

Importance of Spatial Arrangement of Cardiomyocyte Network for Precise and Stable On-Chip Predictive Cardiotoxicity Measurement

Abstract
:1. Introduction
2. Principle
3. Materials and Methods
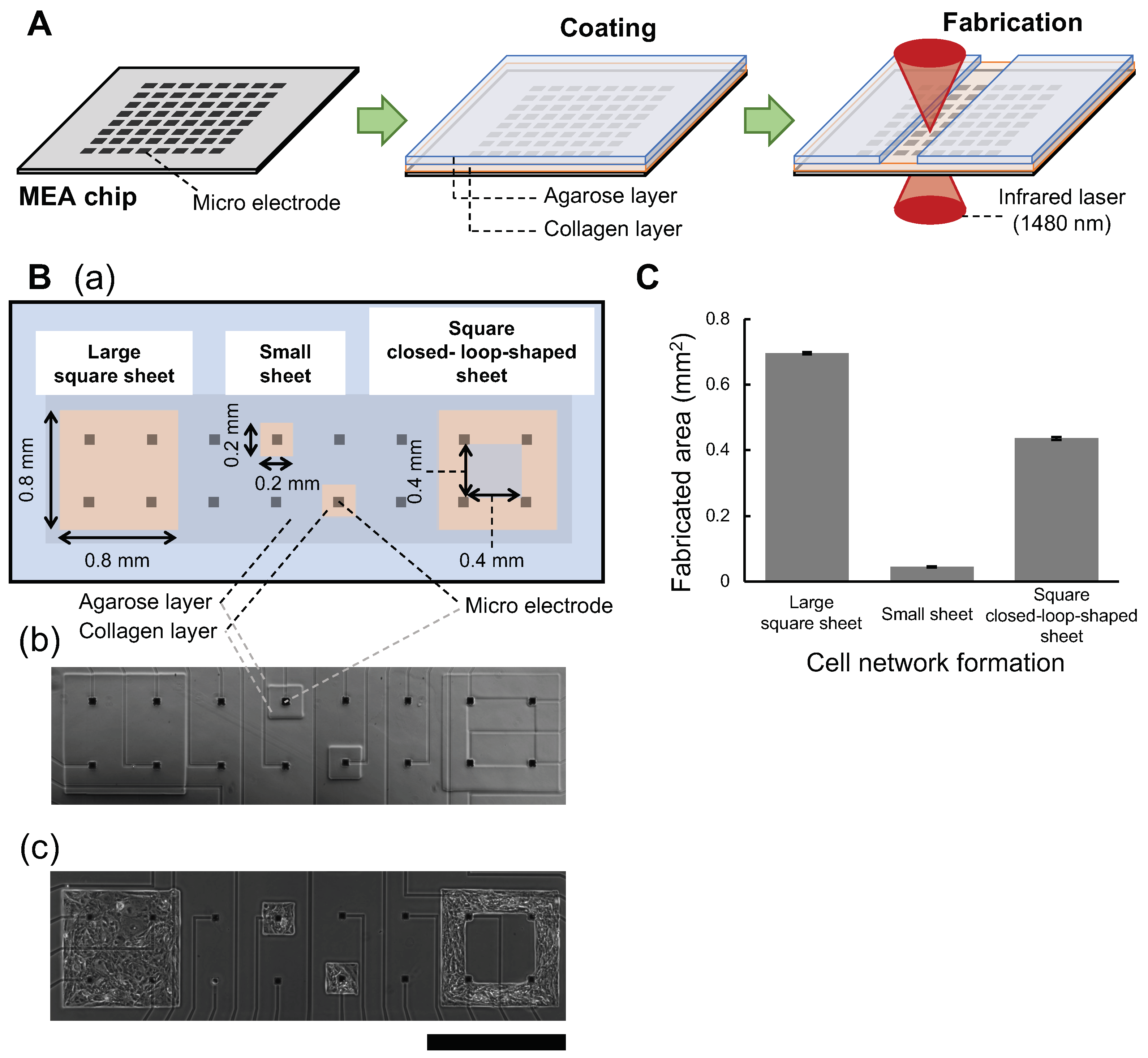
3.1. Agarose Microchamber Formation on Multielectrode Chips
3.2. Human Embryonic Stem Cell-Derived Cardiomyocyte Cultivation
3.3. Test Compound
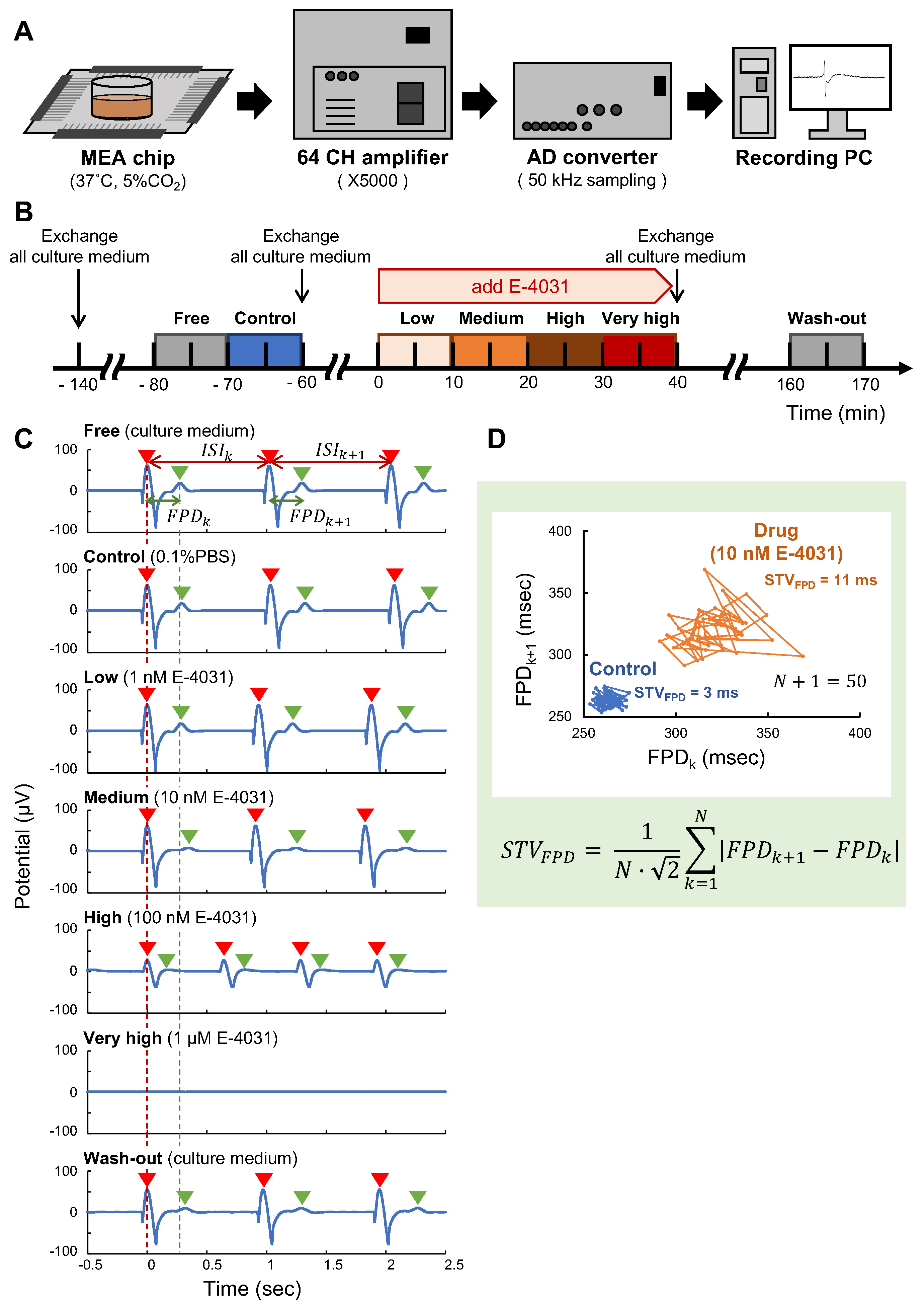
3.4. Field Potential Recordings
3.5. Data Analysis
3.6. Statistical Analysis
4. Results and Discussion
4.1. Fabrication of Three Formats of Cardiomyocyte Cell Network Patterns
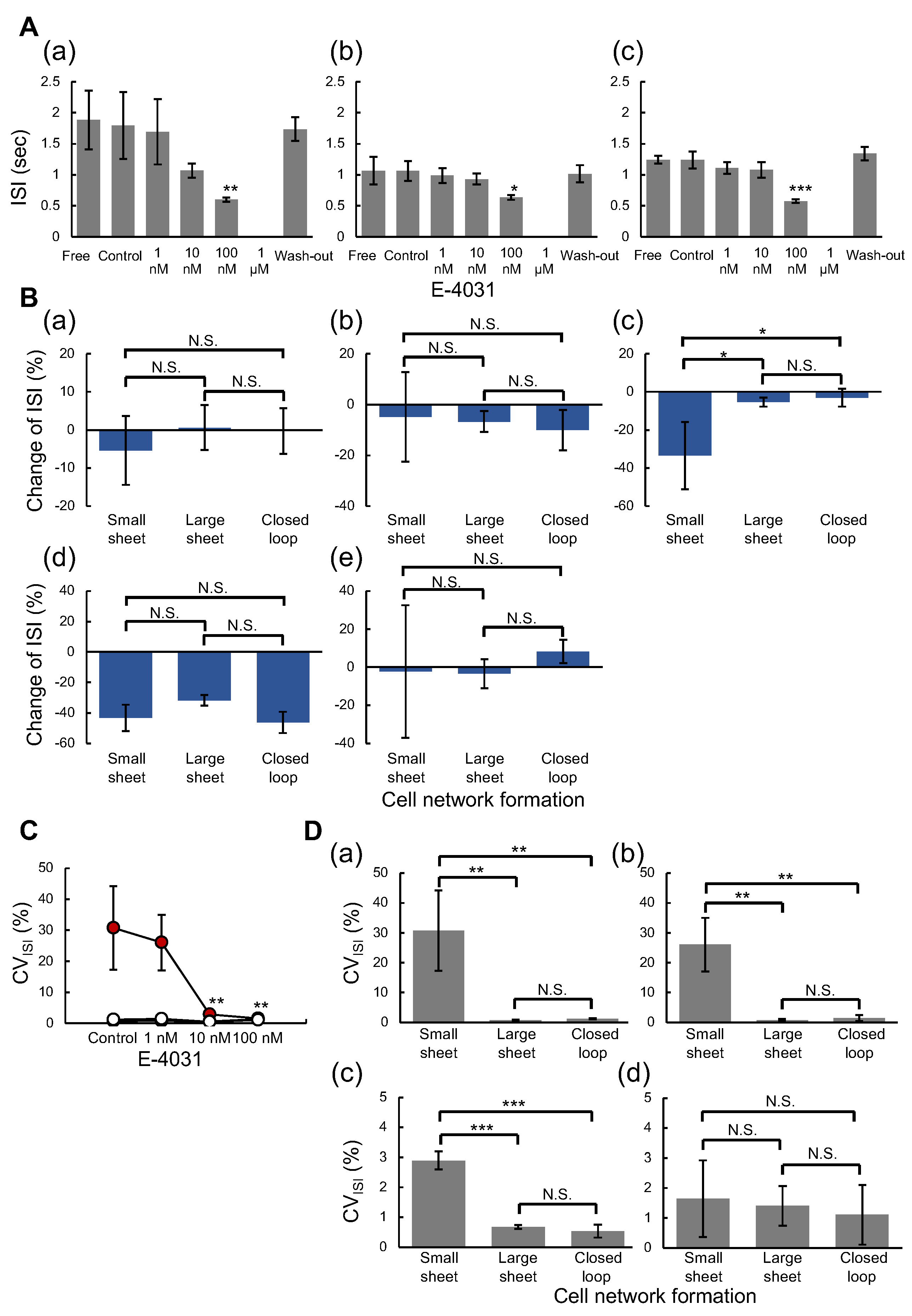
4.2. Measurement of hERG Ion Channel Blocker E-4031 Response in Three Formats of Cardiomyocyte Networks
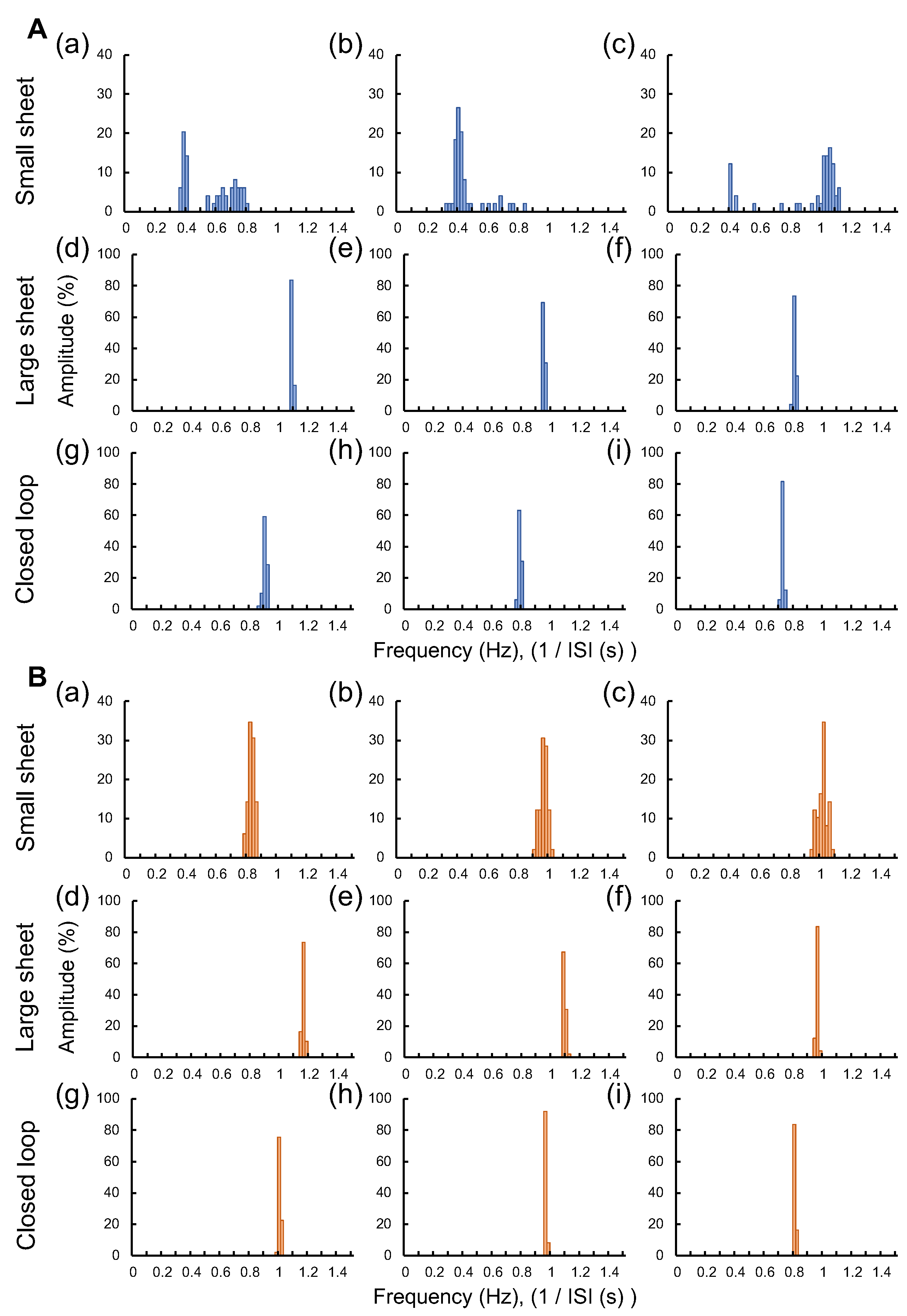
4.3. Effect of hERG Ion Channel Blocker E-4031 on Beating Inter-Spike Intervals of Three Type Cardiomyocyte Networks
4.4. Effect of hERG Ion Channel Blocker E-4031 on Field Potential Duration Time of Cardiomyocyte Networks
4.5. Importance of Geometry-Controlled Cardiomyocyte Network Analysis Using at Least Three Geometry of Network Patterns
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gintant, G. An evaluation of hERG current assay performance: Translating preclinical safety studies to clinical QT prolongation. Pharmacol. Ther. 2011, 129, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Ponti, F.D.; Poluzzi, E.; Cavalli, A.; Recanatini, M.; Montanaro, N.; De Ponti, F.; Poluzzi, E.; Cavalli, A.; Recanatini, M.; Montanaro, N.; et al. Safety of non-antiarrhythmic drugs that prolong the QT interval or induce torsade de pointes: An overview. Drug Saf. Int. J. Med. Toxicol. Drug Exp. 2002, 25, 263–286. [Google Scholar] [CrossRef] [PubMed]
- Giorgi, M.; Bolanos, R.; Gonzalez, C.; Di Girolamo, G. QT interval prolongation: Preclinical and clinical testing arrhythmogenesis in drugs and regulatory Implications. Curr. Drug Saf. 2010, 5, 54–57. [Google Scholar] [CrossRef] [PubMed]
- Gintant, G.; Sager, P.T.; Stockbridge, N. Evolution of strategies to improve preclinical cardiac safety testing. Nat. Rev. Drug Discov. 2016, 15, 457–471. [Google Scholar] [CrossRef]
- Colatsky, T.; Fermini, B.; Gintant, G.; Pierson, J.B.; Sager, P.; Sekino, Y.; Strauss, D.G.; Stockbridge, N. The Comprehensive in Vitro Proarrhythmia Assay (CiPA) initiative—Update on progress. J. Pharmacol. Toxicol. Methods 2016, 81, 15–20. [Google Scholar] [CrossRef] [Green Version]
- Sartipy, P.; Björquist, P.; Strehl, R.; Hyllner, J. The application of human embryonic stem cell technologies to drug discovery. Drug Discov. Today 2007, 12, 688–699. [Google Scholar] [CrossRef]
- Jahnke, H.G.; Steel, D.; Fleischer, S.; Seidel, D.; Kurz, R.; Vinz, S.; Dahlenborg, K.; Sartipy, P.; Robitzki, A.A. A Novel 3D Label-Free Monitoring System of hES-Derived Cardiomyocyte Clusters: A Step Forward to In Vitro Cardiotoxicity Testing. PLoS ONE 2013, 8, e68971. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, T.; Tohyama, S.; Murata, M.; Nomura, F.; Kaneko, T.; Chen, H.; Hattori, F.; Egashira, T.; Seki, T.; Ohno, Y.; et al. In vitro pharmacologic testing using human induced pluripotent stem cell-derived cardiomyocytes. Biochem. Biophys. Res. Commun. 2009, 385, 497–502. [Google Scholar] [CrossRef]
- Shi, Y.; Inoue, H.; Wu, J.C.; Yamanaka, S. Induced pluripotent stem cell technology: A decade of progress. Nat. Rev. Drug Discov. 2016, 16, 115–130. [Google Scholar] [CrossRef] [PubMed]
- Rowe, R.G.; Daley, G.Q. Induced pluripotent stem cells in disease modelling and drug discovery. Nat. Rev. Genet. 2019, 20, 377–388. [Google Scholar] [CrossRef]
- Asahi, Y.; Hamada, T.; Hattori, A.; Matsuura, K.; Odaka, M.; Nomura, F.; Kaneko, T.; Abe, Y.; Takasuna, K.; Sanbuissho, A.; et al. On-chip spatiotemporal electrophysiological analysis of human stem cell derived cardiomyocytes enables quantitative assessment of proarrhythmia in drug development. Sci. Rep. 2018, 8, 14536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Redondo, F.; Kurokawa, J.; Nomura, F.; Kaneko, T.; Hamada, T.; Furukawa, T.; Yasuda, K. A distribution analysis of action potential parameters obtained from patch-clamped human stem cell-derived cardiomyocytes. J. Pharmacol. Sci. 2016, 131, 141–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Z.; Gong, Q.; Ye, B.; Fan, Z.; Makielski, J.C.; Robertson, G.A.; January, C.T. Properties of HERG channels stably expressed in HEK 293 cells studied at physiological temperature. Biophys. J. 1998, 74, 230–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirsch, G.E.; Trepakova, E.S.; Brimecombe, J.C.; Sidach, S.S.; Erickson, H.D.; Kochan, M.C.; Shyjka, L.M.; Lacerda, A.E.; Brown, A.M. Variability in the measurement of hERG potassium channel inhibition: Effects of temperature and stimulus pattern. J. Pharmacol. Toxicol. Methods 2004, 50, 93–101. [Google Scholar] [CrossRef]
- Koivisto, M.; Tolvanen, T.A.; Toimela, T.; Miinalainen, I.; Kiviaho, A.; Kesseli, J.; Nykter, M.; Eklund, L.; Heinonen, T. Functional human cell-based vascularised cardiac tissue model for biomedical research and testing. Sci. Rep. 2022, 12, 13459. [Google Scholar] [CrossRef]
- Kaneko, T.; Nomura, F.; Hamada, T.; Abe, Y.; Takamori, H.; Sakakura, T.; Takasuna, K.; Sanbuissho, A.; Hyllner, J.; Sartipy, P.; et al. On-chip in vitro cell-network pre-clinical cardiac toxicity using spatiotemporal human cardiomyocyte measurement on a chip. Sci. Rep. 2014, 4, 4670. [Google Scholar] [CrossRef] [Green Version]
- Sakamoto, K.; Aoki, S.; Tanaka, Y.; Shimoda, K.; Hondo, Y.; Yasuda, K. Geometric understanding of local fluctuation distribution of conduction time in lined-up cardiomyocyte network in agarose-microfabrication multi-electrode measurement assay. Micromachines 2020, 11, 1105. [Google Scholar] [CrossRef]
- Eng, G.; Lee, B.W.; Protas, L.; Gagliardi, M.; Brown, K.; Kass, R.S.; Keller, G.; Robinson, R.B.; Vunjak-Novakovic, G. Autonomous beating rate adaptation in human stem cell-derived cardiomyocytes. Nat. Commun. 2016, 7, 10312. [Google Scholar] [CrossRef] [Green Version]
- Sakamoto, K.; Hondo, Y.; Takahashi, N.; Tanaka, Y.; Sekine, R.; Shimoda, K.; Watanabe, H.; Yasuda, K. Emergent synchronous beating behavior in spontaneous beating cardiomyocyte clusters. Sci. Rep. 2021, 11, 11869. [Google Scholar] [CrossRef]
- Ji, G.; Barsotti, R.J.; Feldman, M.E.; Kotlikoff, M.I. Stretch-induced Calcium Release in Smooth Muscle. J. Gen. Physiol. 2002, 119, 533–543. [Google Scholar] [CrossRef] [Green Version]
- Hotta, S.; Morimura, K.; Ohya, S.; Muraki, K.; Takeshima, H.; Imaizumi, Y. Ryanodine receptor type 2 deficiency changes excitation-contraction coupling and membrane potential in urinary bladder smooth muscle. J. Physiol. 2007, 582, 489–506. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; He, L.; Zhang, S.M.; Luo, J. Human iPS Cell-derived Tissue Engineered Vascular Graft: Recent Advances and Future Directions. Stem Cell Rev. Rep. 2021, 17, 862–877. [Google Scholar] [CrossRef]
- Yang, X.; Pabon, L.; Murry, C.E. Engineering adolescence: Maturation of human pluripotent stem cell-derived cardiomyocytes. Circ. Res. 2014, 114, 511–523. [Google Scholar] [CrossRef] [Green Version]
- Takada, T.; Sasaki, D.; Matsuura, K.; Miura, K.; Sakamoto, S.; Goto, H.; Ohya, T.; Iida, T.; Homma, J.; Shimizu, T.; et al. Aligned human induced pluripotent stem cell-derived cardiac tissue improves contractile properties through promoting unidirectional and synchronous cardiomyocyte contraction. Biomaterials 2022, 281, 121351. [Google Scholar] [CrossRef]
- Nomura, F.; Kaneko, T.; Hamada, T.; Hattori, A.; Yasuda, K. Quantitative evaluation of closed-loop-shaped cardiomyocyte network by using ring-shaped electrode. Jpn. J. Appl. Phys. 2012, 51, 06FK06. [Google Scholar] [CrossRef]
- Kadota, S.; Minami, I.; Morone, N.; Heuser, J.E.; Agladze, K.; Nakatsuji, N. Development of a reentrant arrhythmia model in human pluripotent stem cell-derived cardiac cell sheets. Eur. Heart J. 2013, 34, 1147–1156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawatou, M.; Masumoto, H.; Fukushima, H.; Morinaga, G.; Sakata, R.; Ashihara, T.; Yamashita, J.K. Modelling Torsade de Pointes arrhythmias in vitro in 3D human iPS cell-engineered heart tissue. Nat. Commun. 2017, 8, 1078. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Small Sheet | Large Square Sheet | Square Closed-Loop-Shaped Sheet | |
|---|---|---|---|
| ISI | + | − | − |
| FPD | − | − | + |
| STV of FPD | − | − | + |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sakamoto, K.; Matsumoto, S.; Abe, N.; Sentoku, M.; Yasuda, K. Importance of Spatial Arrangement of Cardiomyocyte Network for Precise and Stable On-Chip Predictive Cardiotoxicity Measurement. Micromachines 2023, 14, 854. https://doi.org/10.3390/mi14040854
Sakamoto K, Matsumoto S, Abe N, Sentoku M, Yasuda K. Importance of Spatial Arrangement of Cardiomyocyte Network for Precise and Stable On-Chip Predictive Cardiotoxicity Measurement. Micromachines. 2023; 14(4):854. https://doi.org/10.3390/mi14040854
Chicago/Turabian StyleSakamoto, Kazufumi, Suguru Matsumoto, Nanami Abe, Mitsuru Sentoku, and Kenji Yasuda. 2023. "Importance of Spatial Arrangement of Cardiomyocyte Network for Precise and Stable On-Chip Predictive Cardiotoxicity Measurement" Micromachines 14, no. 4: 854. https://doi.org/10.3390/mi14040854