
Seed Priming with Glutamic-Acid-Functionalized Iron Nanoparticles Modulating Response of Vigna radiata (L.) R. Wilczek (Mung Bean) to Induce Osmotic Stress
 , ,
, ,
Abstract
:1. Introduction
2. Methods and Materials
2.1. Synthesis of Glutamic-Acid-Capped Iron Nanoparticles (Glu-FeNPs)
2.2. Characterization of Nanoparticles
2.3. Lab Experimental Details
2.4. Pot Experiment Details
2.5. Measurement of the Vegetative and Yield Parameters
2.6. Measurement of Biochemical Parameters
2.7. Statistical Analysis
3. Results
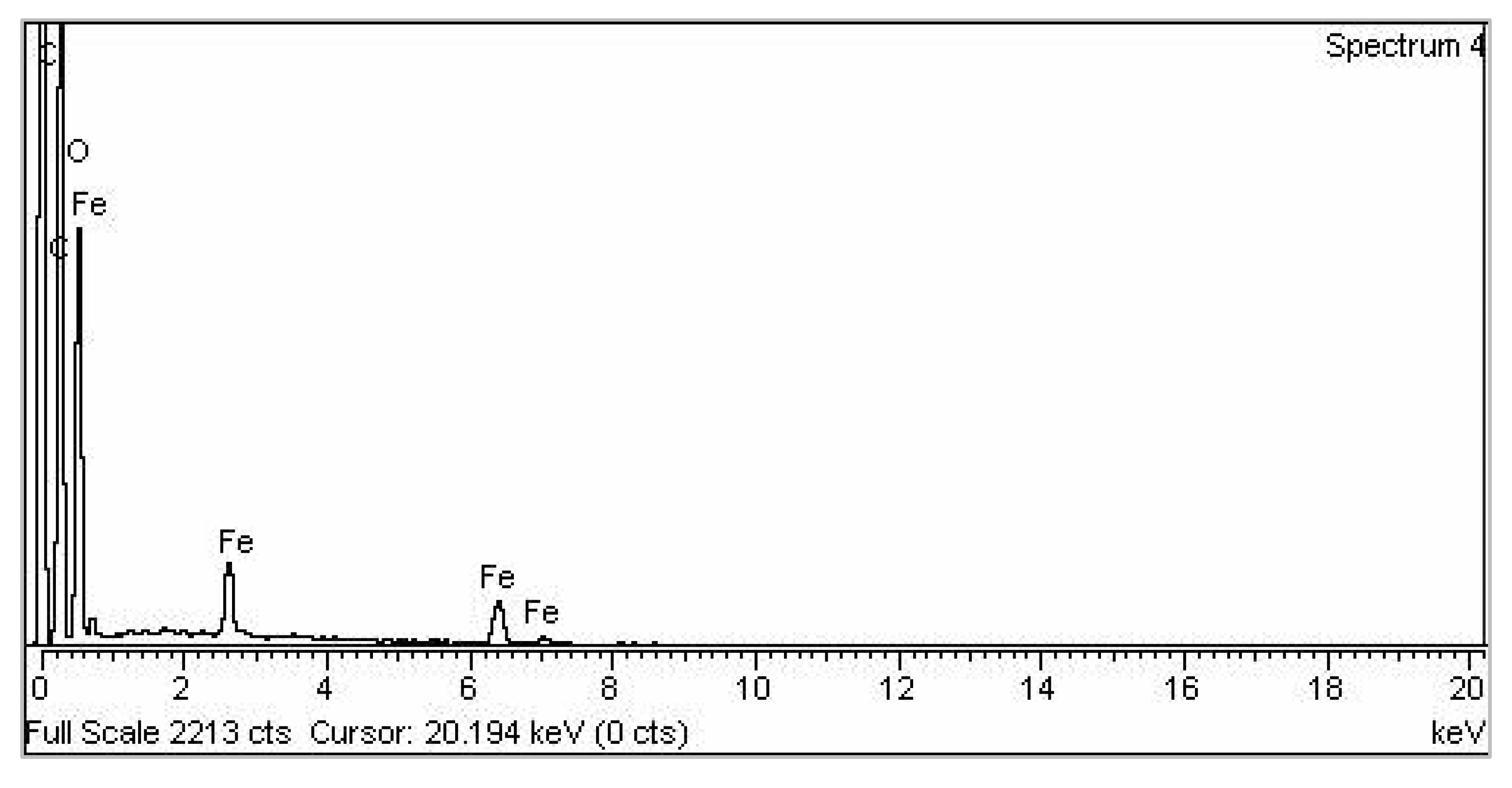
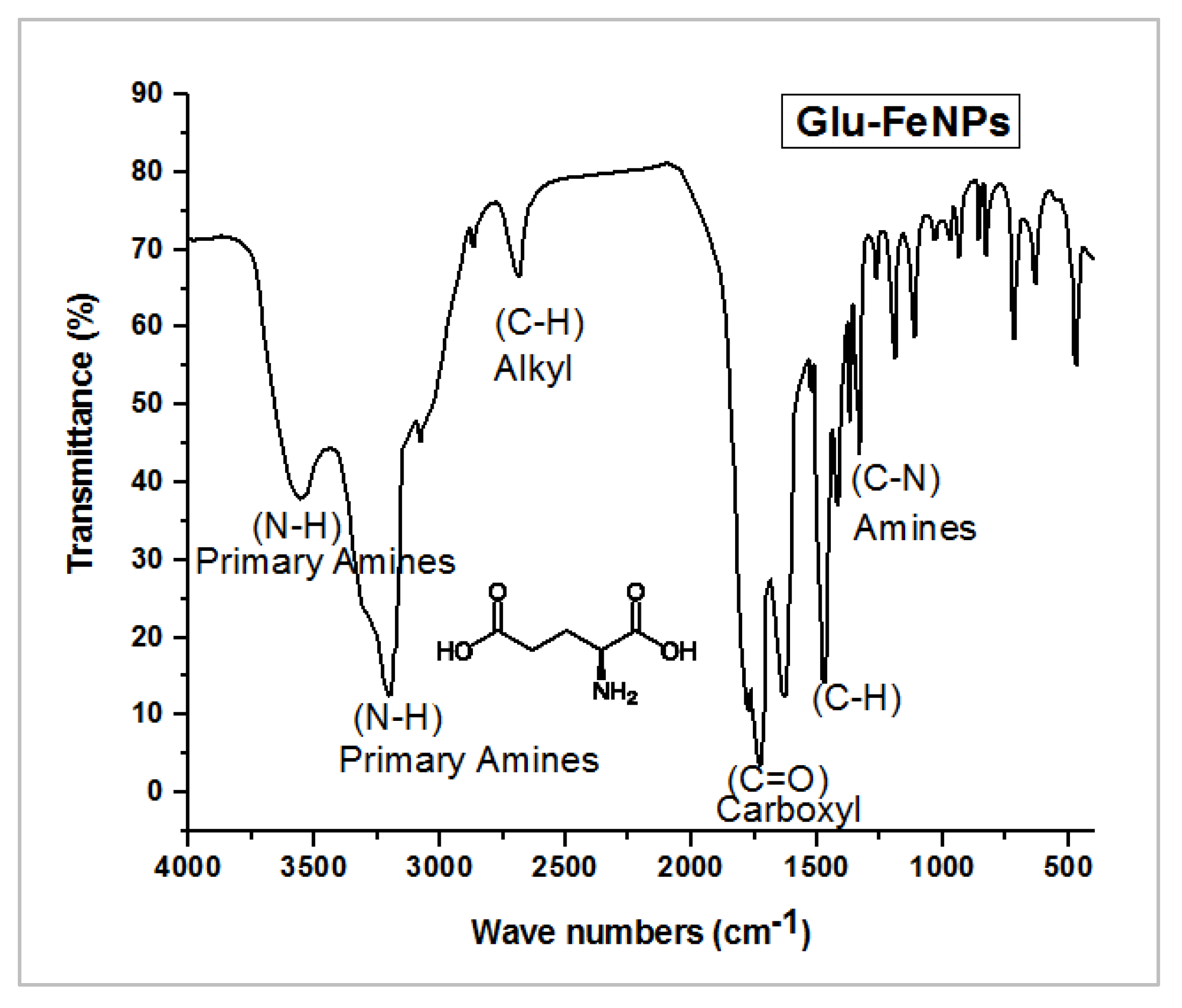
3.1. Characterization of Nanoparticles
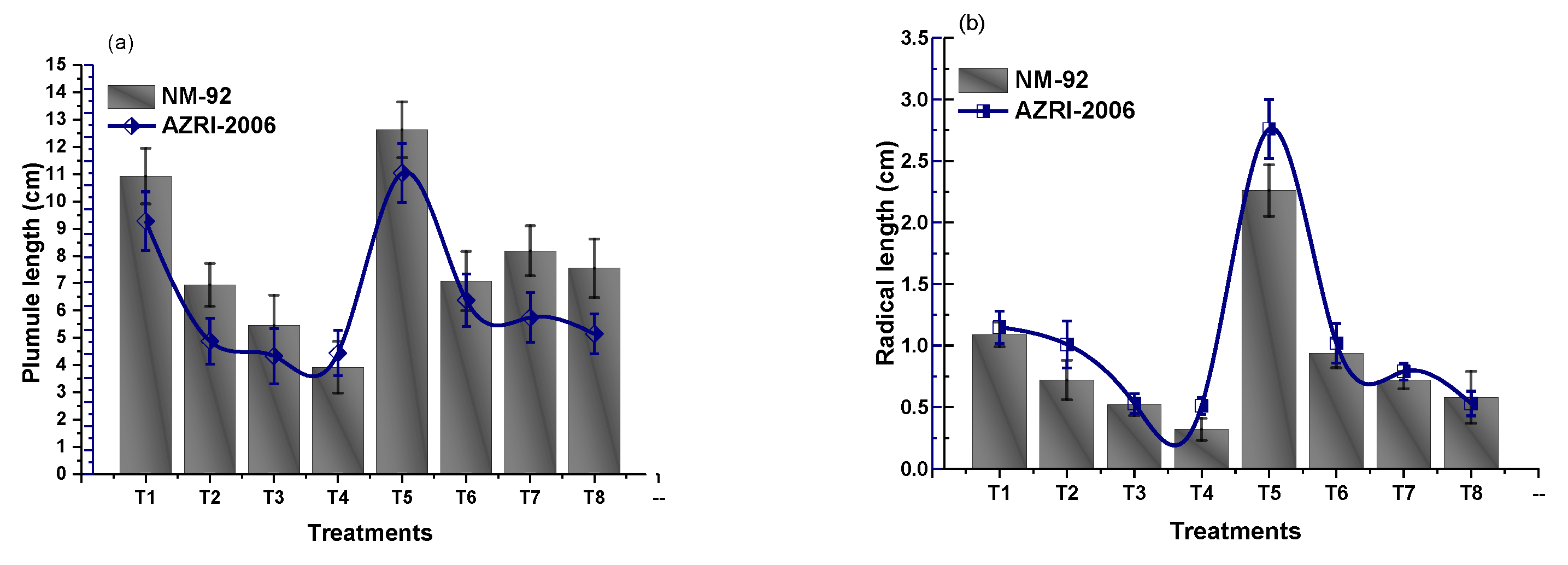
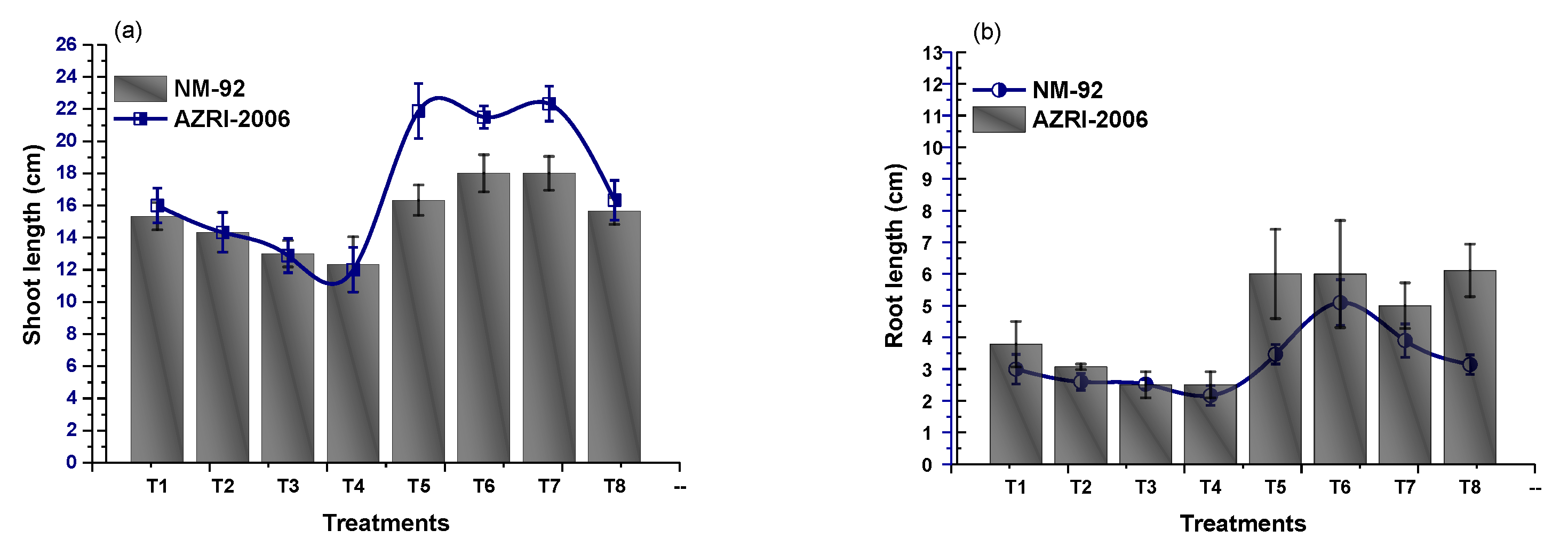
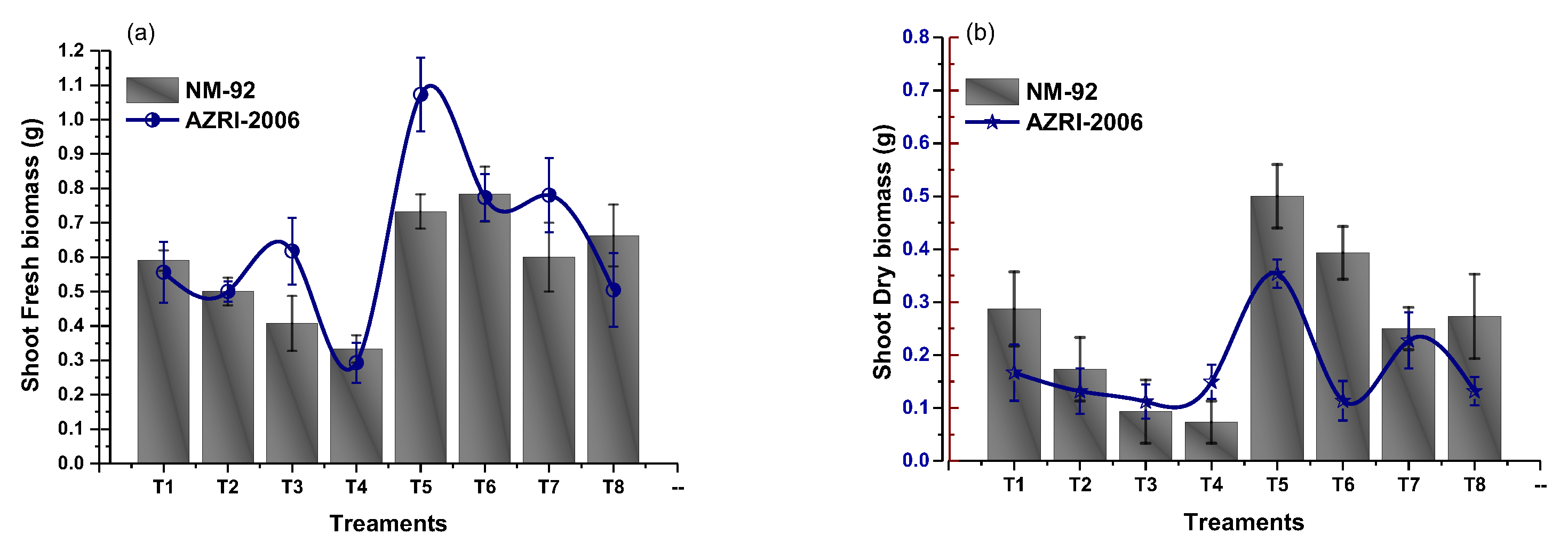
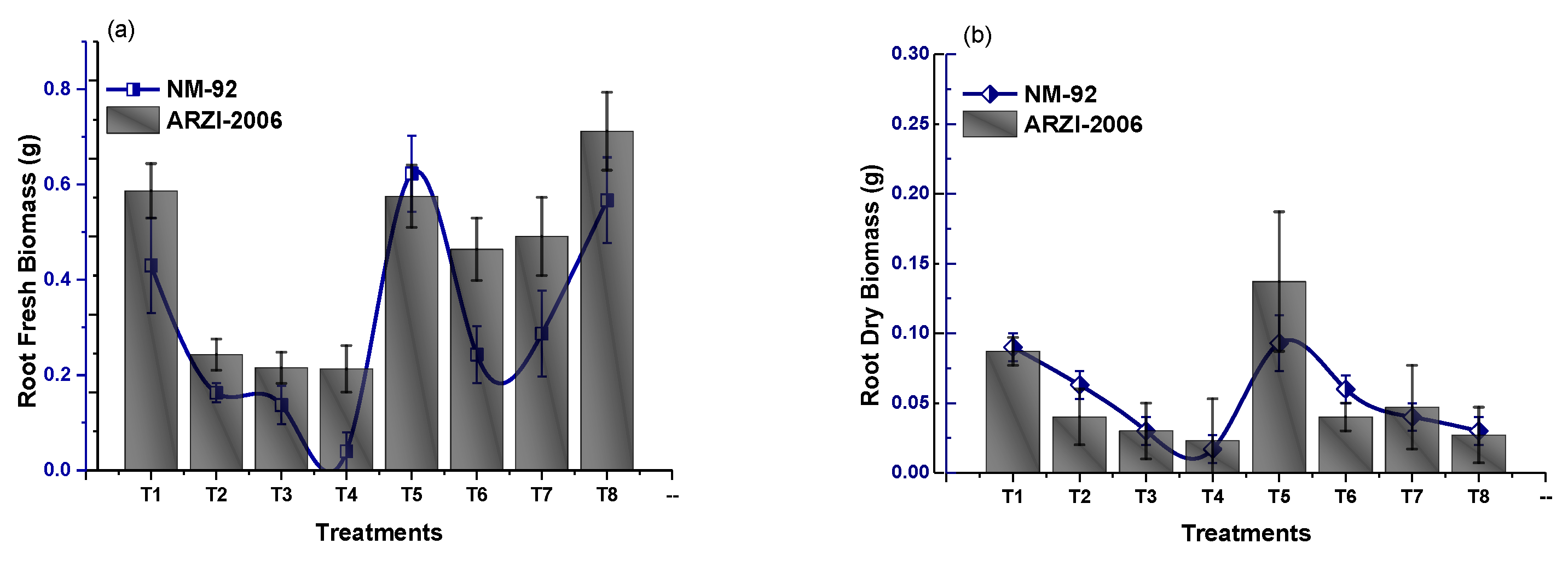
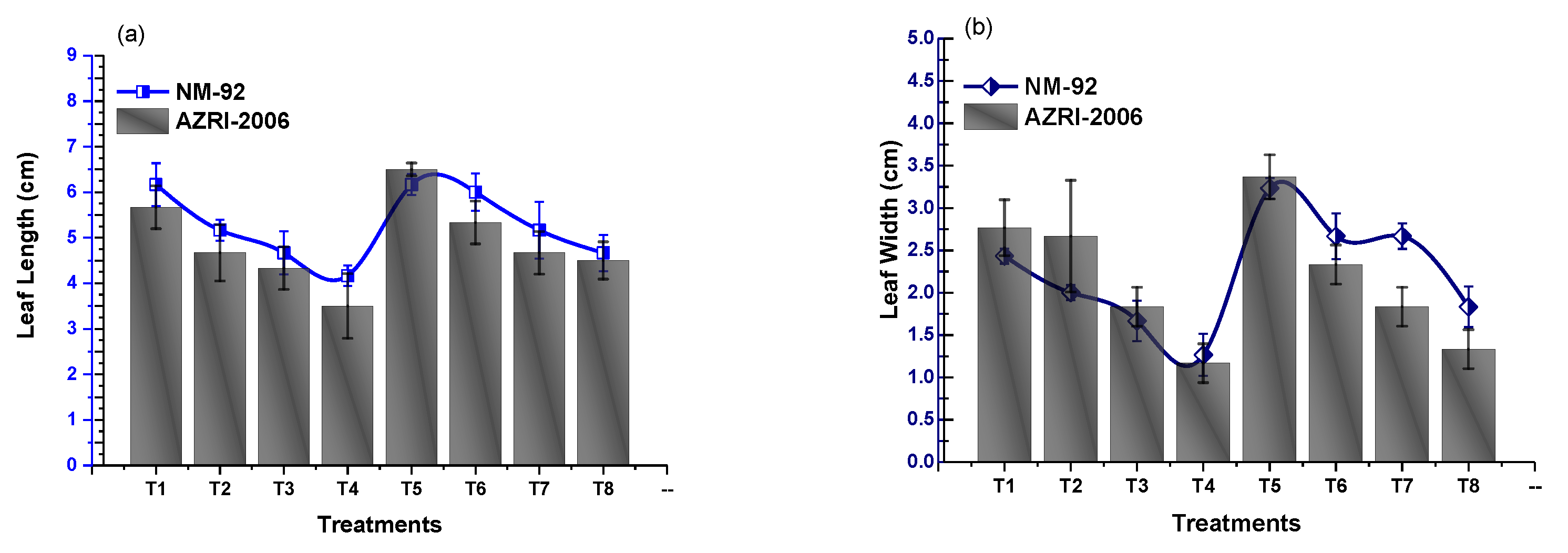
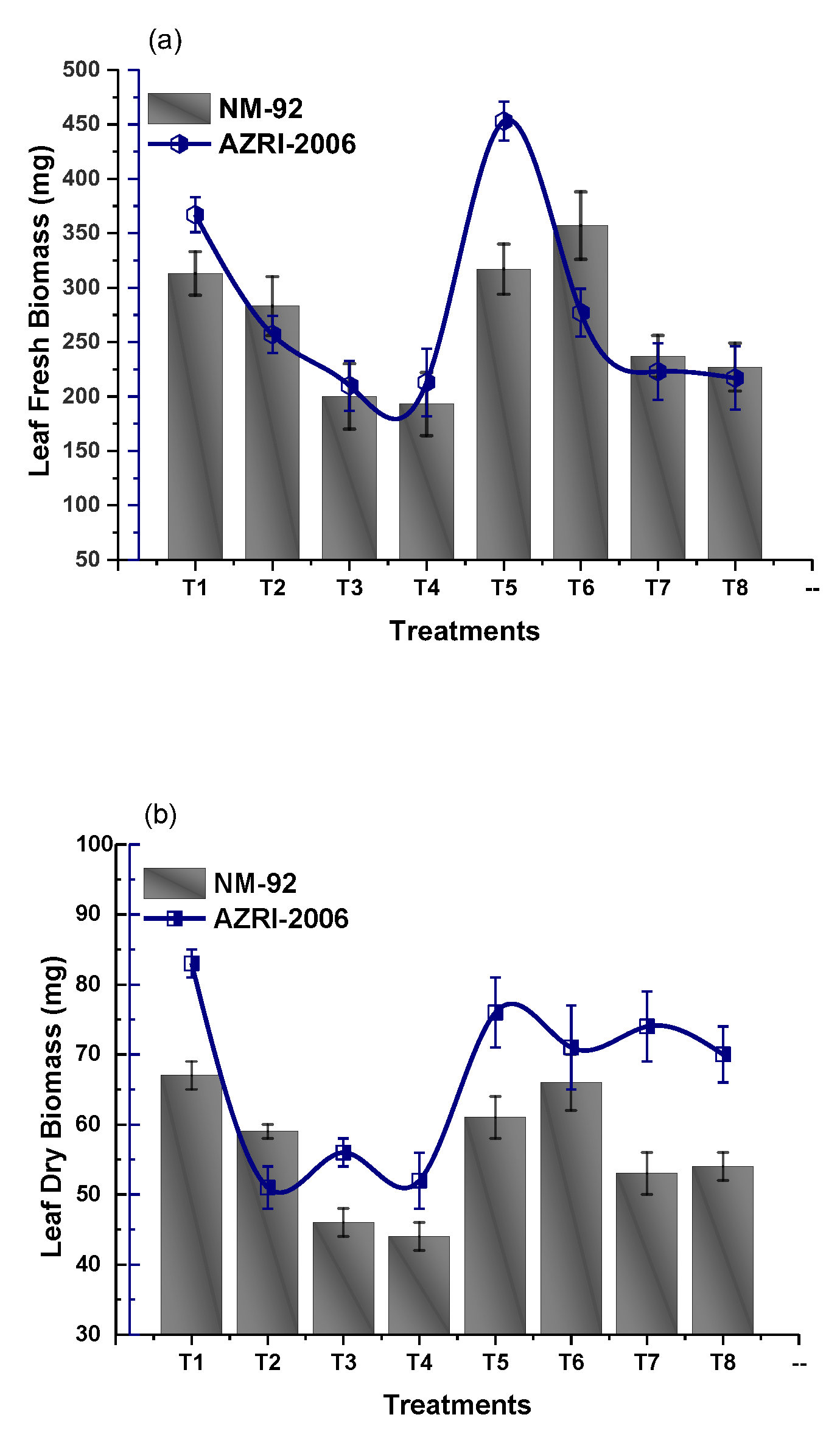
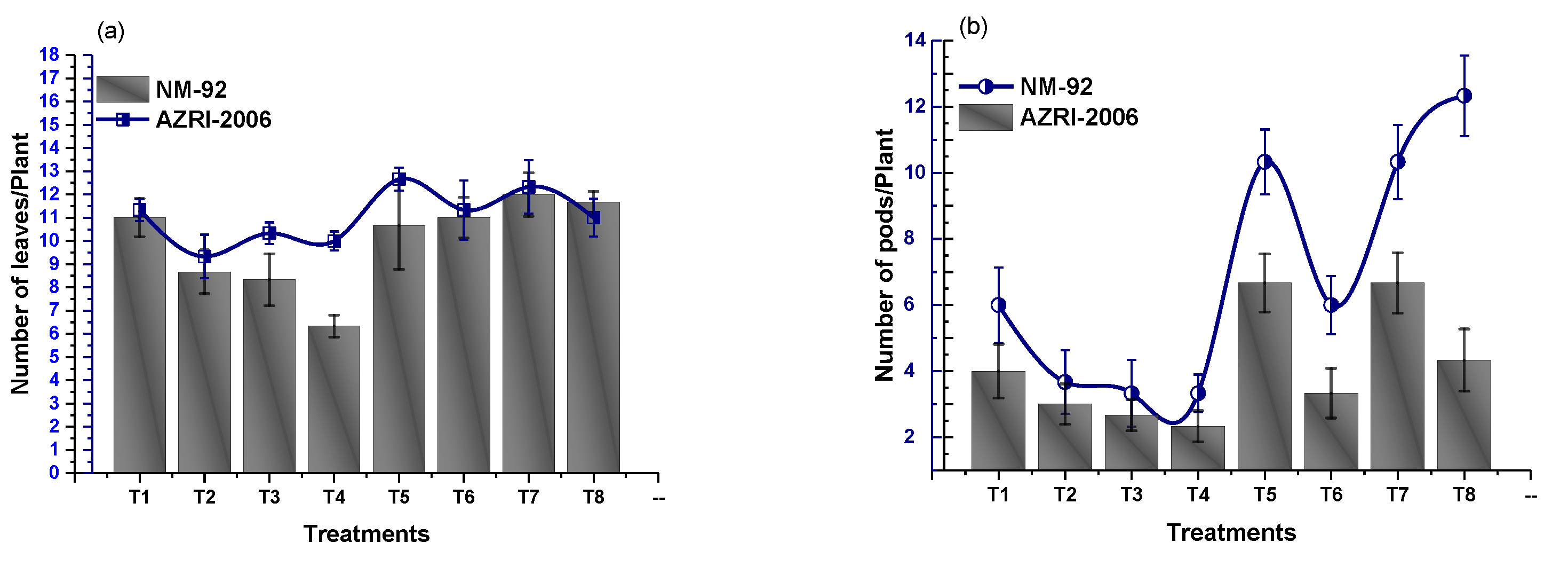
3.2. Effect of Glu-FeNPs on Growth Performance of Mung Bean
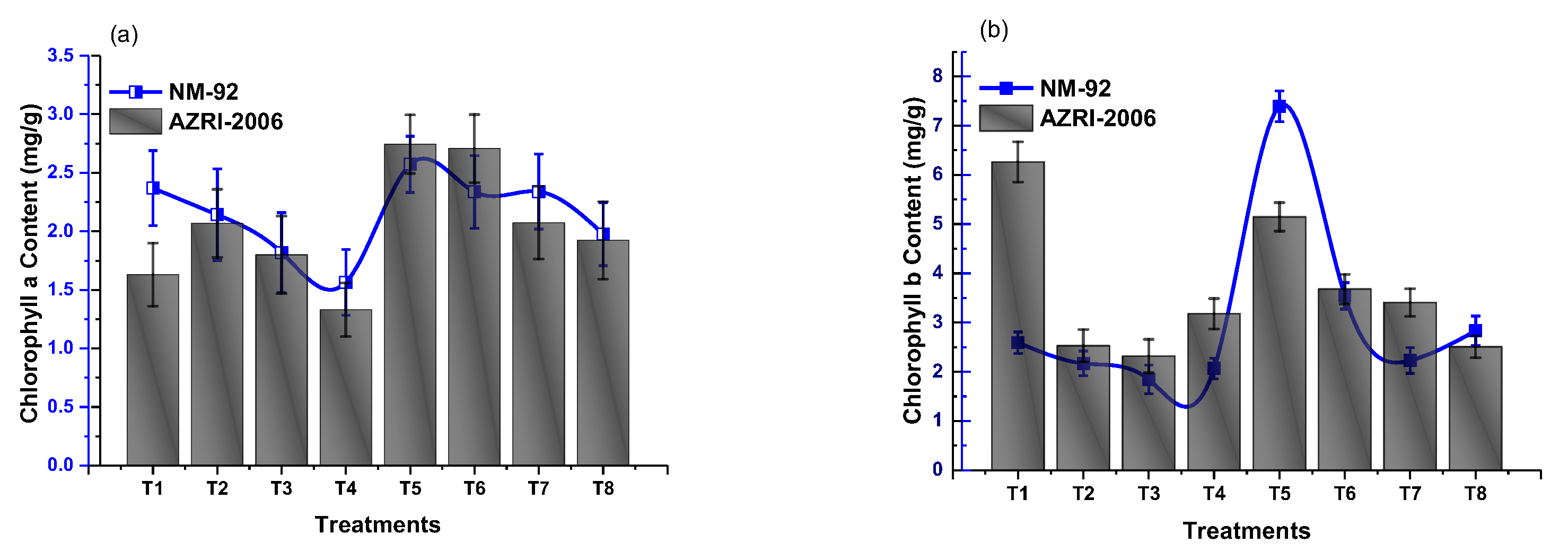
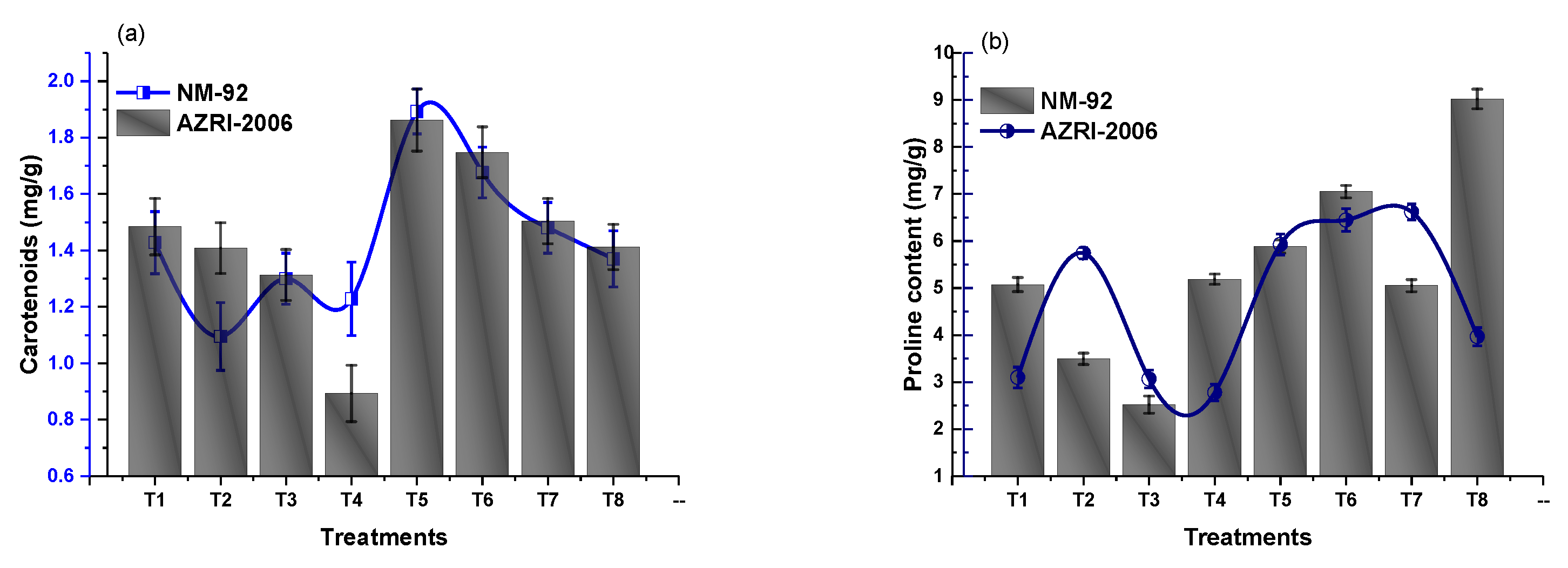
3.3. Effect of Glu-FeNPs on Biochemical Marker
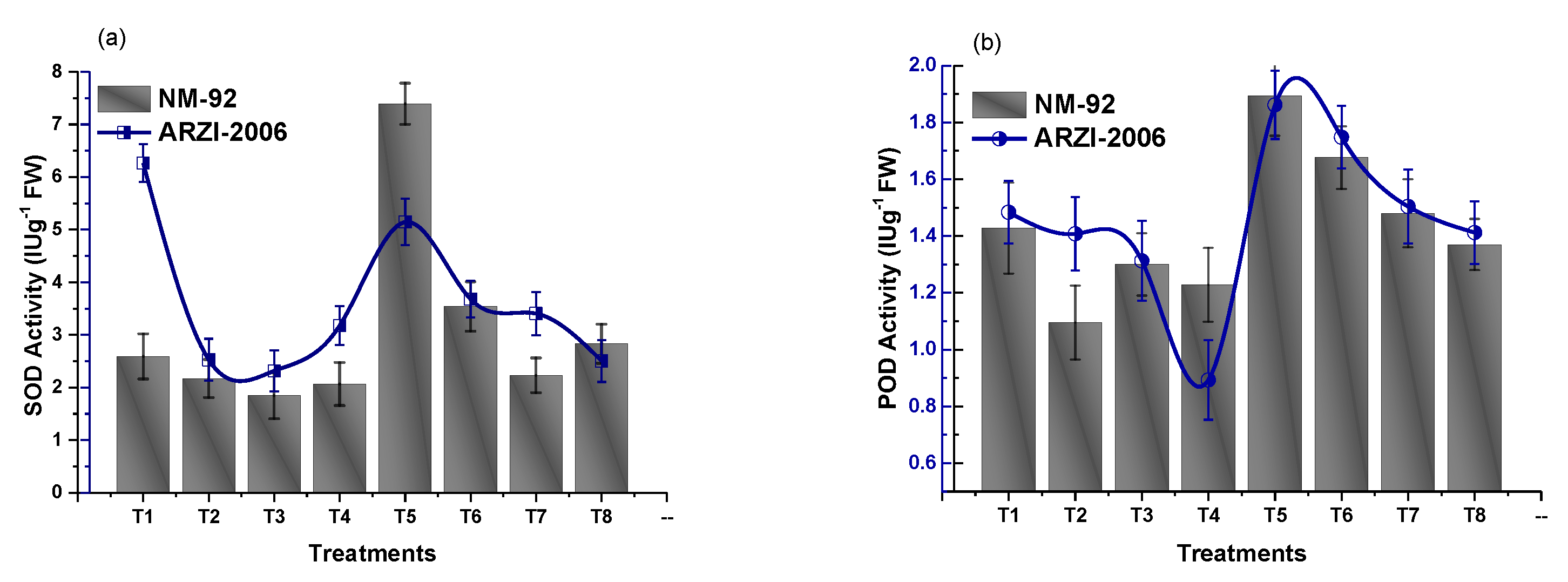
3.4. Effect on Antioxidant Enzymes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, Y.H.; Ying, Y.; Chen, J.; Wang, Y.C. Transgenic Arabidopsis over expressing Mn-SOD enhanced salt-tolerance. Plant Sci. 2014, 167, 671–677. [Google Scholar] [CrossRef]
- GoP. Economic Survey; Government of Pakistan, Finance Division, Economic Adviser’s Wing: Islamabad, Pakistan, 2005; p. 233. [Google Scholar]
- Adjel, F.; Kadi, Z.; Bouzerzour, H.; Benmahammed, A. Salt stress effects on seeds germination and seedling growth of Barley genotypes. J. Agric. Sustain. 2013, 3, 223–237. [Google Scholar]
- Carillo, P.; Cirillo, C.; DeMicco, V.; Arena, C.; Depascale, S.; Rouphael, Y. Morpho-anatomical, physiological and biochemical adaptive responses to saline water of Bougainvillea spectabilis Willd. trained to different canopy shapes. Agric. Water Manag. 2019, 212, 12–22. [Google Scholar] [CrossRef]
- Zeeshan, M.; Meiqin, L.; Shafaque, S.; Paul, H.; Feibo, W. Drain water management for salinity mitigation in irrigated agriculture. Agronomy 2020, 10, 127. [Google Scholar] [CrossRef] [Green Version]
- Rashidzadeh, A.; Olad, A. Slow-released NPK fertilizer encapsulated bNaAlg-g-poly(AA-co-AAm)/MMT superabsorbent nanocomposite. Carbohydr. Polym. 2014, 114, 269–278. [Google Scholar] [CrossRef]
- Jampílek, J.; Kráľová, K.; Novák, P.; Novák, M. Nanobiotechnology in Neurodegenerative Diseases; Springer: Cham, Switzerland, 2019; pp. 65–138. [Google Scholar]
- Chinnamuthu, C.R.; Boopathi, P.M. Nanotechnology and Agroecosystem. Madras. Agric. J. 2019, 96, 17–31. [Google Scholar]
- Baruah, S.; Dutta, J. Nanotechnology applications in sensing and pollution degradation in agriculture. Environ. Chem. Letters J. 2018, 7, 191–204. [Google Scholar] [CrossRef]
- Abbas, G.; Hussain, F.; Anwar, Z.; Khattak, J.Z.K.; Ishaque, U.; Asmat, U. Effects of Iron on the Wheat Crop (Triticum aestivum L.) by Uptake of nitrogen, phosphorus and potassium. Agric. Sci. 2012, 4, 229–235. [Google Scholar]
- Jiang, Z.I.; Liu, C.Y.; Sun, L.W. Catalytic properties of silver nanoparticles supported on silica spheres. J. Am. Chem. Soc. 2014, 71, 2341–2343. [Google Scholar] [CrossRef]
- Haq, T.; Ullah, S.; Ullah, R. Beneficial Effects of Several Nanoparticles on the Growth of Different Plants Species. Curr. Nanosci. 2019, 15, 460–470. [Google Scholar] [CrossRef]
- Frewer, L.J.; Norde, W.; Fischer, A.R.H.; Kampers, F.W.H. Nanotechnology in theAgri-Food Sector: Implications for the Future; Wiley-VCH: Weinheim, Germany, 2015; p. 455. [Google Scholar]
- Perez-de-Luque, A.; Hermosín, M.C.; Bagchi, D.; Bagchi, M.; Moriyama, H. Nanotechnology and Its Use in Agriculture; Wiley-Blackwell: West Sussex, UK, 2013; pp. 299–405. [Google Scholar]
- Elshamy, M.T.; El-Khallal, S.T.; Husseiny, S.M.; Farroh, K.Y. Catalytic properties of Zinc oxide nanoparticles. J. Sci. Res. Sci. 2019, 36, 424–441. [Google Scholar]
- Haq, T.; Ullah, R. Green synthesis and characterization of gold nanoparticles (Au-NPs) using stem extract of Euphorbia neriifolia and evaluation of their antibacterial and antifungal potential. Int. J. Nanosci. 2022, 21, 2250008. [Google Scholar] [CrossRef]
- Mahmoodi, P.; Yarnia, M.; Rashidi, V.; Amirnia, R.; Tarinejhad, A. Positive effect of zinc oxide nanoparticels on tomato plants: A step towards developing “Nano-fertilizers’. Appl. Ecol. Environ. Res. 2018, 16, 4773–4788. [Google Scholar] [CrossRef]
- Kalteh, M.; Alipour, M.Z.; Ashraf, S.; Aliabadi, M.M.; Nosratabadi, A.F. Effect of silica Nanoparticles on Basil (Ocimum basilicum) Under Salinity Stress. J. Chem. Health Risks 2014, 3, 49–55. [Google Scholar]
- Raliya, R.; Biswas, P.; Tarafdar, J.C. TiO2 nanoparticle biosynthesis and its physiological effect on mung bean (Vigna radiata L.). Biotechnol. Rep. 2015, 5, 22–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanatt, S.; Sharma, R.A.K. Antioxidant andantimicrobial activity of legume hulls. Food Res Int. 2011, 44, 3182–3187. [Google Scholar] [CrossRef]
- Imran, K.; Hussain, I.; Khattak, I.; Rehman, A.U.; Ahmad, F.; Zada, H.; Naveed, S. Effect of Iron Nutrient Care Sprayed on Foliage at Different Physiological Growth Stages on Yield and Quality of Some Durum Wheat (Triticum durum L.) varieties in Sandy Soil. Pure Appl. Biol. 2015, 4, 557–567. [Google Scholar]
- Nair, R.; Varghese, S.H.; Nair, B.G.; Maekawa, T.; Yoshida, Y.; Kumar, S.D. Nanoparticulate material delivery to plants. Plant Sci. 2018, 179, 154–163. [Google Scholar] [CrossRef]
- Min, L. Research Advance in Chemical Composionand Pharmacological Action of Mung Bean. Trad Chin. Med. 2017, 5, 18. [Google Scholar]
- Siva, C.V.; Benita, L.F. Iron Oxide Nanoparticles Promotes Agronomic Traits of Ginger (Zingiber officinale Rosc.). Int. J. Adv. Res. Biol. Sci. 2015, 3, 230–237. [Google Scholar]
- Moghadam, A.L.; Vattani, H.; Baghaei, N.; Keshavarz, N. Effect of different levels of fertilizer nanoiron chelates on growth and yield characteristics of twovarieties of spinach (Spinacia oleracea L.): Varamin 88 and viroflay. Res. J. Appl. Sci. Eng. Technol. 2017, 4, 4813–4818. [Google Scholar]
- Srivastava, G.; Das, C.K.; Das, A.; Singh, S.K.; Roy, M.; Kim, H.; Sethy, N.; Kumar, A.; Sharma, S.K.; Singh, R.K.; et al. Seed treatment with iron pyrite (FeS2) nanoparticles increases the production of spinach. RSC Adv. 2014, 4, 58495–58504. [Google Scholar] [CrossRef]
- Askary, M.S.; Talebi, M.; Amini, F. Efect of NaCl and iron oxide nanoparticles on Mentha piperita essential oil composition. Environ. Exp. Biol. 2016, 14, 27–32. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugar and related substances. Anal. Chem. 2016, 28, 350–356. [Google Scholar] [CrossRef]
- Rostami, F.; Ehsanpour, A.A. Application of silver thiosulfate (STS) on silver accumulation and protein pattern of potato (Solanumtubersum L.) under in vitro culture. Ann. Appl. Biol. 2009, 38, 46–54. [Google Scholar]
- Gholami, M.; Rahemi, M.; Kholdebarin, B.; Rastegar, S. Use of rapid screening methods for detecting drought tolerance cultivars of Fig. (Ficus carica L.). Sci. Hortic. Amst. 2012, 148, 109–117. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2012, 7, 405–410. [Google Scholar] [CrossRef]
- Padgett, P.E.; Leonard, R.T. Free amino acid levels and the regulation of nitrate uptake in maize cell suspension cultures. J. Exp. Bot. 1996, 47, 871–883. [Google Scholar] [CrossRef] [Green Version]
- Shekari, G.; Javanmardi, J. Effects of Foliar Application Pure Amino Acid and Amino Acid Containing Fertilizer on Broccoli (Brassica oleracea L. var. italica) Transplant. Adv. Crop. Sci. Tech. 2017, 5, 280. [Google Scholar] [CrossRef]
- Davies, P.J. Plant Hormones: Biosynthesis, Signal Transduction, Action! Springer: New York, NY, USA, 2004. [Google Scholar]
- Bahari, A.; Pirdashti, H.; Yaghubi, M. The effects of amino acid fertilizers spraying on photosynthetic pigments and antioxidant enzymes of wheat (Triticum aestivum L.) under salinity stress. Int. J. Agron. Plant Prod. 2013, 4, 787–793. [Google Scholar]
- Briat, J.F.; Curie, C.; Grignon, F. Iron utilization and metabolism in plants. Curr. Opin. Plant Biol. 2007, 10, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Rout, G.R.; Sahoo, S. Role of iron in plants growth and metabolism. Rev. Agri. Sci. 2015, 3, 124. [Google Scholar] [CrossRef] [Green Version]
- Ali, E.A. Seed germination and seedling growth of Suaeda salsa under salt stress. Asian J. Crop. Sci. 2012, 4, 139–149. [Google Scholar] [CrossRef] [Green Version]
- Bameri, M.; Abdolshahi, R.; Mohammadi-Nejad, G.; Yousefi, U.; Tabatabaie, S.M. Effect of different microelement treatment on wheat (Triticum aestivum) growth and yield. Intl. Res. J. Appl. Basic. Sci. 2013, 3, 219–223. [Google Scholar]
- Hu, J.; Guo, H.; Li, J.; Gan, Q.; Xing, B. omparative impacts of iron oxide nanoparticles and ferric ions on the growth of Citrus maxima. Environ. Pollut. 2017, 221, 199–208. [Google Scholar] [CrossRef] [Green Version]
- Rui, M.; Ma, C.; Hao, Y.; Guo, J.; Rui, Y.; Tang, X.; Zhao, Q.; Fan, X.; Zhang, Z.; Hou, T.; et al. Iron Oxide Nanoparticles as a Potential Iron Fertilizer for Peanut (Arachis hypogaea). Front. Plant Sci. 2016, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, T.; Nishizawa, N.K. Iron uptake, translocation and regulation in higher plants. Annu. Rev. Plant Biol. 2012, 63, 131–152. [Google Scholar] [CrossRef] [Green Version]
- Racuciu, M.; Creanga, D.E. Biocompatible magnetic fluid nanoparticles internalized in vegetal tissue. J. Phys. 2019, 54, 115–124. [Google Scholar]
- Hu, C.A.; Delauney, A.G.; Verma, D.P.S.A. A bifunctional enzyme (delta 1pyrroline-5-carboxylate synthetase) catalyzes the first two steps in proline biosynthesis in plants. Proc. Natl. Acad. Sci. USA 2018, 89, 9354–9358. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.; Shahzad, B.; Kumar, V. Phytohormones regulate accumulation of osmolytes under abiotic stress. Biomolecules 2019, 9, 285. [Google Scholar] [CrossRef] [Green Version]
- Ashraf, M.; Foolad, M.R. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ Exp Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Bowler, T.; Torres, F.I.A.; Terner, J.; Pittman, R.N.; Proffitt, E.; Ward, K.R. Measurement of hemoglobin oxygen saturation using Raman microscopy and 532-nm excitation. J. Appl. Physiol. 2016, 104, 1809–1817. [Google Scholar]
- Takahashi, M.A.; Asada, K. Superoxide anion permeability of phospholipid membrane and chloroplast thylakoids. Arch. Biochem. Bhiophys. 1983, 226, 558–566. [Google Scholar] [CrossRef] [PubMed]
- Das, C.K.; Jangir, H.; Kumar, J.; Verma, S. Sustainable design for NKP equivalent rice production. Nanotechnol. Environ. Eng. 2018, 3, 14. [Google Scholar] [CrossRef]
- Alexandre, M.A.; Abdel-Fattah, G.H.; Shahin, S.M. Magnetic (Fe3O4) nanoparticles reduce heavy metal uptake and mitigate their toxicity in wheat seedling. Sustainability 2017, 9, 790. [Google Scholar]
- Harinasut, P.; Poonsopa, D.; Roengmongkol, K.; Charoensataporn, R. Salinity effects on antioxidant enzymes in mulberry cultivar. Sci. Asia 2003, 29, 109–113. [Google Scholar] [CrossRef]
- Song, F.; Yang, C.; Liu, X.; Li, G. Effect of salt stress on activity of superoxide dismutase (SOD) in Ulmus pumila L. J. For. Res. 2006, 17, 13–16. [Google Scholar] [CrossRef]
















{kind=link}
{kind=link}
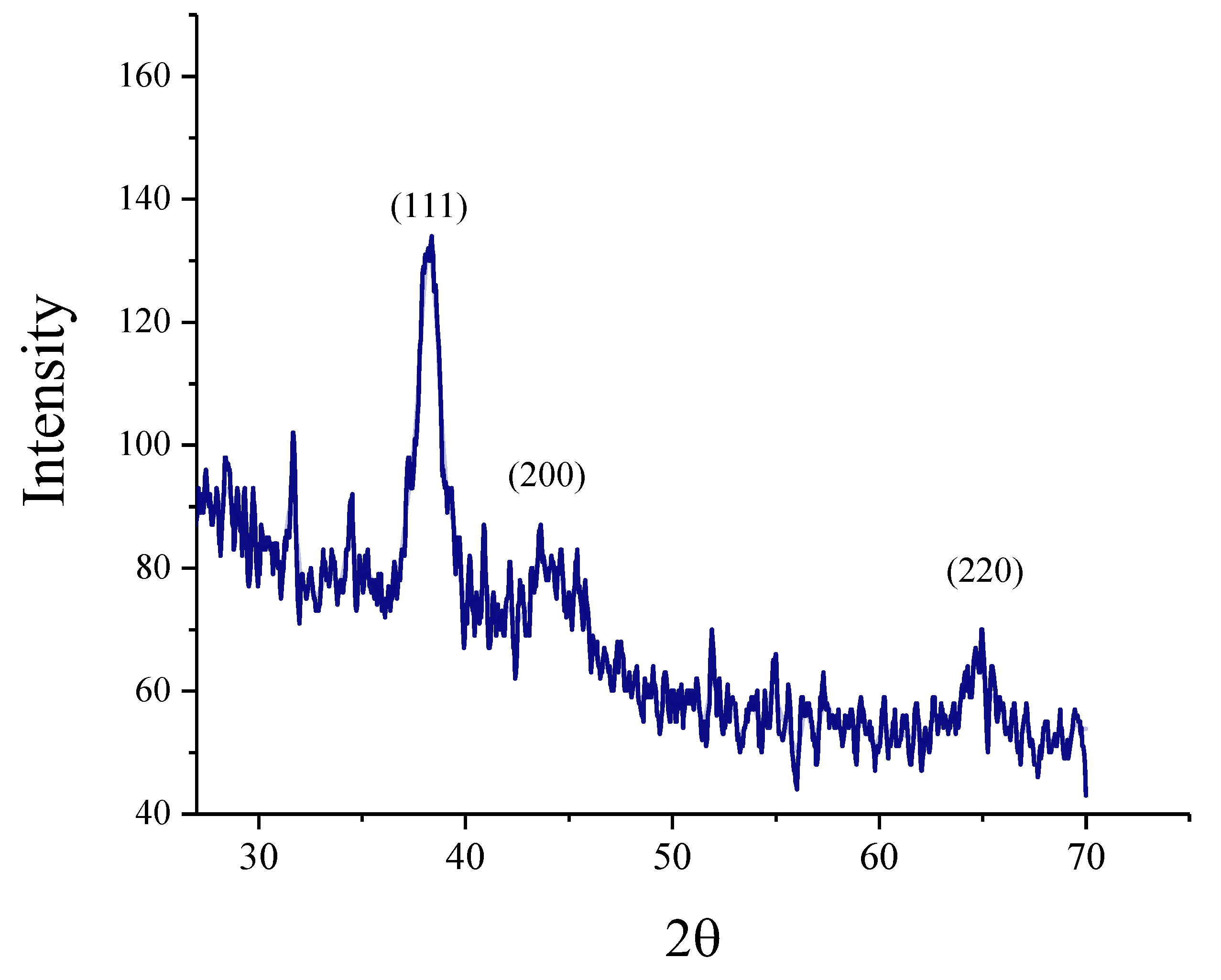{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Salinity Concentration | Glu-FeNPs |
|---|---|---|
| T1 (Control) | Untreated | Untreated |
| T2 | 40 mM | Untreated |
| T3 | 60 Mm | Untreated |
| T4 | 80 Mm | Untreated |
| T5 | Untreated | 150 mg/L |
| T6 | 40 Mm | 150 mg/L |
| T7 | 60 Mm | 150 mg/L |
| T8 | 80 Mm | 150 mg/L |
| Variables | DF | SS | MS | F | P | Sig | CI |
|---|---|---|---|---|---|---|---|
| Plumule Length | 15 | 85.66 | 5.71 | 4.83 | 0.0001 | *** | 95% |
| Radical | 15 | 73.18 | 4.88 | 5.15 | 0.0001 | *** | 95% |
| Shoot Length | 15 | 507.91 | 33.86 | 4.64 | 0.0001 | *** | 95% |
| Shoot Fresh Biomass | 15 | 1.90 | 0.12 | 4.03 | 0.0005 | *** | 95% |
| Shoot Dry Biomass | 15 | 0.88 | 0.06 | 3.23 | 0.0026 | ** | 95% |
| Root Length | 15 | 92.07 | 6.13 | 2.17 | 0.0312 | * | 95% |
| Root Fresh Biomass | 15 | 1.85 | 0.12 | 1.91 | 0.0618 | NS | 95% |
| Root Dry Biomass | 15 | 0.052 | 0.003 | 5.05 | 0.0001 | *** | 95% |
| No. of Leaves/Plant | 15 | 121.91 | 8.12 | 4.24 | 0.0003 | *** | 95% |
| Leaf Length | 15 | 33.71 | 2.24 | 4.11 | 0.0003 | *** | 95% |
| Leaf Width | 15 | 19.14 | 1.27 | 8.38 | 0.0000 | *** | 95% |
| Leaf Area | 15 | 560.08 | 37.33 | 1.64 | 0.1164 | NS | 95% |
| Leaf Fresh Biomass | 15 | 2.76 | 0.18 | 2.05 | 0.0417 | * | 95% |
| Leaf Dry Biomass | 15 | 0.71 | 0.04 | 0.63 | 0.8270 | NS | 95% |
| No. of Pods/Plant | 15 | 529.66 | 35.31 | 5.20 | 0.0000 | *** | 95% |
| Protein Content | 15 | 254.37 | 16.95 | 3.33 | 0.0019 | ** | 95% |
| Proline Content | 15 | 148.33 | 9.88 | 2.27 | 0.0243 | * | 95% |
| Total soluble Sugar | 15 | 76.73 | 5.11 | 1.44 | 0.1874 | NS | 95% |
| Chlorophyll a | 15 | 7.56 | 0.50 | 1.72 | 0.0956 | NS | 95% |
| Chlorophyll b | 15 | 114.98 | 7.66 | 2.31 | 0.0221 | * | 95% |
| Carotenoids | 15 | 3.14 | 0.20 | 5.11 | 0.0000 | *** | 95% |
| SOD | 15 | 7.02 | 0.46 | 1.29 | 0.2609 | NS | 95% |
| POD | 15 | 0.96 | 0.06 | 0.32 | 0.9894 | NS | 95% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ul Haq, T.; Ullah, R.; Khan, M.N.; Nazish, M.; Almutairi, S.M.; Rasheed, R.A. Seed Priming with Glutamic-Acid-Functionalized Iron Nanoparticles Modulating Response of Vigna radiata (L.) R. Wilczek (Mung Bean) to Induce Osmotic Stress. Micromachines 2023, 14, 736. https://doi.org/10.3390/mi14040736
Ul Haq T, Ullah R, Khan MN, Nazish M, Almutairi SM, Rasheed RA. Seed Priming with Glutamic-Acid-Functionalized Iron Nanoparticles Modulating Response of Vigna radiata (L.) R. Wilczek (Mung Bean) to Induce Osmotic Stress. Micromachines. 2023; 14(4):736. https://doi.org/10.3390/mi14040736
Chicago/Turabian StyleUl Haq, Tauheed, Rehman Ullah, Muhammad Nauman Khan, Moona Nazish, Saeedah Musaed Almutairi, and Rabab Ahmed Rasheed. 2023. "Seed Priming with Glutamic-Acid-Functionalized Iron Nanoparticles Modulating Response of Vigna radiata (L.) R. Wilczek (Mung Bean) to Induce Osmotic Stress" Micromachines 14, no. 4: 736. https://doi.org/10.3390/mi14040736