adhesion kinase (FAK) signaling. Cell Death Discov. 2019, 5, 109. [Google Scholar] [CrossRef] [PubMed]
- Shin, V.Y.; Chen, J.; Cheuk, I.W.; Siu, M.T.; Ho, C.W.; Wang, X.; Jin, H.; Kwong, A. Long non-coding RNA NEAT1 confers oncogenic role in triple-negative breast cancer through modulating chemoresistance and cancer stemness. Cell Death Dis. 2019, 10, 270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, S.; Wang, F.; Shen, J.; Sun, Y.; Xu, W.; Lu, J.; Wei, M.; Xu, C.; Wu, C.; Zhang, Z.; et al. Long non-coding RNA metastasis associated in lung adenocarcinoma transcript 1 derived miniRNA as a novel plasma-based biomarker for diagnosing prostate cancer. Eur. J. Cancer 2013, 49, 2949–2959. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Yin, C.; Dang, Y.; Ye, F.; Zhang, G. Identification of the long non-coding RNA H19 in plasma as a novel biomarker for diagnosis of gastric cancer. Sci. Rep. 2015, 5, 11516. [Google Scholar] [CrossRef]
- Trimarchi, T.; Bilal, E.; Ntziachristos, P.; Fabbri, G.; Dalla-Favera, R.; Tsirigos, A.; Aifantis, I. Genome-wide mapping and characterization of Notch-regulated long noncoding RNAs in acute leukemia. Cell 2014, 158, 593–606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhan, A.; Soleimani, M.; Mandal, S.S. Long Noncoding RNA and Cancer: A New Paradigm. Cancer Res. 2017, 77, 3965–3981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, M.C.; Spitale, R.C.; Chang, H.Y. Long intergenic noncoding RNAs: New links in cancer progression. Cancer Res. 2011, 71, 3–7. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Wang, X. Role of long noncoding RNAs in malignant disease (Review). Mol. Med. Rep. 2016, 13, 1463–1469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, X.; Malouf, G.G.; Chen, Y.; Zhang, J.; Yao, H.; Valero, V.; Weinstein, J.N.; Spano, J.P.; Meric-Bernstam, F.; Khayat, D.; et al. Comprehensive analysis of long non-coding RNAs in human breast cancer clinical subtypes. Oncotarget 2014, 5, 9864–9876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Sharma, S.; Watabe, K. Roles of lncRNA in breast cancer. Front. Biosci. 2015, 7, 94–108. [Google Scholar] [CrossRef]
- Shaath, H.; Toor, S.M.; Nair, V.S.; Elkord, E.; Alajez, N.M. Transcriptomic Analyses Revealed Systemic Alterations in Gene Expression in Circulation and Tumor Microenvironment of Colorectal Cancer Patients. Cancers 2019, 11, 1994. [Google Scholar] [CrossRef] [Green Version]
- Bray, N.L.; Pimentel, H.; Melsted, P.; Pachter, L. Near-optimal probabilistic RNA-seq quantification. Nat. Biotechnol. 2016, 34, 525–527. [Google Scholar] [CrossRef] [PubMed]
- Olsson, A.; Venkatasubramanian, M.; Chaudhri, V.K.; Aronow, B.J.; Salomonis, N.; Singh, H.; Grimes, H.L. Single-cell analysis of mixed-lineage states leading to a binary cell fate choice. Nature 2016, 537, 698–702. [Google Scholar] [CrossRef]
- Hallett, R.M.; Dvorkin-Gheva, A.; Bane, A.; Hassell, J.A. A gene signature for predicting outcome in patients with basal-like breast cancer. Sci. Rep. 2012, 2, 227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perou, C.M.; Sorlie, T.; Eisen, M.B.; van de Rijn, M.; Jeffrey, S.S.; Rees, C.A.; Pollack, J.R.; Ross, D.T.; Johnsen, H.; Akslen, L.A.; et al. Molecular portraits of human breast tumours. Nature 2000, 406, 747–752. [Google Scholar] [CrossRef] [PubMed]
- Vishnubalaji, R.; Alajez, N.M. Transcriptional landscape associated with triple negative breast cancer (TNBC) resistance to neoadjuvant chemotherapy revealed by single cell RNA-Seq. Mol. Ther.-Oncolytics 2021, 23, 151–162. [Google Scholar] [CrossRef]
- Vishnubalaji, R.; Alajez, N.M. Epigenetic regulation of triple negative breast cancer (TNBC) by TGF-beta signaling. Sci. Rep. 2021, 11, 15410. [Google Scholar] [CrossRef] [PubMed]
- Shaath, H.; Vishnubalaji, R.; Elango, R.; Khattak, S.; Alajez, N.M. Single-cell long noncoding RNA (lncRNA) transcriptome implicates MALAT1 in triple-negative breast cancer (TNBC) resistance to neoadjuvant chemotherapy. Cell Death Discov. 2021, 7, 23. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Sui, S.; Wu, H.; Zhang, J.; Zhang, X.; Xu, S.; Pang, D. The transcriptional landscape of lncRNAs reveals the oncogenic function of LINC00511 in ER-negative breast cancer. Cell Death Dis. 2019, 10, 599. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Mao, X.; Luo, F.; Wang, J. LINC00511 promotes gastric cancer progression by regulating SOX4 and epigenetically repressing PTEN to activate PI3K/AKT pathway. J. Cell. Mol. Med. 2021, 25, 9112–9127. [Google Scholar] [CrossRef] [PubMed]
- Cui, N.; Sun, Q.; Liu, H.; Li, L.; Guo, X.; Shi, Y.; Jing, C.; Qin, C.; Zhao, Y. Long non-coding RNA LINC00511 regulates the expression of microRNA-625-5p and activates signal transducers and activators of transcription 3 (STAT3) to accelerate the progression of gastric cancer. Bioengineered 2021, 12, 2915–2927. [Google Scholar] [CrossRef]
- Hu, Y.; Zhang, Y.; Ding, M.; Xu, R. LncRNA LINC00511 Acts as an Oncogene in Colorectal Cancer via Sponging miR-29c-3p to Upregulate NFIA. OncoTargets Ther. 2020, 13, 13413–13424. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Tian, M.; Liu, J.; Wang, K. LINC00511 facilitates Temozolomide resistance of glioblastoma cells via sponging miR-126-5p and activating Wnt/beta-catenin signaling. J. Biochem. Mol. Toxicol. 2021, 35, e22848. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Tao, B.; Li, L.; Liu, P.; Xia, K.; Zhong, C. LINC00511 knockdown suppresses glioma cell malignant progression through miR-15a-5p/AEBP1 axis. Brain Res. Bull. 2021, 173, 82–96. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Xu, Y.; Liu, X.; Fu, Y.; Zhu, K.; Niu, Z.; Liu, J.; Qian, C. Upregulation of LINC00511 expression by DNA hypomethylation promotes the progression of breast cancer. Gland Surg. 2021, 10, 1418–1430. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Li, X.; Yang, S.; Huang, M.; Wei, S.; Ma, Y.; Li, Y.; Wu, B.; Jin, H.; Li, B.; et al. LINC00511 drives invasive behavior in hepatocellular carcinoma by regulating exosome secretion and invadopodia formation. J. Exp. Clin. Cancer Res. 2021, 40, 183. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.M.; Zhang, X.L.; Mao, M.H.; Li, Y.P.; Zhang, X.Y.; Xue, D.W.; Liu, Y.L. LINC00511/miRNA-143-3p Modulates Apoptosis and Malignant Phenotype of Bladder Carcinoma Cells via PCMT1. Front. Cell Dev. Biol. 2021, 9, 650999. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.; Li, Z.; Xue, L.; Li, G.; Zhang, C.; Cai, Z.; Li, H.; Guo, R. DIAPH3 promoted the growth, migration and metastasis of hepatocellular carcinoma cells by activating beta-catenin/TCF signaling. Mol. Cell Biochem. 2018, 438, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Xiang, G.; Weiwei, H.; Erji, G.; Haitao, M. DIAPH3 promotes the tumorigenesis of lung adenocarcinoma. Exp. Cell Res. 2019, 385, 111662. [Google Scholar] [CrossRef] [PubMed]
- Rong, Y.; Gao, J.; Kuang, T.; Chen, J.; Li, J.A.; Huang, Y.; Xin, H.; Fang, Y.; Han, X.; Sun, L.Q.; et al. DIAPH3 promotes pancreatic cancer progression by activating selenoprotein TrxR1-mediated antioxidant effects. J. Cell. Mol. Med. 2021, 25, 2163–2175. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J. Diaphanous-related formin-3 overexpression inhibits the migration and invasion of triple-negative breast cancer by inhibiting RhoA-GTP expression. Biomed. Pharmacother. 2017, 94, 439–445. [Google Scholar] [CrossRef]
- Lv, Y.; Lv, X.; Yang, H.; Qi, X.; Wang, X.; Li, C.; Shang, X.; Guo, H.; Zhang, J.; Zhang, Y. LncRNA SNHG6/miR-125b-5p/BMPR1B Axis: A New Therapeutic Target for Triple-Negative Breast Cancer. Front. Oncol. 2021, 11, 678474. [Google Scholar] [CrossRef] [PubMed]
- Sane, A.; Sridhar, S.; Sanyal, K.; Ghosh, S.K. Shugoshin ensures maintenance of the spindle assembly checkpoint response and efficient spindle disassembly. Mol. Microbiol. 2021, 116, 1079–1098. [Google Scholar] [CrossRef] [PubMed]
- Asad Samani, M.; Peymani, M. Changes in the Expression of SGO1 and SGO1-AS1 Genes in Colorectal Tumor Tissues, Compared to Healthy Tissues. J. Arak Univ. Med. Sci. 2021, 24, 168–179. [Google Scholar] [CrossRef]
- Nasim, N.; Ghafouri-Fard, S.; Soleimani, S.; Esfandi, F.; Shirkhoda, M.; Safaei, M.; Oskooei, V.K.; Taheri, M.; Raheb, J. Assessment of SGO1 and SGO1-AS1 contribution in breast cancer. Hum. Antib. 2019, 27, 279–284. [Google Scholar] [CrossRef]
- Zhang, H.; Ren, L.; Ding, Y.; Li, F.; Chen, X.; Ouyang, Y.; Zhang, Y.; Zhang, D. Hyaluronan-mediated motility receptor confers resistance to chemotherapy via TGFbeta/Smad2-induced epithelial-mesenchymal transition in gastric cancer. FASEB J. 2019, 33, 6365–6377. [Google Scholar] [CrossRef]
- Schatz-Siemers, N.; Chen, Y.T.; Chen, Z.; Wang, D.; Ellenson, L.H.; Du, Y.N. Expression of the Receptor for Hyaluronic Acid-Mediated Motility (RHAMM) in Endometrial Cancer is Associated With Adverse Histologic Parameters and Tumor Progression. Appl. Immunohistochem. Mol. Morphol. 2020, 28, 453–459. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Ji, X.; Wang, H. Targeting Long Noncoding RNA HMMR-AS1 Suppresses and Radiosensitizes Glioblastoma. Neoplasia 2018, 20, 456–466. [Google Scholar] [CrossRef]
- Chu, Z.P.; Dai, J.; Jia, L.G.; Li, J.; Zhang, Y.; Zhang, Z.Y.; Yan, P. Increased expression of long noncoding RNA HMMR-AS1 in epithelial ovarian cancer: An independent prognostic factor. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 8145–8150. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Sheng, Z.; Chen, Y.; Wang, J. LncRNA HMMR-AS1 promotes proliferation and metastasis of lung adenocarcinoma by regulating MiR-138/sirt6 axis. Aging (Albany NY) 2019, 11, 3041–3054. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Ma, J.; Cheng, Y.; Zhang, H.; Luo, W.; Zhang, H. HMMR antisense RNA 1, a novel long noncoding RNA, regulates the progression of basal-like breast cancer cells. Breast Cancer (Dove Med. Press) 2016, 8, 223–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Battistelli, C.; Garbo, S.; Riccioni, V.; Montaldo, C.; Santangelo, L.; Vandelli, A.; Strippoli, R.; Tartaglia, G.G.; Tripodi, M.; Cicchini, C. Design and Functional Validation of a Mutant Variant of the LncRNA HOTAIR to Counteract Snail Function in Epithelial-to-Mesenchymal Transition. Cancer Res. 2021, 81, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Zuroske, T.; Watts, J.K. RNA therapeutics on the rise. Nat. Rev. Drug Discov. 2020, 19, 441–442. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| B | S.E. | Wald | df | Sig. | Exp(B) | 95% C.I. for EXP(B) | |||
|---|---|---|---|---|---|---|---|---|---|
| Lower | Upper | ||||||||
| Step 1 a | ENST00000448208.5 | 5.227 | 0.636 | 67.531 | 1 | 0.000 | 186.228 | 53.534 | 647.832 |
| Constant | −2.794 | 0.400 | 48.786 | 1 | 0.000 | 0.061 | |||
| Step 2 b | ENST00000448208.5 | 3.516 | 0.711 | 24.418 | 1 | 0.000 | 33.643 | 8.342 | 135.681 |
| ENST00000521666.1 | 4.179 | 0.914 | 20.906 | 1 | 0.000 | 65.290 | 10.887 | 391.561 | |
| Constant | −3.367 | 0.466 | 52.301 | 1 | 0.000 | 0.034 | |||
| Step 3 c | ENST00000448208.5 | 3.392 | 0.764 | 19.704 | 1 | 0.000 | 29.725 | 6.648 | 132.912 |
| ENST00000521666.1 | 4.198 | 0.935 | 20.153 | 1 | 0.000 | 66.557 | 10.646 | 416.090 | |
| ENST00000650510.1 | 3.222 | 0.983 | 10.743 | 1 | 0.001 | 25.069 | 3.651 | 172.103 | |
| Constant | −4.074 | 0.563 | 52.441 | 1 | 0.000 | 0.017 | |||
| Step 4 d | ENST00000425820.1 | 1.262 | 0.587 | 4.622 | 1 | 0.032 | 3.533 | 1.118 | 11.166 |
| ENST00000448208.5 | 3.146 | 0.836 | 14.166 | 1 | 0.000 | 23.242 | 4.516 | 119.606 | |
| ENST00000521666.1 | 3.085 | 1.036 | 8.861 | 1 | 0.003 | 21.858 | 2.868 | 166.574 | |
| ENST00000650510.1 | 3.338 | 0.983 | 11.525 | 1 | 0.001 | 28.175 | 4.100 | 193.615 | |
| Constant | −4.498 | 0.617 | 53.132 | 1 | 0.000 | 0.011 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shaath, H.; Elango, R.; Alajez, N.M. Molecular Classification of Breast Cancer Utilizing Long Non-Coding RNA (lncRNA) Transcriptomes Identifies Novel Diagnostic lncRNA Panel for Triple-Negative Breast Cancer. Cancers 2021, 13, 5350. https://doi.org/10.3390/cancers13215350
Shaath H, Elango R, Alajez NM. Molecular Classification of Breast Cancer Utilizing Long Non-Coding RNA (lncRNA) Transcriptomes Identifies Novel Diagnostic lncRNA Panel for Triple-Negative Breast Cancer. Cancers. 2021; 13(21):5350. https://doi.org/10.3390/cancers13215350
Chicago/Turabian StyleShaath, Hibah, Ramesh Elango, and Nehad M. Alajez. 2021. "Molecular Classification of Breast Cancer Utilizing Long Non-Coding RNA (lncRNA) Transcriptomes Identifies Novel Diagnostic lncRNA Panel for Triple-Negative Breast Cancer" Cancers 13, no. 21: 5350. https://doi.org/10.3390/cancers13215350