
Neurotoxicity of Silver Nanoparticles and Non-Linear Development of Adaptive Homeostasis with Age

 ,
,  , and
, and
Abstract
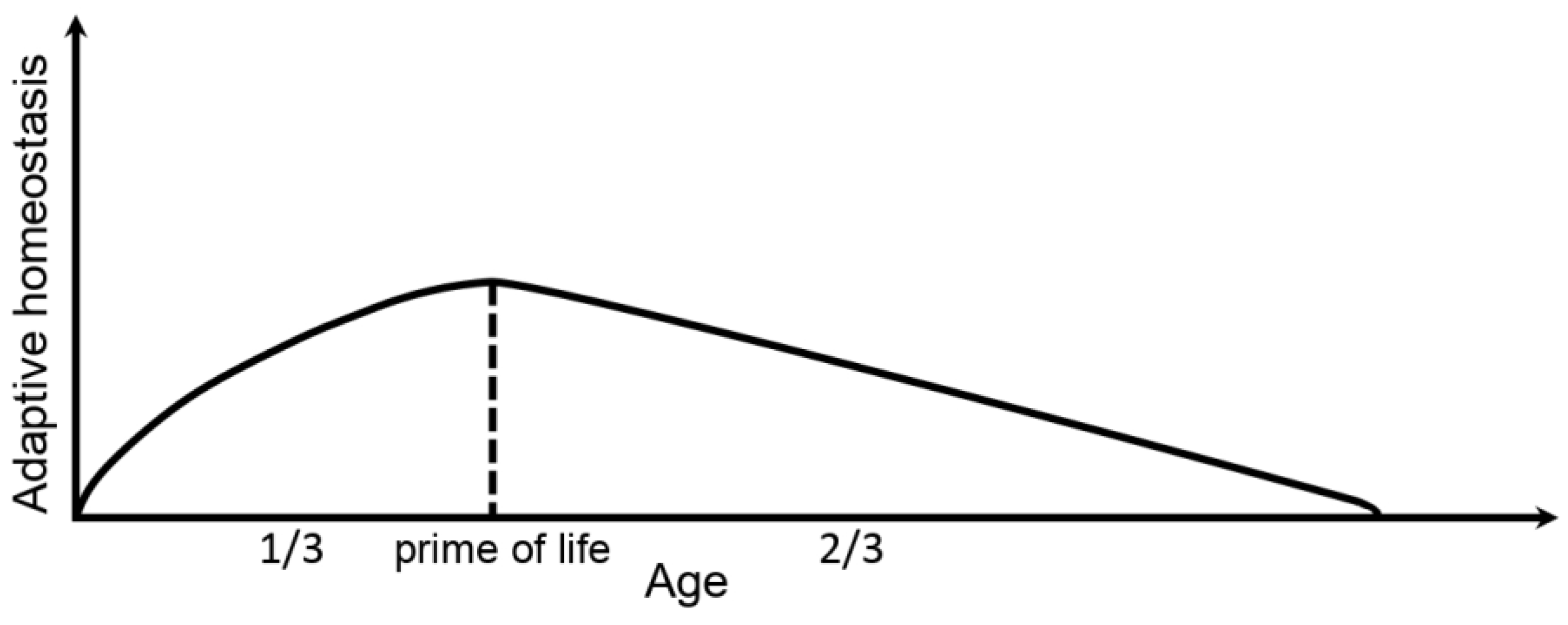
:1. Introduction
2. Materials and Methods
3. Experiment Scheme
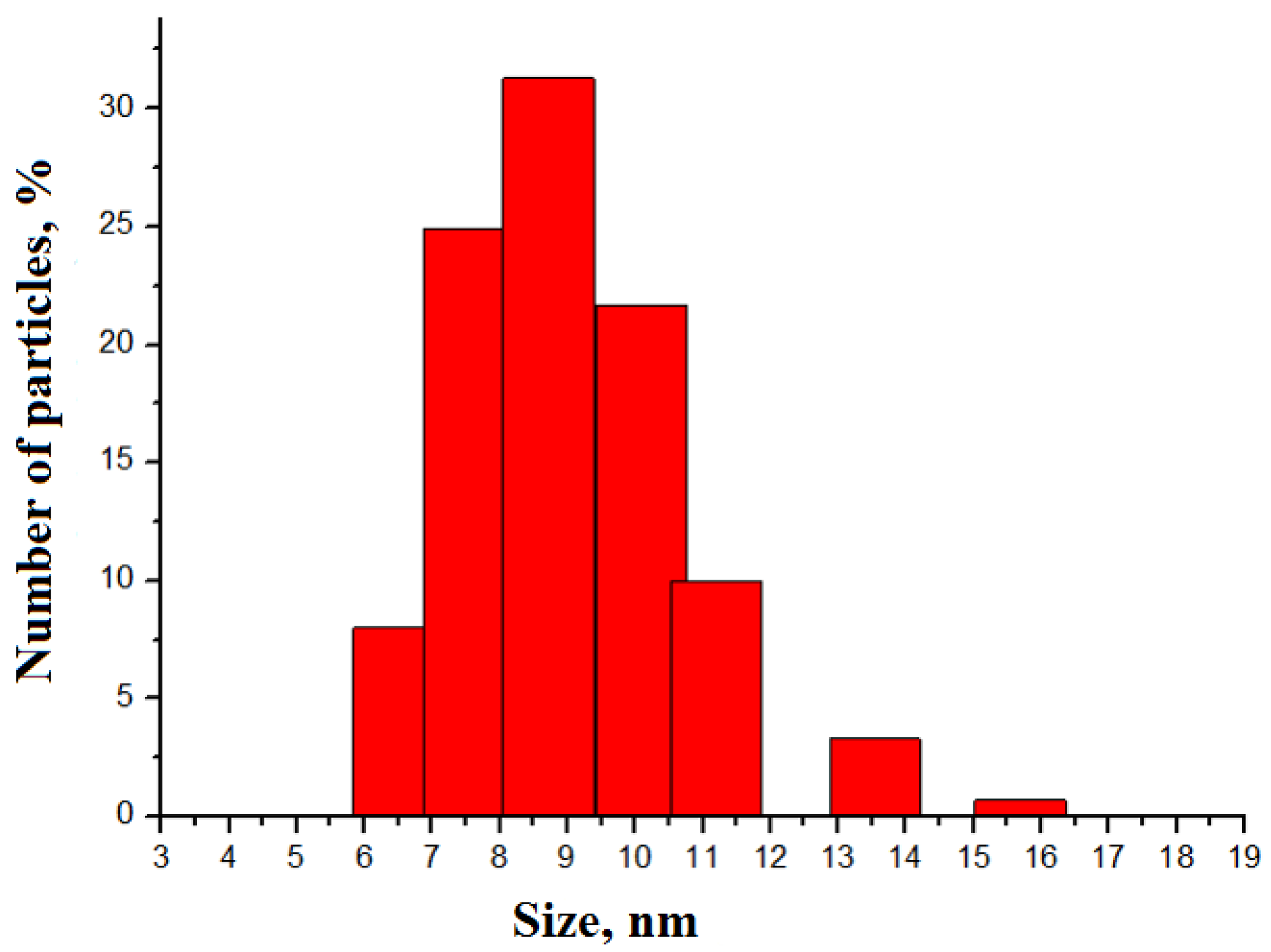
3.1. Study of Nanoparticle’s Morphology
3.2. Animal Treatment and Preparation Administration
3.3. Behavioral Tests
3.3.1. Open Field
3.3.2. Elevated Plus Maze
3.3.3. Light-Dark Box
3.4. Silver Content Measurement in Organs
3.5. Statistical Analysis
4. Results
4.1. Nanoparticles
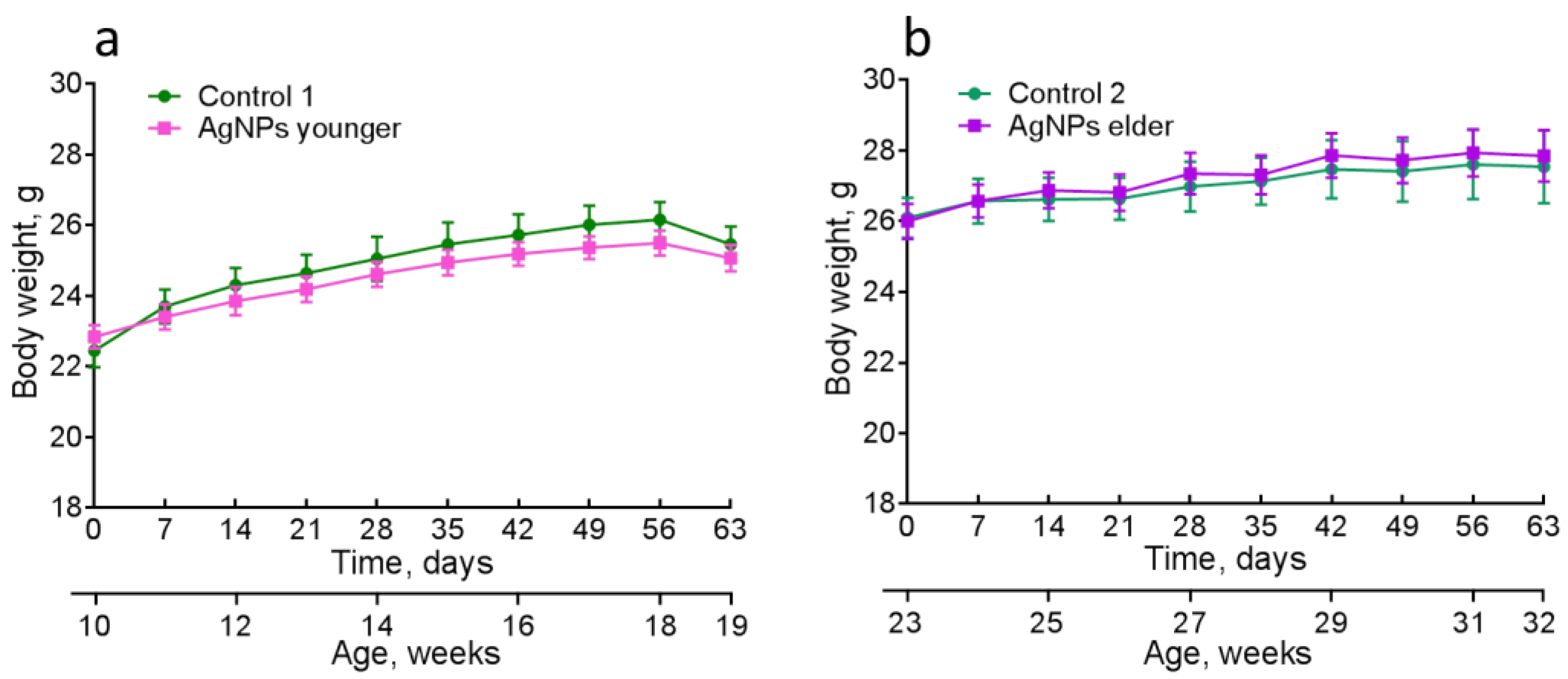
4.2. Physiological Characteristics
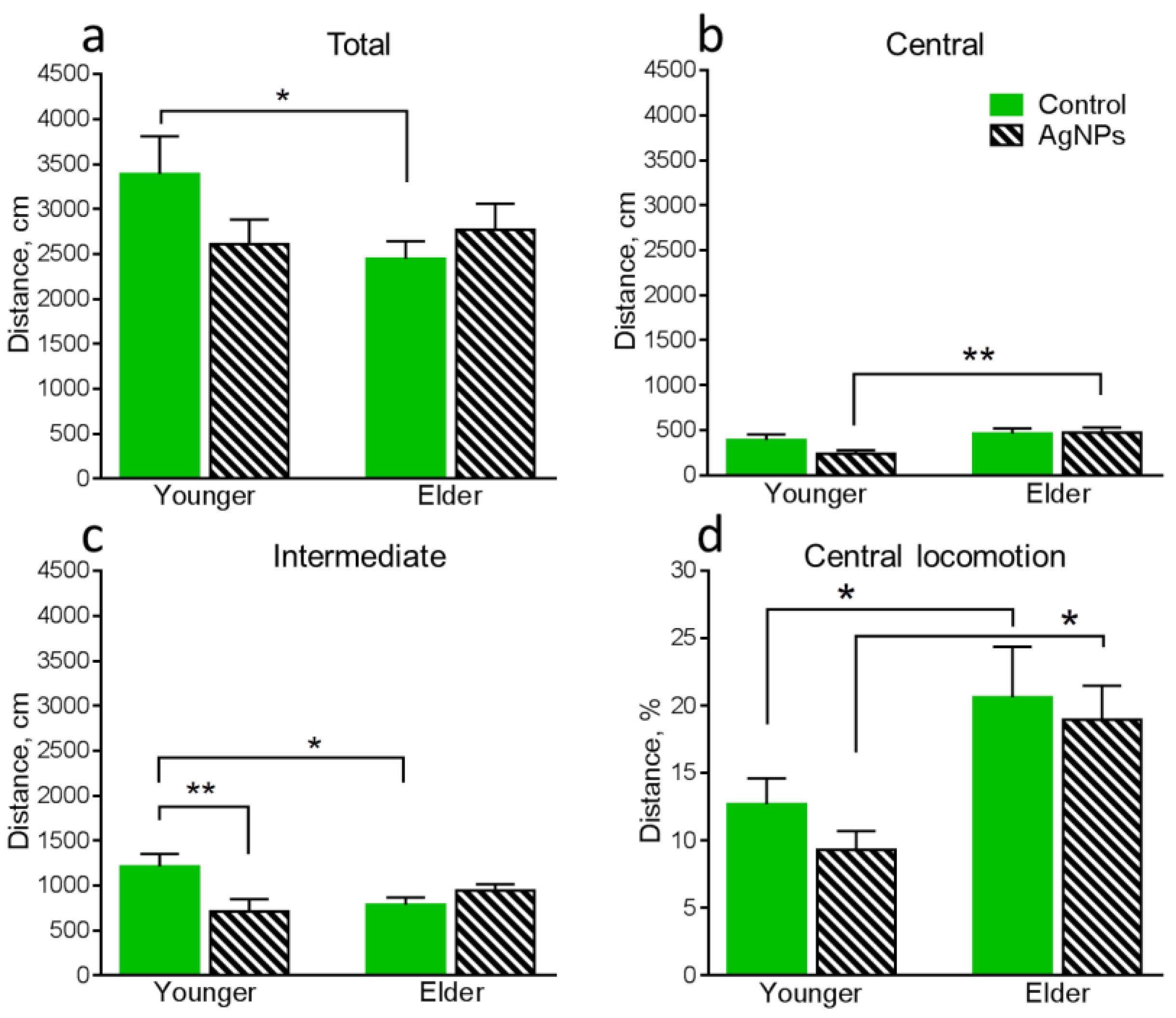
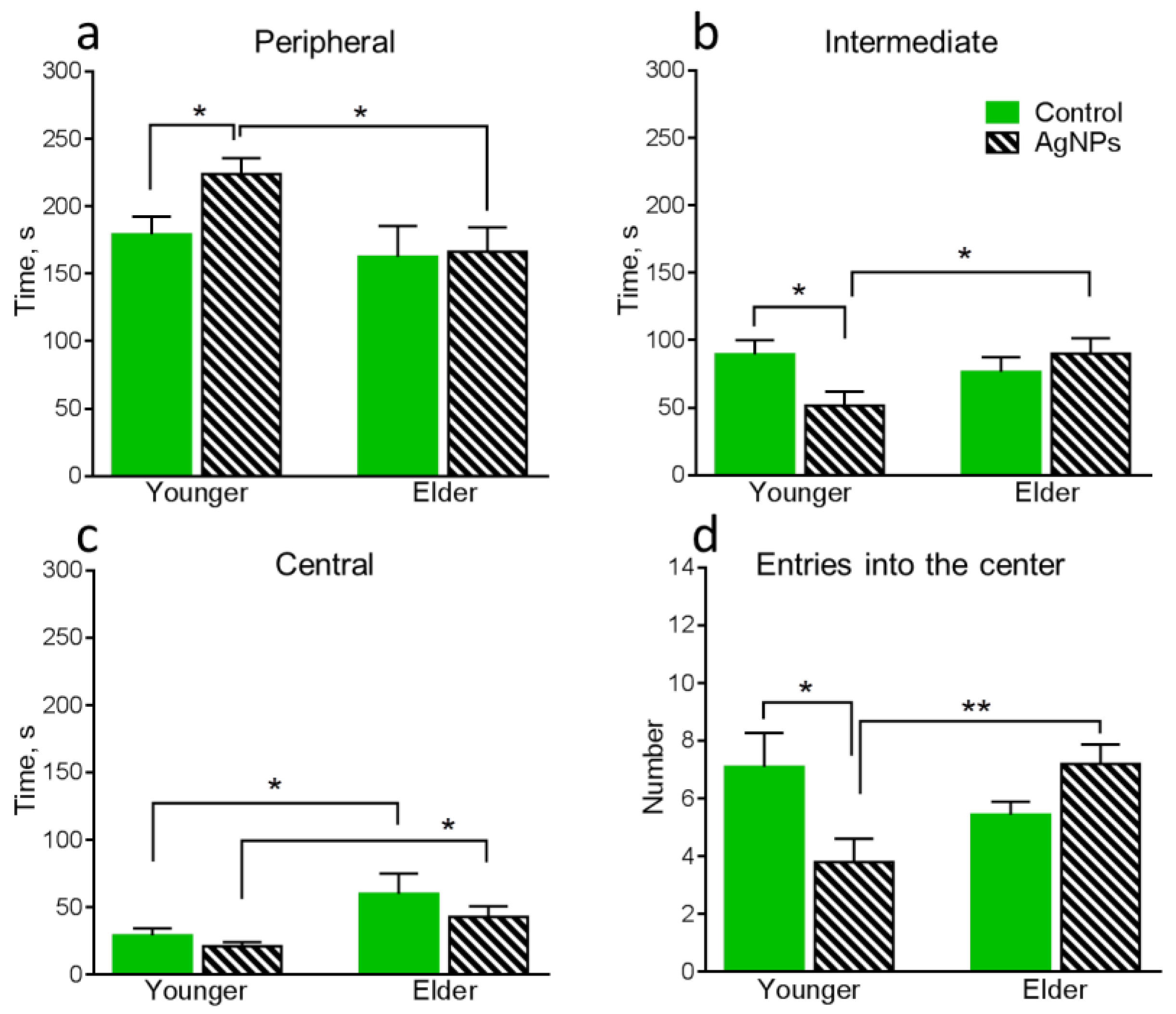
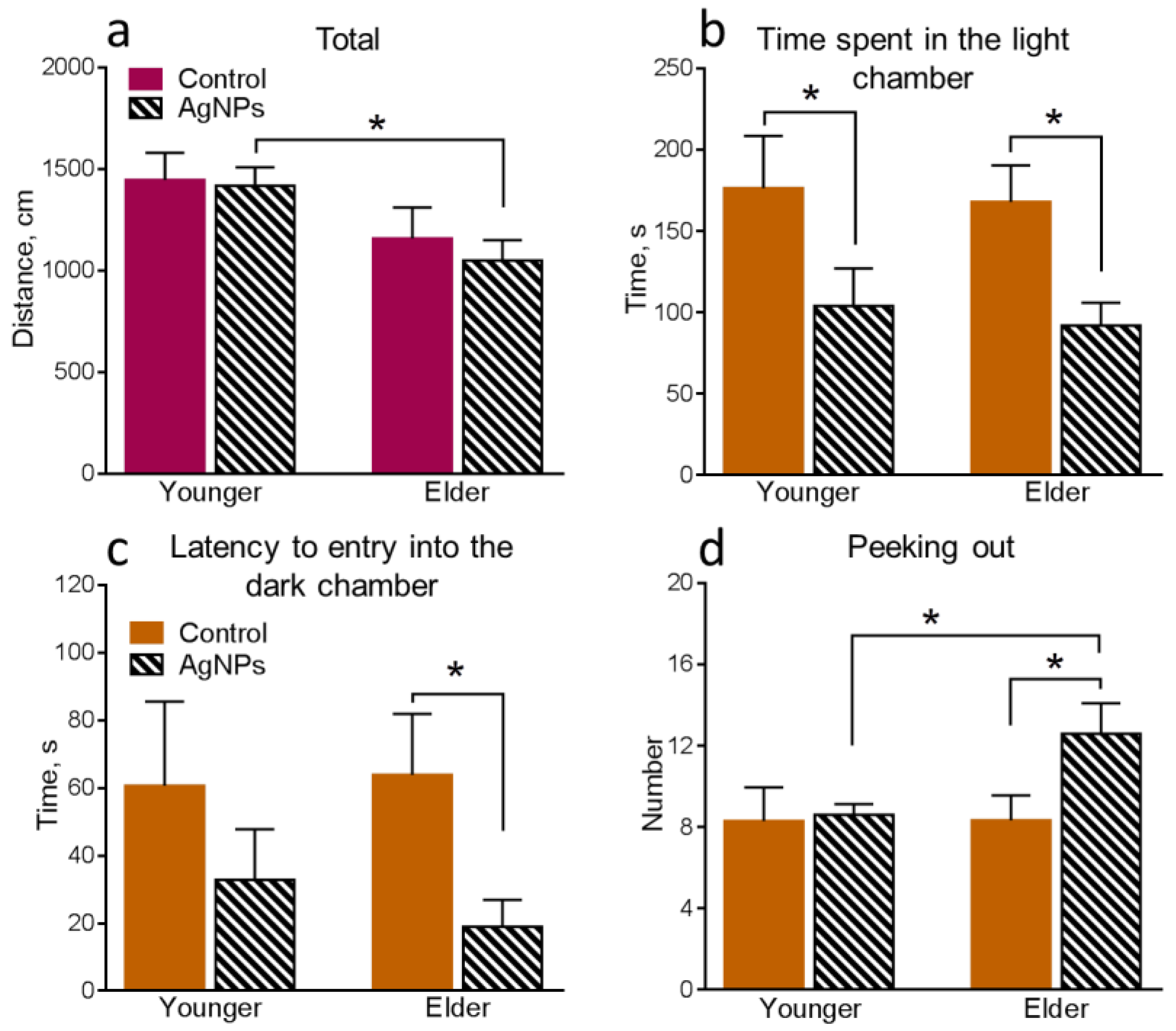
4.3. Behavioral Functions of Animals
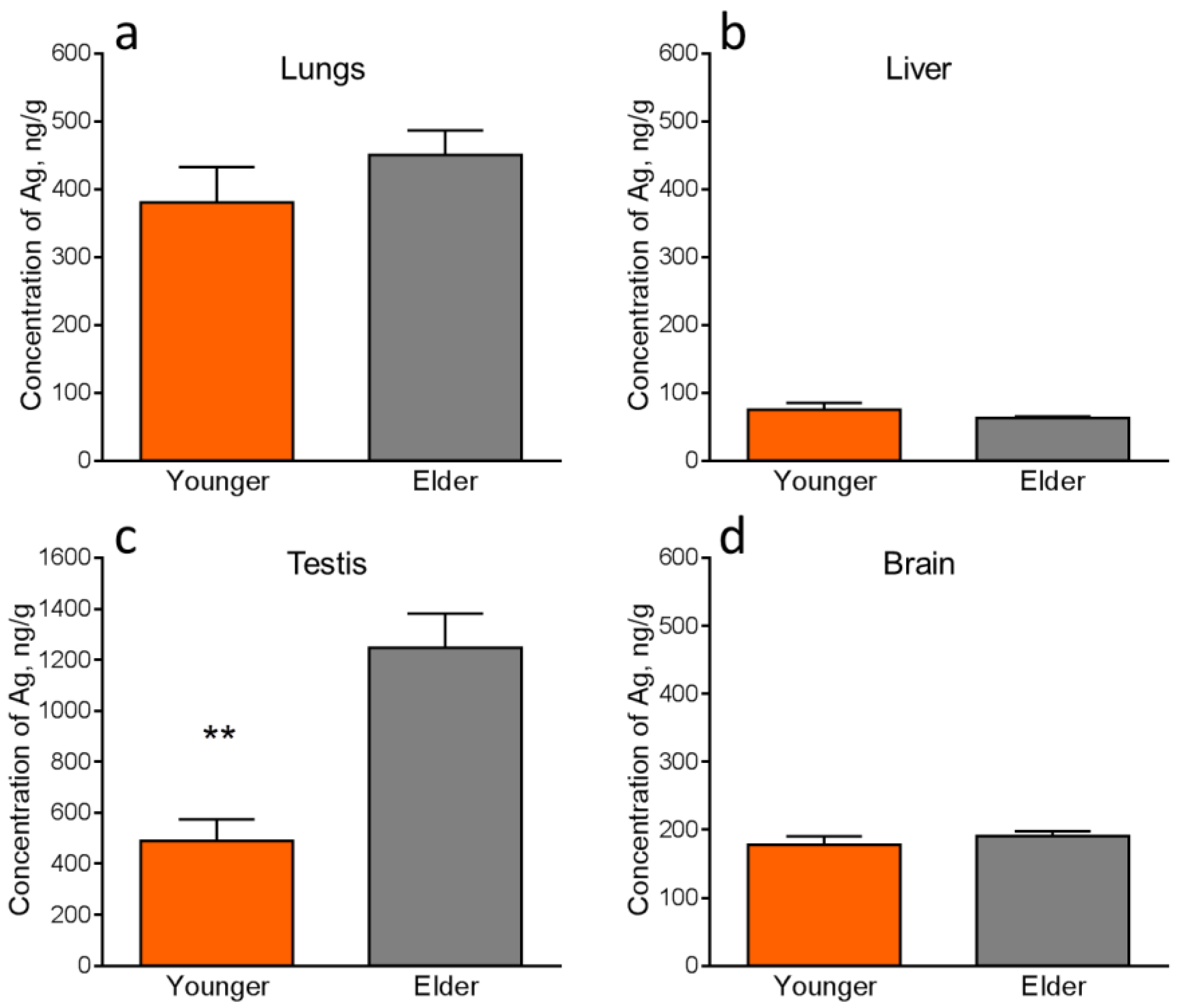
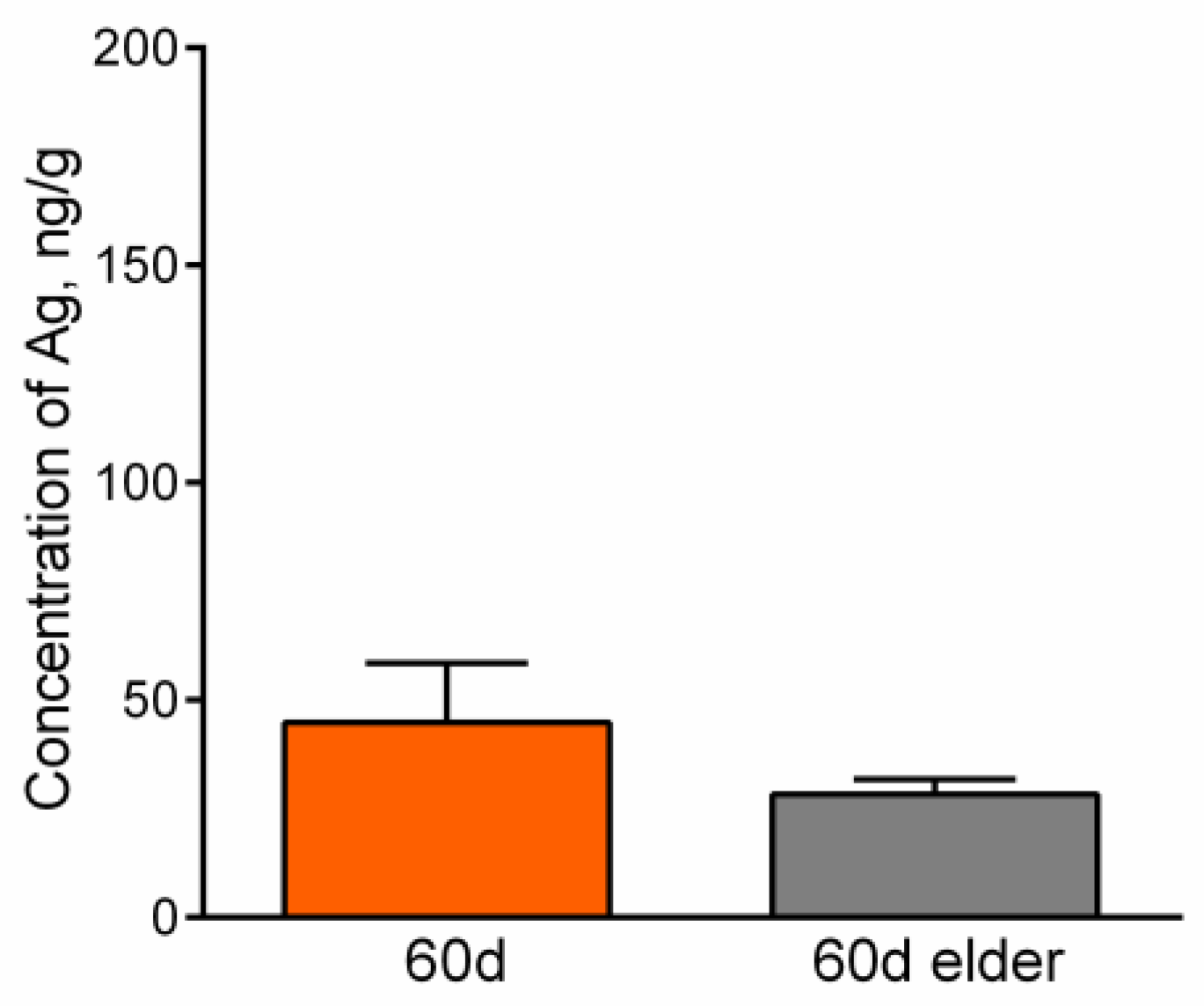
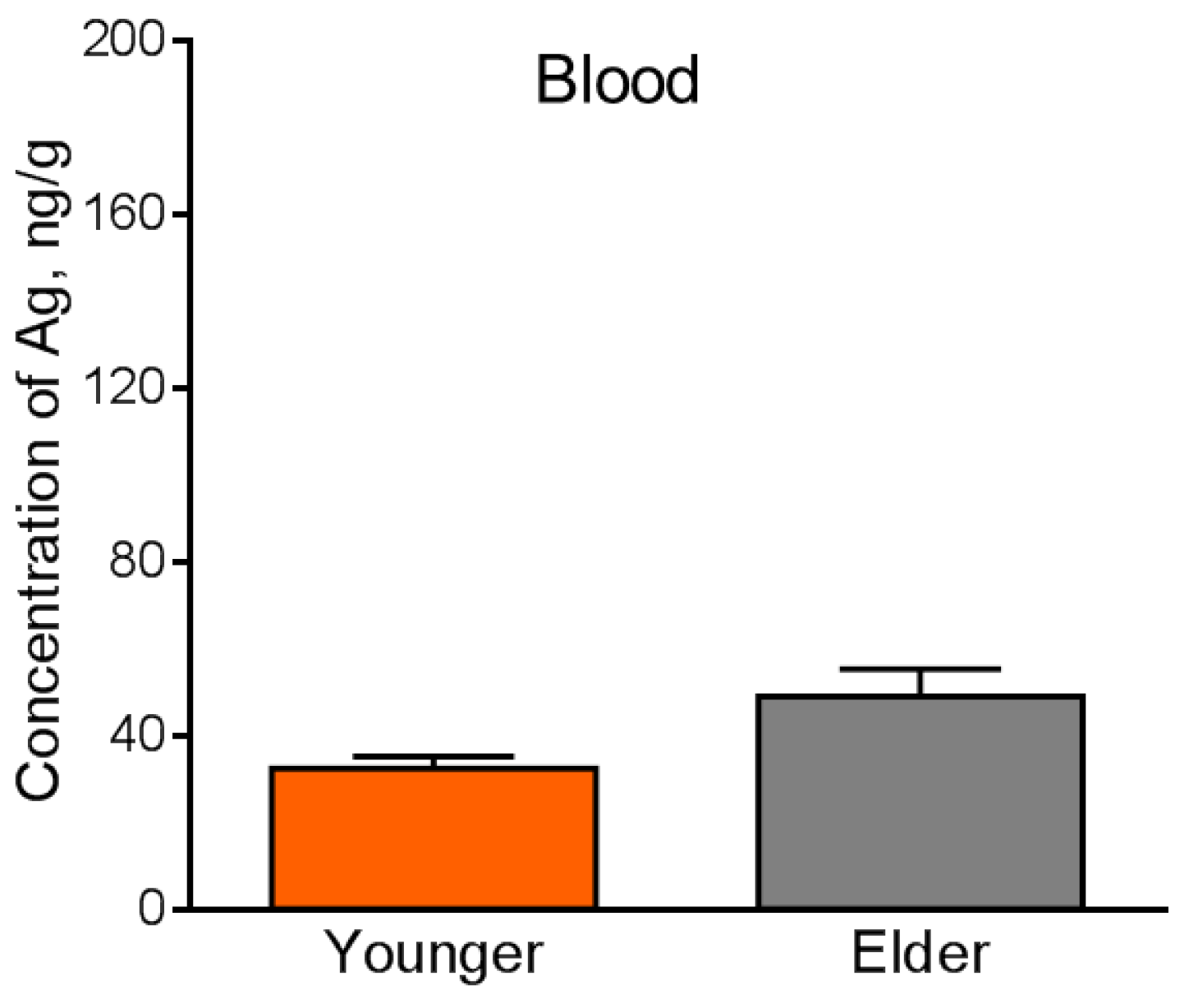
4.4. Silver Accumulation in the Internal Organs
5. Discussion
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Davies, K.J.A. Adaptive homeostasis. Mol. Aspects Med. 2016, 49, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Pomatto, L.C.D.; Davies, K.J.A. The role of declining adaptive homeostasis in ageing. J. Physiol. 2017, 595, 7275–7309. [Google Scholar] [CrossRef] [PubMed]
- Oranskii, I.E.; Makaryan, A.N.; Kochergin, Y.V. Biorythms and Life (Clocks inside us). In Kamyshilovskaya Tiporgafia; Uprinformpechati Pravitelstva Sverdlovskoi Oblasti: Ekaterinburg, Russia, 1980; p. 268. (In Russian) [Google Scholar]
- Dayem, A.A.; Hossain, M.K.; Lee, S.B.; Kim, K.; Saha, S.K.; Yang, G.M.; Cho, S.G. The Role of Reactive Oxygen Species (ROS) in the Biological Activities of Metallic Nanoparticles. Int. J. Mol. Sci. 2017, 18, 120. [Google Scholar] [CrossRef] [PubMed]
- Antsiferova, A.A.; Kashkarov, P.K.; Kovalchuk, M.V. Effect of Different Forms of Silver on Biological Objects. Nanobiotechnol. Rep. 2022, 17, 155–164. [Google Scholar] [CrossRef]
- Xu, L.; Wang, Y.Y.; Huang, J.; Chen, C.Y.; Wang, Z.X.; Xie, H. Silver nanoparticles: Synthesis, medical applications and biosafety. Theranostics 2020, 10, 8996–9031. [Google Scholar] [CrossRef]
- Railean, V.; Buszewska-Forajta, M.; Rodzik, A.; Gołębiowski, A.; Pomastowski, P.; Buszewski, B. In Vivo Efficacy of Wound Healing under External (Bio)AgNCs Treatment: Localization Case Study in Liver and Blood Tissue. Int. J. Mol. Sci. 2023, 24, 434. [Google Scholar] [CrossRef]
- Desai, A.S.; Singh, A.; Edis, Z.; Haj Bloukh, S.; Shah, P.; Pandey, B.; Agrawal, N.; Bhagat, N. An In Vitro and In Vivo Study of the Efficacy and Toxicity of Plant-Extract-Derived Silver Nanoparticles. J. Funct. Biomater. 2022, 13, 54. [Google Scholar] [CrossRef]
- Janzadeh, A.; Behroozi, Z.; Saliminia, F.; Janzadeh, N.; Arzani, H.; Tanha, K.; Hamblin, R.; Ramezani, F. Neurotoxicity of silver nanoparticles in the animal brain: A systematic review and meta-analysis. Forensic Toxicol. 2022, 40, 49–63. [Google Scholar] [CrossRef]
- Haase, A.; Rott, S.; Mantion, A.; Graf, P.; Plendl, J.; Andreas, F.; Thunemann, A.F.; Wolfgang, P.; Meier, W.P.; Andreas Taubert, A.; et al. Effects of silver nanoparticles on primary mixed neural cell cultures: Uptake, oxidative stress and acute calcium responses. Toxicol. Sci. 2012, 126, 457–468. [Google Scholar] [CrossRef]
- Elsharkawy, E.E.; Abd El-Nasser, M.; Kamaly, H.F. Silver nanoparticles testicular toxicity in rat. Environ. Toxicol. Pharmacol. 2019, 70, 103194. [Google Scholar] [CrossRef]
- Faedmaleki, F.H.; Shirazi, F.; Salarian, A.A.; Ahmadi Ashtiani, H.; Rastegar, H. Toxicity Effect of Silver Nanoparticles on Mice Liver Primary Cell Culture and HepG2 Cell Line. Iran J. Pharm. Res. 2014, 13, 235–242. [Google Scholar] [PubMed]
- van der Zande, M.; Vandebriel, R.J.; Van Doren, E.; Kramer, E.; Herrera Rivera, Z.; Serrano-Rojero, C.S.; Gremmer, E.R.; Mast, J.; Peters, R.J.B.; Hollman, P.C.H.; et al. Distribution, Elimination, and Toxicity of Silver Nanoparticles and Silver Ions in Rats after 28-Day Oral Exposure. ACS Nano 2012, 6, 7427–7442. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Kim, Y.S.; Song, K.S.; Ryu, H.R.; Sung, J.H.; Park, J.D.; Park, H.M.; Song, N.W.; Shin, B.S.; Marshak, D.; et al. Biopersistence of silver nanoparticles in tissues from Sprague–Dawley rats. Part Fibre Toxicol. 2013, 10, 36. [Google Scholar] [CrossRef] [PubMed]
- Antsiferova, A.; Buzulukov, Y.; Demin, V.; Kashkarov, P.; Kovalchuk, M.; Petritskaya, E. Extremely low level of Ag nanoparticle excretion from mice brain in in vivo experiments. IOP Conf. Ser. Mater. Sci. Eng. 2015, 98, 012003. [Google Scholar] [CrossRef]
- Antsiferova, A.A.; Kopaeva, M.Y.; Kochkin, V.N.; Kashkarov, P.K. Kinetics of Silver Accumulation in Tissues of Laboratory Mice after Long-Term Oral Administration of Silver Nanoparticles. Nanomaterials 2021, 11, 3204. [Google Scholar] [CrossRef]
- Antsiferova, A.; Kopaeva, M.; Kashkarov, P. Effects of Prolonged Silver Nanoparticle Exposure on the Contextual Cognition and Behavior of Mammals. Materials 2018, 11, 558. [Google Scholar] [CrossRef]
- Greish, K.; Alqahtani, A.A.; Alotaibi, A.F.; Abdulla, A.M.; Bukelly, A.T.; Alsobyani, F.M.; Alharbi, G.H.; Alkiyumi, I.S.; Aldawish, M.M.; Alshahrani, T.F.; et al. The Effect of Silver Nanoparticles on Learning, Memory and Social Interaction in BALB/C Mice. Int. J. Environ. Res. Public Health 2019, 16, 148. [Google Scholar] [CrossRef]
- Dziendzikowska, K.; Węsierska, M.; Gromadzka-Ostrowska, J.; Wilczak, J.; Oczkowski, M.; Męczyńska-Wielgosz, S.; Kruszewski, M. Silver Nanoparticles Impair Cognitive Functions and Modify the Hippocampal Level of Neurotransmitters in a Coating-Dependent Manner. Int. J. Mol. Sci. 2021, 22, 12706. [Google Scholar] [CrossRef]
- Antsiferova, A.A.; Kopaeva, M.Y.; Kochkin, V.N.; Kashkarov, P.K.; Kovalchuk, M.V. Disturbance in Mammalian Cognition Caused by Accumulation of Silver in Brain. Toxics 2021, 9, 30. [Google Scholar] [CrossRef]
- Dąbrowska-Bouta, B.; Sulkowski, G.; Strużyński, W. Prolonged Exposure to Silver Nanoparticles Results in Oxidative Stress in Cerebral Myelin. Neurotox Res. 2019, 35, 495. [Google Scholar] [CrossRef]
- Ahmed, S.M.; Abdelrahman, S.A.; Shalaby, S.M. Evaluating the effect of silver nanoparticles on testes of adult albino rats (histological, immunohistochemical and biochemical study). J. Mol. Hist. 2017, 48, 9–27. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.-Y.; Su, Y.-C.; Cheng, F.-Y.; Yan, S.-J.; Wang, Y.-J. Lifetime bioaccumulation of silver nanoparticles accelerates functional aging by inactivating antioxidant pathways, an effect reversed by pterostilbene. Environ. Sci. Nano 2021, 8, 3774. [Google Scholar] [CrossRef]
- Ferdous, Z.; Nemmar, A. Health Impact of Silver Nanoparticles: A Review of the Biodistribution and Toxicity Following Various Routes of Exposure. Int. J. Mol. Sci. 2020, 21, 2375. [Google Scholar] [CrossRef]
- Dziendzikowska, K.; Wilczak, J.; Grodzicki, W.; Gromadzka-Ostrowska, J.; Węsierska, M.; Kruszewski, M. Coating-Dependent Neurotoxicity of Silver Nanoparticles-An In Vivo Study on Hippocampal Oxidative Stress and Neurosteroids. Int. J. Mol. Sci. 2022, 23, 1365. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Carter, D.A.; Fen Leo, B.; Ruenraroengsak, P.; Chen, S.; Goode, A.E.; Theodorou, I.G.; Fan Chung, K.; Carzaniga, R.; Shaffer, M.S.P.; Dexter, D.T.; et al. Silver nanoparticles reduce brain inflammation and related neurotoxicity through induction of H2S-synthesizing enzymes. Sci. Rep. 2017, 7, 42871. [Google Scholar] [CrossRef] [PubMed]
- Available online: http://vector-vita.narod.ru/products.html (accessed on 3 January 2023).
- Flurkey, K.; Currer, J.M.; Harrison, D.E. Mouse models in aging research. In The Mouse in Biomedical Research, 2nd ed.; Fox, J.G., Barthold, S.W., Davisson, M.T., Newcomer, C.E., Quimby, F.W., Smith, A.L., Eds.; Academic Press: Burlington, MA, USA, 2007; pp. 637–672. [Google Scholar] [CrossRef]
- Kraeuter, A.K.; Guest, P.C.; Sarnyai, Z. The Elevated Plus Maze Test for Measuring Anxiety-Like Behavior in Rodents. Methods Mol. Biol. 2019, 1916, 69–74. [Google Scholar] [CrossRef]
- Kulesskaya, N.; Voikar, V. Assessment of mouse anxiety-like behavior in the light-dark box and open-field arena: Role of equipment and procedure. Physiol. Behav. 2014, 133, 30–38. [Google Scholar] [CrossRef]
- Campos-Cardoso, R.; Godoy, L.D.; Lazarini-Lopes, W.; Novaes, L.S.; Dos Santos, N.B.; Perfetti, J.G.; Garcia-Cairasco, N.; Munhoz, C.D.; Padovan, C.M. Exploring the light/dark box test: Protocols and implications for neuroscience research. J. Neurosci. Methods 2023, 384, 109748. [Google Scholar] [CrossRef]
- Kopaeva, M.Y.; Alchinova, I.B.; Cherepov, A.B.; Demorzhi, M.S.; Nesterenko, M.V.; Zarayskaya, I.Y.; Karganov, M.Y. New Properties of a Well-Known Antioxidant: Pleiotropic Effects of Human Lactoferrin in Mice Exposed to Gamma Irradiation in a Sublethal Dose. Antioxidants 2022, 11, 1833. [Google Scholar] [CrossRef]
- Sun, B.; Lorang, C.; Qin, S.; Zhang, Y.; Liu, K.; Li, G.; Sun, Z.; Francke, A.; Utleg, A.G.; Hu, Z.; et al. Mouse Organ-Specific Proteins and Functions. Cells 2021, 10, 3449. [Google Scholar] [CrossRef]
- Demin, V.F.; Antsiferova, A.A.; Buzulukov, Y.P.; Demin, V.A.; Soloviev, V.Y. Nuclear physical method for the detection of chemical elements in biological and other samples using activation by charged particles. Med. Radiol. Radiat. Saf. 2015, 60, 60–65. [Google Scholar]
- TERMINIX. Available online: https://www.terminix.com/rodents/mice/habitat/ (accessed on 29 November 2022).
- Xue, Y.; Zhang, S.; Huang, Y.; Zhang, T.; Liu, X.; Hu, Y.; Zhang, Z.; Tang, M. Acute toxic effects and gender-related biokinetics of silver nanoparticles following an intravenous injection in mice. J. Appl. Toxicol. 2012, 32, 890–899. [Google Scholar] [CrossRef] [PubMed]
- Martin, K.L.; Blizzard, L.; Srikanth, V.K.; Wood, A.; Thomson, R.; Sanders, L.M.; Callisaya, M.L. Cognitive Function Modifies the Effect of Physiological Function on the Risk of Multiple Falls—A Population-Based Study. J. Gerontol. Ser. A 2013, 68, 1091–1097. [Google Scholar] [CrossRef] [PubMed]
- Mandolesi, L.; Polverino, A.; Montuori, S.; Foti, F.; Ferraioli, G.; Sorrentino, P.; Sorrentino, G. Effects of Physical Exercise on Cognitive Functioning and Wellbeing: Biological and Psychological Benefits. Front. Psychol. 2018, 9, 509. [Google Scholar] [CrossRef]
- Available online: https://www.sciencealert.com/does-iq-decline-as-we-age-one-type-of-intelligence-peaks-in-your-twenties (accessed on 3 January 2023).
- Aizawa, K. Cognition and behavior. Synthese 2017, 194, 4269–4288. [Google Scholar] [CrossRef]
- Cornaggia, C.M.; Beghi, M.; Provenzi, M. Correlation between Cognition and Behavior in Epilepsy. Epilepsia 2006, 47, 34–39. [Google Scholar] [CrossRef]
- Wang, E.; Huang, Y.; Du, Q.; Sun, Y. Alterations in reproductive parameters and gene expression in Balb/c mice testes after exposure to silver nanoparticles. Andrologia 2021, 53, e13841. [Google Scholar] [CrossRef]
- Fathi, N.; Hoseinipanah, S.M.; Alizadeh, Z.; Assari, M.J.; Moghimbeigi, A.; Mortazavi, M.; Hosseini, M.H.; Bahmanzadeh, M. The effect of silver nanoparticles on the reproductive system of adult male rats: A morphological, histological and DNA integrity study. Adv. Clin. Exp. Med. 2019, 28, 299–305. [Google Scholar] [CrossRef]
- Kose, O.; Mantecca, P.; Costa, A.; Carrière, M. Putative adverse outcome pathways for silver nanoparticle toxicity on mammalian male reproductive system: A literature review. Part Fibre Toxicol. 2023, 20, 1. [Google Scholar] [CrossRef]
- Zinkovskaya, I.; Ivlieva, A.L.; Petritskaya, E.N.; Rogatkin, D.A. An unexpected effect of long-term oral administration of silver nanoparticles on fertility in mice. Ecol. Cheloveka 2020, 27, 23–30. (In Russian) [Google Scholar] [CrossRef]
- Melnik, E.A.; Buzulukov, Y.P.; Demin, V.F.; Demin, V.A.; Gmoshinski, I.V.; Tyshko, N.V.; Tutelyan, V.A. Transfer of Silver Nanoparticles through the Placenta and Breast Milk during in vivo Experiments on Rats. Acta Nat. 2013, 5, 107–115. [Google Scholar] [CrossRef]
- Ramírez-Sánchez, M.; Prieto, I.; Segarra, A.B.; Banegas, I.; Martínez-Cañamero, M.; Domínguez-Vías, G.; de Gasparo, M. Brain Asymmetry: Towards an Asymmetrical Neurovisceral Integration. Symmetry 2021, 13, 2409. [Google Scholar] [CrossRef]
- Segarra, A.B.; Prieto, I.; Banegas, I.; Martínez-Cañamero, M.; de Gasparo, M.; Ramírez-Sánchez, M. Blood Pressure Correlates Asymmetrically with Neuropeptidase Activities of the Left and Right Frontal Cortices. Symmetry 2021, 13, 105. [Google Scholar] [CrossRef]
- Jindal, R.D.; Jennings, J.R. Grand challenges in psychosomatic research. Front. Psychiatry 2010, 1, 131. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Element | Ratio |
|---|---|---|
| 1 | C | 0.62 |
| 2 | O | 0.0165 |
| 3 | Al | 0.0006 |
| 4 | Ag | 0.039 |
| Organ | Group | Concentration, ng/g | Mass, ng |
|---|---|---|---|
| Heart | Younger | <11.2 | <1.7 |
| Elder | <15 | <2.4 | |
| Spleen | Younger | <24.6 | <1.5 |
| Elder | <76.4 | <4.6 |
| Mass, ng | ||||||
|---|---|---|---|---|---|---|
| Organ | Brain | Testes | Lungs | Liver | Kidneys | Blood |
| Younger | 85 ± 19 | 88 ± 31.5 | 78 ± 34 | 102 ± 41 | 18 ± 8 | 20 ± 4 |
| Elder | 82 ± 5 | 219 ± 58.5 | 97 ± 9 | 98 ± 13 | 12 ± 1 | 37 ± 12 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Antsiferova, A.A.; Kopaeva, M.Y.; Kochkin, V.N.; Reshetnikov, A.A.; Kashkarov, P.K. Neurotoxicity of Silver Nanoparticles and Non-Linear Development of Adaptive Homeostasis with Age. Micromachines 2023, 14, 984. https://doi.org/10.3390/mi14050984
Antsiferova AA, Kopaeva MY, Kochkin VN, Reshetnikov AA, Kashkarov PK. Neurotoxicity of Silver Nanoparticles and Non-Linear Development of Adaptive Homeostasis with Age. Micromachines. 2023; 14(5):984. https://doi.org/10.3390/mi14050984
Chicago/Turabian StyleAntsiferova, Anna A., Marina Yu. Kopaeva, Vyacheslav N. Kochkin, Alexander A. Reshetnikov, and Pavel K. Kashkarov. 2023. "Neurotoxicity of Silver Nanoparticles and Non-Linear Development of Adaptive Homeostasis with Age" Micromachines 14, no. 5: 984. https://doi.org/10.3390/mi14050984