Coumarins and Coumarin-Related Compounds in Pharmacotherapy of Cancer

 ,
,  and
and 
Abstract
:1. Introduction
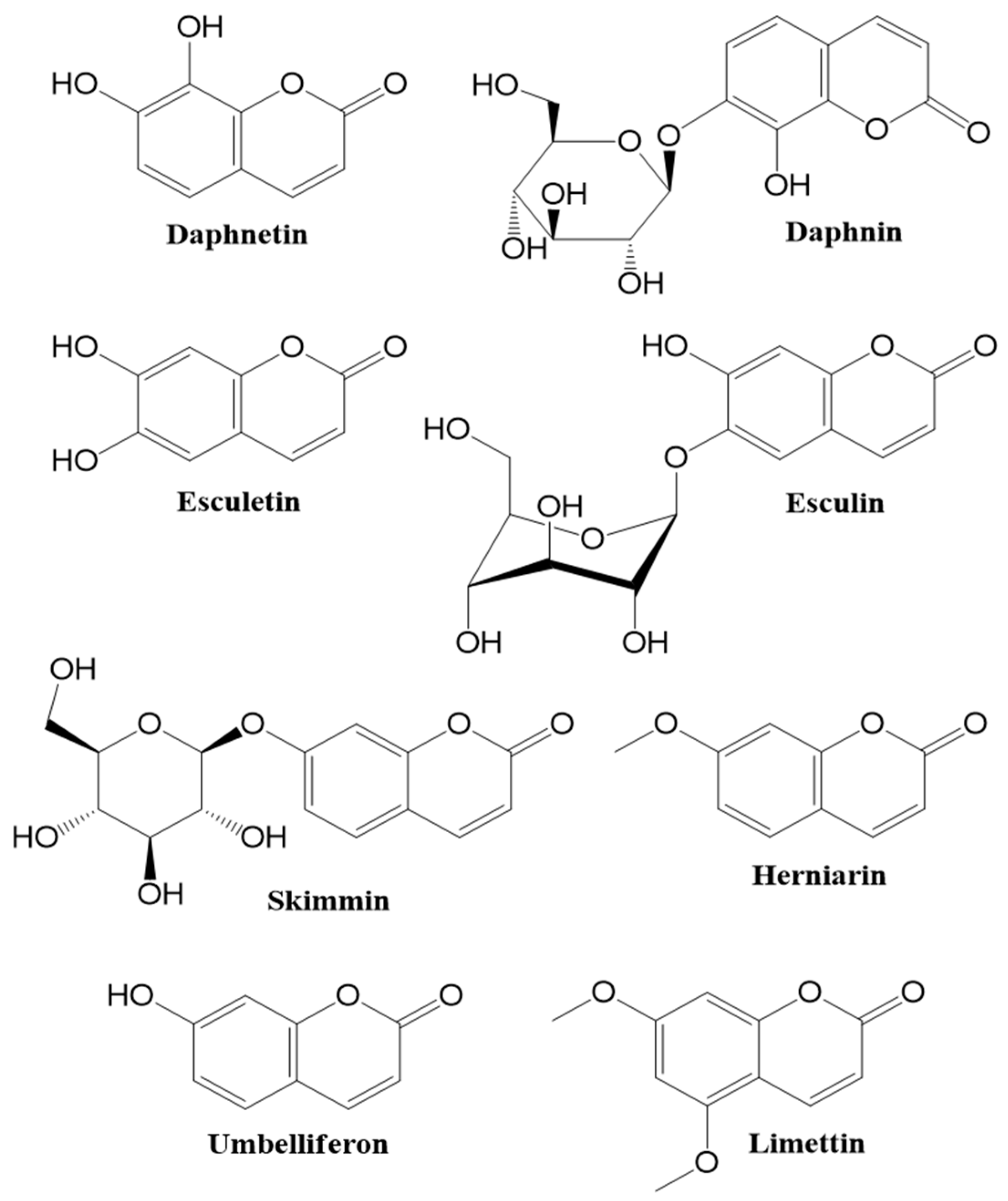
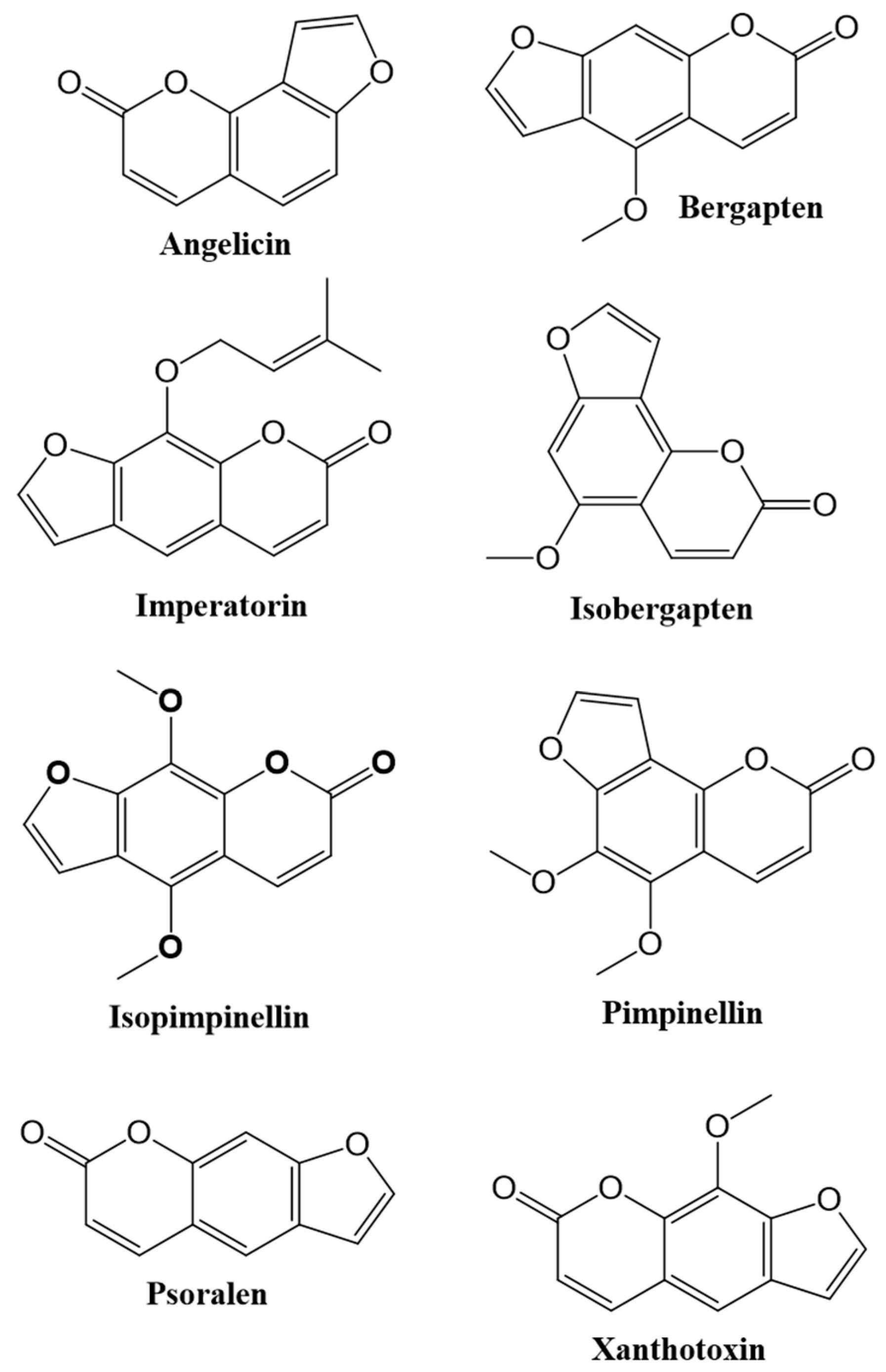
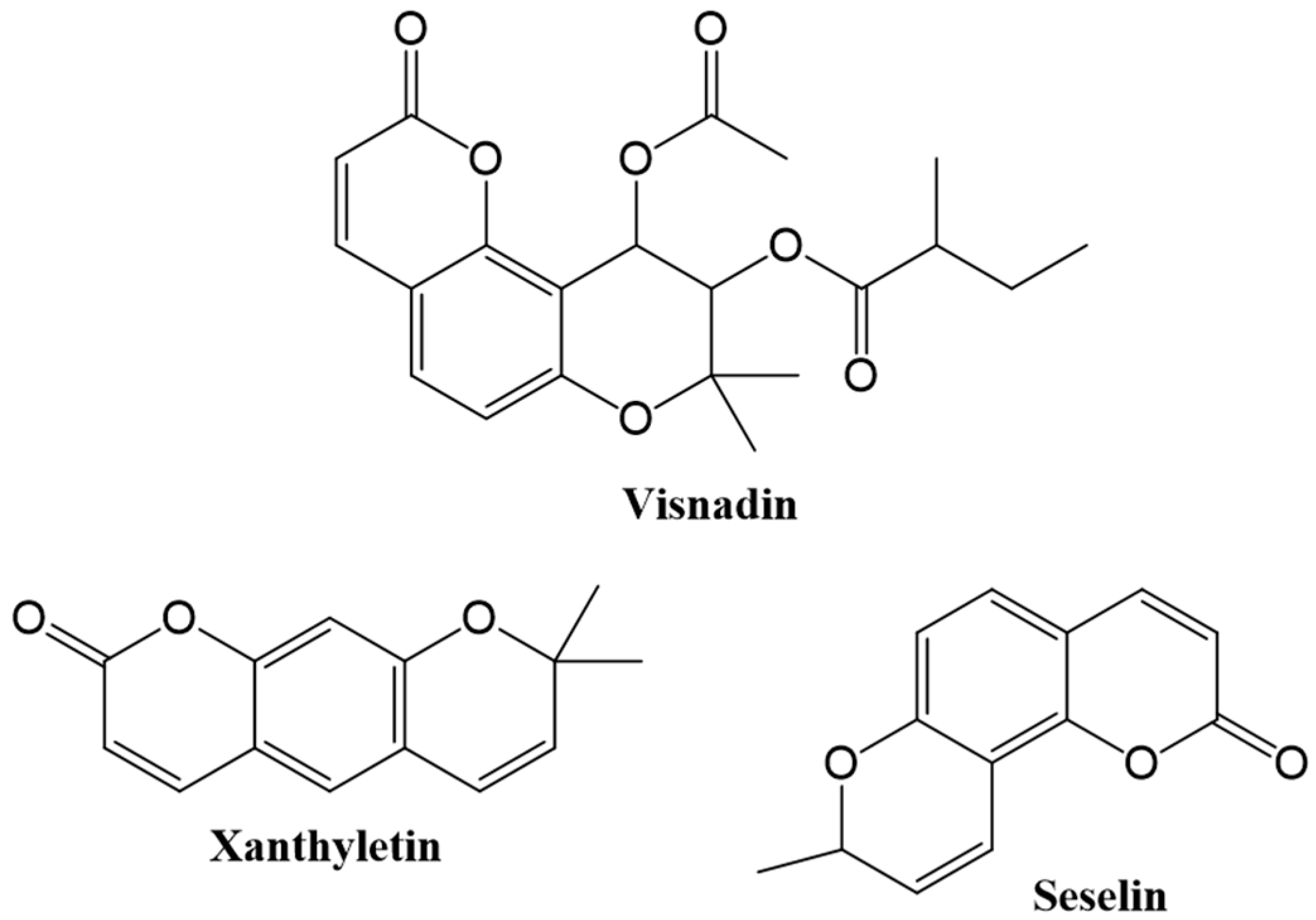
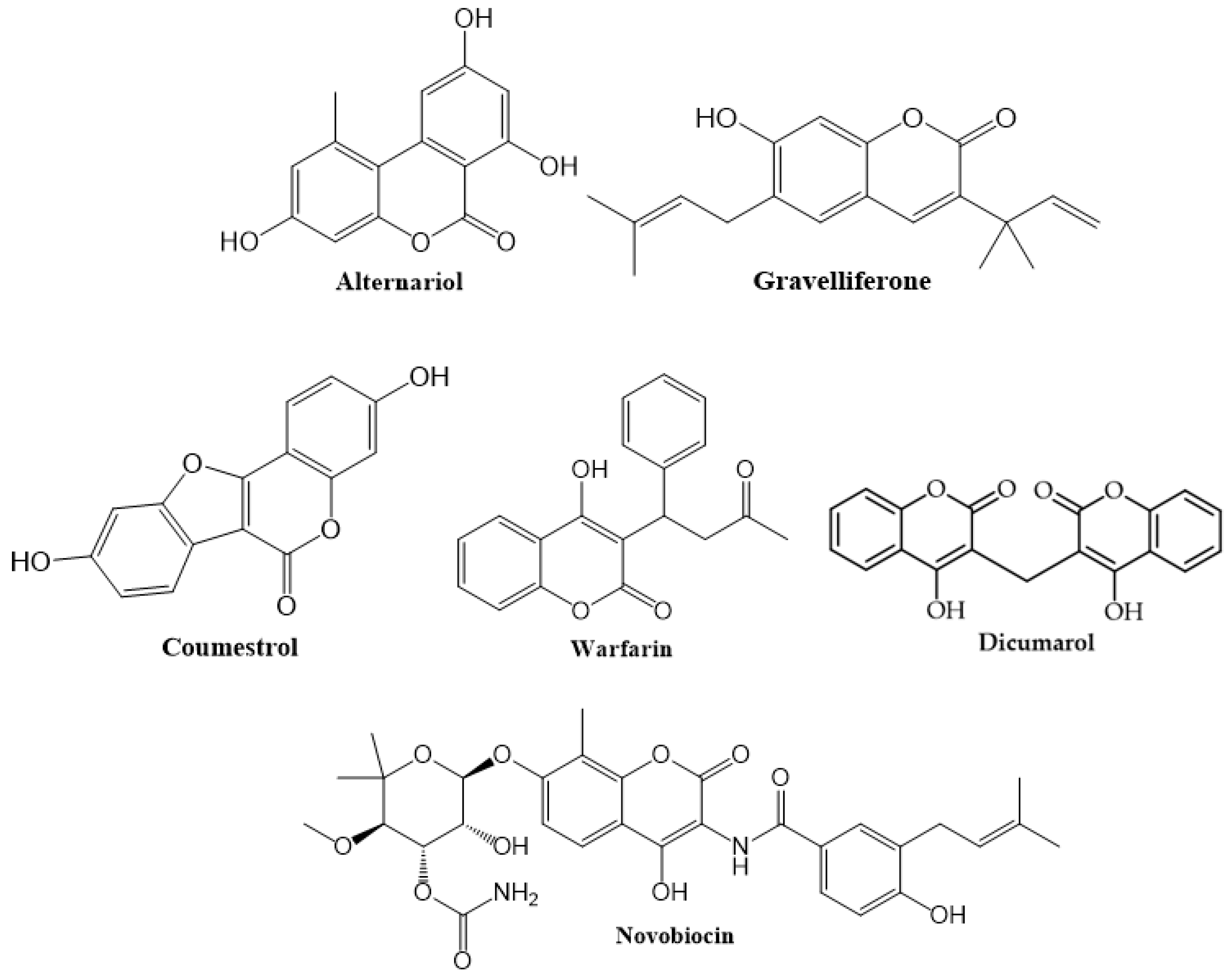
1.1. Occurence
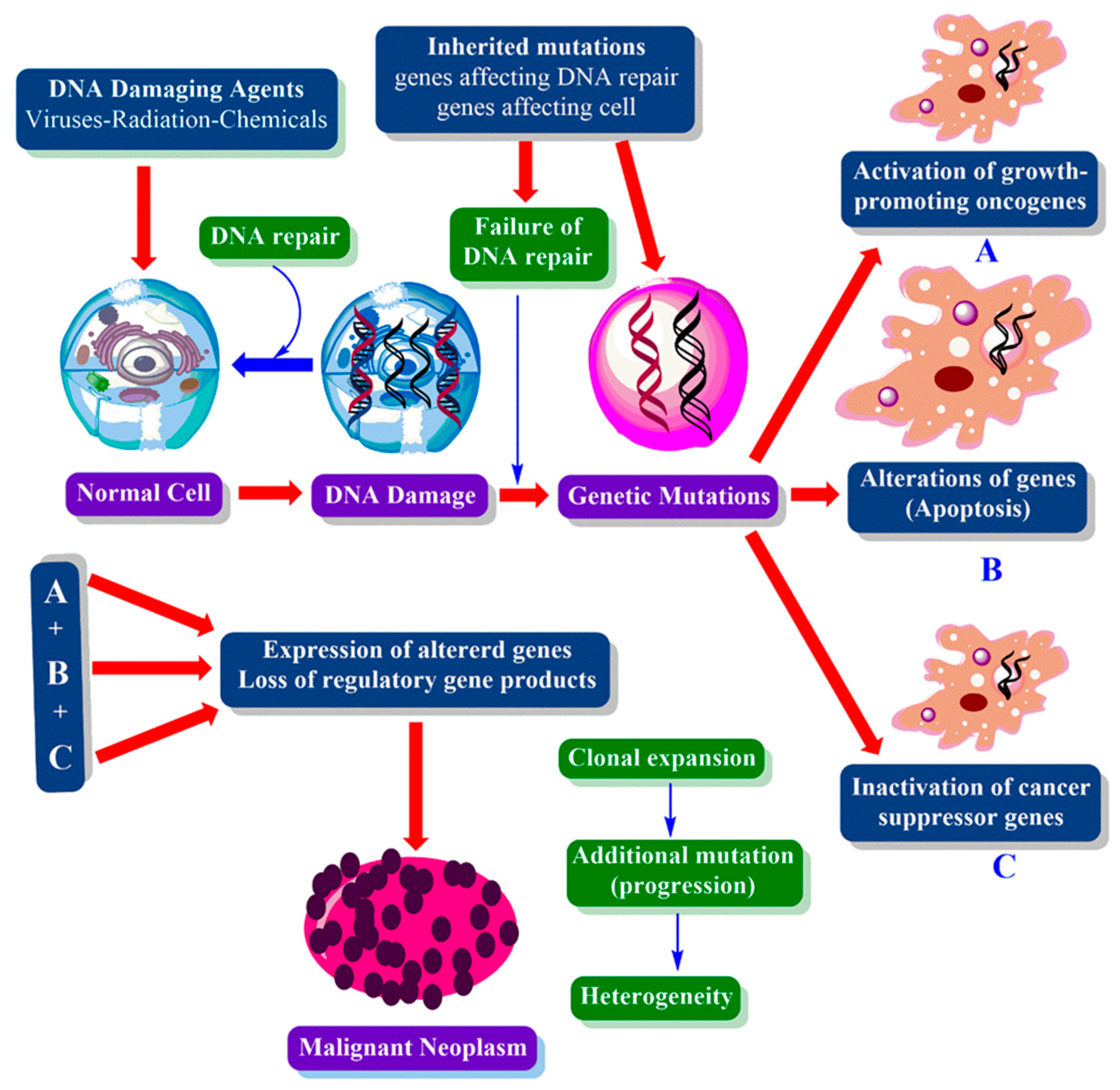
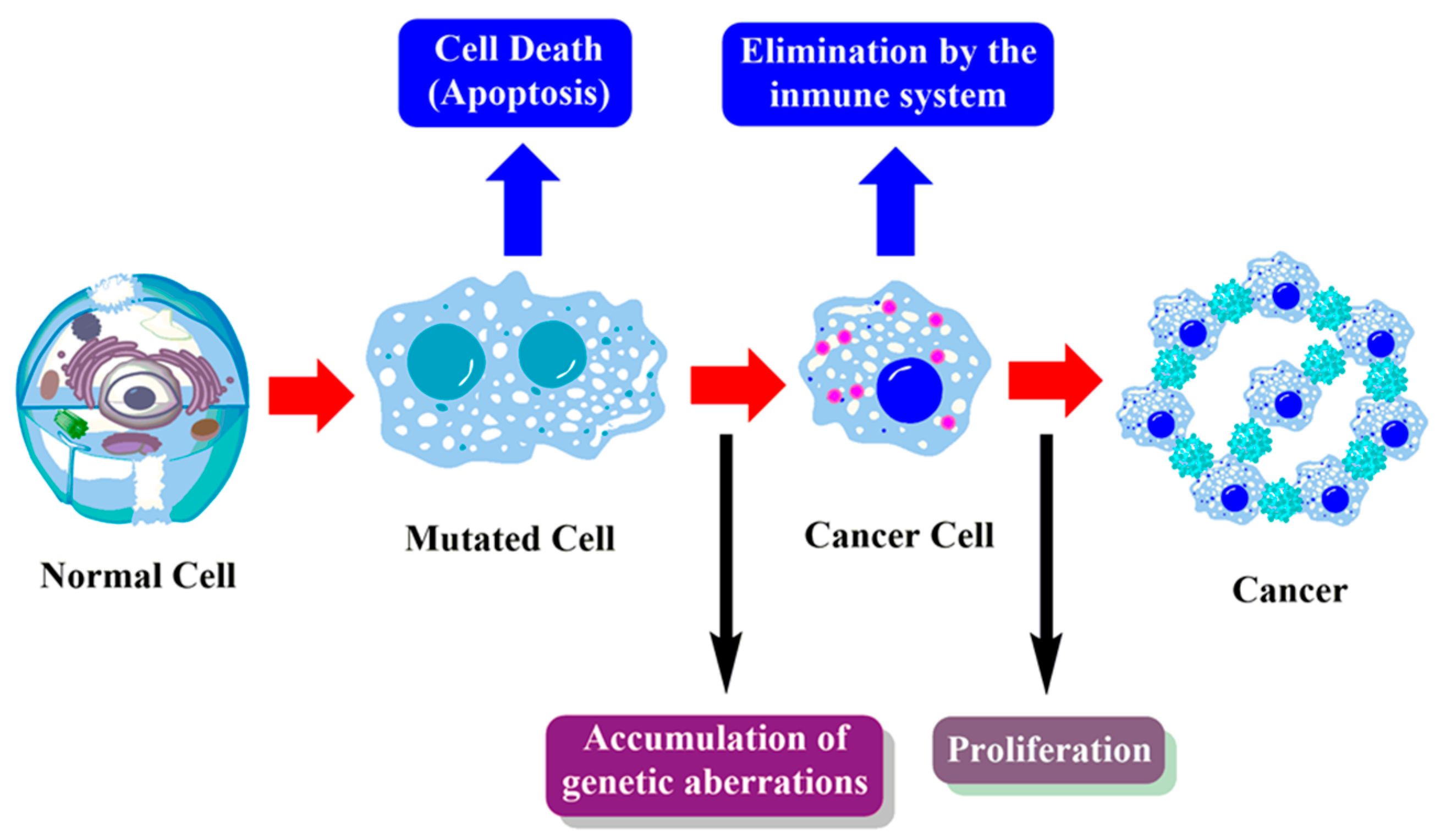
1.2. Cancer
1.3. Cancer Treatment
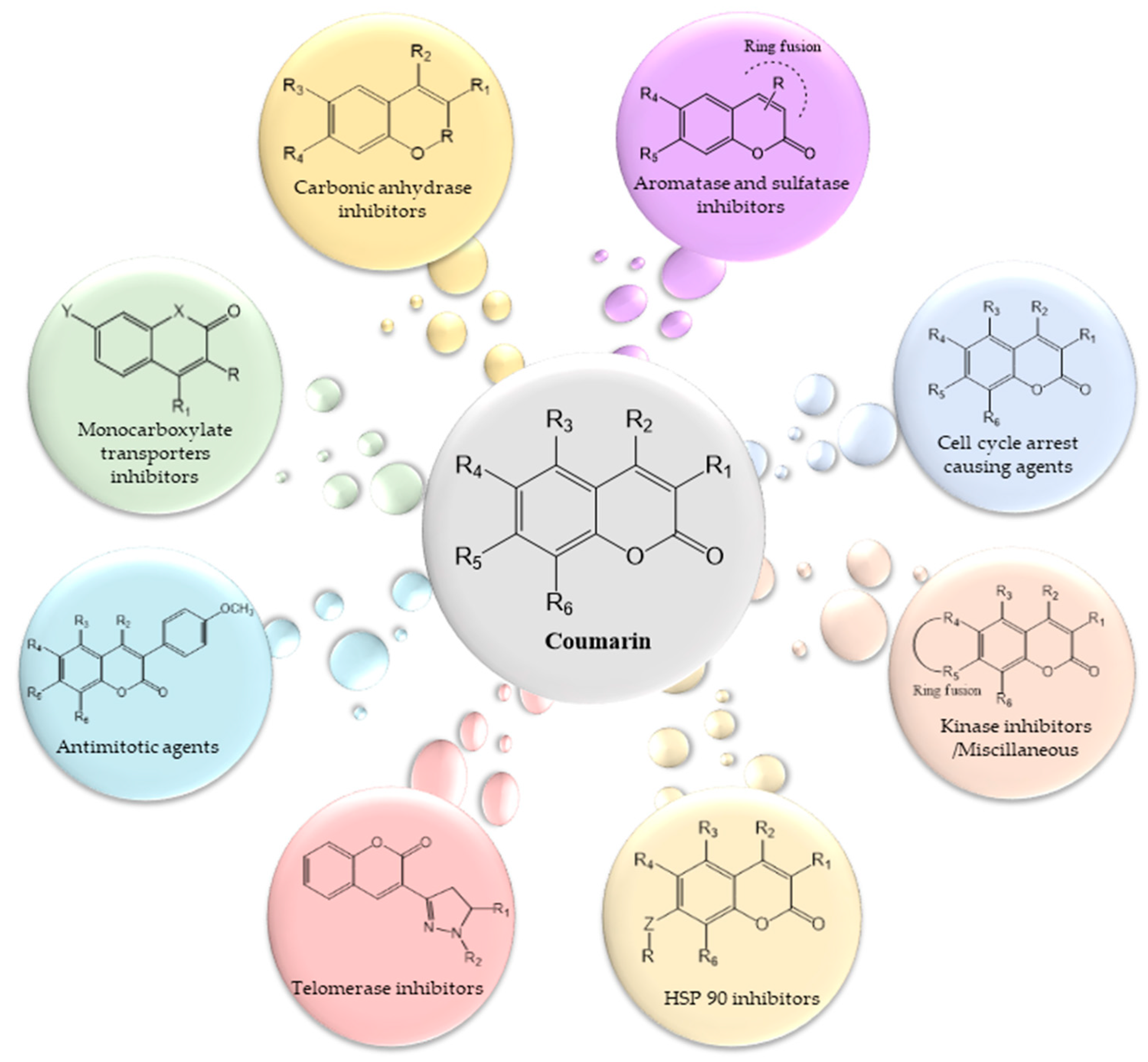
2. Roles of Coumarins in Anticancer Activity
2.1. Coumarins in Breast Cancer
2.2. Coumarins in Leukemia
2.3. Coumarins in Malignant Melanoma
2.4. Coumarins in Renal Cell Carcinoma
2.5. Coumarins in Prostate Cancer
2.6. Coumarins and Other Cancers
3. Pharmacokinetics
4. Toxicology
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bruneton, J. Immunotoxicity of Epicutaneously Applied Anti-Coagulant Rodenticide Warfarin; Intercept Ltd.: Hampshire, UK, 1999; pp. 245–263. [Google Scholar]
- Bruneton, J. Pharmacognosy, Phytochemistry, Medicinal Plants, 2nd ed.; Intercept Ltd.: Hampshire, UK, 1999; pp. 263–277. [Google Scholar]
- Ojala, T. Biological Screening of Plant Coumarins. Ph.D. Thesis, University of Helsinki, Helsinki, Finland, 2001. [Google Scholar]
- Lacy, A.; O’Kennedy, R. Studies on coumarins and coumarin-related compounds to determine their therapeutic role in the treatment of cancer. Curr. Pharm. Des. 2004, 10, 3797–3811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, R.D.H.; Mendez, J.; Brown, S.A. The Natural Coumarins Occurrence, Chemistry and Biochemistry, Chichester; John Wiley and Sons Ltd.: New York, NY, USA, 1982. [Google Scholar]
- Lake, B. Synthesis & pharmacological investigation of 4-hydroxy coumarin derivatives & shown as anti-coagulant. Food Chem. Tox. 1999, 3, 412–423. [Google Scholar]
- Cooke, D.; Fitzpatrick, B.; O’ Kennedy, R.; McCormack, T.; Egan, D. Coumarin: Biochemical Profile and Recent Developments; John Wiley & Sons: New York, NY, USA, 1997; Volume 3, pp. 311–322. [Google Scholar]
- Cooke, D.; O’Kennedy, R. Comparison of the Tetrazolium Salt Assay for Succinate Dehydrogenase with the Cytosensor Microphysiometer in the Assessment of Compound Toxicities. Anal. Biochem. 1999, 274, 188–194. [Google Scholar] [CrossRef] [PubMed]
- Vianna, D.R.; Hamerski, L.; Figueiró, F.; Bernardi, A.; Visentin, L.C.; Pires, E.N.; Teixeira, H.F.; Salbego, C.G.; Eifler-Lima, V.L.; Battastini, A.M.; et al. Selective cytotoxicity and apoptosis induction in glioma cell lines by 5-oxygenated-6,7-methylenedioxycoumarins from Pterocaulon species. Eur. J. Med. Chem. 2012, 57, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Harvey, R.G.; Cortex, C.; Ananthanarayan, T.P.; Schmolka, S. A new coumarin synthesis and its utilization for the synthesis of polycyclic coumarin compounds with anticarcinogenic properties. J. Org. Chem. 1988, 53, 3936–3943. [Google Scholar] [CrossRef]
- Kostova, I.; Momekov, G.; Tzanova, T.; Karaivanova, M. Synthesis, characterization, and cytotoxic activity of new lanthanum (III) complexes of bis-coumarins. Bioinorg. Chem. Appl. 2006, 25651, 1–9. [Google Scholar] [CrossRef]
- Al-Haiza, M.A.; Mostafa, M.S.; El-Kady, M.Y. Synthesis and Biological Evaluation of Some New Coumarin Derivatives. Molecules 2003, 8, 275–286. [Google Scholar] [CrossRef]
- Musiciki, B.; Periers, A.M.; Laurin, P.; Ferroud, D.; Benedetti, Y.; Lachaud, S.; Chatreaux, F.; Haesslein, J.L.; Lltis, A.; Pierre, C.; et al. Improved antibacterial activities of coumarin antibiotics bearing 5’,5’-dialkylnoviose: Biological activity of RU79115. Bioorg. Med. Chem. Lett. 2000, 10, 1695–1699. [Google Scholar] [CrossRef]
- Tosun, A.; Kupelı, E.; Yesılada, E. Anti-Inflammatory and Antinociceptive Activity of Coumarins from Seseli gummiferum subsp. corymbosum (Apiaceae). Z. Nat. C. 2009, 64c, 56–62. [Google Scholar] [CrossRef]
- Küpeli, E.; Tosun, A.; Yesilada, E. Anti-inflammatory and Antinociceptive Activities of Seseli L. Species (Apiaceae) Growing in Turkey. J. Ethnopharmacol. 2006, 104, 310–314. [Google Scholar] [CrossRef]
- Fylaktakidou, K.C.; Hadipavlou-Litina, D.J.; Litinas, K.E.; Nicolaides, D.N. Natural and synthetic coumarin derivatives with anti-inflammatory/ antioxidant activities. Curr. Pharm. Des. 2004, 10, 3813–3833. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.; Kin, J.; Park, O.S. A convenient one-pot synthesis of 4-hydroxycoumarin, 4-hydroxythiocoumarin, and 4-hydroxyquinolin-2(1H)-one. Synth. Commun. 2001, 31, 1195–1200. [Google Scholar] [CrossRef]
- Hoult, J.R.; Payá, M. Pharmacological and biochemical actions of simple coumarins: Natural products with therapeutic potential. Gen. Pharmacol 1996, 27, 713–722. [Google Scholar] [CrossRef]
- Madhavan, G.R.; Balraju, V.; Malleshasm, B.; Chakrabarti, R.; Lohray, V.B. Novel coumarin derivatives of heterocyclic compounds as lipid-lowering agents. Bioorg. Med. Chem. Lett. 2003, 13, 2547–2551. [Google Scholar] [CrossRef]
- Moffet, R.S. Central Nervous System Depressants. VII. Pyridyl Coumarins. J. Med. Chem. 1964, 7, 446–449. [Google Scholar] [CrossRef]
- Paya, M.; Halliwell, B.; Hoult, J.R. Interactions of a series of coumarins with reactive oxygen species. Scavenging of superoxide, hypochlorous acid and hydroxyl radicals. Biochem. Pharmacol. 1992, 44, 205–214. [Google Scholar] [CrossRef]
- Murray, R.D. The naturally occurring coumarins. Fortschr. Chem. Org. Naturst. 2002, 83, 1. [Google Scholar]
- Casley-Smith, J.R.; Morgan, R.G.; Piller, N.B. Treatment of lymphedema of the arms and legs with 5,6-benzo-[alpha]-pyrone. N. Engl. J. Med. 1993, 76, 1158–1163. [Google Scholar] [CrossRef]
- Casley-Smith, J.R.; Casley-Smith, J.R. Frequency of coumarin hepatotoxicity. Med. J. Aus. 1995, 162, 391. [Google Scholar] [CrossRef]
- Beinssen, A.P. Possible coumarin hepatotoxicity. Med. J. Aus. 1994, 161, 725. [Google Scholar] [CrossRef]
- Cox, D.; O’Kennedy, R.; Thornes, R.D. The rarity of liver toxicity in patients treated with coumarin (1,2-benzopyrone). Hum. Toxicol. 1989, 8, 501–506. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.M.; Ellwein, L.B. Genetic errors, cell proliferation, and carcinogenesis. Cancer Res. 1991, 51, 6493–6505. [Google Scholar] [PubMed]
- Fentem, J.H.; Fry, J.R. Species differences in the metabolism and hepatotoxicity of coumarin. Comp. Biochem. Physiol. C. 1993, 104, 1–8. [Google Scholar] [CrossRef]
- Myers, R.B.; Parker, M.; Grizzle, W.E. The Effects of Coumarin and Suramin on the Growth of Malignant Renal and Prostatic Cell Lines. J. Cancer Res. Clin. Oncol. 1994, 120, 11–13. [Google Scholar] [CrossRef]
- Carotti, A.; Carriri, A.; Chimichi, S.; Boccalini, M.; Cosimelli, B.; Gnerre, C.; Carrupt, P.A.; Testa, B. Natural and synthetic geiparvarins are strong and selective MAO-B inhibitors. Synthesis and SAR studies. Bioorg. Med. Chem. Lett. 2002, 72, 3551–3555. [Google Scholar] [CrossRef]
- Okamoto, T.; Kobayashi, T.; Yoshida, S. Chemical aspects of coumarin compounds for the prevention of hepatocellular carcinomas. Curr. Med. Chem. Anticancer Agents 2005, 5, 47–51. [Google Scholar] [CrossRef]
- Marshall, M.E.; Ryles, M.; Butler, K.; Weiss, L. Treatment of advanced renal cell carcinoma (RCC) with coumarin and cimetidine: Longterm follow-up of patients treated on a phase I trial. J. Cancer Res. Clin. Oncol. 1994, 120, 535–538. [Google Scholar]
- Stanchev, S.; Momekov, G.; Jensen, F.; Manolov, I. Synthesis, computational study and cytotoxic activity of new 4-hydroxycoumarin derivatives. Eur. J. Med. Chem. 2008, 43, 694–706. [Google Scholar] [CrossRef]
- Thornes, R.D.; Daly, L.; Lynch, G.; Breslin, B.; Browne, H.; Browne, H.Y.; Corrigan, T.; Daly, P.; Edwards, G.; Gaffney, E.; et al. Treatment With Coumarin to Prevent or Delay Recurrence of Malignant Melanoma. Cancer Res. Clin. Oncol. 1994, 120, 32–34. [Google Scholar] [CrossRef]
- Mohler, J.L.; Gomella, L.G.; Crawford, E.D.; Glode, L.M.; Zippe, C.D.; Fair, W.R.; Marshall, M.E. Phase II Evaluation of Coumarin (1,2-Benzopyrone) in Metastatic Prostatic Carcinoma. Prostate 1992, 20, 123–131. [Google Scholar] [CrossRef]
- Budzisz, E.; Brzezinska, E.; Krajewska, U.; Rozalski, M. Cytotoxic Effects, Alkylating Properties and Molecular Modelling of Coumarin Derivatives and Their Phosphonic Analogues. Eur. J. Med. Chem. 2003, 38, 597–603. [Google Scholar] [CrossRef]
- Kostova, I.; Raleva, S.; Genova, P.; Argirova, R. Structure-Activity Relationships of Synthetic Coumarins as HIV-1 Inhibitors. Bioinorg. Chem. Appl. 2006, 68274, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, U.S.; Steffen, B.; Siegers, C.P. Antitumour-Activities of Coumarin, 7-Hydroxycoumarin and its Glucuronide in Several Human Tumour Cell Lines. Res. Commun. Mol. Pathol. Pharmacol. 1998, 99, 193–206. [Google Scholar] [PubMed]
- Keating, G.; O’Kennedy, R. The Chemistry and Occurrence of Coumarins. Coumarins: Biology, Applications and Mode of Action; O’Kennedy, R., Thornes, R.D., Eds.; John Wiley & Sons: Chichester, UK, 1997; pp. 23–66. [Google Scholar]
- Trease, G.E. A Textbook of Pharmacognosy, 8th ed.; Baillierie Tindall: London, UK, 1961. [Google Scholar]
- Guenther, E. The Oils Vol II; D. Van Ltd.: New York, NY, USA, 1975. [Google Scholar]
- Zechmeister, L. Fortsohritte der Chemie Organischer Naturstoffe, Naturally Occuring Coumarins, Herausgeber Editor Redacteur; Springer: Wien, Austria, 1952. [Google Scholar]
- Ahmad, J.; Shamsuddin, K.M.; Zaiman, A. A Pyranocoumarin from Atalantia ceylanica. Phytochemistry 1984, 23, 2098–2099. [Google Scholar] [CrossRef]
- Kresge, N.; Simoni, R.D.; Hill, R.L. Hemorrhagic Sweet Clover Disease, Dicumarol, and Warfarin: The Work of Karl Paul Link. J. Biol. Chem. 2005, 280, e5. [Google Scholar]
- Pan, L.; Chai, H.; Kinghorn, A.D. The Continuing Search for Antitumor Agents from Higher Plants. Phytochem. Lett. 2010, 3, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Majnooni, M.B.; Fakhri, S.; Shokoohinia, Y.; Mojarrab, M.; Kazemi-Afrakoti, S.; Farzaei, M.H. Isofraxidin: Synthesis, Biosynthesis, Isolation, Pharmacokinetic and Pharmacological Properties. Molecules 2020, 25, 2040. [Google Scholar] [CrossRef]
- Pádua, D.; Rocha, E.; Gargiulo, D.; Ramos, A.A. Bioactive Compounds from Brown Seaweeds: Phloroglucinol, Fucoxanthin and Fucoidan as Promising Therapeutic Agents Against Breast Cancer. Phytochem. Lett. 2015, 14, 91–98. [Google Scholar] [CrossRef]
- Bourgaud, F.; Hehn, A.; Larbat, R.; Doerper, S.; Gontier, E.; Kellner, S.; Matern, U. Biosynthesis of coumarins in plants: A major pathway still to be unravelled for cytochrome P450 enzymes. Phytochem. Rev. 2006, 5, 293–308. [Google Scholar] [CrossRef]
- Blackadar, C.B. Historical review of the causes of cancer. World J. Clin. Oncol. 2016, 7, 54–86. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2015. CA Cancer J. Clin. 2015, 65, 5–29. [Google Scholar] [CrossRef] [PubMed]
- Fitzmaurice, C.; Dicker, D.; Pain, A.; Hamavid, H.; Moradi-Lakeh, M.; MacIntyre, M.F.; Allen, C.; Hansen, G.; Woodbrook, R.; Wolfe, C.; et al. Global Burden of Disease Cancer Collaboration, The Global Burden of Cancer 2013. JAMA Oncol. 2015, 1, 505–527. [Google Scholar] [PubMed]
- Pavlopoulou, A.; Spandidos, D.A.; Michalopoulos, I. Human cancer databases (review). Oncol. Rep. 2015, 33, 3–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khoobi, M.; Foroumadi, A.; Emami, S.; Safavi, M.; Dehghan, G.; Alizadeh, B.; Ramzani, A.; Ardastani, S.; Shafiee, A. Coumarin based bioactive compounds: Facile synthesis and biological evaluation of coumarin-fused 1,4-Thiazepines. Chem. Bio. Drug Des. 2011, 78, 580–586. [Google Scholar] [CrossRef] [PubMed]
- Carter, S.K.; Bakowski, M.T.; Hellman, K. Chemotherapy of Cancer, 3rd ed.; Wiley & Sons: New York, UY, USA, 1989. [Google Scholar]
- Semini, G.; Hildmann, A.; Haefen, C.; Danker, K. Glycosidated phospholipids—A promising group of anti-tumour lipids. Anti. Cancer Agents Med. Chem. 2014, 14, 607–617. [Google Scholar] [CrossRef]
- Kokron, O.; Maca, S.; Gasser, G.; Schmidt, P.R. Synthesis of novel coumarin 3-(N-aryl)-sulfonamides & evaluated for their anti-cancer activity and reported in vitro inhibitory activity on human platelet aggregation. Oncology 1991, 48, 91–102. [Google Scholar]
- Marshall, M.E.; Butler, K.; Fried, A. Screening of synthetic new hetrocyclic derivatives of 3-formyl-4-hydroxy coumarin and evaluated for Anti-Inflammatory activity in albino rats. Mol. Biother. 1991, 3, 158–170. [Google Scholar]
- Bosland, M.C. Synthesis of vanillin ethers from bromo- methylcoumarins as anti-inflammatory agents. San Diego Acad. Press 1991, 6, 162–177. [Google Scholar]
- Sandhu, S.; Bansal, Y.; Silakari, O.; Bansal, G. Coumarin hybrids as novel therapeutic agents. Bioorg. Med. Chem. 2014, 22, 3806–3814. [Google Scholar] [CrossRef]
- Mann, J. Natural products in cancer chemotherapy: Past, present and future. Nat. Rev. Can. 2002, 2, 143–148. [Google Scholar] [CrossRef]
- Farabegoli, F.; Papi, A.; Bartolini, G.; Ostan, R.; Orlandi, M. (-)-Epigallocatechin-3-gallate downregulates Pg-P and BCRP in a tamoxifen resistant MCF-7 cell line. Phytomedicine 2010, 17, 356–362. [Google Scholar] [CrossRef] [PubMed]
- Nautiyal, J.; Banerjee, S.; Kanwar, S.S.; Yu, Y.; Patel, B.B.; Sarkar, F.H.; Majumdar, A.P. Curcumin enhances dasatinib-induced inhibition of growth and transformation of colon cancer cells. Int. J. Cancer. 2011, 128, 951–961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sen, S.; Sharma, H.; Singh, N. Curcumin enhances Vinorelbine mediated apoptosis in NSCLC cells by the mitochondrial pathway. Biochem. Biophys. Res. Commun. 2005, 331, 1245–1252. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.Z.; Yuan, C.S. Potential role of ginseng in the treatment of colorectal cancer. Am. J. Chin. Med. 2008, 36, 1019–1028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banerjee, S.; Kambhampati, S.; Haque, I.; Banerjee, S.K. Pomegranate sensitizes Tamoxifen action in ER-α positive breast cancer cells. J. Cell Commun. Signal. 2011, 5, 317–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishii, T.; Teramoto, S.; Matsuse, T. GSTP1 affects chemoresistance against camptothecin in human lung adenocarcinoma cells. Cancer Lett. 2004, 216, 89–102. [Google Scholar] [CrossRef]
- Nakashima, K.; Virgona, N.; Miyazawa, M.; Watanabe, T.; Yano, T. The tocotrienol-rich fraction from rice bran enhances cisplatin-induced cytotoxicity in human mesothelioma H28 cells. Phytother. Res. 2010, 24, 1317–1321. [Google Scholar] [CrossRef]
- Finn, G.; Kenealy, E.; Creaven, B.; Egan, D. In vitro Cytotoxic Potential and Mechanism of Action of Selected Coumarins, Using Human Renal Cell Lines. Cancer Lett. 2002, 183, 61–68. [Google Scholar] [CrossRef]
- Loprinzi, C.L.; Kugler, J.W.; Sloan, J.A.; Rooke, T.W.; Quella, S.K.; Novotny, P.; Mowat, R.B.; Michalak, J.C.; Stella, P.J.; Levitt, R.; et al. Lack of effect of coumarin in women with lymphedema after treatment for breast cancer. N. Engl. J. Med. 1999, 340, 346–350. [Google Scholar] [CrossRef]
- Grotz, K.A.; Wustenberg, P.; Kohnen, R.; Al-Nawas, B.; Henneicke-von Zepelin, H.H.; Bockisch, A.; Kutzner, J.; Naser-Hijazi, B.; Belz, G.G.; Wagner, W. Prophylaxis Of Radiogenic Sialadenitis and Mucositis by Coumarin/Troxerutine in Patients with Head and Neck Cancer-A Prospective, Randomized, Placebo-Controlled, Double-Blind Study. Br. J. Oral Maxillofac. Surg. 2001, 39, 34–39. [Google Scholar] [CrossRef]
- Agarwal, R. Synthesis & biological screening of some novel coumarin derivatives. Biochem. Pharmacol. 2000, 6, 1042–1051. [Google Scholar]
- Marshall, M.E.; Butler, K.; Hermansen, D. Treatment of Hormone-Refractory Stage-D Carcinoma of Prostate with Coumarin (1,2- Benzopyrone) and Cimetidine: A Pilot Study. Prostate 1990, 17, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Mahler, J.L.; Gomella, L.G.; Crawford, E.D.; Glode, L.M.; Zippe, C.D.; Fair, W.R. Synthesis of novel coumarin bisindole derivatives & reported as anti-hyperlipidimic activity. Prostate 1992, 20, 112–123. [Google Scholar]
- Egan, D.; James, P.; Cooke, D.; O’Kennedy, R. Studies on the Cytostatic and Cytotoxic Effects and Mode of Action of 8-Nitro-7-Hydroxycoumarin. Cancer Lett. 1997, 118, 201–211. [Google Scholar] [CrossRef]
- Yang, J.; Liu, X.; Bhalla, K.; Kim, C.N.; Ibrado, A.M.; Cai, J. Prevention of apoptosis by bcl-2; release of cytochrome c from mitochondria blocked. Science 1997, 275, 1129–1132. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, H.; Kong, Y.Y.; Yoshida, R.; Elia, A.J.; Hakem, R.; Penninger, J.M.; Mak, T.W. Apaf-1 is required for mitochondrial pathways of apoptosis and brain development. Cell 1998, 94, 739–750. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, H.; Wang, H.G. The protein kinase PKB/Akt regulates cell survival and apoptosis by inhibiting Bax conformational change. Oncogene 2001, 20, 7780–7786. [Google Scholar] [CrossRef] [Green Version]
- Baez, C.; Leon, F.; Santos, E. Effects of coumarin and 7-OH- coumarin on bcl-2 and Bax expression in two human lung cancer cell lines in vitro. J. Cell. Biol. Int. 2009, 29, 703–708. [Google Scholar]
- Wellbrock, C.; Karasarides, M.; Marais, R. The RAF proteins take centre stage. Nat. Rev. Mol. Cell Biol. 2004, 5, 875–885. [Google Scholar] [CrossRef]
- Dumitru, C.D.; Ceci, J.D.; Tsatsanis, C.; Kontoyiannis, D.; Stamatakis, K.; Lin, J.H.; Patriotis, C.; Jenkins, N.A.; Copeland, N.G.; Kollias, G.; et al. TNF-alpha induction by LPS is regulated post transcriptionally via a Tpl2/ERK-dependent pathway. Cell 2000, 103, 1071–1083. [Google Scholar] [CrossRef] [Green Version]
- Ohren, J.F.; Chen, H.; Pavlovsky, A.; Whitehead, C.; Zhang, E.; Kuffa, P.; Yan, C.; McConnell, P.; Spessard, C.; Banotai, C.; et al. Structures of human MAP kinase kinase 1 (MEK1) and MEK2 describe novel non-competitive kinase inhibition. Nat. Struct. Mol. Biol. 2004, 11, 1192–1197. [Google Scholar] [CrossRef] [PubMed]
- Goel, A.; Prasad, A.; Parmar, V.; Ghosh, B.; Saini, N. 7,8-Dihydroxy-4-methylcoumarin induces apoptosis of human lung adenocarcinoma cells by ROS-independent mitochondrial pathway through partial inhibition of ERK/MAPK signaling. J. FEBS. Lett. 2007, 581, 2447–2454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valente, S.; Bana, E.; Viry, E.; Bagrel, D.; Kirsch, G. Synthesis and biological evaluation of novel coumarin-based inhibitors of Cdc25 phosphatses. Bioorg. Med. Chem. Lett. 2010, 20, 5827–5830. [Google Scholar] [CrossRef] [PubMed]
- Boutros, R.; Lobjois, V.; Ducommun, B. CDC25 phoshatases in cancer cells: Key players? Good targets? Nat. Rev. Cancer 2007, 7, 495–507. [Google Scholar] [CrossRef] [PubMed]
- Rudolph, J. Cdc25 Phosphatases: Structure, Specificity, and Mechanism. Biochemistry 2007, 46, 3595–3600. [Google Scholar] [CrossRef]
- Niida, H.; Nakanishi, M. DNA damage checkpoints in mammals. Mutagenesis 2006, 21, 3–9. [Google Scholar] [CrossRef]
- Aressy, B.; Ducommun, B. Cell cycle control by the CDC25 phosphatases. Anticancer Agents Med. Chem. 2008, 8, 818–824. [Google Scholar] [CrossRef]
- Kacar, A.K.; Bahadori, F.; Tekkeli, S.E.K.; Topcu, G.; Bolkent, S. Investigation of cell death mechanism and activity of esculetin-loaded PLGA nanoparticles on insulinoma cells in vitro. J. Pharm. Pharmacol. 2020, 72, 592–606. [Google Scholar] [CrossRef]
- Stacey, D.; DeGrasse, C.; Johnston, L. Addressing the support needs of women at high risk for breast cancer: Evidence-based care by advanced practice nurses. Oncol. Nurs. Forum 2002, 29, 77–84. [Google Scholar] [CrossRef]
- Jemal, A.; Bray, F.; Center, M.M.; Ferlay, J.; Ward, E.; Forman, D. Global cancer statistics. CA Cancer J. Clin. 2011, 61, 69–90. [Google Scholar] [CrossRef] [Green Version]
- Parkin, D.M.; Bray, F.; Ferlay, J.; Pisani, P. Estimating the world cancer burden: Globocan 2000. Int. J. Cancer 2001, 94, 153–156. [Google Scholar] [CrossRef] [PubMed]
- Wooster, R.; Weber, B.L. Breast and ovarian cancer. N. Engl. J. Med. 2003, 348, 2339–2347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foulkes, W.D. Inherited susceptibility to common cancers. N. Engl. J. Med. 2008, 359, 2143–2153. [Google Scholar] [CrossRef]
- Beral, V.; Bull, D.; Doll, R.; Peto, R.; Reeves, G. Breast cancer and breastfeeding: Collaborative reanalysis of individual data from 47 epidemiological studies in 30 countries, including 50 302 women with breast cancer and 96 973 women without the disease. Lancet 2002, 360, 187–195. [Google Scholar]
- Yersal, O.; Barutca, S. Biological subtypes of breast cancer: Prognostic and therapeutic implications. World J. Clin. Oncol. 2014, 5, 412–424. [Google Scholar] [CrossRef]
- De Ronde, J.J.; Hannemann, J.; Halfwerk, H.; Mulder, L.; Straver, M.E.; Vrancken Peeters, M.J.; Wesseling, J.; van de Vijver, M.; Wessels, L.F.; Rodenhuis, S. Concordance of clinical and molecular breast cancer subtyping in the context of preoperative chemotherapy response. Breast Cancer Res. Treat. 2010, 119, 119–126. [Google Scholar] [CrossRef] [Green Version]
- Perou, C.M.; Sørlie, T.; Eisen, M.B.; van de Rijn, M.; Jeffrey, S.S.; Rees, C.A.; Pollack, J.R.; Ross, D.T.; Johnsen, H.; Akslen, L.A.; et al. Molecular portraits of human breast tumours. Nature 2000, 406, 747–752. [Google Scholar] [CrossRef] [PubMed]
- Ignatiadis, M.; Bedard, P.; Haibe-Kains, B.; Haibe-Kains, B.; Singhal, S.; Loi, S.; Criscitiello, C.; Desmedt, C.; Bontempi, G.; Piccart, M.; et al. A meta-analysis of gene expression profiling studies identifies clinically relevant oncogenic pathways in basal like breast cancer. Cancer Res. 2009, 69, 106. [Google Scholar]
- Tran, B.; Bedard, P.L. Luminal-B breast cancer and novel therapeutic targets. Breast Cancer Res. 2011, 13, 221. [Google Scholar] [CrossRef] [Green Version]
- Fu, X.; Osborne, C.K.; Schiff, R. Biology and therapeutic potential of PI3K signaling in ER+/HER2-negative breast cancer. Breast 2013, 22, 12–18. [Google Scholar] [CrossRef]
- Eliyatkın, N.; Özgür, H.; Erçetin, P.; Aktaş, S.; Küpelioğlu, A. Meme karsinomlarında HER-2 durumunun immünohistokimyasal ve moleküler analizlerle değerlendirilmesi. Tepecik Eğit. Araşt. Hast. Derg. 2015, 25, 19–27. [Google Scholar]
- Moasser, M.M. The oncogene HER2: Its signaling and transforming functions and its role in human cancer pathogenesis. Oncogene 2007, 26, 6469–6487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutierrez, C.; Schiff, R. HER2: Biology, detection, and clinical implications. Arch. Pathol. Lab. Med. 2011, 135, 55–62. [Google Scholar] [PubMed]
- Tsutsui, S.; Ohno, S.; Murakami, S.; Kataoka, A.; Kinoshita, J.; Hachitanda, Y. Prognostic significance of the coexpression of p53 protein and c-erbB2 in breast cancer. Am. J. Surg. 2003, 185, 165–167. [Google Scholar] [CrossRef]
- Diniz, G.; Irkkan, Ç.; Kelten, C.; Özekinci, S. HER2 değerlendirmesindeki ipucu ve tuzaklar. Tepecik Eğit. Araşt. Hast. Derg. 2015, 25, 7–12. [Google Scholar]
- Anderson, W.F.; Rosenberg, P.S.; Prat, A.; Perou, C.M.; Sherman, M.E. How many etiological subtypes of breast cancer: Two, three, four, or more? J. Natl. Cancer Inst. 2014, 106, 165. [Google Scholar] [CrossRef] [Green Version]
- Wesolowski, R.; Ramaswamy, B. Gene expression profiling: Changing face of breast cancer classification and management. Gene Expr. 2011, 15, 105–115. [Google Scholar] [CrossRef]
- Putti, T.C.; El-Rehim, D.M.A.; Rakha, E.A.; Paish, C.E.; Lee, A.H.S.; Pinder, S.E.; Ellis, I.O. Estrogen receptor-negative breast carcinomas: A review of morphology and immunophenotypical analysis. Mod. Pathol. 2005, 18, 26–35. [Google Scholar] [CrossRef]
- Kesse-Adu, R.; Shousha, S. Myoepithelial markers are expressed in at least 29% of oestrogen receptor negative invasive breast carcinoma. Mod. Pathol. 2004, 17, 646–652. [Google Scholar] [CrossRef] [Green Version]
- Simpson, E.R.; Mahendroo, M.S.; Means, G.D.; Kilgore, M.W.; Hinshelwood, M.M.; Graham-Lorence, S.; Amarneh, B.; Ito, Y.; Fisher, C.R.; Michael, M.D.; et al. Aromatase cytochrome P450, the enzyme responsible for estrogen biosynthesis. Endocr. Rev. 1994, 15, 342–355. [Google Scholar]
- Coombes, R.C.; Gibson, L.; Hall, E.; Emson, M.; Bliss, J. Aromatase inhibitors as adjuvant therapies in patients with breast cancer. J. Steroid Biochem. Mol. Biol. 2003, 86, 309–311. [Google Scholar] [CrossRef]
- Henderson, I.C.; Canellos, G.P. Cancer of the breast: The past decade (first of two parts). N. Engl. J. Med. 1980, 302, 17–30. [Google Scholar] [CrossRef] [PubMed]
- Hamelers, I.; Schaik, R.; Sussenbach, J.S.; Steenbergh, P.H. 17beta-Estradiol responsiveness of MCF-7 laboratory strains is dependent on an autocrine signal activating the IGF type I receptor. Cancer Cell Int. 2003, 3, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strott, C.A. Sulfonation and molecular action. Endocr. Rev. 2002, 23, 703–732. [Google Scholar] [CrossRef] [PubMed]
- Utsumi, T.; Yoshimura, N.; Takeuchi, S.; Ando, J.; Maruta, M.; Maeda, K.; Harada, N. Steroid sulfatase expression is an independent predictor of recurrence in human breast cancer. Cancer Res. 1999, 59, 377–381. [Google Scholar] [PubMed]
- Miyoshi, Y.; Ando, A.; Hasegawa, S.; Ishitobi, M.; Taguchi, T.; Tamaki, Y.; Noguchi, S. High expression of steroid sulfatase mRNA predicts poor prognosis in patients with estrogen receptor-positive breast cancer. Clin. Cancer Res. 2003, 9, 2288–2293. [Google Scholar]
- Suzuki, T.; Nakata, T.; Miki, Y.; Kaneko, C.; Moriya, T.; Ishida, T.; Akinaga, S.; Hirakawa, H.; Kimura, M.; Sasano, H. Estrogen sulfotransferase and steroid sulfatase in human breast carcinoma. Cancer Res. 2003, 63, 2762–2770. [Google Scholar]
- Yoshimura, N.; Harada, N.; Bukholm, I.; Karesen, R.; Borresen-Dale, A.L.; Kristensen, V.N. Intratumoural mRNA expression of genes from the oestradiol metabolic pathway and clinical and histopathological parameters of breast cancer. Breast Cancer Res. 2004, 6, R46–R55. [Google Scholar] [CrossRef] [Green Version]
- Jonat, W.; Howell, A.; Blomqvist, C.; Eiermann, W.; Winblad, G.; Tyrrell, C.; Mauriac, L.; Roche, H.; Lundgren, S.; Hellmund, R.; et al. A randomised trial comparing two doses of the new selective aromatase inhibitor anastrozole (Arimidex) with megestrol acetate in postmenopausal patients with advanced breast cancer. Eur. J. Cancer 1996, 32A, 404–412. [Google Scholar] [CrossRef]
- Santner, S.J.; Feil, P.D.; Santen, R.J. In situ estrogen production via the estrone sulfatase pathway in breast tumors: Relative importance versus the aromatase pathway. J. Clin. Endocrinol. Metab. 1984, 59, 29–33. [Google Scholar] [CrossRef]
- Yamamoto, T.; Kitawaki, J.; Urabe, M.; Honjo, H.; Tamura, T.; Noguchi, T.; Okada, H.; Sasaki, H.; Tada, A.; Yoshiteru, T.; et al. Estrogen productivity of endometrium and endometrial cancer tissue; influence of aromatase on proliferation of endometrial cancer cells. J. Steroid Biochem. Mol. Biol. 1993, 44, 463–468. [Google Scholar] [CrossRef]
- Purohit, A.; Woo, L.W.; Chander, S.K.; Newman, S.P.; Ireson, C.; Ho, Y.; Grasso, A.; Leese, M.P.; Potter, B.V.; Reed, M.J. Steroid sulphatase inhibitors for breast cancer therapy. J. Steroid Biochem. Mol. Biol. 2003, 86, 423–432. [Google Scholar] [CrossRef]
- Winum, J.Y.; Scozzafava, A.; Montero, J.L.; Supuran, C.T. Therapeutic applications of sulfamates. Expert Opin. Ther. Patents 2004, 14, 1273–1308. [Google Scholar] [CrossRef]
- Scozzafava, A.; Owa, T.; Mastrolorenzo, A.; Supuran, C.T. Anticancer and antiviral sulfonamides. Curr. Med. Chem. 2003, 70, 925–953. [Google Scholar] [CrossRef] [PubMed]
- Nussbaumer, P.; Billich, A. Steroid sulfatase inhibitors. Med. Res. Rev. 2004, 24, 529–576. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Cho, M.; Karlsberg, K.; Zhou, D.; Yuan, Y.C. Biochemical and biological characterization of a novel anti-aromatase coumarin derivative. J. Biol. Chem. 2004, 279, 48071–48078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, D.; Liehr, J.G. Estrogen, DNA damage and mutations. Mutat. Res. 1999, 424, 107–115. [Google Scholar] [CrossRef]
- Liehr, J.G. Genotoxicity of the steroidal oestrogens oestrone and oestradiol: Possible mechanism of uterine and mammary cancer development. Hum. Reprod. Update 2001, 7, 273–281. [Google Scholar] [CrossRef] [Green Version]
- Russo, J.; Russo, I.H. Genotoxicity of steroidal estrogens. Trends Endocrinol. Metab. 2004, 15, 211–214. [Google Scholar] [CrossRef]
- Cuzick, J.; Sestak, I.; Forbes, J.F.; Dowsett, M.; Knox, J.; Cawthorn, S.; Saunders, C.; Roche, N.; Mansel, R.E.; von Minckwitz, G.; et al. IBIS-II investigators, Anastrozole for prevention of breast cancer in highrisk postmenopausal women (IBIS-II): An international, double-blind, randomised placebocontrolled trial. Lancet 2014, 383, 1041–1048. [Google Scholar] [CrossRef]
- Leonetti, F.; Favia, A.; Rao, A.; Aliano, R.; Paluszcak, A.; Hartmann, R.W.; Carotti, A. Design, synthesis, and 3D QSAR of novel potent and selective aromatase inhibitors. J. Med. Chem. 2004, 47, 6792–6803. [Google Scholar] [CrossRef] [PubMed]
- Recanatini, M.; Cavalli, A. Comparative molecular field analysis of non-steroidal aromatase inhibitors: An extended model for two different structural classes. Bioorg. Med. Chem. 1998, 6, 377–388. [Google Scholar] [CrossRef]
- Gotteland, M.; May, E.; May-Levin, F.; Contesso, G.; Delarue, J.C.; Mouriesse, H. Estrogen receptors (ER) in human breast cancer. The significance of a new prognostic factor based on both ER protein and ER mRNA contents. Cancer 1994, 74, 864–871. [Google Scholar] [CrossRef]
- Soubeyran, I.; Quenel, N.; Mauriac, L.; Durand, M.; Bonichon, F.; Coindre, J.M. Variation of hormonal receptor, pS2, c-erbB-2 and GSTpi contents in breast carcinomas under tamoxifen: A study of 74 cases. Br. J. Cancer 1996, 73, 735–743. [Google Scholar] [CrossRef] [Green Version]
- Devraj, R.; Barrett, J.F.; Fernandez, J.A.; Katzenellenbogen, J.A.; Cushman, M. Design, synthesis, and biological evaluation of ellipticine-estradiol conjugates. J. Med. Chem. 1996, 39, 3367–3374. [Google Scholar] [CrossRef] [PubMed]
- Upeslacis, J.; Hinman, L. Chemical modification of antibodies for cancer chemotherapy. Annu. Rep. Med. Chem. 1988, 23, 151–160. [Google Scholar]
- Blatter, W.A.; Lambert, J.M.; Goldmacher, V.S. Realizing the full potential of immunotoxins. Cancer Cells 1989, 1, 50–55. [Google Scholar]
- Krohn, K.; Kulikowski, K.; Leclercq, G. Diethylstilbestrol-linked cytotoxic agents: Synthesis and binding affinity for estrogen receptors. J. Med. Chem. 1989, 32, 1532–1538. [Google Scholar] [CrossRef]
- Schmidt, B.F.; Hernandez, L.; Rouzer, C.; Czerwinski, G.; Chmurny, G.; Michejda, C.J. Peptide-linked 1,3-dialkyl-3-acyltriazenes: Gastrin receptor directed antineoplastic alkylating agents. J. Med. Chem. 1994, 37, 3812–3818. [Google Scholar] [CrossRef]
- Varga, J.M.; Asato, N.; Lande, S.; Lerner, A.B. Melanotropin-daunomycin conjugate shows receptor-mediated cytotoxicity in cultured murine melanoma cells. Nature 1977, 267, 56–58. [Google Scholar] [CrossRef]
- Nakagawa-Goto, K.; Yamada, K.; Nakamura, S.; Chen, T.H.; Chiang, P.C.; Bastow, K.F.; Wang, S.C.; Spohn, B.; Hung, M.C.; Lee, F.Y.; et al. Antitumor agents. 258. Syntheses and evaluation of dietary antioxidant--taxoid conjugates as novel cytotoxic agents. Bioorg. Med. Chem. Lett. 2007, 17, 5204–5209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, N.; Dubuc, C.; Rousseau, J.; Benard, F.; van Lier, J.E. Synthesis, characterization, and estrogen receptor binding affinity of flavone-, indole-, and furan-estradiol conjugates. Bioorg. Med. Chem. Lett. 2007, 17, 3212–3216. [Google Scholar] [CrossRef] [PubMed]
- Ali, H.; Ahmed, N.; Tessier, G.; van Lier, J.E. Synthesis and biological activities of nucleoside-estradiol conjugates. Bioorg. Med. Chem. Lett. 2006, 16, 317–319. [Google Scholar] [CrossRef] [PubMed]
- James, D.A.; Swamy, N.; Paz, N.; Hanson, R.N.; Ray, R. Synthesis and estrogen receptor binding affinity of a porphyrin-estradiol conjugate for targeted photodynamic therapy of cancer. Bioorg. Med. Chem. Lett. 1999, 9, 2379–2384. [Google Scholar] [CrossRef]
- Swamy, N.; James, D.A.; Mohr, S.C.; Hanson, R.N.; Ray, R. An estradiol-porphyrin conjugate selectively localizes into estrogen receptor-positive breast cancer cells. Bioorg. Med. Chem. 2002, 10, 3237–3243. [Google Scholar] [CrossRef]
- Liu, C.; Strobl, J.S.; Bane, S.; Schilling, J.K.; McCracken, M.; Chatterjee, S.K.; Rahim-Bata, R.; Kingston, D.G. Design, synthesis, and bioactivities of steroid-linked taxol analogues as potential targeted drugs for prostate and breast cancer. J. Nat. Prod. 2004, 67, 152–159. [Google Scholar] [CrossRef]
- Musa, M.A.; Cooperwood, J.S.; Khan, M.O.F. A Review of Coumarin Derivatives in Pharmacotherapy of Breast Cancer. Curr. Med. Chem. 2008, 15, 2664–2679. [Google Scholar] [CrossRef] [Green Version]
- Cui, N.; Lin, D.-D.; Shen, Y.; Shi, J.-G.; Wang, B.; Zhao, M.-Z.; Zheng, L.; Chen, H.; Shi, J.-H. Triphenylethylene-Coumarin Hybrid TCH-5c Suppresses Tumorigenic Progression in Breast Cancer Mainly Through the Inhibition of Angiogenesis. Anti. Cancer Agents Med. Chem. 2019, 19, 1253–1261. [Google Scholar] [CrossRef]
- Cooke, D. Studies on the Mode of Action of Coumarins (Coumarin, 6-Hydroxycoumarin, 7-Hydroxycoumarin and Esculetin) at a Cellular Level. Ph.D. Thesis, Dublin City University, Dublin, Ireland, 1999. [Google Scholar]
- Wang, C.J.; Hsieh, Y.J.; Chu, C.Y.; Lin, Y.L.; Tseng, T.H. Inhibition of Cell Cycle Progression in Human Leukemic HL-60 Cells by Esculetin. Cancer Lett. 2002, 160, 107–113. [Google Scholar]
- Jimenez-Orozco, F.; Lopez-Gonzalez, J.S.; Nieto-Rodriguez, A.; Velasco-Velazquez, M.A.; Molina-Guarneros, J.A.; Mendoza-Patino, N.; García-Mondragón, M.J.; Elizalde-Galvan, P.; León-Cedeño, F.; Mandoki, J.J. Decrease of Cyclin D1 in the Human Lung Adenocarcinoma Cell Line A-427 by 7-hydroxycoumarin. Lung Cancer 2001, 34, 185–194. [Google Scholar] [CrossRef]
- Mojarrab, M.; Emami, S.A.; Delazar, A.; Tayarani-Najaran, Z. Cytotoxic Properties of Three Isolated Coumarin-hemiterpene Ether Derivatives from Artemisia armeniaca Lam. IJPR 2017, 16, 221–229. [Google Scholar] [PubMed]
- Rigel, D.S. The incidence of malignant melanoma in the United States: Issues as we approach the 21th century. J. Am. Acad. Dermatol. 1996, 77, 533–537. [Google Scholar] [CrossRef]
- Godar, D.E. Worldwide increasing incidences of cutaneous malignant melanoma. J. Skin Cancer 2011, 858425, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, B.K.; Kricker, A. How much melanoma is caused by sun exposure? Melanoma Res. 1993, 3, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Wcislo, G.; Szczylik, C. In process citation. Pol. Merkuriusz. Lek. 2003, 15, 5–8. [Google Scholar]
- Green, A.; Trichopoulos, D. Skin cancer. In Textbook of Cancer Epidemiology; Adami, H.O., Hunter, D., Trichopoulos, D., Eds.; Oxford University Press: New York, NY, USA, 2002; pp. 281–300. [Google Scholar]
- Balch, C.M. Cutaneous Melanoma, 3rd ed.; Quality Medical Publishing Inc: St. Louis, MO, USA, 1998. [Google Scholar]
- Thornes, R.D.; Edlow, D.W., Jr.; Wood, S. Inhibition of Locomotion in Cancer Cells In Vivo By Anticoagulant Therapy. 1. Effect of Sodium Warfarin on V2 Cancer Cells, Granulocytes, Lymphocytes and Macrophages in Rabbits. Johns Hopkins Med. J. 1968, 123, 305–316. [Google Scholar]
- Thornes, R.D. Clinical and Biological Observations Associated with Coumarins. Coumarins: Biology, Applications and Mode of Action; O’Kennedy, R., Thornes, R.D., Eds.; John Wiley & Sons: Chichester, UK, 1997; pp. 255–265. [Google Scholar]
- Velasco-Velazquez, M.A.; Agramonte-Hevia, J.; Barrera, D.; Jimenez- Orozco, A.; Garcia-Mondragon, M.J.; Mendoza-Patino, N.; Landa, A.; Mandoki, J. 4-Hydroxycoumarin Disorganizes the Actin Cytoskeleton in B16-F10 Melanoma Cells but not in B82 Fibroblasts, Decreasing their Adhesion to Extracellular Matrix Proteins and Motility. Cancer Lett. 2003, 198, 179–186. [Google Scholar] [CrossRef]
- Hollingsworth, J.M.; Miller, D.C.; Daignault, S.; Hollenbeck, B.K. Five-year survival after surgical treatment for kidney cancer: A population-based competing risk analysis. Cancer 2007, 109, 1763–1768. [Google Scholar] [CrossRef] [Green Version]
- Russo, P.; O’Brien, M.F. Surgical intervention in patients with metastatic renal cell carcinoma: Metastasectomy and cytoreductive nephrectomy. Urol. Clin. N. Am. 2008, 35, 679–686. [Google Scholar] [CrossRef]
- Hoffman, N.E.; Gillet, M.D.; Cheville, J.C. Difference in organ system of distant metastses by mRCC subtypes. J. Urol. 2008, 180, 474–477. [Google Scholar] [CrossRef]
- Ebbinghaus, S.W.; Mohler, J.L.; Marshall, M.E. Renal Cell Carcinoma: The background, Rationale and Current Development of Coumarin (1,2-benzopyrone) as a Potential Therapeutic Agent. Coumarins: Biology, Applications and Mode of Action; O’Kennedy, R., Thornes, R.D., Eds.; John Wiley & Sons: Chichester, UK, 1997; pp. 209–239. [Google Scholar]
- Mickisch, G.H. Urologic approaches to metastatic renal cell carcinoma. Onkologie 2001, 24, 122–126. [Google Scholar] [CrossRef]
- Vuky, J.; Motzer, R.J. Cytokine Therapy in Renal Cell Cancer. Urol. Oncol. 2000, 5, 249–257. [Google Scholar] [CrossRef]
- Motzer, R.J.; Mazumdar, M.; Bacik, J.; Russo, P.; Berg, W.J.; Metz, E.M. Effect of cytokine therapy on survival for patients with advanced renal cell carcinoma. J. Clin. Oncol. 2000, 18, 1928–1935. [Google Scholar] [CrossRef] [PubMed]
- Rini, B.I.; Atkins, M.B. Resistance to targeted therapy in renal-cell carcinoma. Lancet Oncol. 2009, 10, 992–1000. [Google Scholar] [CrossRef]
- Thornes, R.D.; Lynch, G.; Sheehan, M.W. Cimetidine and Coumarin Therapy of Melanoma. Lancet 1982, 2, 328. [Google Scholar] [CrossRef]
- Dexeus, F.H.; Logothetis, C.J.; Sella, A.; Fitz, K.; Amato, R.; Reuben, J.M.; Dozier, N. Phase II Study of Coumarin and Cimetidine in Patients with Metastatic Renal-Cell Carcinoma. J. Clin. Oncol. 1990, 8, 325–329. [Google Scholar] [CrossRef] [PubMed]
- Kokron, O.; Maca, S.; Gasser, G.; Schmidt, P.R. Cimetidine and Coumarin Therapy of Renal Cell Carcinoma. Oncology 1991, 48, 102–106. [Google Scholar] [CrossRef]
- Marshall, M.E.; Butler, K.; Fried, A. Phase I Evaluation of Coumarin (1,2-Benzopyrone) and Cimetidine in Patients with Advanced Malignancies. Mol. Biother. 1991, 3, 170–178. [Google Scholar]
- Kattimani, P.P.; Kamble, R.R.; Kariduraganavar, M.Y.; Dorababu, A.; Hunnur, R.K. Synthesis, characterization and in vitro anticancer evaluation of novel 1,2,4-triazolin-3-one derivatives. Eur. J. Med. Chem. 2013, 62, 232–240. [Google Scholar] [CrossRef]
- Champelovier, P.; Barbier, P.; Daras, E.; Douillard, S.; Toussaint, B.; Persoon, V.; Combes, S. The indolylcoumarin COUFIN exhibits potent activity against renal carcinoma cells without affecting hematopoietic system. Anti. Cancer Agents Med. Chem. 2014, 14, 862–871. [Google Scholar] [CrossRef]
- Bosland, M.C. Prostate Cancer, Encyclopaedia of Human Biology. San Diego Acad. Press 1991, 6, 177–190. [Google Scholar]
- Agarwal, R. Cell Signalling and Regulators of Cell Cycle as Molecular Targets for Prostate Cancer Prevention by Dietary Agents. Biochem. Pharmacol. 2000, 60, 1051–1059. [Google Scholar] [CrossRef]
- Taniguchi, K.; Funasaki, M.; Kishida, A.; Sadhu, S.K.; Ahmed, F.; Ishibashi, M.; Ohsaki, A. Two new coumarins and a new xanthone from the leaves of Rhizophora mucronate. Bioorg. Med. Chem. Lett. 2018, 28, 1063–1106. [Google Scholar] [CrossRef] [PubMed]
- Jantamat, P.; Weerapreeyakul, N.; Puthongking, P. Cytotoxicity and Apoptosis Induction of Coumarins and Carbazole Alkaloids from Clausena harmandiana. Molecules 2019, 24, 3385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Hua, J.; Zhou, J.; Zhang, H.; Zhu, H.; Cheng, Y.; Gust, R. Synthesis and in vitro antitumor activity of novel scopoletin derivatives. Bioorg. Med. Chem. Lett. 2012, 22, 5008–5012. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lv, Z.; Zhong, H.; Geng, D.; Zhang, M.; Zhang, T.; Li, Y.; Li, K. Convenient synthesis of novel geiparvarin analogs with potential anti-cancer activity via click chemistry. Eur. J. Med. Chem. 2012, 53, 356–363. [Google Scholar] [CrossRef]
- Zhu, J.J.; Jiang, J.G. Pharmacological and nutritional effects of natural coumarins and their structure–activity relationships. Mol. Nutr. Food Res. 2018, 62, 1701073. [Google Scholar] [CrossRef]
- Jain, P.K.; Joshi, H. Coumarin: Chemical and pharmacological profile. J. Appl. Pharm. Sci. 2012, 2, 236–240. [Google Scholar]
- Lake, B. Coumarin Metabolism, Toxicity and Carcinogenicity: Relevance for Human Risk Assessment. Food Chem. Tox. 1999, 37, 423–453. [Google Scholar] [CrossRef]
- Ritschel, W.A.; Grummich, K.W.; Kaul, S.; Hardt, T.J. Biopharmaceutical Parameters of Coumarin and 7- hydroxycoumarin. Die Pharma. Ind. 1981, 43, 271–276. [Google Scholar]
- Egan, D.; O’Kennedy, R. Spectrofluorimetric Method for the Quantification of 7-Hydroxycoumarin in Urine and Plasma Using Both Extracted and Unextracted Samples. Analyst 1993, 118, 201–203. [Google Scholar] [CrossRef] [PubMed]
- Dempsey, E.; O’Sullivan, C.; Smyth, M.R.; Egan, D.; O’Kennedy, R.; Wang, J. Development of an Electrochemical Biosensor for 7-hydroxycoumarin. Analyst 1993, 118, 411–414. [Google Scholar] [CrossRef] [PubMed]
- Keating, G. Biosensor-Based Studies on Coumarins. Ph.D. Thesis, Dublin City University, Dublin, Ireland, 1998. [Google Scholar]
- Pelkonen, O.; Rautio, A.; Raunio, H.; Pasanen, M. CYP2A6: A Human Coumarin 7-hydroxylase. Toxicology 2000, 144, 139–147. [Google Scholar] [CrossRef]
- Pelkonen, O.; Maenpaa, J.; Taavitsainen, P.; Rautio, A.; Raunio, H. Inhibition and Induction of Human Cytochrome P450 (CYP) Enzymes. Xenobiotica 1998, 28, 1203–1253. [Google Scholar] [CrossRef] [PubMed]
- Pelkonen, O.; Raunio, H.; Rautio, A.; Pasanen, M.; Lang, M.A. The Metabolism of Coumarin. Coumarins: Biology, Applications and Mode of Action; O’Kennedy, R., Thornes, R.D., Eds.; John Wiley & Sons: Chichester, UK, 1997; pp. 67–92. [Google Scholar]
- Taavitsainen, P. Cytochrome P450 Isoform-Specific In Vitro Methods to Predict Drug Metabolism and Interactions; Oulu University Press: Oulu, Finland, 2001. [Google Scholar]
- Bogan, D.; Deasy, B.; O’Kennedy, R.; Smyth, M. Determination of Free and Total 7-hydroxycoumarin in Urine and Serum by Capillary Electrophoresis. J. Chromatogr. B 1995, 663, 371–378. [Google Scholar] [CrossRef]
- Rautio, A.; Kraul, H.; Kojo, A.; Salmela, E.; Pelkonen, O. Interindividual Variability in Coumarin 7-Hydroxylation in Healthy Individuals. Pharmacogenetics 1992, 2, 227–233. [Google Scholar] [CrossRef]
- Egan, D.; O’Kennedy, R. The Production and Characterisation of Anti-7-Hydroxycoumarin Antibodies and Their Use in the Development of an Enzyme-Linked Immunosorbent Assay. J. Ir. Coll. Phys. Surg. 1993, 22, 72. [Google Scholar]
- Bogan, D.; Deasy, B.; O’Kennedy, R.; Smyth, M. The Use of Capillary Electrophoresis for Studying Interspecies Differences in Coumarin Metabolism in Liver Microsomes. Xenobiotica 1996, 26, 437–448. [Google Scholar] [CrossRef]
- Pineo, G.F.; Hull, R.D. Adverse effects of coumarin anticoagulants. Drug Saf. 1993, 9, 263–271. [Google Scholar] [CrossRef]
- Aronson, J.K. Coumarin Anticoagulants. Meyler’s Side Effects of Drugs; Elsevier Science: Amsterdam, The Netherlands, 2016; pp. 702–737. [Google Scholar]
- Hart, J.P.; Catterall, A.; Dodds, R.A.; Klenerman, L.; Shearer, M.J.; Bitensky, L.; Chayen, J. Circulating vitamin K1 levels in fractured neck of femur. Lancet 1984, 2, 283. [Google Scholar] [CrossRef]
- Ginsberg, J.S.; Hirsh, J. Use of antithrombotic drugs during pregnancy. Chest 1992, 102, 385S–390S. [Google Scholar] [CrossRef] [PubMed]
- Byrden, T. The Elucidation of the Mode of Action of Coumarin and Some Specified Derivatives. Master’s Thesis, Dublin City University, Dublin, Ireland, 1995. [Google Scholar]
- Egan, D.; O’Kennedy, R.; Moran, E.; Cox, D.; Prosser, E.; Thornes, R.D. The Pharmacology, Metabolism, Analysis and Applications of Coumarin and Coumarin-Related Compounds. Drug Metab. Rev. 1990, 22, 503–529. [Google Scholar] [CrossRef] [PubMed]
- Deasy, B. Development of Novel Analytical Methods to Study the Metabolism of Coumarin. Ph.D. Thesis, Dublin City University, Dublin, Ireland, 1996. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Demographic Features | Gender, Age, Race/Ethnicity |
|---|---|
| Reproductive story | The age of menarche, number of birth, first full-term pregnancy age, menopause age, lactation, infertility, miscarriage |
| Familial/genetic factors | Family history, known or suspected BRCA1/2, p53, PTEN, or other gene mutations related to breast cancer risk |
| Environmental factors | Radiotherapy to the thorax before 30 years of age, hormone replacement therapy, alcohol use, socioeconomic level, etc. |
| Other factors | Personal history of breast cancer, number of breast biopsies, atypical hyperplasia or lobular carcinoma in situ, dense breast structure, body mass index (BMI) |
| Physical conditions | Light skin color, blond or red hair, light eyes, freckles, or skin burns easily in the sun Sunburn history (formation of one or more severe, vesicle-forming sunburn) Having many or unusual moles |
| Familial/genetic factors | Family history of melanoma People diagnosed with melanoma in close relatives such as parents, children or siblings |
| Environmental factors | Life close to the equator or at high altitude Exposure to UV rays from the sun or solarium |
| Weakened immune system | People with weakened immunity (e.g., those with organ transplants) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Küpeli Akkol, E.; Genç, Y.; Karpuz, B.; Sobarzo-Sánchez, E.; Capasso, R. Coumarins and Coumarin-Related Compounds in Pharmacotherapy of Cancer. Cancers 2020, 12, 1959. https://doi.org/10.3390/cancers12071959
Küpeli Akkol E, Genç Y, Karpuz B, Sobarzo-Sánchez E, Capasso R. Coumarins and Coumarin-Related Compounds in Pharmacotherapy of Cancer. Cancers. 2020; 12(7):1959. https://doi.org/10.3390/cancers12071959
Chicago/Turabian StyleKüpeli Akkol, Esra, Yasin Genç, Büşra Karpuz, Eduardo Sobarzo-Sánchez, and Raffaele Capasso. 2020. "Coumarins and Coumarin-Related Compounds in Pharmacotherapy of Cancer" Cancers 12, no. 7: 1959. https://doi.org/10.3390/cancers12071959