
High miR-30 Expression Associates with Improved Breast Cancer Patient Survival and Treatment Outcome

 , , , ,
, , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients
2.2. miRNA In Situ Hybridization
2.3. Gene Expression and Pathway Enrichment Analyses
2.4. Sensitization Screening and Drug Response
2.5. Analysis of Tumor Phenotype and Patient Survival
3. Results
3.1. High miR-30d Expression Is Associated with Aggressive Clinical and Pathological Features of the Tumors
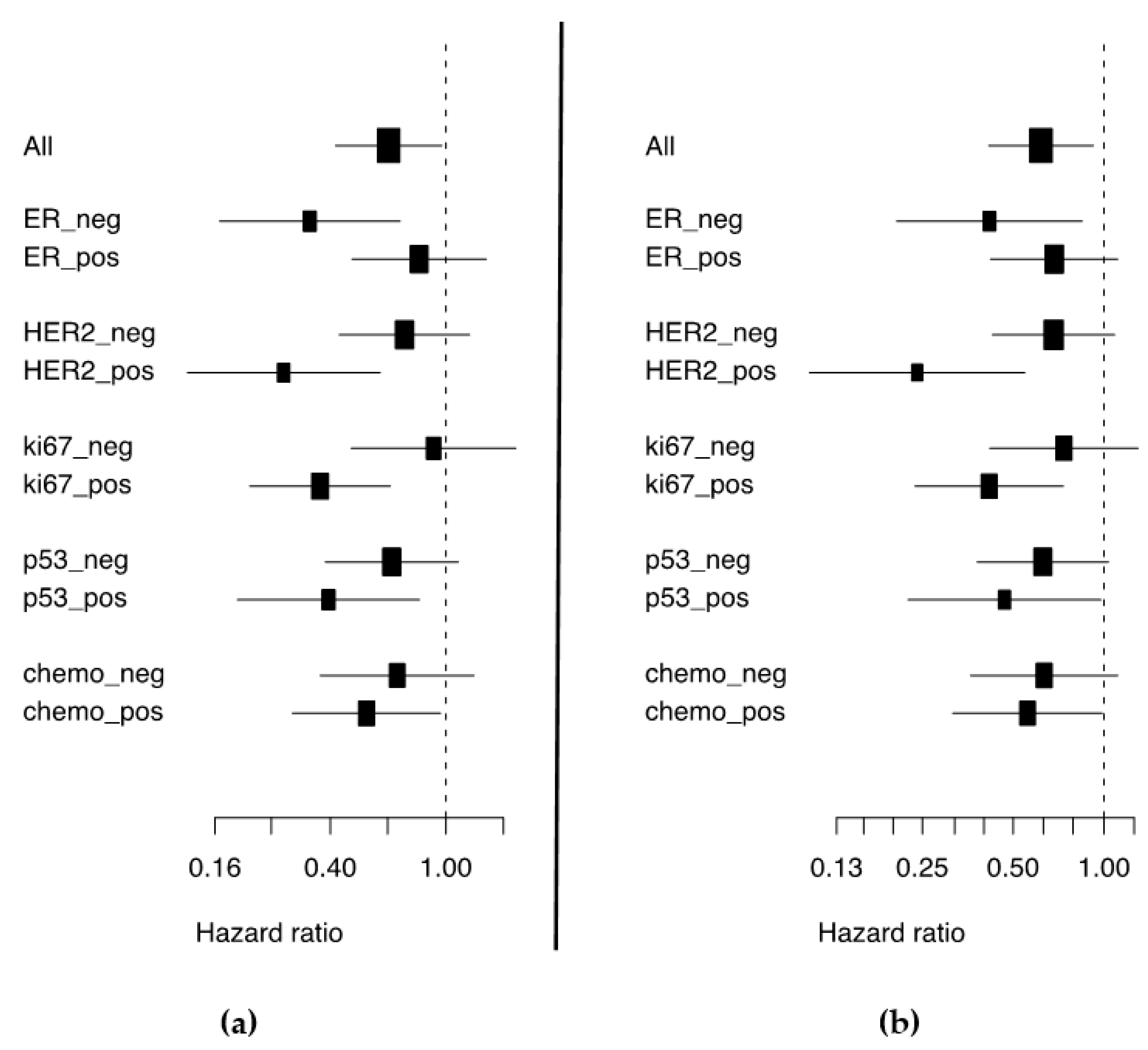
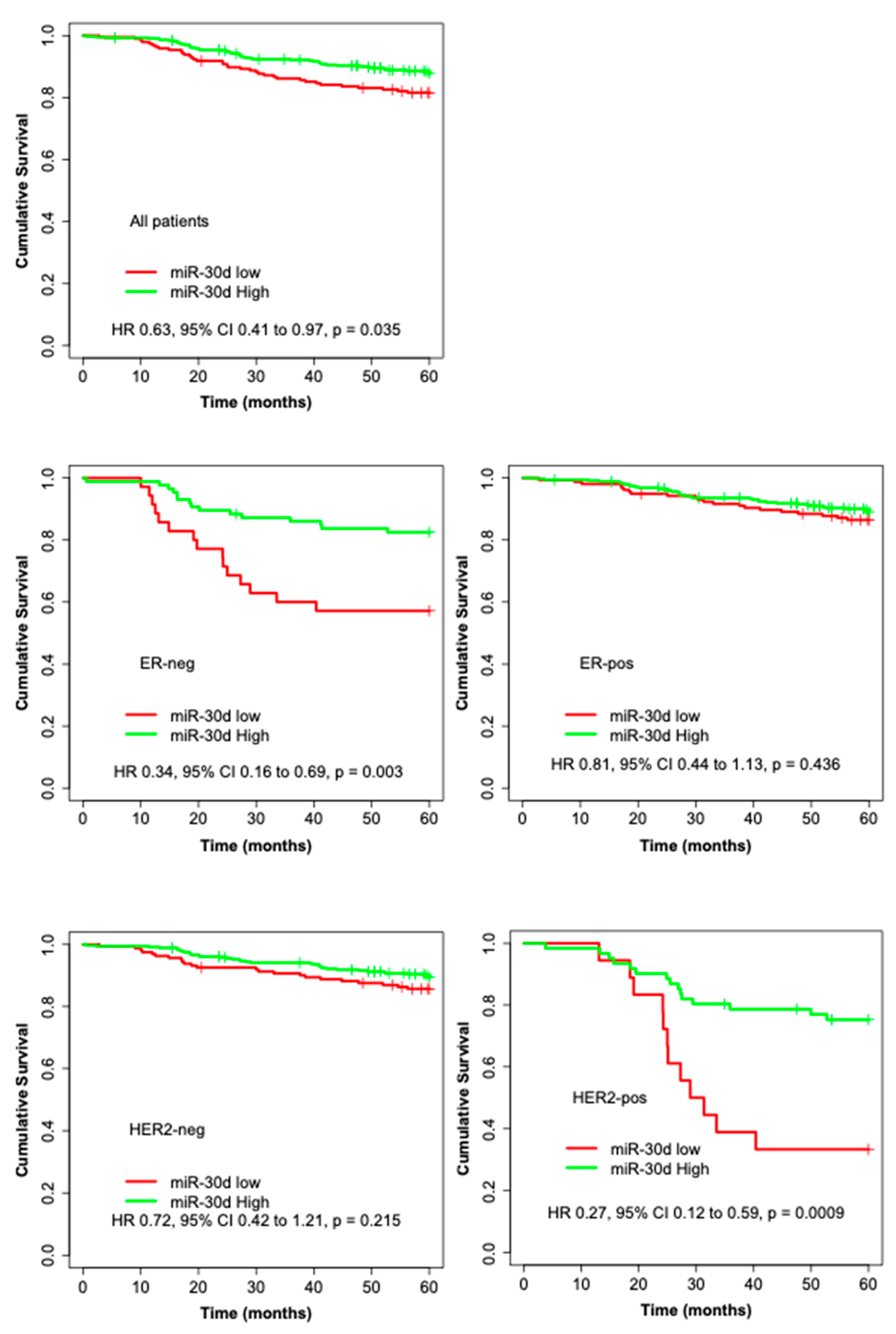
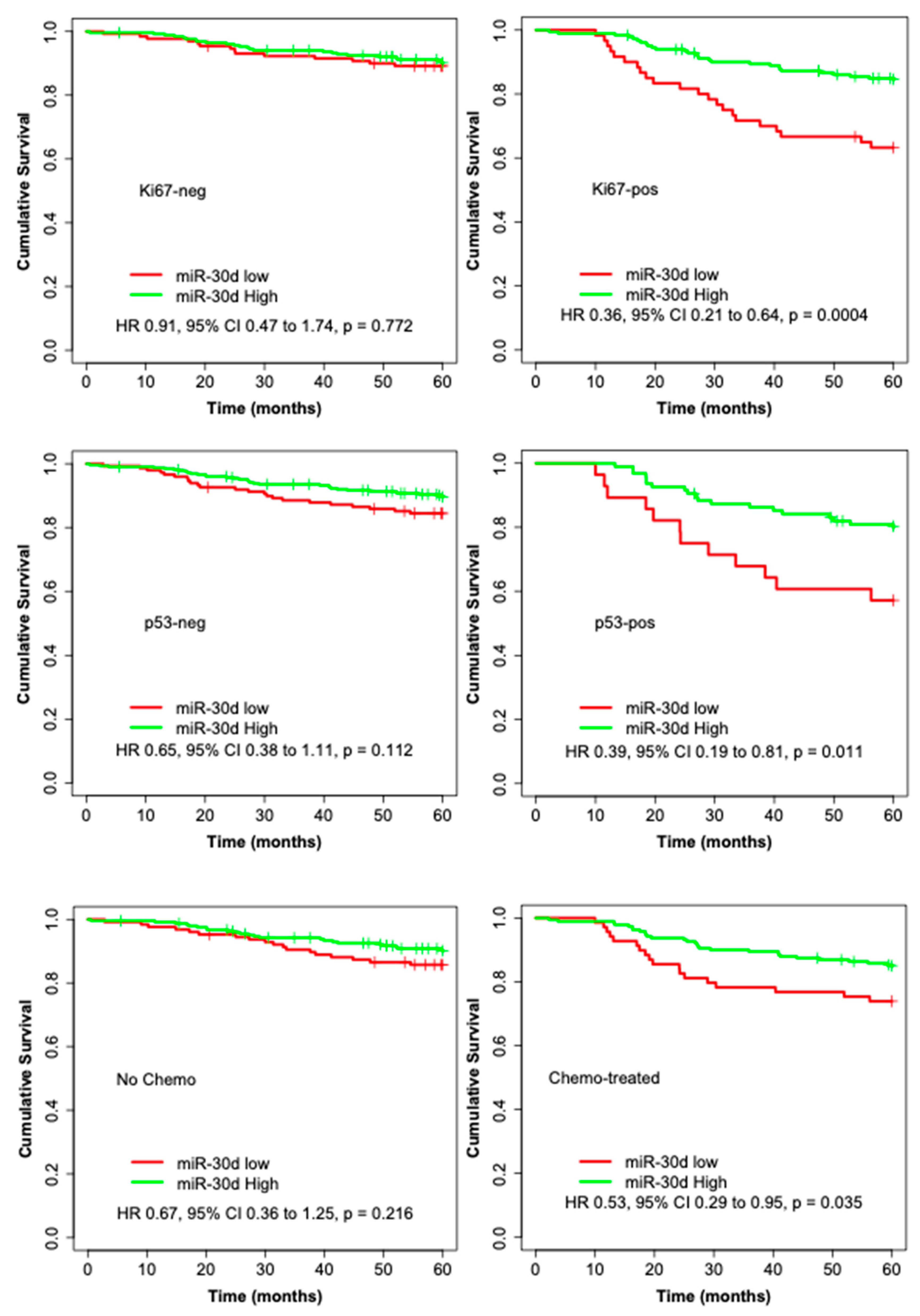
3.2. High miR-30d Expression Is an Independent Predictor of Improved Survival in Breast Cancer Patients and among Subgroups
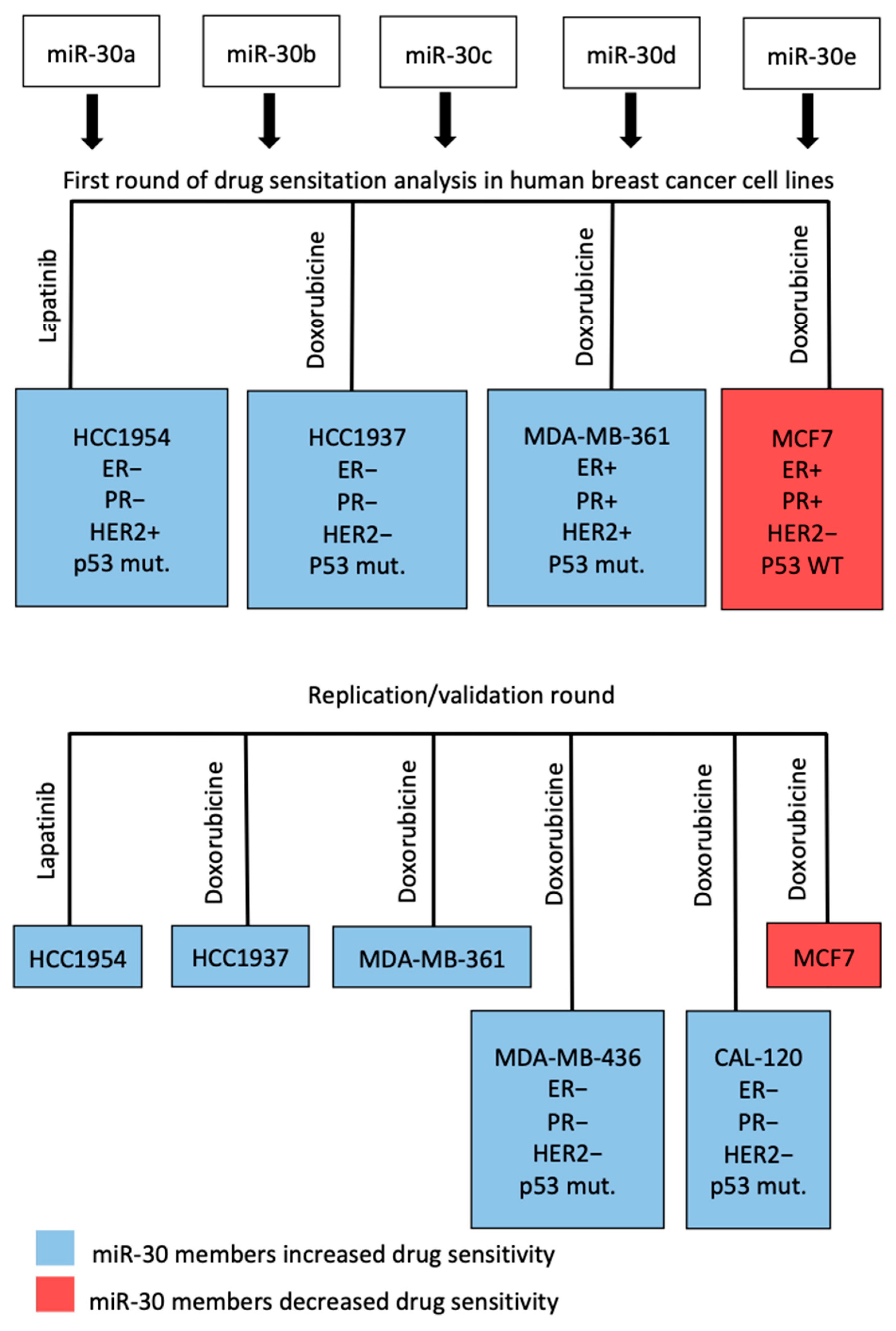
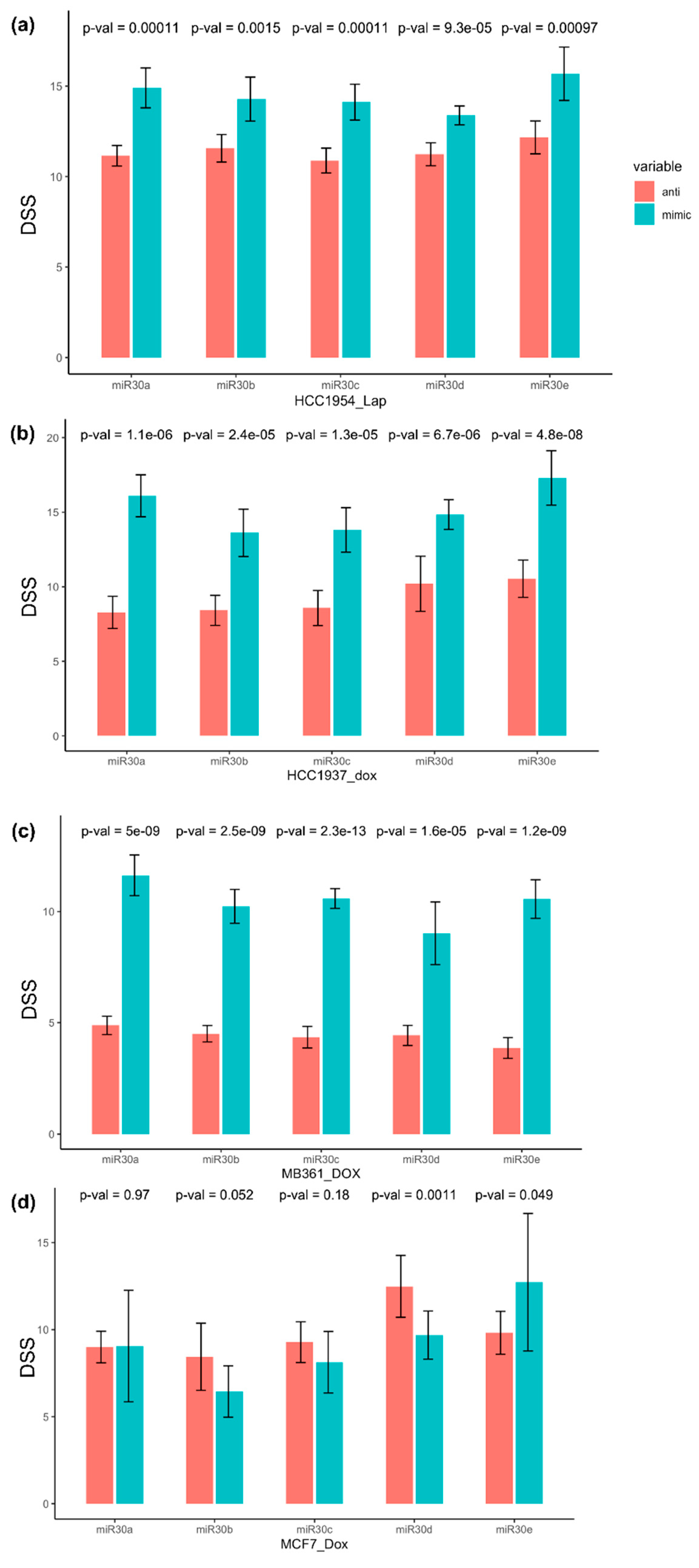
3.3. miR-30 Family Members Sensitized Breast Cancer Cell Lines to Doxorubicin and Lapatinib
3.4. The Putative Targets of the miR-30 Family Members in Breast Cancers Are Associated with Cellular Growth, Proliferation, and Motility
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Calin, G.A.; Sevignani, C.; Dumitru, C.D.; Hyslop, T.; Noch, E.; Yendamuri, S.; Shimizu, M.; Rattan, S.; Bullrich, F.; Negrini, M.; et al. Human microRNA genes are frequently located at fragile sites and genomic regions involved in cancers. Proc. Natl. Acad. Sci. USA 2004, 101, 2999–3004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quesne, J.L.; Jones, J.; Warren, J.; Dawson, S.J.; Ali, H.R.; Bardwell, H.; Blows, F.; Pharoah, P.; Caldas, C. Biological and prognostic associations of miR-205 and let-7b in breast cancer revealed by in situ hybridization analysis of micro-RNA expression in arrays of archival tumour tissue. J. Pathol. 2012, 227, 306–314. [Google Scholar] [CrossRef]
- Jonas, S.; Izaurralde, E. Towards a molecular understanding of microRNA-mediated gene silencing. Nat. Rev. Genet. 2015, 16, 421–433. [Google Scholar] [CrossRef]
- Ameres, S.L.; Zamore, P.D. Diversifying microRNA sequence and function. Nat. Rev. Mol. Cell Biol. 2013, 14, 475–488. [Google Scholar] [CrossRef] [PubMed]
- Baek, D.; Villen, J.; Shin, C.; Camargo, F.D.; Gygi, S.P.; Bartel, D.P. The impact of microRNAs on protein output. Nature 2008, 455, 64–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, W.; Liu, B.; Qu, S.; Liang, G.; Luo, W.; Gong, C. MicroRNAs and cancer: Key paradigms in molecular therapy. Oncol. Lett. 2018, 15, 2735–2742. [Google Scholar] [CrossRef] [Green Version]
- Kurozumi, S.; Yamaguchi, Y.; Kurosumi, M.; Ohira, M.; Matsumoto, H.; Horiguchi, J. Recent trends in microRNA research into breast cancer with particular focus on the associations between microRNAs and intrinsic subtypes. J. Hum. Genet. 2017, 62, 15–24. [Google Scholar] [CrossRef] [Green Version]
- Mertens-Talcott, S.U.; Chintharlapalli, S.; Li, X.; Safe, S. The oncogenic microRNA-27a targets genes that regulate specificity protein transcription factors and the G2-M checkpoint in MDA-MB-231 breast cancer cells. Cancer Res. 2007, 67, 11001–11011. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.; Zhang, H.W.; Lu, M.H.; He, X.H.; Li, Y.; Gu, H.; Liu, M.F.; Wang, E.D. MicroRNA-155 functions as an OncomiR in breast cancer by targeting the suppressor of cytokine signaling 1 gene. Cancer Res. 2010, 70, 3119–3127. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Kaur, S.; Greshock, J.; Lassus, H.; Zhong, X.; Wang, Y.; Leminen, A.; Shao, Z.; Hu, X.; Liang, S.; et al. A combined array-based comparative genomic hybridization and functional library screening approach identifies mir-30d as an oncomir in cancer. Cancer Res. 2012, 72, 154–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feliciano, A.; Castellvi, J.; Artero-Castro, A.; Leal, J.A.; Romagosa, C.; Hernandez-Losa, J.; Peg, V.; Fabra, A.; Vidal, F.; Kondoh, H.; et al. miR-125b acts as a tumor suppressor in breast tumorigenesis via its novel direct targets ENPEP, CK2-alpha, CCNJ, and MEGF9. PLoS ONE 2013, 8, e76247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fong, M.Y.; Zhou, W.; Liu, L.; Alontaga, A.Y.; Chandra, M.; Ashby, J.; Chow, A.; O’Connor, S.T.; Li, S.; Chin, A.R.; et al. Breast-cancer-secreted miR-122 reprograms glucose metabolism in premetastatic niche to promote metastasis. Nat. Cell Biol. 2015, 17, 183–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tavazoie, S.F.; Alarcon, C.; Oskarsson, T.; Padua, D.; Wang, Q.; Bos, P.D.; Gerald, W.L.; Massague, J. Endogenous human microRNAs that suppress breast cancer metastasis. Nature 2008, 451, 147–152. [Google Scholar] [CrossRef] [Green Version]
- Kozomara, A.; Griffiths-Jones, S. miRBase: Annotating high confidence microRNAs using deep sequencing data. Nucleic Acids Res. 2014, 42, D68–D73. [Google Scholar] [CrossRef] [Green Version]
- Syrjakoski, K.; Vahteristo, P.; Eerola, H.; Tamminen, A.; Kivinummi, K.; Sarantaus, L.; Holli, K.; Blomqvist, C.; Kallioniemi, O.P.; Kainu, T.; et al. Population-based study of BRCA1 and BRCA2 mutations in 1035 unselected Finnish breast cancer patients. J. Natl. Cancer Inst. 2000, 92, 1529–1531. [Google Scholar] [CrossRef] [Green Version]
- Kilpivaara, O.; Bartkova, J.; Eerola, H.; Syrjakoski, K.; Vahteristo, P.; Lukas, J.; Blomqvist, C.; Holli, K.; Heikkila, P.; Sauter, G.; et al. Correlation of CHEK2 protein expression and c.1100delC mutation status with tumor characteristics among unselected breast cancer patients. Int. J. Cancer 2005, 113, 575–580. [Google Scholar] [CrossRef]
- Tommiska, J.; Eerola, H.; Heinonen, M.; Salonen, L.; Kaare, M.; Tallila, J.; Ristimaki, A.; von Smitten, K.; Aittomaki, K.; Heikkila, P.; et al. Breast cancer patients with p53 Pro72 homozygous genotype have a poorer survival. Clin. Cancer Res. 2005, 11, 5098–5103. [Google Scholar] [CrossRef] [Green Version]
- Vehmanen, P.; Friedman, L.S.; Eerola, H.; McClure, M.; Ward, B.; Sarantaus, L.; Kainu, T.; Syrjakoski, K.; Pyrhonen, S.; Kallioniemi, O.P.; et al. Low proportion of BRCA1 and BRCA2 mutations in Finnish breast cancer families: Evidence for additional susceptibility genes. Hum. Mol. Genet. 1997, 6, 2309–2315. [Google Scholar] [CrossRef] [Green Version]
- Vahteristo, P.; Eerola, H.; Tamminen, A.; Blomqvist, C.; Nevanlinna, H. A probability model for predicting BRCA1 and BRCA2 mutations in breast and breast-ovarian cancer families. Br. J. Cancer 2001, 84, 704–708. [Google Scholar] [CrossRef] [Green Version]
- Vahteristo, P.; Bartkova, J.; Eerola, H.; Syrjakoski, K.; Ojala, S.; Kilpivaara, O.; Tamminen, A.; Kononen, J.; Aittomaki, K.; Heikkila, P.; et al. A CHEK2 genetic variant contributing to a substantial fraction of familial breast cancer. Am. J. Hum. Genet. 2002, 71, 432–438. [Google Scholar] [CrossRef] [Green Version]
- Elston, C.W.; Ellis, I.O. Pathological prognostic factors in breast cancer. I. The value of histological grade in breast cancer: Experience from a large study with long-term follow-up. Histopathology 1991, 19, 403–410. [Google Scholar] [CrossRef]
- Tanner, M.; Gancberg, D.; Di Leo, A.; Larsimont, D.; Rouas, G.; Piccart, M.J.; Isola, J. Chromogenic in situ hybridization: A practical alternative for fluorescence in situ hybridization to detect HER-2/neu oncogene amplification in archival breast cancer samples. Am. J. Pathol. 2000, 157, 1467–1472. [Google Scholar] [CrossRef]
- Ahlin, C.; Aaltonen, K.; Amini, R.M.; Nevanlinna, H.; Fjallskog, M.L.; Blomqvist, C. Ki67 and cyclin A as prognostic factors in early breast cancer. What are the optimal cut-off values? Histopathology 2007, 51, 491–498. [Google Scholar] [CrossRef] [PubMed]
- Dvinge, H.; Git, A.; Graf, S.; Salmon-Divon, M.; Curtis, C.; Sottoriva, A.; Zhao, Y.; Hirst, M.; Armisen, J.; Miska, E.A.; et al. The shaping and functional consequences of the microRNA landscape in breast cancer. Nature 2013, 497, 378–382. [Google Scholar] [CrossRef] [PubMed]
- Curtis, C.; Shah, S.P.; Chin, S.F.; Turashvili, G.; Rueda, O.M.; Dunning, M.J.; Speed, D.; Lynch, A.G.; Samarajiwa, S.; Yuan, Y.; et al. The genomic and transcriptomic architecture of 2,000 breast tumours reveals novel subgroups. Nature 2012, 486, 346–352. [Google Scholar] [CrossRef]
- Yadav, B.; Pemovska, T.; Szwajda, A.; Kulesskiy, E.; Kontro, M.; Karjalainen, R.; Majumder, M.M.; Malani, D.; Murumagi, A.; Knowles, J.; et al. Quantitative scoring of differential drug sensitivity for individually optimized anticancer therapies. Sci. Rep. 2014, 4, 5193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potdar, S.; Ianevski, A.; Mpindi, J.P.; Bychkov, D.; Fiere, C.; Ianevski, P.; Yadav, B.; Wennerberg, K.; Aittokallio, T.; Kallioniemi, O.; et al. Breeze: An integrated quality control and data analysis application for high-throughput drug screening. Bioinformatics 2020, 36, 3602–3604. [Google Scholar] [CrossRef] [PubMed]
- Fagerholm, R.; Khan, S.; Schmidt, M.K.; Garcia-Closas, M.; Heikkila, P.; Saarela, J.; Beesley, J.; Jamshidi, M.; Aittomaki, K.; Liu, J.; et al. TP53-based interaction analysis identifies cis-eQTL variants for TP53BP2, FBXO28, and FAM53A that associate with survival and treatment outcome in breast cancer. Oncotarget 2017, 8, 18381–18398. [Google Scholar] [CrossRef]
- Konecny, G.E.; Pauletti, G.; Untch, M.; Wang, H.J.; Mobus, V.; Kuhn, W.; Thomssen, C.; Harbeck, N.; Wang, L.; Apple, S.; et al. Association between HER2, TOP2A, and response to anthracycline-based preoperative chemotherapy in high-risk primary breast cancer. Breast Cancer Res. Treat. 2010, 120, 481–489. [Google Scholar] [CrossRef]
- Jarvinen, T.A.; Tanner, M.; Barlund, M.; Borg, A.; Isola, J. Characterization of topoisomerase II alpha gene amplification and deletion in breast cancer. Genes Chromosomes Cancer 1999, 26, 142–150. [Google Scholar] [CrossRef]
- Fang, Y.; Shen, H.; Cao, Y.; Li, H.; Qin, R.; Chen, Q.; Long, L.; Zhu, X.L.; Xie, C.J.; Xu, W.L. Involvement of miR-30c in resistance to doxorubicin by regulating YWHAZ in breast cancer cells. Braz. J. Med. Biol. Res. 2014, 47, 60–69. [Google Scholar] [CrossRef] [Green Version]
- Bockhorn, J.; Dalton, R.; Nwachukwu, C.; Huang, S.; Prat, A.; Yee, K.; Chang, Y.F.; Huo, D.; Wen, Y.; Swanson, K.E.; et al. MicroRNA-30c inhibits human breast tumour chemotherapy resistance by regulating TWF1 and IL-11. Nat. Commun. 2013, 4, 1393. [Google Scholar] [CrossRef] [Green Version]
- Sestito, R.; Cianfrocca, R.; Rosano, L.; Tocci, P.; Semprucci, E.; Di Castro, V.; Caprara, V.; Ferrandina, G.; Sacconi, A.; Blandino, G.; et al. miR-30a inhibits endothelin A receptor and chemoresistance in ovarian carcinoma. Oncotarget 2016, 7, 4009–4023. [Google Scholar] [CrossRef]
- Guo, F.; Chen, H.; Chang, J.; Zhang, L. Mutation R273H confers p53 a stimulating effect on the IGF-1R-AKT pathway via miR-30a suppression in breast cancer. Biomed. Pharmacother. 2016, 78, 335–341. [Google Scholar] [CrossRef] [PubMed]
- di Gennaro, A.; Damiano, V.; Brisotto, G.; Armellin, M.; Perin, T.; Zucchetto, A.; Guardascione, M.; Spaink, H.P.; Doglioni, C.; Snaar-Jagalska, B.E.; et al. A p53/miR-30a/ZEB2 axis controls triple negative breast cancer aggressiveness. Cell Death Differ. 2018, 25, 2165–2180. [Google Scholar] [CrossRef] [PubMed]
- di Gennaro, A.; Damiano, V.; Brisotto, G.; Armellin, M.; Perin, T.; Zucchetto, A.; Guardascione, M.; Spaink, H.P.; Doglioni, C.; Snaar-Jagalska, B.E.; et al. Correction to: A p53/miR-30a/ZEB2 axis controls triple negative breast cancer aggressiveness. Cell Death Differ. 2019, 26, 2493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, C.W.; Wang, H.W.; Chang, C.W.; Chu, H.W.; Chen, C.Y.; Yu, J.C.; Chao, J.I.; Liu, H.F.; Ding, S.L.; Shen, C.Y. MicroRNA-30a inhibits cell migration and invasion by downregulating vimentin expression and is a potential prognostic marker in breast cancer. Breast Cancer Res. Treat. 2012, 134, 1081–1093. [Google Scholar] [CrossRef]
- Chang, C.W.; Yu, J.C.; Hsieh, Y.H.; Yao, C.C.; Chao, J.I.; Chen, P.M.; Hsieh, H.Y.; Hsiung, C.N.; Chu, H.W.; Shen, C.Y.; et al. MicroRNA-30a increases tight junction protein expression to suppress the epithelial-mesenchymal transition and metastasis by targeting Slug in breast cancer. Oncotarget 2016, 7, 16462–16478. [Google Scholar] [CrossRef] [Green Version]
- Kumarswamy, R.; Mudduluru, G.; Ceppi, P.; Muppala, S.; Kozlowski, M.; Niklinski, J.; Papotti, M.; Allgayer, H. MicroRNA-30a inhibits epithelial-to-mesenchymal transition by targeting Snai1 and is downregulated in non-small cell lung cancer. Int. J. Cancer 2012, 130, 2044–2053. [Google Scholar] [CrossRef]
- Zhou, Q.; Yang, M.; Lan, H.; Yu, X. miR-30a negatively regulates TGF-beta1-induced epithelial-mesenchymal transition and peritoneal fibrosis by targeting Snai1. Am. J. Pathol. 2013, 183, 808–819. [Google Scholar] [CrossRef]
- Zhang, N.; Wang, X.; Huo, Q.; Sun, M.; Cai, C.; Liu, Z.; Hu, G.; Yang, Q. MicroRNA-30a suppresses breast tumor growth and metastasis by targeting metadherin. Oncogene 2014, 33, 3119–3128. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.; Settleman, J. EMT, cancer stem cells and drug resistance: An emerging axis of evil in the war on cancer. Oncogene 2010, 29, 4741–4751. [Google Scholar] [CrossRef] [Green Version]
- van Staalduinen, J.; Baker, D.; Ten Dijke, P.; van Dam, H. Epithelial-mesenchymal-transition-inducing transcription factors: New targets for tackling chemoresistance in cancer? Oncogene 2018, 37, 6195–6211. [Google Scholar] [CrossRef]
- Kawaguchi, T.; Yan, L.; Qi, Q.; Peng, X.; Gabriel, E.M.; Young, J.; Liu, S.; Takabe, K. Overexpression of suppressive microRNAs, miR-30a and miR-200c are associated with improved survival of breast cancer patients. Sci. Rep. 2017, 7, 15945. [Google Scholar] [CrossRef] [Green Version]
- Peurala, H.; Greco, D.; Heikkinen, T.; Kaur, S.; Bartkova, J.; Jamshidi, M.; Aittomaki, K.; Heikkila, P.; Bartek, J.; Blomqvist, C.; et al. MiR-34a expression has an effect for lower risk of metastasis and associates with expression patterns predicting clinical outcome in breast cancer. PLoS ONE 2011, 6, e26122. [Google Scholar] [CrossRef]
- Gaziel-Sovran, A.; Segura, M.F.; Di Micco, R.; Collins, M.K.; Hanniford, D.; Vega-Saenz de Miera, E.; Rakus, J.F.; Dankert, J.F.; Shang, S.; Kerbel, R.S.; et al. miR-30b/30d regulation of GalNAc transferases enhances invasion and immunosuppression during metastasis. Cancer Cell 2011, 20, 104–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, J.; Liang, L.; Huang, S.; Ding, J.; Tan, N.; Zhao, Y.; Yan, M.; Ge, C.; Zhang, Z.; Chen, T.; et al. MicroRNA-30d promotes tumor invasion and metastasis by targeting Galphai2 in hepatocellular carcinoma. Hepatology 2010, 51, 846–856. [Google Scholar] [CrossRef] [PubMed]
- Mayya, V.K.; Duchaine, T.F. On the availability of microRNA-induced silencing complexes, saturation of microRNA-binding sites and stoichiometry. Nucleic Acids Res. 2015, 43, 7556–7565. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | miR-30d n (%) | p | ||
|---|---|---|---|---|
| Total | Low 1 | High 1 | ||
| Tumor histology | ||||
| Ductal | 838 (70.2%) | 233 (64.5%) | 605 (72.7%) | 0.003 2 |
| Lobular | 222 (18.6%) | 90 (24.9%) | 132 (15.9%) | |
| Medullary | 15 (1.3%) | 3 (0.8%) | 12 (1.4%) | |
| Other | 118 (9.9%) | 35 (9.7%) | 83 (10.0%) | |
| Grade | ||||
| 1 | 281 (23.9%) | 96 (27.0%) | 185 (22.5%) | 0.0002 |
| 2 | 547 (46.5%) | 183 (51.5%) | 364 (44.3%) | |
| 3 | 349 (29.7%) | 76 (21.4%) | 273 (33.2%) | |
| Tumor size | ||||
| 1 | 694 (58.9%) | 213 (60.2%) | 481 (58.3%) | 0.417 |
| 2 | 410 (34.8%) | 115 (32.5%) | 295 (35.8%) | |
| 3 | 40 (3.4%) | 16 (4.5%) | 24 (2.9%) | |
| 4 | 35 (3.0%) | 10 (2.8%) | 25 (3.0%) | |
| Nodal status 3 | ||||
| Negative | 645 (54.9%) | 214 (61.0%) | 431 (52.4%) | 0.007 |
| positive | 529 (45.1%) | 137 (39.0%) | 392 (47.6%) | |
| M 4 | ||||
| Negative | 1147 (97.0%) | 347 (96.9%) | 800 (97.0%) | 1.000 |
| Positive | 36 (3.0%) | 11 (3.1%) | 25 (3.0%) | |
| ER status | ||||
| Negative | 234 (20.6%) | 70 (21.0%) | 164 (20.5%) | 0.872 |
| Positive | 900 (79.4%) | 263 (79.0%) | 637 (79.5%) | |
| PR status | ||||
| Negative | 379 (33.5%) | 124 (37.0%) | 255 (32.0%) | 0.113 |
| Positive | 753 (66.5%) | 211 (63.0%) | 542 (68.0%) | |
| p53 tumor status | ||||
| Negative | 899 (80.1%) | 264 (82.8%) | 635 (79.0%) | 0.160 |
| Positive | 224 (19.9%) | 55 (17.2%) | 169 (21.0%) | |
| HER2 status | ||||
| Negative | 984 (87.1%) | 298 (90.0%) | 686 (85.9%) | 0.064 |
| Positive | 146 (12.9%) | 33 (10.0%) | 113 (14.1%) | |
| Ki-67 (proliferation marker) | ||||
| Negative | 786 (67.2%) | 250 (72.5%) | 536 (65.0%) | 0.014 |
| Positive | 383 (32.8%) | 95 (27.5%) | 288 (35.0%) | |
| ER/PR/HER2 status 5 | ||||
| ER/PR+ and HER2− | 778 (72.6%) | 232 (76.1%) | 546 (71.2%) | 0.032 |
| ER/PR+ and HER2+ | 91 (8.5%) | 14 (4.6%) | 77 (10.0%) | |
| ER/PR− and HER2+ | 52 (4.9%) | 17 (5.6%) | 35 (4.6%) | |
| ER− and PR− and HER2− | 151 (14.1%) | 42 (13.8%) | 109 (14.2%) | |
| Covariate | 5-Year BDDM | 10-Year BCS | ||||
|---|---|---|---|---|---|---|
| p | HR | 95%CI | p | HR | 95%CI | |
| Grade | 0.0001 | 2.28 | 1.45–3.49 | 0.0004 | 2.06 | 1.37–3.09 |
| T | 1.7 × 10−6 | 1.58 | 1.24–2.01 | 0.001 | 1.50 | 1.17–1.93 |
| N | 1.1 × 10−6 | 2.95 | 1.77–4.91 | 1.9 × 10−5 | 3.11 | 1.84–5.23 |
| M | - | - | - | 5.8 × 10−8 | 6.31 | 3.24–12.2 |
| ER | 0.60 | 1.16 | 0.66–2.045 | 0.50 | 0.82 | 0.46–1.45 |
| Ki67 | 0.97 | 0.97 | 0.57–1.71 | 0.70 | 0.90 | 0.55–1.48 |
| HER2 | 0.38 | 1.62 | 0.95–2.78 | 0.17 | 1.48 | 0.85–2.57 |
| p53 | 0.44 | 1.32 | 0.77–2.25 | 0.23 | 1.37 | 0.81–2.32 |
| miR-30d | 0.0004 | 0.44 | 0.28–0.69 | 0.006 | 0.54 | 0.35–0.84 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jamshidi, M.; Fagerholm, R.; Muranen, T.A.; Kaur, S.; Potdar, S.; Khan, S.; Netti, E.; Mpindi, J.-P.; Yadav, B.; Kiiski, J.I.; et al. High miR-30 Expression Associates with Improved Breast Cancer Patient Survival and Treatment Outcome. Cancers 2021, 13, 2907. https://doi.org/10.3390/cancers13122907
Jamshidi M, Fagerholm R, Muranen TA, Kaur S, Potdar S, Khan S, Netti E, Mpindi J-P, Yadav B, Kiiski JI, et al. High miR-30 Expression Associates with Improved Breast Cancer Patient Survival and Treatment Outcome. Cancers. 2021; 13(12):2907. https://doi.org/10.3390/cancers13122907
Chicago/Turabian StyleJamshidi, Maral, Rainer Fagerholm, Taru A. Muranen, Sippy Kaur, Swapnil Potdar, Sofia Khan, Eliisa Netti, John-Patrick Mpindi, Bhagwan Yadav, Johanna I. Kiiski, and et al. 2021. "High miR-30 Expression Associates with Improved Breast Cancer Patient Survival and Treatment Outcome" Cancers 13, no. 12: 2907. https://doi.org/10.3390/cancers13122907