Cisplatin Decreases ENaC Activity Contributing to Renal Salt Wasting Syndrome



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
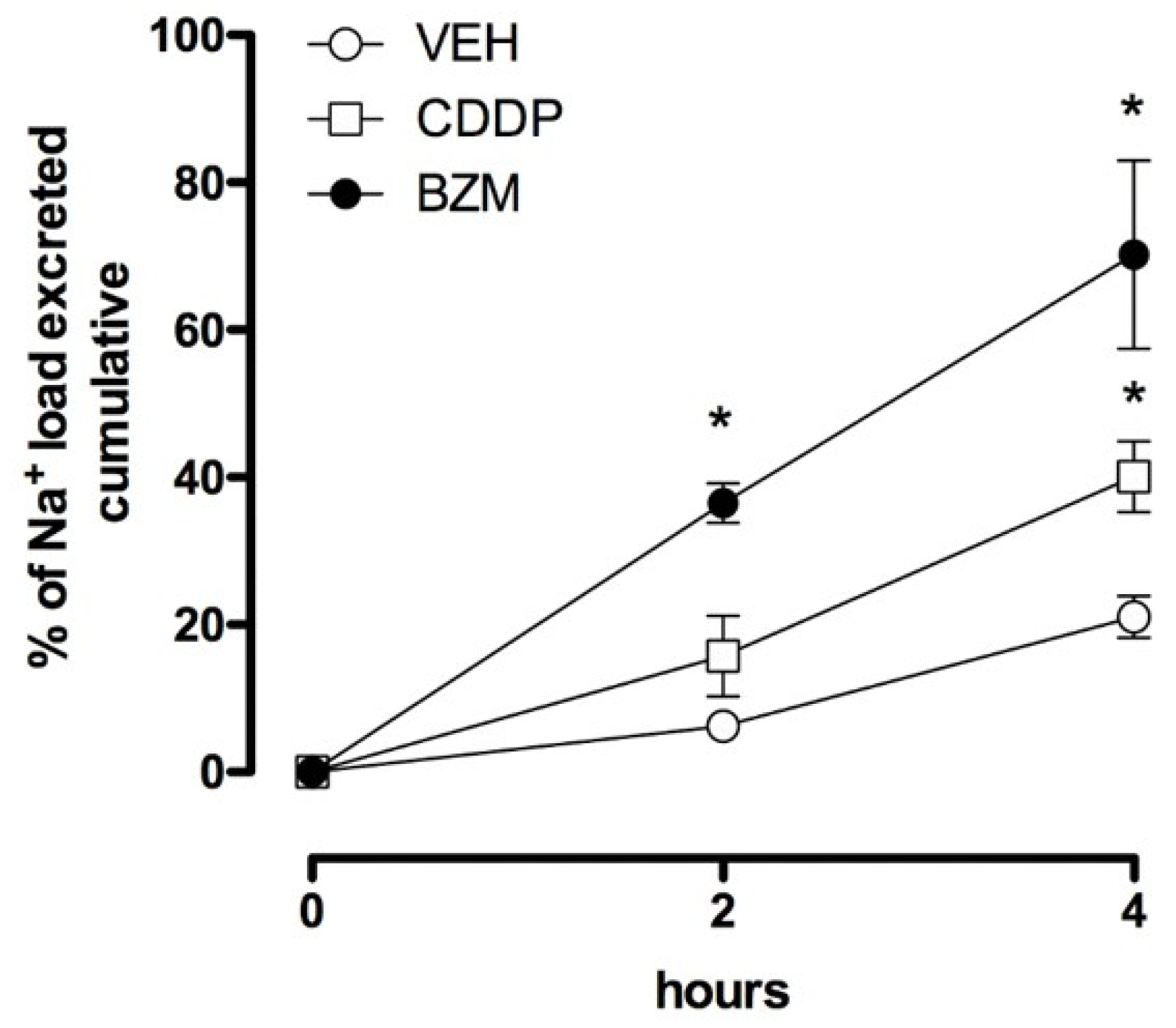
2.1. CDDP Increases Urinary Sodium Excretion
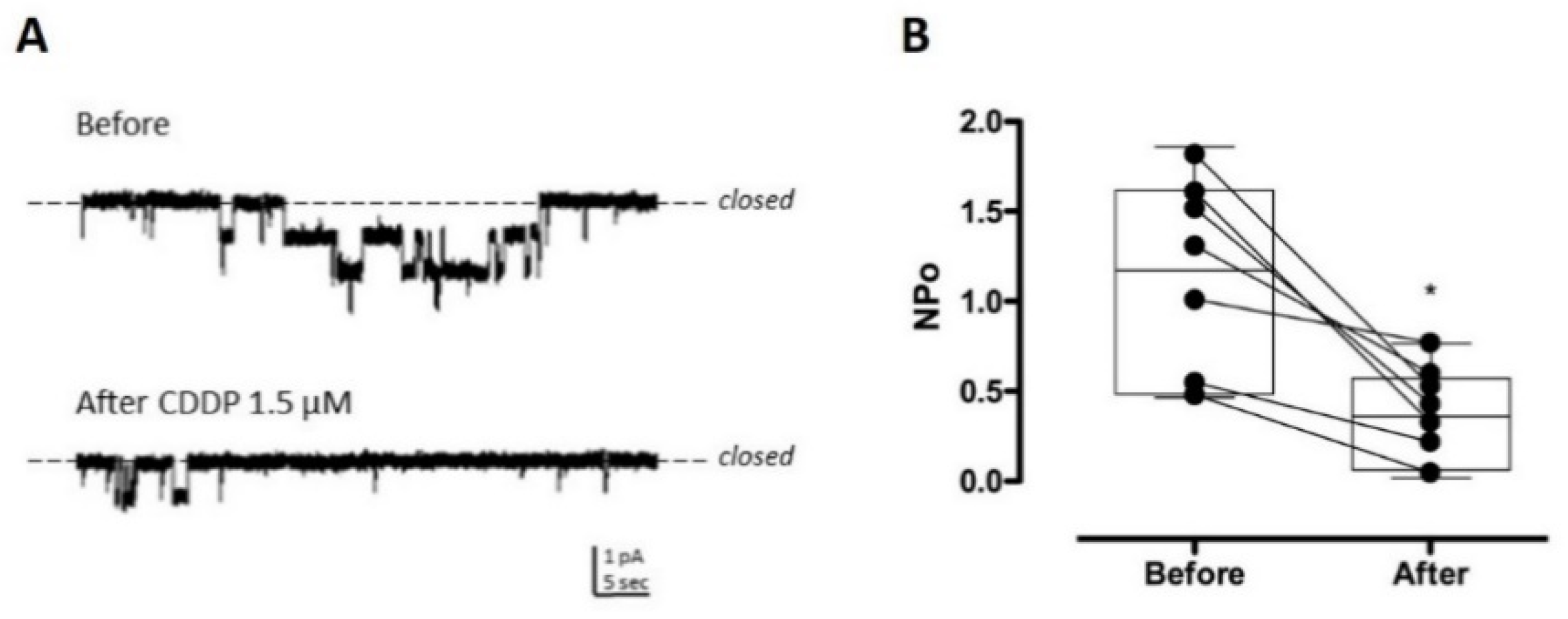
2.2. CDDP Reduces ENaC Activity in Split-Open Tubules
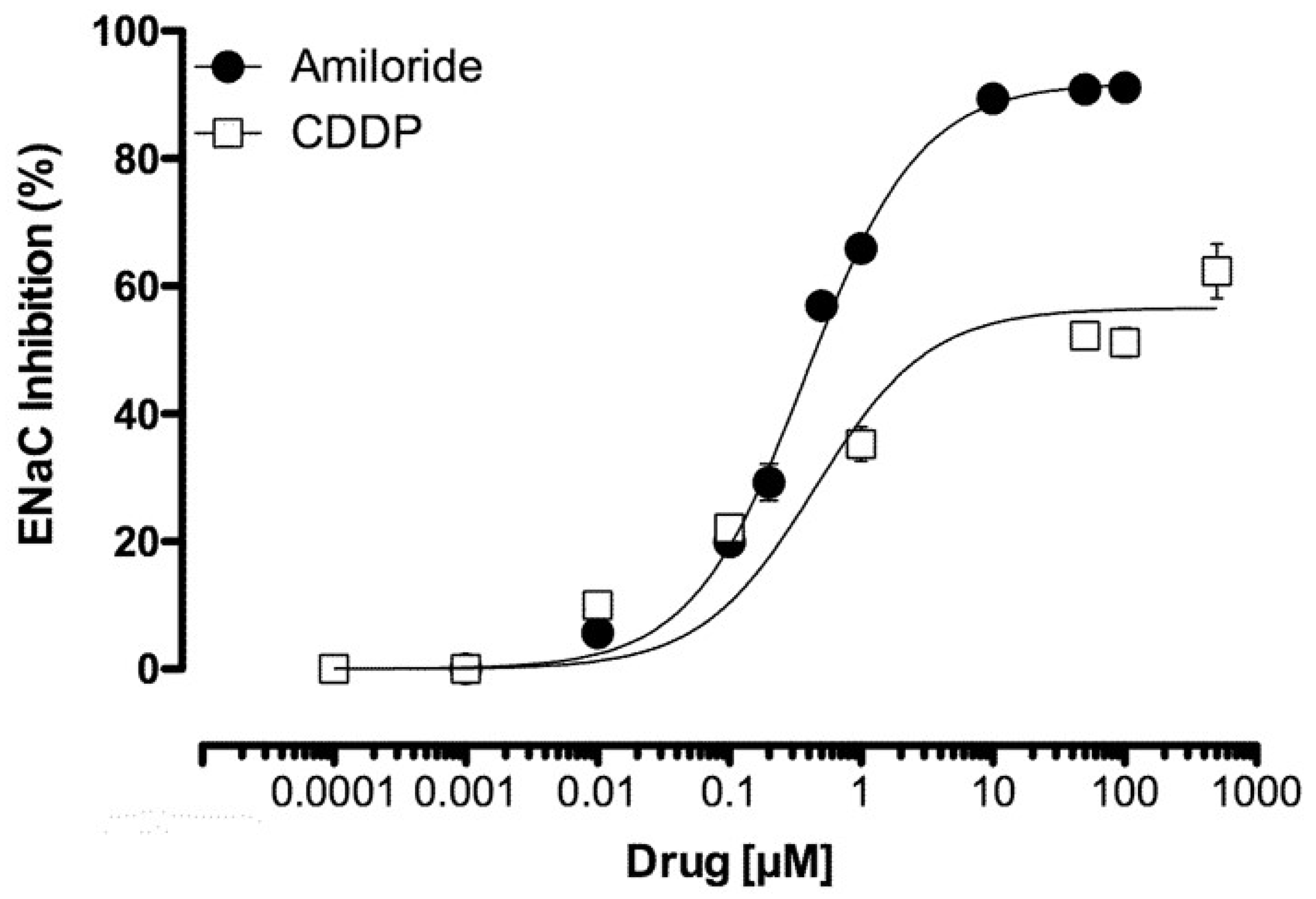
2.3. CDDP Is a Partial Inhibitor of ENaC
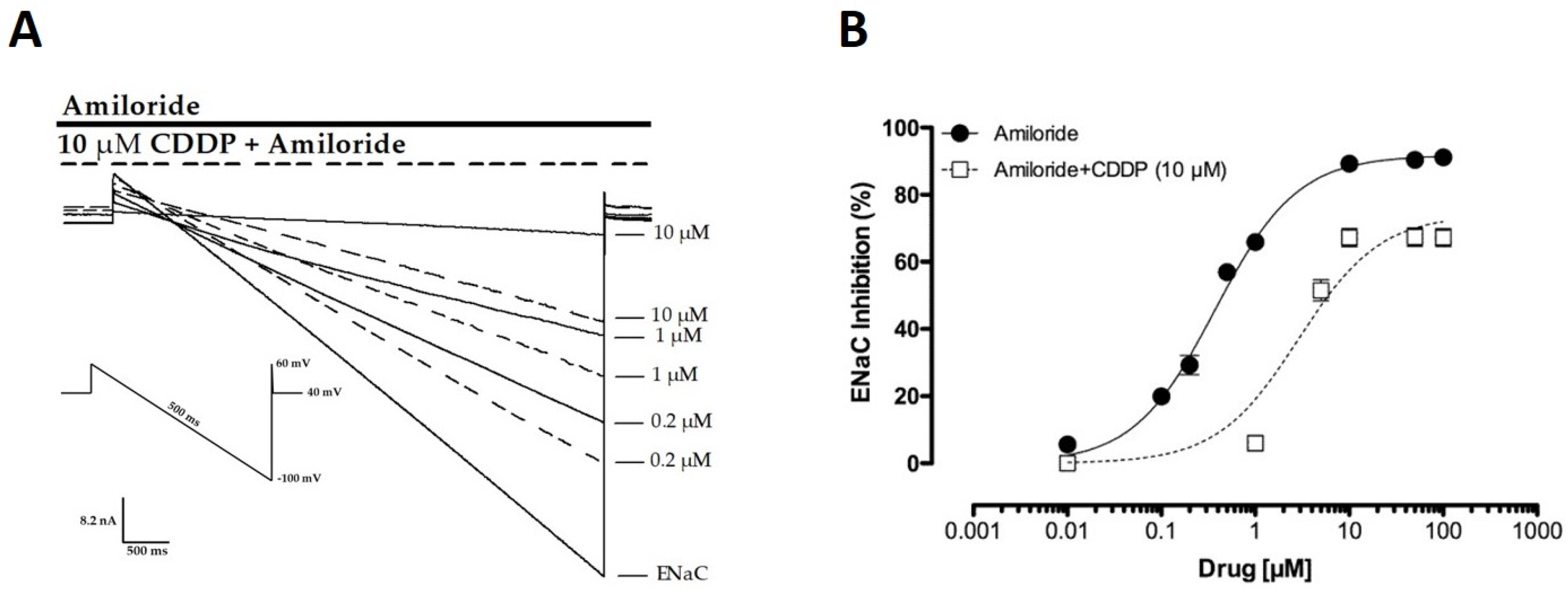
2.4. CDDP Competes with Amiloride for Inhibition of ENaC
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Metabolic Cage Experiments
4.3. Split-Open Tubule Preparation and Single-Channel Patch Clamp Electrophysiology
4.4. ENaC Expression in CHO Cells and Whole-Cell Patch Clamp Electrophysiology
4.5. Statistics
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Chu, G. Cellular responses to cisplatin. The roles of DNA-binding proteins and DNA repair. J. Biol. Chem. 1994, 269, 787–790. [Google Scholar] [PubMed]
- Siddik, Z.H. Cisplatin: Mode of cytotoxic action and molecular basis of resistance. Oncogene 2003, 22, 7265–7279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dasari, S.; Tchounwou, P.B. Cisplatin in cancer therapy: Molecular mechanisms of action. Eur. J. Pharmacol. 2014, 740, 364–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pabla, N.; Dong, Z. Cisplatin nephrotoxicity: Mechanisms and renoprotective strategies. Kidney Int. 2008, 73, 994–1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciarimboli, G.; Ludwig, T.; Lang, D.; Pavenstädt, H.; Koepsell, H.; Piechota, H.-J.; Haier, J.; Jaehde, U.; Zisowsky, J.; Schlatter, E. Cisplatin Nephrotoxicity Is Critically Mediated via the Human Organic Cation Transporter 2. Am. J. Pathol. 2005, 167, 1477–1484. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, T.; Yonezawa, A.; Hashimoto, S.; Katsura, T.; Inui, K.-I. Disruption of multidrug and toxin extrusion MATE1 potentiates cisplatin-induced nephrotoxicity. Biochem. Pharmacol. 2010, 80, 1762–1767. [Google Scholar] [CrossRef] [Green Version]
- Huličiak, M.; Vacek, J.; Sebela, M.; Orolinova, E.; Znaleziona, J.; Havlikova, M.; Kubala, M. Covalent binding of cisplatin impairs the function of Na(+)/K(+)-ATPase by binding to its cytoplasmic part. Biochem. Pharmacol. 2012, 83, 1507–1513. [Google Scholar] [CrossRef]
- Šeflová, J.; Cechova, P.; Stenclova, T.; Sebela, M.; Kubala, M. Identification of cisplatin-binding sites on the large cytoplasmic loop of the Na(+)/K(+)-ATPase. J. Enzym. Inhib. Med. Chem. 2018, 33, 701–706. [Google Scholar] [CrossRef] [Green Version]
- Kishore, B.K.; Krane, C.M.; Di Iulio, D.; Menon, A.G.; Cacini, W. Expression of renal aquaporins 1, 2, and 3 in a rat model of cisplatin-induced polyuria. Kidney Int. 2000, 58, 701–711. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.W.; Lee, J.U.; Nah, M.Y.; Kang, D.G.; Ahn, K.Y.; Lee, H.S.; Choi, K.C. Cisplatin decreases the abundance of aquaporin water channels in rat kidney. J. Am. Soc. Nephrol. 2001, 12, 875–882. [Google Scholar]
- Hanukoglu, I.; Hanukoglu, A. Epithelial sodium channel (ENaC) family: Phylogeny, structure–function, tissue distribution, and associated inherited diseases. Gene 2016, 579, 95–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roush, G.C.; Sica, M.A. Diuretics for Hypertension: A Review and Update. Am. J. Hypertens. 2016, 29, 1130–1137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexander, R.T.; Dimke, H. Effect of diuretics on renal tubular transport of calcium and magnesium. Am. J. Physiol. Physiol. 2017, 312, F998–F1015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunt, T.; Atherton-Watson, H.C.; Axford, J.; Collingwood, S.P.; Coote, K.J.; Cox, B.; Czarnecki, S.; Danahay, H.; Devereux, N.; Howsham, C.; et al. Discovery of a novel chemotype of potent human ENaC blockers using a bioisostere approach. Part 1: Quaternary amines. Bioorganic Med. Chem. Lett. 2012, 22, 929–932. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Eaton, D.C.; Helms, M.N. Effect of divalent heavy metals on epithelial Na+ channels in A6 cells. Am. J. Physiol. Physiol. 2007, 293, F236–F244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamdi, T.; Latta, S.; Jallad, B.; Kheir, F.; Alhosaini, M.N.; Patel, A. Cisplatin-Induced Renal Salt Wasting Syndrome. South. Med. J. 2010, 103, 793–799. [Google Scholar] [CrossRef]
- Inamori, M.; Inohara, H.; Horii, A. Differential diagnosis of hyponatremia induced by cisplatin-containing chemotherapy: Syndrome of inappropriate secretion of antidiuretic hormone (SIADH) or renal salt wasting syndrome (RSWS). Acta Oto Laryngol. Case Rep. 2016, 1, 33–35. [Google Scholar] [CrossRef]
- Pham, P.C.T.; Reddy, P.; Qaqish, S.; Kamath, A.; Rodriguez, J.; Bolos, D.; Zalom, M.; Pham, P.-T. Cisplatin-Induced Renal Salt Wasting Requiring over 12 Liters of 3% Saline Replacement. Case Rep. Nephrol. 2017, 2017, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Iida, T.; Makino, Y.; Okamoto, K.; Yoshikawa, N.; Makino, I.; Nakamura, T.; Tanaka, H. Functional modulation of the mineralocorticoid receptor by cis-diamminedichloroplatinum (II). Kidney Int. 2000, 58, 1450–1460. [Google Scholar] [CrossRef] [Green Version]
- Vardanyan, R.; Hruby, V. Synthesis of Essential Drugs. Synth. Essent. Drugs 2006. [Google Scholar] [CrossRef]
- Szturz, P.; Wouters, K.; Kiyota, N.; Tahara, M.; Prabhash, K.; Noronha, V.; Adelstein, D.; Van Gestel, D.; Vermorken, J.B. Low-Dose vs. High-Dose Cisplatin: Lessons Learned From 59 Chemoradiotherapy Trials in Head and Neck Cancer. Front. Oncol. 2019, 9, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakakibara, N.; Suzuki, K.; Kaneta, H.; Yoshimura, Y.; Deyama, Y.; Matsumoto, A.; Fukuda, H. Inhibition of Na+,K+-ATPase by cisplatin and its recovery by 2-mercaptoethanol in human squamous cell carcinoma cells. Anti Cancer Drugs 1999, 10, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Mironova, E.; Chen, Y.; Pao, A.C.; Roos, K.P.; Kohan, N.E.; Bugaj, V.; Stockand, J.D. Activation of ENaC by AVP contributes to the urinary concentrating mechanism and dilution of plasma. Am. J. Physiol. 2015, 308, F237–F243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mironova, E.; Bugay, V.; Pochynyuk, O.; Staruschenko, A.; Stockand, J.D. Recording Ion Channels in Isolated, Split-Opened Tubules. Methods Mol. Biol. 2013, 998, 341–353. [Google Scholar] [PubMed] [Green Version]
- Gamper, N.; Stockand, J.D.; Shapiro, M.S. Subunit-Specific Modulation of KCNQ Potassium Channels by Src Tyrosine Kinase. J. Neurosci. 2003, 23, 84–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staruschenko, A.; Adams, E.; Booth, R.E.; Stockand, J.D. Epithelial Na+ Channel Subunit Stoichiometry. Biophys. J. 2005, 88, 3966–3975. [Google Scholar] [CrossRef] [Green Version]
- Staruschenko, A.; Nichols, A.; Medina, J.L.; Camacho, P.; Zheleznova, N.N.; Stockand, J.D. Rho Small GTPases Activate the Epithelial Na+ Channel. J. Biol. Chem. 2004, 279, 49989–49994. [Google Scholar] [CrossRef] [Green Version]
- Pochynyuk, O.; Staruschenko, A.; Tong, Q.; Medina, J.; Stockand, J.D. Identification of a Functional Phosphatidylinositol 3,4,5-Trisphosphate Binding Site in the Epithelial Na+Channel. J. Biol. Chem. 2005, 280, 37565–37571. [Google Scholar] [CrossRef] [Green Version]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soares, A.G.; Mironova, E.; Archer, C.R.; Contreras, J.; Stockand, J.D.; Abd El-Aziz, T.M. Cisplatin Decreases ENaC Activity Contributing to Renal Salt Wasting Syndrome. Cancers 2020, 12, 2140. https://doi.org/10.3390/cancers12082140
Soares AG, Mironova E, Archer CR, Contreras J, Stockand JD, Abd El-Aziz TM. Cisplatin Decreases ENaC Activity Contributing to Renal Salt Wasting Syndrome. Cancers. 2020; 12(8):2140. https://doi.org/10.3390/cancers12082140
Chicago/Turabian StyleSoares, Antonio G., Elena Mironova, Crystal R. Archer, Jorge Contreras, James D. Stockand, and Tarek Mohamed Abd El-Aziz. 2020. "Cisplatin Decreases ENaC Activity Contributing to Renal Salt Wasting Syndrome" Cancers 12, no. 8: 2140. https://doi.org/10.3390/cancers12082140