The Biological Function of Sigma-2 Receptor/TMEM97 and Its Utility in PET Imaging Studies in Cancer


Abstract
:1. Introduction
2. Molecular Cloning of the σ2R
3. Evaluation of the σ2R PET Radiotracer for Assessing Proliferative Status and Predicting Drug Response
3.1. Evaluation of σ2R PET Radiotracer for Assessing the Proliferative Status and Growth Rate of Solid Tumors
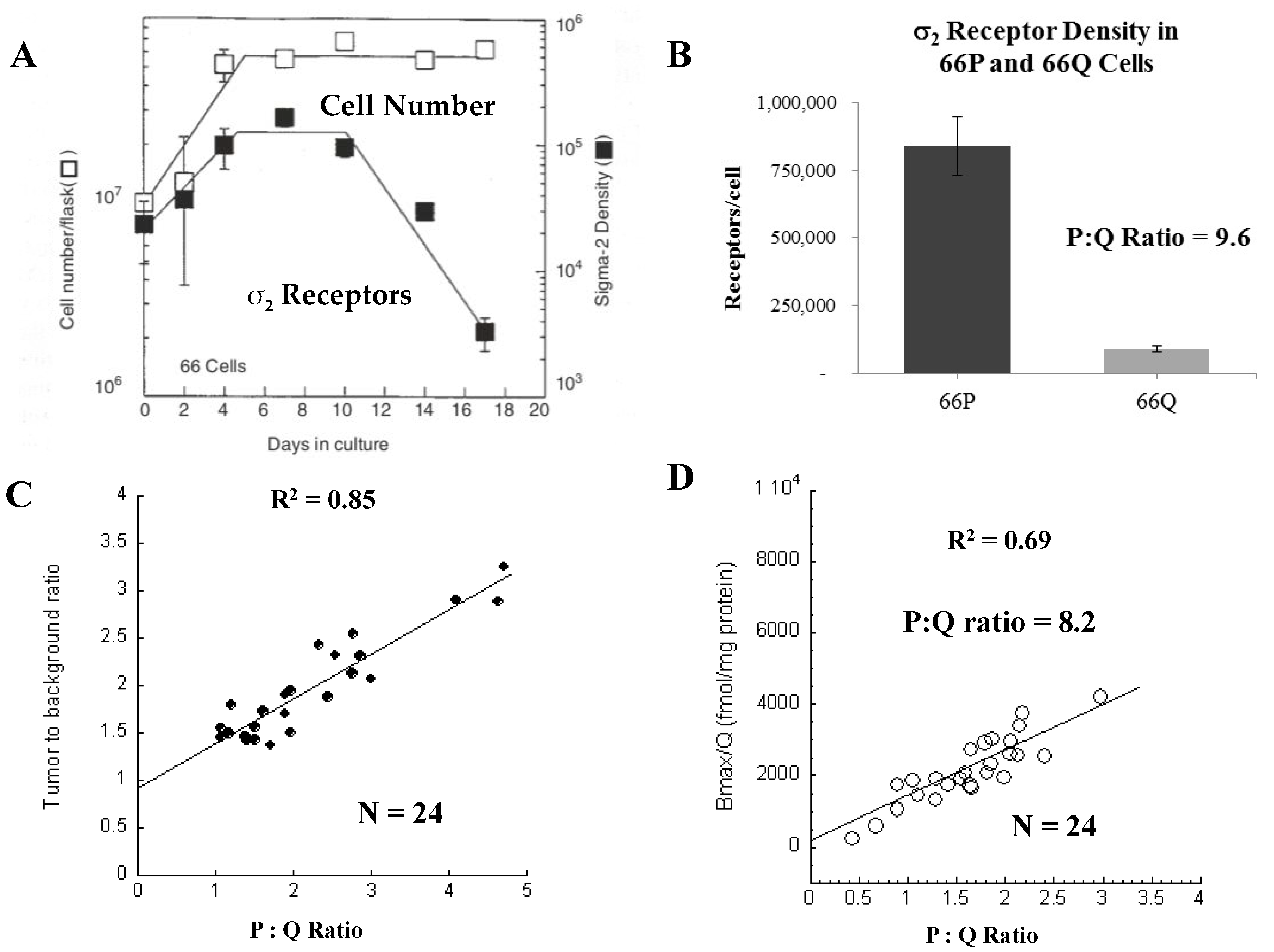
3.1.1. Evaluation of the σ2R as a Proliferation Biomarker
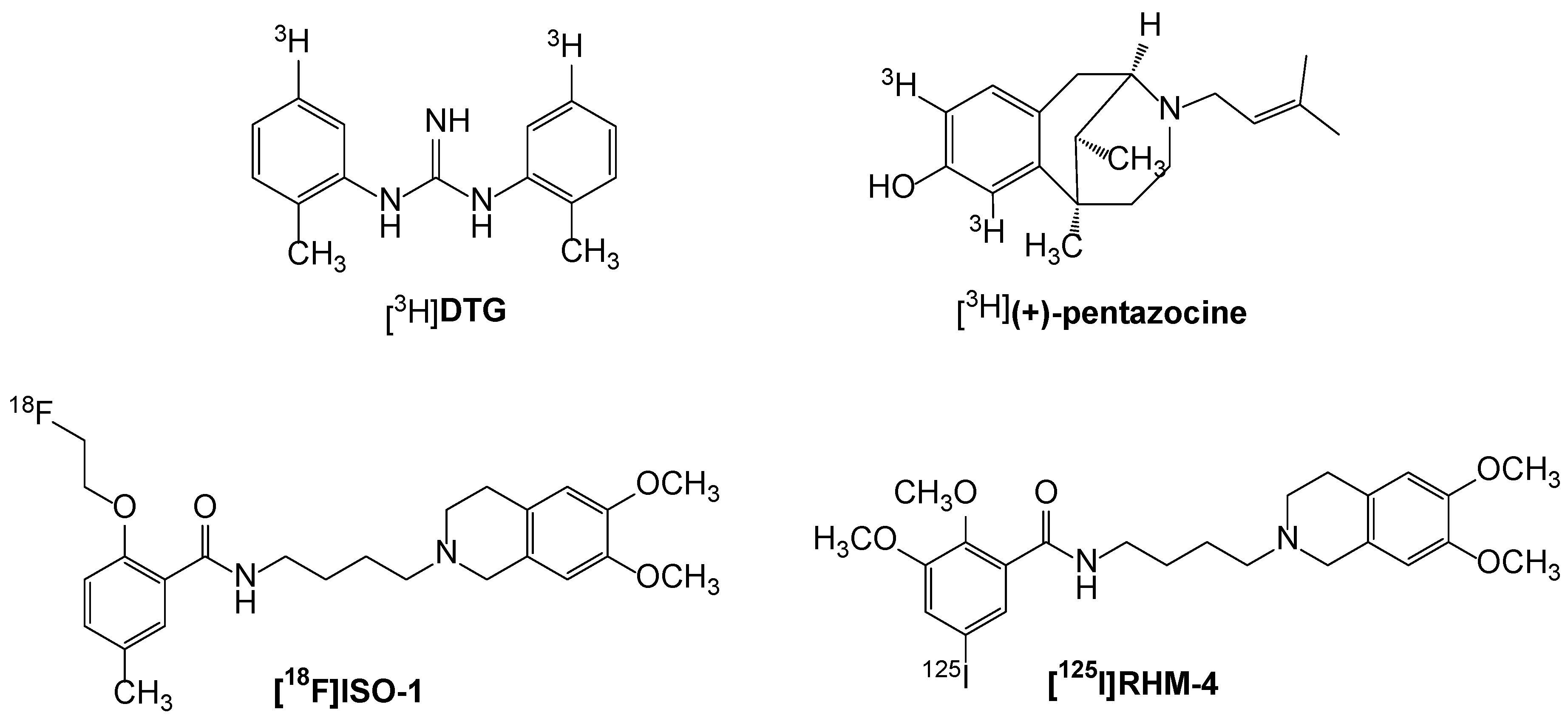
3.1.2. Development of σ2R Radioligands
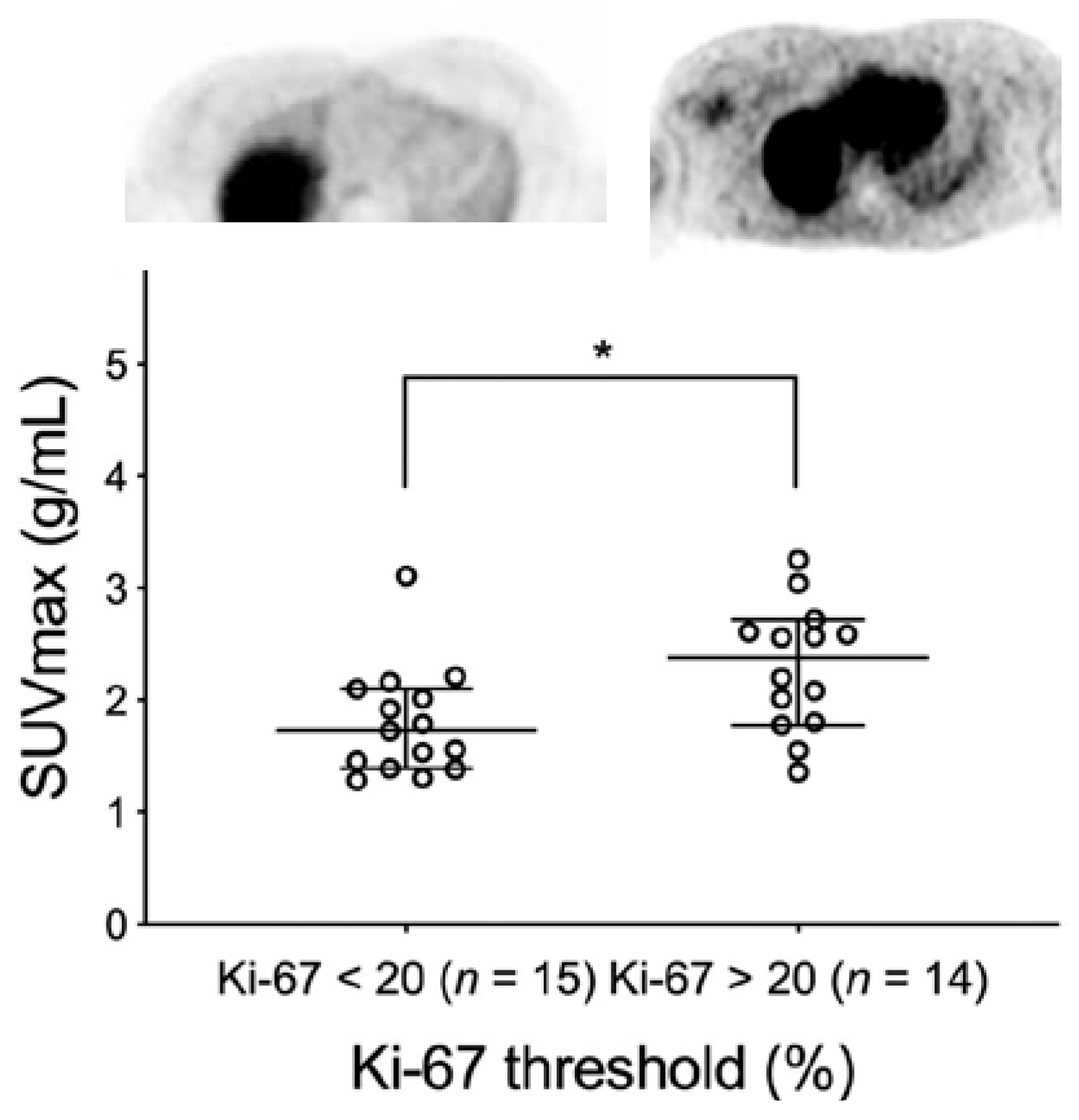
3.1.3. Clinical Studies with [18F]ISO-1
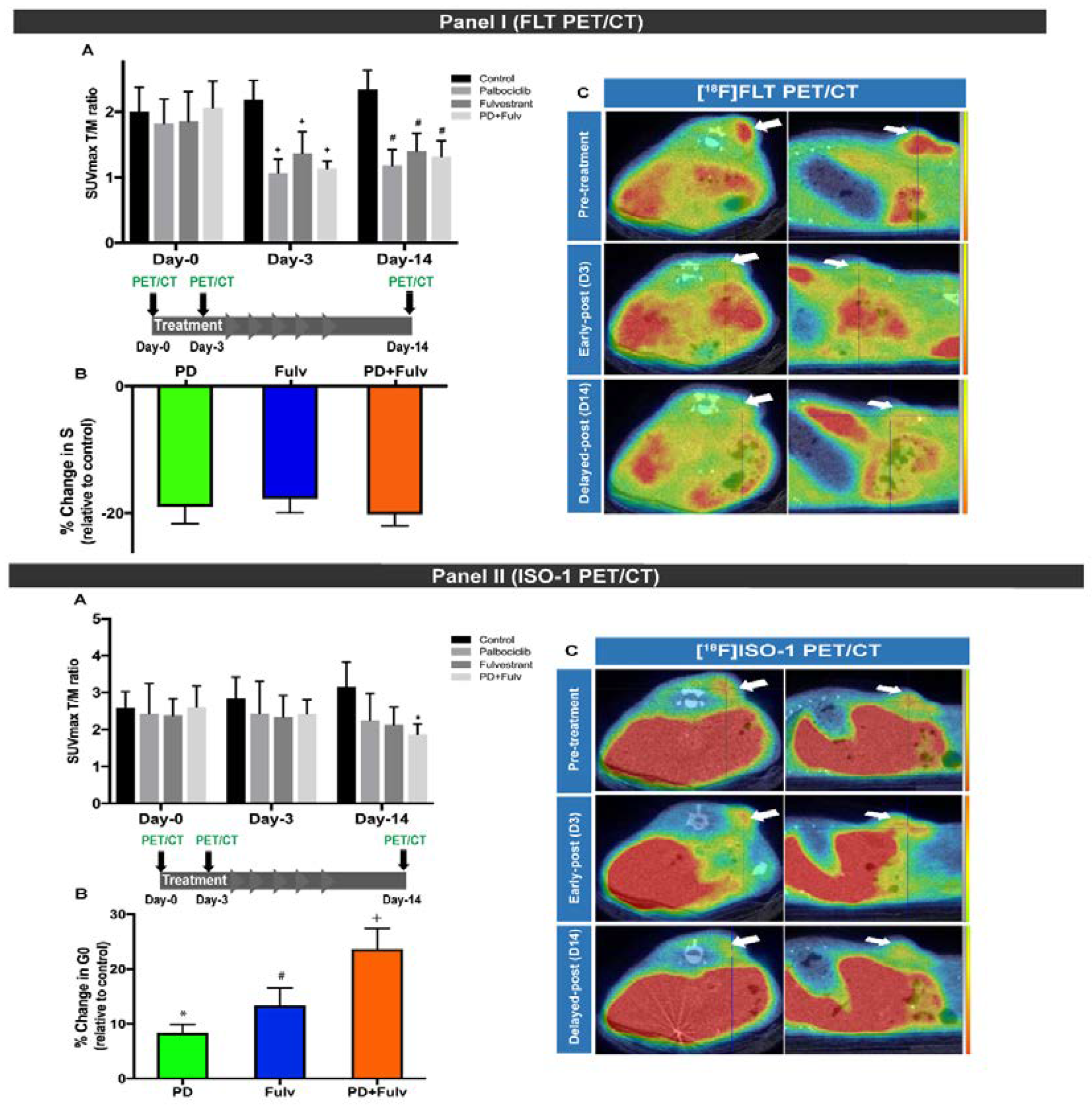
3.2. Evaluation of [18F]ISO-1 for the Monitoring and Predicting Response to Therapy
4. Biological Characteristics and Functions of σ2R/TMEM97
4.1. Subcellular Localization of σ2R/TMEM97
4.2. Biological Characterization and Roles of TMEM97 in Cancer Proliferation
4.2.1. Differential Expression of TMEM97 in Normal and Cancer Tissues
4.2.2. Potential Requirement of TMEM97 in Cancer Proliferation
4.2.3. Regulation of TMEM97 Expression in Cancer
4.2.4. TMEM97 Does Not Mediate σ2R Ligand Cytotoxicity
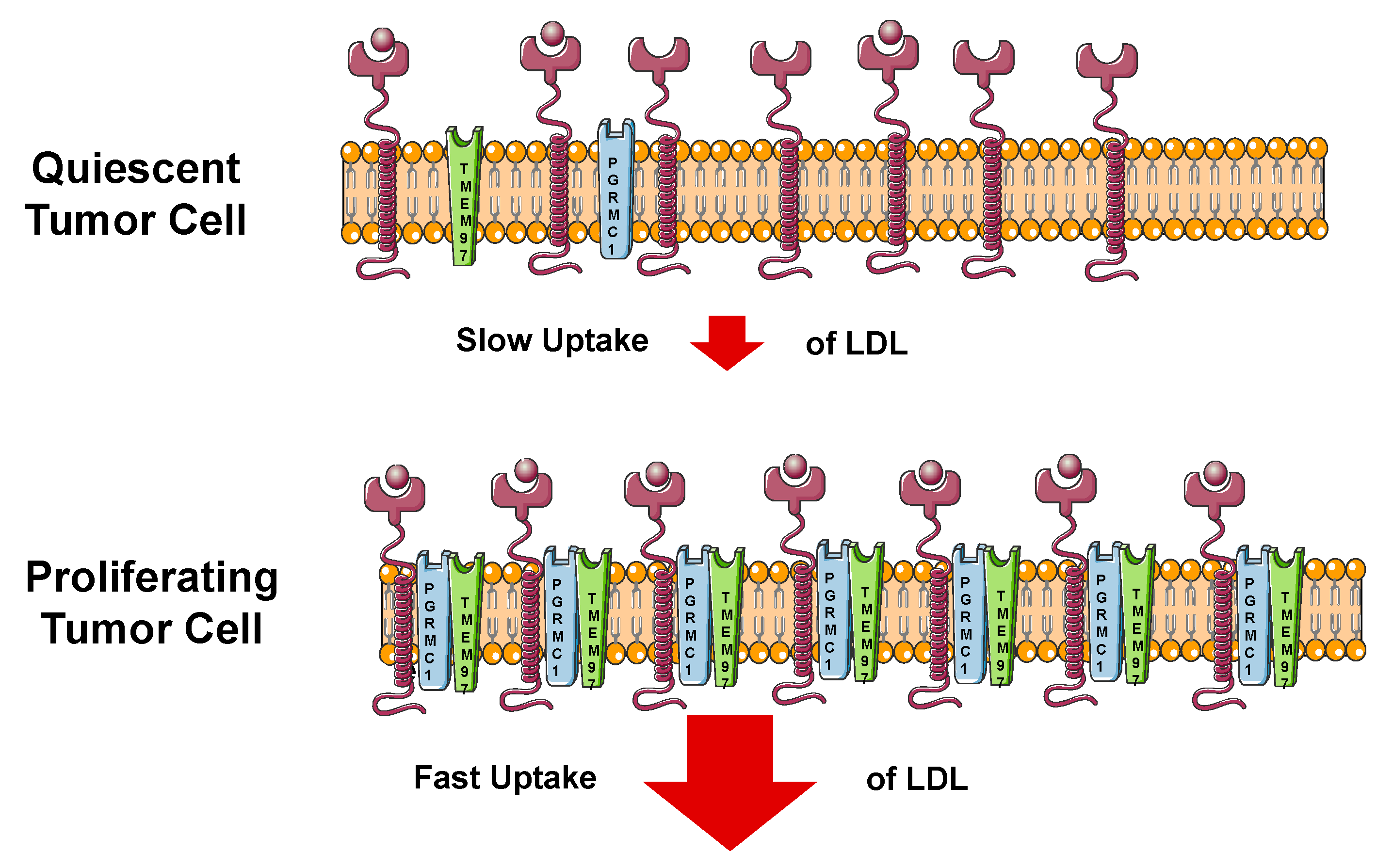
4.3. Biological Roles of TMEM97 in Cholesterol Homeostasis
4.4. TMEM97 in the Interplay of Cancer and Cholesterol Homeostasis
5. Conclusions
Funding
Conflicts of Interest
References
- Martin, W.R.; Eades, C.G.; Thompson, J.A.; Huppler, R.E.; Gilbert, P.E. The effects of morphine- and nalorphine-like drugs in the nondependent and morphine-dependent chronic spinal dog. J. Pharmacol. Exp. Ther. 1976, 197, 517–532. [Google Scholar]
- Matsumoto, R.R. σ Receptors: Historical perspective and background. In Sigma Receptors: Chemistry, Cell Biology and Clinical Implications; Su, T.-P., Matsumoto, R.R., Bowen, W.D., Eds.; Springer: Boston, MA, USA, 2007; pp. 1–23. [Google Scholar]
- Hellewell, S.B.; Bowen, W.D. A sigma-like binding site in rat pheochromocytoma (PC12) cells: Decreased affinity for (+)-benzomorphans and lower molecular weight suggest a different sigma receptor form from that of guinea pig brain. Brain Res. 1990, 527, 244–253. [Google Scholar] [CrossRef]
- Hellewell, S.B.; Bruce, A.; Feinstein, G.; Orringer, J.; Williams, W.; Bowen, W.D. Rat liver and kidney contain high densities of sigma 1 and sigma 2 receptors: Characterization by ligand binding and photoaffinity labeling. Eur. J. Pharmacol. 1994, 268, 9–18. [Google Scholar] [CrossRef]
- Hanner, M.; Moebius, F.F.; Flandorfer, A.; Knaus, H.G.; Striessnig, J.; Kempner, E.; Glossmann, H. Purification, molecular cloning, and expression of the mammalian sigma1-binding site. Proc. Natl. Acad. Sci. USA 1996, 93, 8072–8077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, H.R.; Kruse, A.C. The Molecular Function of sigma Receptors: Past, Present, and Future. Trends Pharmacol. Sci. 2019, 40, 636–654. [Google Scholar] [CrossRef] [PubMed]
- Alon, A.; Schmidt, H.R.; Wood, M.D.; Sahn, J.J.; Martin, S.F.; Kruse, A.C. Identification of the gene that codes for the sigma2 receptor. Proc. Natl. Acad. Sci. USA 2017, 114, 7160–7165. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Zeng, C.; Chu, W.; Pan, F.; Rothfuss, J.M.; Zhang, F.; Tu, Z.; Zhou, D.; Zeng, D.; Vangveravong, S.; et al. Identification of the PGRMC1 protein complex as the putative sigma-2 receptor binding site. Nat. Commun. 2011, 2, 380. [Google Scholar] [CrossRef]
- Chu, U.B.; Mavlyutov, T.A.; Chu, M.L.; Yang, H.; Schulman, A.; Mesangeau, C.; McCurdy, C.R.; Guo, L.W.; Ruoho, A.E. The Sigma-2 Receptor and Progesterone Receptor Membrane Component 1 are Different Binding Sites Derived From Independent Genes. EBioMedicine 2015, 2, 1806–1813. [Google Scholar] [CrossRef] [Green Version]
- Pati, M.L.; Groza, D.; Riganti, C.; Kopecka, J.; Niso, M.; Berardi, F.; Hager, S.; Heffeter, P.; Hirai, M.; Tsugawa, H.; et al. Sigma-2 receptor and progesterone receptor membrane component 1 (PGRMC1) are two different proteins: Proofs by fluorescent labeling and binding of sigma-2 receptor ligands to PGRMC1. Pharmacol. Res. 2017, 117, 67–74. [Google Scholar] [CrossRef]
- Riad, A.; Zeng, C.; Weng, C.C.; Winters, H.; Xu, K.; Makvandi, M.; Metz, T.; Carlin, S.; Mach, R.H. Sigma-2 Receptor/TMEM97 and PGRMC-1 Increase the Rate of Internalization of LDL by LDL Receptor through the Formation of a Ternary Complex. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef]
- Wheeler, K.T.; Wang, L.M.; Wallen, C.A.; Childers, S.R.; Cline, J.M.; Keng, P.C.; Mach, R.H. Sigma-2 receptors as a biomarker of proliferation in solid tumours. Br. J. Cancer 2000, 82, 1223–1232. [Google Scholar] [CrossRef] [PubMed]
- Mach, R.H.; Smith, C.R.; al-Nabulsi, I.; Whirrett, B.R.; Childers, S.R.; Wheeler, K.T. Sigma 2 receptors as potential biomarkers of proliferation in breast cancer. Cancer Res. 1997, 57, 156–161. [Google Scholar] [PubMed]
- Wallen, C.A.; Higashikubo, R.; Dethlefsen, L.A. Murine mammary tumour cells in vitro. I. The development of a quiescent state. Cell Tissue Kinet. 1984, 17, 65–77. [Google Scholar] [CrossRef] [PubMed]
- Wallen, C.A.; Higashikubo, R.; Dethlefsen, L.A. Murine mammary tumour cells in vitro. II. Recruitment of quiescent cells. Cell Tissue Kinet. 1984, 17, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Al-Nabulsi, I.; Mach, R.H.; Wang, L.M.; Wallen, C.A.; Keng, P.C.; Sten, K.; Childers, S.R.; Wheeler, K.T. Effect of ploidy, recruitment, environmental factors, and tamoxifen treatment on the expression of sigma-2 receptors in proliferating and quiescent tumour cells. Br. J. Cancer 1999, 81, 925–933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, C.; McDonald, E.S.; Mach, R.H. Molecular probes for imaging the sigma-2 receptor: In vitro and in vivo imaging studies. In Sigma Proteins: Evolution of the Concept of Sigma Receptors; Kim, F.J., Pasternak, G.W., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 309–330. [Google Scholar]
- Mach, R.H.; Zeng, C.; Hawkins, W.G. The sigma2 receptor: A novel protein for the imaging and treatment of cancer. J. Med. Chem. 2013, 56, 7137–7160. [Google Scholar] [CrossRef] [Green Version]
- Dehdashti, F.; Laforest, R.; Gao, F.; Shoghi, K.I.; Aft, R.L.; Nussenbaum, B.; Kreisel, F.H.; Bartlett, N.L.; Cashen, A.; Wagner-Johnston, N.; et al. Assessment of cellular proliferation in tumors by PET using 18F-ISO-1. J. Nucl. Med. 2013, 54, 350–357. [Google Scholar] [CrossRef] [Green Version]
- McDonald, E.S.; Doot, R.K.; Young, A.J.; Schubert, E.K.; Pryma, D.A.; Farwell, M.D.; Tchou, J.; Nayak, A.; Ziober, A.; Feldman, M.D.; et al. Breast Cancer (18)F-ISO-1 Uptake as a Marker of Proliferation Status. J. Nucl. Med. 2019. [Google Scholar] [CrossRef]
- Shoghi, K.I.; Xu, J.; Su, Y.; He, J.; Rowland, D.; Yan, Y.; Garbow, J.R.; Tu, Z.; Jones, L.A.; Higashikubo, R.; et al. Quantitative receptor-based imaging of tumor proliferation with the sigma-2 ligand [(18)F]ISO-1. PLoS ONE 2013, 8, e74188. [Google Scholar] [CrossRef] [Green Version]
- Loibl, S.; Turner, N.C.; Ro, J.; Cristofanilli, M.; Iwata, H.; Im, S.A.; Masuda, N.; Loi, S.; Andre, F.; Harbeck, N.; et al. Palbociclib Combined with Fulvestrant in Premenopausal Women with Advanced Breast Cancer and Prior Progression on Endocrine Therapy: PALOMA-3 Results. Oncologist 2017, 22, 1028–1038. [Google Scholar] [CrossRef] [Green Version]
- Elmi, A.; Makvandi, M.; Weng, C.C.; Hou, C.; Clark, A.S.; Mach, R.H.; Mankoff, D.A. Cell-Proliferation Imaging for Monitoring Response to CDK4/6 Inhibition Combined with Endocrine-Therapy in Breast Cancer: Comparison of [(18)F]FLT and [(18)F]ISO-1 PET/CT. Clin. Cancer Res. 2019, 25, 3063–3073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vilner, B.J.; John, C.S.; Bowen, W.D. Sigma-1 and sigma-2 receptors are expressed in a wide variety of human and rodent tumor cell lines. Cancer Res. 1995, 55, 408–413. [Google Scholar] [PubMed]
- Gebreselassie, D.; Bowen, W.D. Sigma-2 receptors are specifically localized to lipid rafts in rat liver membranes. Eur. J. Pharmacol. 2004, 493, 19–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, C.; Vangveravong, S.; Xu, J.; Chang, K.C.; Hotchkiss, R.S.; Wheeler, K.T.; Shen, D.; Zhuang, Z.P.; Kung, H.F.; Mach, R.H. Subcellular localization of sigma-2 receptors in breast cancer cells using two-photon and confocal microscopy. Cancer Res. 2007, 67, 6708–6716. [Google Scholar] [CrossRef] [Green Version]
- Kayed, H.; Kleeff, J.; Ding, J.; Hammer, J.; Giese, T.; Zentgraf, H.; Buchler, M.W.; Friess, H. Expression analysis of MAC30 in human pancreatic cancer and tumors of the gastrointestinal tract. Histol. Histopathol. 2004, 19, 1021–1031. [Google Scholar] [CrossRef]
- Zhao, Z.R.; Zhang, L.J.; He, X.Q.; Zhang, Z.Y.; Zhang, F.; Li, F.; Pei, Y.B.; Hu, Y.M.; Wang, M.W.; Sun, X.F. Significance of mRNA and protein expression of MAC30 in progression of colorectal cancer. Chemotherapy 2011, 57, 394–401. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.Y.; Zhao, Z.R.; Adell, G.; Jarlsfelt, I.; Cui, Y.X.; Kayed, H.; Kleeff, J.; Wang, M.W.; Sun, X.F. Expression of MAC30 in rectal cancers with or without preoperative radiotherapy. Oncology 2006, 71, 259–265. [Google Scholar] [CrossRef]
- Moparthi, S.B.; Arbman, G.; Wallin, A.; Kayed, H.; Kleeff, J.; Zentgraf, H.; Sun, X.F. Expression of MAC30 protein is related to survival and biological variables in primary and metastatic colorectal cancers. Int. J. Oncol. 2007, 30, 91–95. [Google Scholar] [CrossRef] [Green Version]
- Lewander, A.; Gao, J.; Adell, G.; Zhang, H.; Sun, X.F. Expression of NF-kappaB p65 phosphorylated at serine-536 in rectal cancer with or without preoperative radiotherapy. Radiol. Oncol. 2011, 45, 279–284. [Google Scholar] [CrossRef] [Green Version]
- Ding, H.; Gui, X.; Lin, X.; Chen, R.; Ma, T.; Sheng, Y.; Cai, H.; Fen, Y. The Prognostic Effect of MAC30 Expression on Patients with Non-Small Cell Lung Cancer Receiving Adjuvant Chemotherapy. Technol. Cancer Res. Treat. 2017, 16, 645–653. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Zhou, F.; Ji, X.; Ren, K.; Shan, Y.; Mao, X.; Fen, Y.; Chen, R.; Ding, H.; Fu, X. The prognostic role of MAC30 in advanced gastric cancer patients receiving platinum-based chemotherapy. Future Oncol. 2017, 13, 2691–2696. [Google Scholar] [CrossRef] [PubMed]
- Xiao, M.; Li, H.; Yang, S.; Huang, Y.; Jia, S.; Wang, H.; Wang, J.; Li, Z. Expression of MAC30 protein is related to survival and clinicopathological variables in breast cancer. J. Surg. Oncol. 2013, 107, 456–462. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Li, H.; Liu, Y.; Ning, X.; Meng, F.; Xiao, M.; Wang, D.; Lou, G.; Zhang, Y. Elevated expression of MAC30 predicts lymph node metastasis and unfavorable prognosis in patients with epithelial ovarian cancer. Med. Oncol. 2013, 30, 324. [Google Scholar] [CrossRef]
- Ramalho-Carvalho, J.; Goncalves, C.S.; Graca, I.; Bidarra, D.; Pereira-Silva, E.; Salta, S.; Godinho, M.I.; Gomez, A.; Esteller, M.; Costa, B.M.; et al. A multiplatform approach identifies miR-152-3p as a common epigenetically regulated onco-suppressor in prostate cancer targeting TMEM97. Clin. Epigenet. 2018, 10, 40. [Google Scholar] [CrossRef] [PubMed]
- Murphy, M.; Pykett, M.J.; Harnish, P.; Zang, K.D.; George, D.L. Identification and characterization of genes differentially expressed in meningiomas. Cell Growth Differ. 1993, 4, 715–722. [Google Scholar]
- Qu, T.; Zhao, Y.; Chen, Y.; Jin, S.; Fang, Y.; Jin, X.; Sun, L.; Ma, Y. Down-regulated MAC30 expression inhibits breast cancer cell invasion and EMT by suppressing Wnt/beta-catenin and PI3K/Akt signaling pathways. Int. J. Clin. Exp. Pathol. 2019, 12, 1888–1896. [Google Scholar]
- Qiu, G.; Sun, W.; Zou, Y.; Cai, Z.; Wang, P.; Lin, X.; Huang, J.; Jiang, L.; Ding, X.; Hu, G. RNA interference against TMEM97 inhibits cell proliferation, migration, and invasion in glioma cells. Tumour. Biol. 2015, 36, 8231–8238. [Google Scholar] [CrossRef]
- Xu, X.Y.; Zhang, L.J.; Yu, Y.Q.; Zhang, X.T.; Huang, W.J.; Nie, X.C.; Song, G.Q. Down-regulated MAC30 expression inhibits proliferation and mobility of human gastric cancer cells. Cell Physiol. Biochem. 2014, 33, 1359–1368. [Google Scholar] [CrossRef]
- Wilcox, C.B.; Feddes, G.O.; Willett-Brozick, J.E.; Hsu, L.C.; DeLoia, J.A.; Baysal, B.E. Coordinate up-regulation of TMEM97 and cholesterol biosynthesis genes in normal ovarian surface epithelial cells treated with progesterone: Implications for pathogenesis of ovarian cancer. BMC Cancer 2007, 7, 223. [Google Scholar] [CrossRef] [Green Version]
- Atalay, A.; Crook, T.; Ozturk, M.; Yulug, I.G. Identification of genes induced by BRCA1 in breast cancer cells. Biochem. Biophys. Res. Commun. 2002, 299, 839–846. [Google Scholar] [CrossRef]
- Maack, S.; Knuechel, R.; Hofstaedter, F.; Stark, A.; Schlegel, J. Detection of differentially expressed genes in human bladder cancer cells using arbitrarily primed PCR of RNA. Int. J. Oncol. 1997, 11, 383–387. [Google Scholar] [CrossRef] [PubMed]
- Zeng, C.; Weng, C.C.; Schneider, M.E.; Puentes, L., Jr.; Riad, A.; Xu, K.; Makvandi, M.; Jin, L.; Hawkins, W.G.; Mach, R.H. TMEM97 and PGRMC1 do not mediate sigma-2 ligand-induced cell death. Cell Death Discov. 2019, 5, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartz, F.; Kern, L.; Erz, D.; Zhu, M.; Gilbert, D.; Meinhof, T.; Wirkner, U.; Erfle, H.; Muckenthaler, M.; Pepperkok, R.; et al. Identification of cholesterol-regulating genes by targeted RNAi screening. Cell Metab. 2009, 10, 63–75. [Google Scholar] [CrossRef] [Green Version]
- Ebrahimi-Fakhari, D.; Wahlster, L.; Bartz, F.; Werenbeck-Ueding, J.; Praggastis, M.; Zhang, J.; Joggerst-Thomalla, B.; Theiss, S.; Grimm, D.; Ory, D.S.; et al. Reduction of TMEM97 increases NPC1 protein levels and restores cholesterol trafficking in Niemann-pick type C1 disease cells. Hum. Mol. Genet. 2016, 25, 3588–3599. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Pulido, L.; Ponting, C.P. TM6SF2 and MAC30, new enzyme homologs in sterol metabolism and common metabolic disease. Front Genet. 2014, 5, 439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabitova, L.; Gorin, A.; Astsaturov, I. Molecular pathways: Sterols and receptor signaling in cancer. Clin. Cancer Res. 2014, 20, 28–34. [Google Scholar] [CrossRef] [Green Version]
- Silvente-Poirot, S.; Poirot, M. Cancer. Cholesterol and cancer, in the balance. Science 2014, 343, 1445–1446. [Google Scholar] [CrossRef]
- Abate, C.; Niso, M.; Berardi, F. Sigma-2 receptor: Past, present and perspectives on multiple therapeutic exploitations. Future Med. Chem. 2018, 10, 1997–2018. [Google Scholar] [CrossRef]
- Oyer, H.M.; Sanders, C.M.; Kim, F.J. Small-Molecule Modulators of Sigma1 and Sigma2/TMEM97 in the Context of Cancer: Foundational Concepts and Emerging Themes. Front. Pharmacol. 2019, 10, 1141. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ligand | Control (µM) Mean ± SD | TMEM97 KO (µM) Mean ± SD | PGRMC1 KO (µM) Mean ± SD | Double KO (µM) Mean ± SD |
|---|---|---|---|---|
| Siramesine | 12.1 ± 1.7 | 10.3 ± 1.49 | 10.7 ± 0.7 | 10.0 ± 1.3 |
| SW43 | 33.7 ± 8.6 | 30.4 ± 3.4 | 33.2 ± 1.6 | 32.8 ± 3.8 |
| PB28 | 58.2 ± 5.9 | 59.8 ± 10.7 | 59.4 ± 8.0 | 57.4 ± 6.9 |
| RHM-4 | >200 | >200 | >200 | >200 |
| ISO-1 | >200 | >200 | >200 | >200 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeng, C.; Riad, A.; Mach, R.H. The Biological Function of Sigma-2 Receptor/TMEM97 and Its Utility in PET Imaging Studies in Cancer. Cancers 2020, 12, 1877. https://doi.org/10.3390/cancers12071877
Zeng C, Riad A, Mach RH. The Biological Function of Sigma-2 Receptor/TMEM97 and Its Utility in PET Imaging Studies in Cancer. Cancers. 2020; 12(7):1877. https://doi.org/10.3390/cancers12071877
Chicago/Turabian StyleZeng, Chenbo, Aladdin Riad, and Robert H. Mach. 2020. "The Biological Function of Sigma-2 Receptor/TMEM97 and Its Utility in PET Imaging Studies in Cancer" Cancers 12, no. 7: 1877. https://doi.org/10.3390/cancers12071877