
An Introduction to Nanopore Sequencing: Past, Present, and Future Considerations

{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Nanopore Sequencing Principles
3. Biological Nanopores
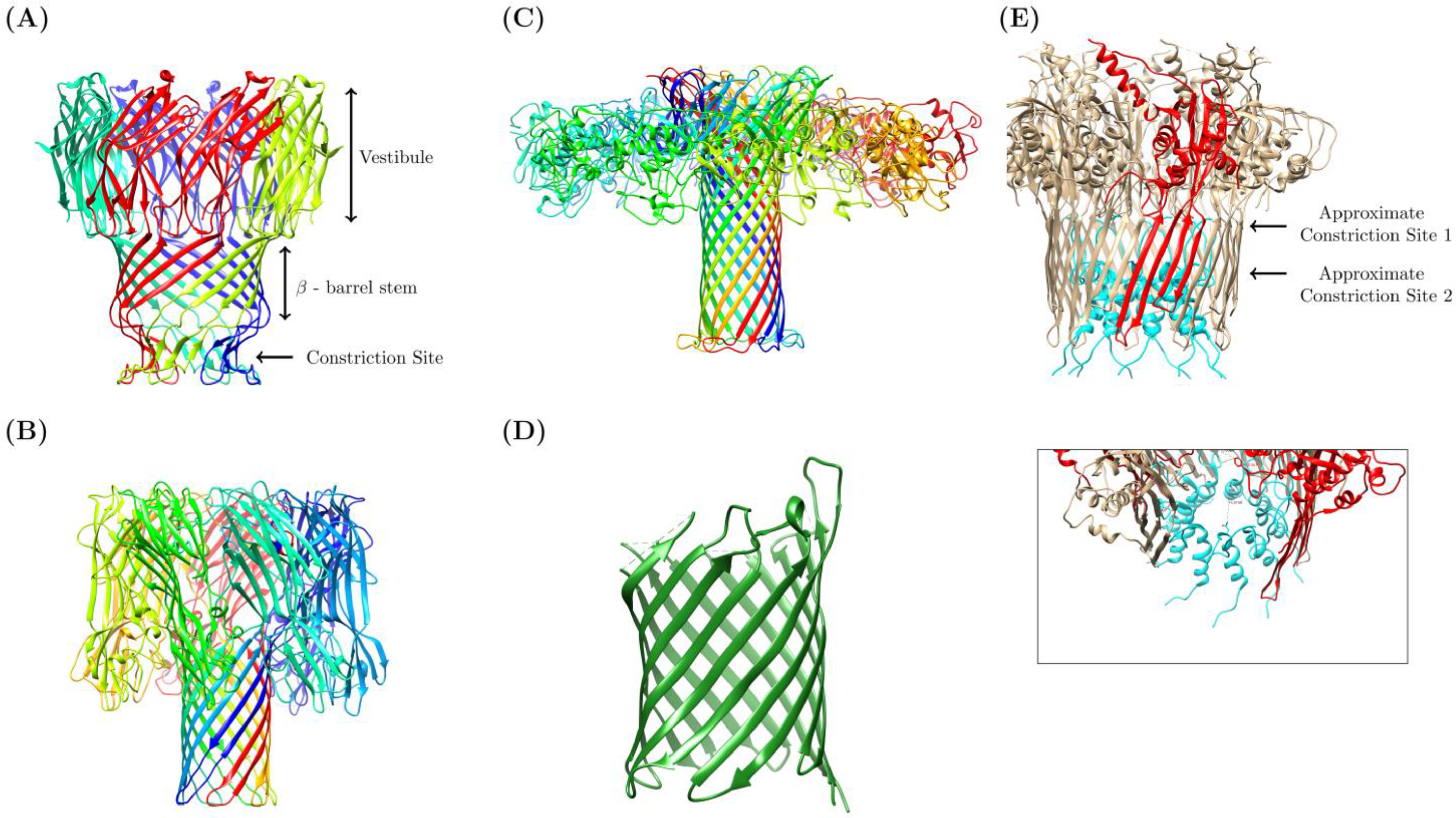
3.1. Biological Nanopore Variants
3.2. Slowing Translocation Speeds in Biological Nanopores
4. Solid-State Nanopores
4.1. Fabrication Techniques
4.2. Materials for Construction
4.3. Controlling Noise and Translocation Speeds in Solid-State Nanopores
5. Oxford Nanopore Technologies vs. Other NGS Platforms
5.1. Short vs. Long Reads
5.2. ONT vs. PacBio
6. Library Preparation Considerations for Nucleic Acid Sequencing
6.1. Library Preparation Overview
6.2. Library Preparation Modifications for Diverse Samples
7. Clinical Applications: Nucleic Acid Sequencing on Nanopores
7.1. Genomics and Structural Variants
7.1.1. Genome Assembly
7.1.2. Fusion Gene Detection
7.1.3. Short Tandem Repeat Detection
7.1.4. DNA Nicks
7.2. Epigenetic Modifications
7.3. Infectious Disease Detection
7.4. RNA Sequencing
7.4.1. Full-Length Transcript Assembly
7.4.2. MicroRNA Detection and Quantification
7.4.3. CircRNAs
7.4.4. Single Cell RNA Sequencing
8. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Shi, W.; Friedman, A.K.; Baker, L.A. Nanopore Sensing. Anal. Chem. 2017, 89, 157–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Company History. Available online: https://nanoporetech.com/about-us/history (accessed on 14 January 2023).
- Song, L.; Hobaugh, M.R.; Shustak, C.; Cheley, S.; Cheley, S.; Cheley, S.; Bayley, H.; Gouaux, J.E.; Gouaux, J.E. Structure of Staphylococcal α-Hemolysin, a Heptameric Transmembrane Pore. Science 1996, 274, 1859–1866. [Google Scholar] [CrossRef] [PubMed]
- Kasianowicz, J.J.; Brandin, E.; Branton, D.; Deamer, D.W. Characterization of Individual Polynucleotide Molecules Using a Membrane Channel. Proc. Natl. Acad. Sci. USA 1996, 93, 13770–13773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Branton, D.; Deamer, D.W.; Marziali, A.; Bayley, H.; Benner, S.A.; Butler, T.; Di Ventra, M.; Garaj, S.; Hibbs, A.; Huang, X.; et al. The Potential and Challenges of Nanopore Sequencing. Nat. Biotechnol. 2008, 26, 1146–1153. [Google Scholar] [CrossRef]
- Deamer, D.; Akeson, M.; Branton, D. Three Decades of Nanopore Sequencing. Nat. Biotechnol. 2016, 34, 518–524. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Wu, L.; Qiao, Y.; Tu, J.; Lu, Z. Microfluidic Systems Applied in Solid-State Nanopore Sensors. Micromachines 2020, 11, 332. [Google Scholar] [CrossRef] [Green Version]
- Stoloff, D.H.; Wanunu, M. Recent Trends in Nanopores for Biotechnology. Curr. Opin. Biotechnol. 2013, 24, 699–704. [Google Scholar] [CrossRef] [Green Version]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera: A Visualization System for Exploratory Research and Analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [Green Version]
- How Nanopore Sequencing Works. Available online: https://nanoporetech.com/support/how-it-works (accessed on 14 January 2023).
- Deamer, D.W.; David, W.; Branton, D. Characterization of Nucleic Acids by Nanopore Analysis. Acc. Chem. Res. 2002, 35, 817–825. [Google Scholar] [CrossRef] [Green Version]
- Ashkenasy, N.; Sánchez-Quesada, J.; Bayley, H.; Ghadiri, M.R. Recognizing a Single Base in an Individual DNA Strand: A Step Toward DNA Sequencing in Nanopores. Angew Chem Int Ed Engl. 2005, 44, 1401–1404. [Google Scholar] [CrossRef] [Green Version]
- Stoddart, D.; Maglia, G.; Mikhailova, E.; Heron, A.J.; Bayley, H. Multiple Base-Recognition Sites in a Biological Nanopore: Two Heads Are Better than One. Angew. Chem. Int. Ed. 2010, 49, 556–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iacovache, I.; De Carlo, S.; Cirauqui, N.; Dal Peraro, M.; van der Goot, F.G.; Zuber, B. Cryo-EM Structure of Aerolysin Variants Reveals a Novel Protein Fold and the Pore-Formation Process. Nat. Commun. 2016, 7, 12062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, C.; Ying, Y.-L.; Hu, Z.-L.; Liao, D.-F.; Tian, H.; Long, Y.-T. Discrimination of Oligonucleotides of Different Lengths with a Wild-Type Aerolysin Nanopore. Nat. Nanotechnol. 2016, 11, 713–718. [Google Scholar] [CrossRef] [PubMed]
- Liang, B.; Tamm, L.K. Structure of Outer Membrane Protein G by Solution NMR Spectroscopy. Proc. Natl. Acad. Sci. USA 2007, 104, 16140–16145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.; Khalid, S.; Mark, S.P.; Sansom, M.S.P.; Bayley, H. Outer Membrane Protein G: Engineering a Quiet Pore for Biosensing. Proc. Natl. Acad. Sci. USA 2008, 105, 6272–6277. [Google Scholar] [CrossRef] [Green Version]
- Sanganna Gari, R.R.; Seelheim, P.; Liang, B.; Tamm, L.K. Quiet Outer Membrane Protein G (OmpG) Nanopore for Biosensing. ACS Sens. 2019, 4, 1230–1235. [Google Scholar] [CrossRef]
- Faller, M.; Niederweis, M.; Schulz, G.E. The Structure of a Mycobacterial Outer-Membrane Channel. Science 2004, 303, 1189–1192. [Google Scholar] [CrossRef]
- Manrao, E.A.; Derrington, I.M.; Pavlenok, M.; Niederweis, M.; Gundlach, J.H. Nucleotide Discrimination with DNA Immobilized in the MspA Nanopore. PLoS ONE 2011, 6, e25723. [Google Scholar] [CrossRef] [Green Version]
- Manrao, E.A.; Derrington, I.M.; Laszlo, A.H.; Langford, K.W.; Hopper, M.K.; Gillgren, N.; Pavlenok, M.; Niederweis, M.; Gundlach, J.H. Reading DNA at Single-Nucleotide Resolution with a Mutant MspA Nanopore and Phi29 DNA Polymerase. Nat. Biotechnol. 2012, 30, 349–353. [Google Scholar] [CrossRef] [Green Version]
- Craig, J.M.; Laszlo, A.H.; Brinkerhoff, H.; Derrington, I.M.; Noakes, M.T.; Nova, I.C.; Tickman, B.I.; Doering, K.; de Leeuw, N.F.; Gundlach, J.H. Revealing Dynamics of Helicase Translocation on Single-Stranded DNA Using High-Resolution Nanopore Tweezers. Proc. Natl. Acad. Sci. USA 2017, 114, 11932–11937. [Google Scholar] [CrossRef] [Green Version]
- Brown, C.G.; Clarke, J. Nanopore Development at Oxford Nanopore. Nat. Biotechnol. 2016, 34, 810–811. [Google Scholar] [CrossRef] [PubMed]
- Carter, J.M.; Hussain, S. Robust long-read native DNA sequencing using the ONT CsgG Nanopore. Wellcome Open Res. 2018, 2, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Stephenson, W.; Razaghi, R.; Busan, S.; Weeks, K.M.; Timp, W.; Smibert, P. Direct Detection of RNA Modifications and Structure Using Single-Molecule Nanopore Sequencing. Cell Genom. 2022, 2, 100097. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Shi, H.; Zhang, X.; Zhang, X.; Huang, Y. Cryo-EM Structure of the Nonameric CsgG-CsgF Complex and Its Implications for Controlling Curli Biogenesis in Enterobacteriaceae. PLoS Biol. 2020, 18, e3000748. [Google Scholar] [CrossRef] [PubMed]
- Van der Verren, S.E.; Van Gerven, N.; Jonckheere, W.; Hambley, R.; Singh, P.; Kilgour, J.; Jordan, M.; Wallace, E.J.; Jayasinghe, L.; Remaut, H. A Dual-Constriction Biological Nanopore Resolves Homonucleotide Sequences with High Fidelity. Nat. Biotechnol. 2020, 38, 1415–1420. [Google Scholar] [CrossRef] [PubMed]
- Derrington, I.M.; Craig, J.M.; Stava, E.; Laszlo, A.H.; Ross, B.C.; Brinkerhoff, H.; Nova, I.C.; Doering, K.; Tickman, B.I.; Ronaghi, M.; et al. Subangstrom Single-Molecule Measurements of Motor Proteins Using a Nanopore. Nat. Biotechnol. 2015, 33, 1073–1075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caldwell, C.C.; Spies, M. Helicase SPRNTing through the Nanopore. Proc. Natl. Acad. Sci. USA 2017, 114, 11809–11811. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhao, Y.; Bollas, A.; Wang, Y.; Au, K.F. Nanopore Sequencing Technology, Bioinformatics and Applications. Nat. Biotechnol. 2021, 39, 1348–1365. [Google Scholar] [CrossRef]
- Xue, L.; Yamazaki, H.; Ren, R.; Wanunu, M.; Ivanov, A.P.; Edel, J.B. Solid-State Nanopore Sensors. Nat. Rev. Mater. 2020, 5, 931–951. [Google Scholar] [CrossRef]
- Mohapatra, S.; Lin, C.-T.; Feng, X.A.; Basu, A.; Ha, T. Single-Molecule Analysis and Engineering of DNA Motors. Chem. Rev. 2020, 120, 36–78. [Google Scholar] [CrossRef]
- Li, J.; Stein, D.; McMullan, C.J.; Branton, D.; Aziz, M.J.; Golovchenko, J.A. Ion-Beam Sculpting at Nanometre Length Scales. Nature 2001, 412, 166–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fürjes, P. Controlled Focused Ion Beam Milling of Composite Solid State Nanopore Arrays for Molecule Sensing. Micromachines 2019, 10, 774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muhammad Sajeer, P.; Nukala, P.; Manoj, M. Varma TEM Based Applications in Solid State Nanopores: From Fabrication to Liquid in-Situ Bio-Imaging. Micron 2022, 162, 103347. [Google Scholar] [CrossRef] [PubMed]
- Bafna, J.A.; Soni, G.V. Fabrication of Low Noise Borosilicate Glass Nanopores for Single Molecule Sensing. PLoS ONE 2016, 11, e0157399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Bell, N.A.W.; Hernández-Ainsa, S.; Thacker, V.V.; Thackray, A.M.; Bujdoso, R.; Keyser, U.F. Single Protein Molecule Detection by Glass Nanopores. ACS Nano 2013, 7, 4129–4134. [Google Scholar] [CrossRef]
- Chen, Z.; Jiang, Y.; Dunphy, D.R.; Adams, D.P.; Hodges, C.; Liu, N.; Zhang, N.; Xomeritakis, G.; Jin, X.; Aluru, N.R.; et al. DNA Translocation through an Array of Kinked Nanopores. Nat. Mater. 2010, 9, 667–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waugh, M.; Briggs, K.; Gunn, D.; Gibeault, M.; King, S.; Ingram, Q.; Jimenez, A.M.; Berryman, S.; Lomovtsev, D.; Andrzejewski, L.; et al. Solid-State Nanopore Fabrication by Automated Controlled Breakdown. Nat. Protoc. 2020, 15, 122–143. [Google Scholar] [CrossRef]
- Kwok, H.; Briggs, K.; Tabard-Cossa, V. Nanopore Fabrication by Controlled Dielectric Breakdown. PLoS ONE 2014, 9, e92880. [Google Scholar] [CrossRef] [Green Version]
- Deng, T.; Li, M.; Wang, Y.; Liu, Z. Development of Solid-State Nanopore Fabrication Technologies. Sci. Bull. 2015, 60, 304–319. [Google Scholar] [CrossRef]
- Fried, J.P.; Swett, J.L.; Nadappuram, B.P.; Mol, J.A.; Edel, J.B.; Ivanov, A.P.; Yates, J.R. In Situ Solid-State Nanopore Fabrication. Chem. Soc. Rev. 2021, 50, 4974–4992. [Google Scholar] [CrossRef]
- Lei, X.; Zhang, J.; Hong, H.; Yuan, Z.; Liu, Z. Controllable Shrinking Fabrication of Solid-State Nanopores. Micromachines 2022, 13, 923. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Gershow, M.; Stein, D.; Brandin, E.; Golovchenko, J.A. DNA Molecules and Configurations in a Solid-State Nanopore Microscope. Nat. Mater. 2003, 2, 611–615. [Google Scholar] [CrossRef] [PubMed]
- Song, B.; Schneider, G.F.; Xu, Q.; Pandraud, G.; Dekker, C.; Zandbergen, H. Atomic-Scale Electron-Beam Sculpting of Near-Defect-Free Graphene Nanostructures. Nano Lett. 2011, 11, 2247–2250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Storm, A.J.; Chen, J.; Ling, X.S.; Zandbergen, H.W.; Zandbergen, H.W.; Dekker, C. Fabrication of Solid-State Nanopores with Single-Nanometre Precision. Nat. Mater. 2003, 2, 537–540. [Google Scholar] [CrossRef] [PubMed]
- Storm, A.J.; Chen, J.H.; Ling, X.S.; Zandbergen, H.W.; Dekker, C. Electron-Beam-Induced Deformations of SiO2 Nanostructures. J. Appl. Phys. 2005, 98, 014307. [Google Scholar] [CrossRef] [Green Version]
- van den Hout, M.; Hall, A.R.; Wu, M.Y.; Zandbergen, H.W.; Dekker, C.; Dekker, N.H. Controlling Nanopore Size, Shape and Stability. Nanotechnology 2010, 21, 115304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tahvildari, R.; Beamish, E.; Tabard-Cossa, V.; Godin, M. Integrating Nanopore Sensors within Microfluidic Channel Arrays Using Controlled Breakdown. Lab. Chip 2015, 15, 1407–1411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Yang, Q.; Wang, Z. The Evolution of Nanopore Sequencing. Front. Genet. 2015, 5, 449. [Google Scholar] [CrossRef] [Green Version]
- Fragasso, A.; Schmid, S.; Dekker, C. Comparing Current Noise in Biological and Solid-State Nanopores. ACS Nano 2020, 14, 1338–1349. [Google Scholar] [CrossRef] [Green Version]
- Eggenberger, O.M.; Ying, C.; Mayer, M. Surface Coatings for Solid-State Nanopores. Nanoscale 2019, 11, 19636–19657. [Google Scholar] [CrossRef] [Green Version]
- Wasfi, A.; Awwad, F.; Ayesh, A.I. Graphene-Based Nanopore Approaches for DNA Sequencing: A Literature Review. Biosens. Bioelectron. 2018, 119, 191–203. [Google Scholar] [CrossRef] [PubMed]
- Venta, K.; Shemer, G.; Puster, M.; Rodríguez-Manzo, J.A.; Balan, A.; Rosenstein, J.K.; Shepard, K.; Drndić, M. Differentiation of Short, Single-Stranded DNA Homopolymers in Solid-State Nanopores. ACS Nano 2013, 7, 4629–4636. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Manzo, J.A.; Puster, M.; Nicolaï, A.; Meunier, V.; Drndić, M. DNA Translocation in Nanometer Thick Silicon Nanopores. ACS Nano 2015, 9, 6555–6564. [Google Scholar] [CrossRef] [PubMed]
- Chien, C.-C.; Shekar, S.; Niedzwiecki, D.J.; Shepard, K.L.; Drndić, M. Single-Stranded DNA Translocation Recordings through Solid-State Nanopores on Glass Chips at 10 MHz Measurement Bandwidth. ACS Nano 2019, 13, 10545–10554. [Google Scholar] [CrossRef] [PubMed]
- Heerema, S.J.; Dekker, C. Graphene Nanodevices for DNA Sequencing. Nat. Nanotechnol. 2016, 11, 127–136. [Google Scholar] [CrossRef] [Green Version]
- Garaj, S.; Liu, S.; Golovchenko, J.A.; Branton, D. Molecule-Hugging Graphene Nanopores. Proc. Natl. Acad. Sci. USA 2013, 110, 12192–12196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prasongkit, J.; Jungthawan, S.; Amorim, R.G.; Scheicher, R.H. Single-Molecule DNA Sequencing Using Two-Dimensional Ti2C(OH)2 MXene Nanopores: A First-Principles Investigation. Nano Res. 2022, 15, 9843–9849. [Google Scholar] [CrossRef]
- Smeets, R.M.M.; Keyser, U.F.; Wu, M.Y.; Dekker, N.H.; Dekker, C. Nanobubbles in Solid-State Nanopores. Phys. Rev. Lett. 2006, 97, 088101. [Google Scholar] [CrossRef] [Green Version]
- Jia, Z.; Choi, J.; Park, S. Surface Charge Density-Dependent DNA Capture through Polymer Planar Nanopores. ACS Appl. Mater. Interfaces 2018, 10, 40927–40937. [Google Scholar] [CrossRef]
- Elibol, K.; Susi, T.; O′Brien, M.; Bayer, B.C.; Pennycook, T.J.; McEvoy, N.; Duesberg, G.S.; Meyer, J.C.; Kotakoski, J. Grain Boundary-Mediated Nanopores in Molybdenum Disulfide Grown by Chemical Vapor Deposition. Nanoscale 2017, 9, 1591–1598. [Google Scholar] [CrossRef] [Green Version]
- Asatekin, A.; Gleason, K.K. Polymeric Nanopore Membranes for Hydrophobicity-Based Separations by Conformal Initiated Chemical Vapor Deposition. Nano Lett. 2011, 11, 677–686. [Google Scholar] [CrossRef] [PubMed]
- Lepoitevin, M.; Ma, T.; Bechelany, M.; Janot, J.-M.; Balme, S. Functionalization of Single Solid State Nanopores to Mimic Biological Ion Channels: A Review. Adv. Colloid Interface Sci. 2017, 250, 195–213. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Tang, H.; Yang, C.; Li, Y. Selective Single Molecule Nanopore Sensing of MicroRNA Using PNA Functionalized Magnetic Core–Shell Fe 3 O 4 –Au Nanoparticles. Anal. Chem. 2019, 91, 7965–7970. [Google Scholar] [CrossRef] [PubMed]
- Kircher, M.; Kelso, J. High-Throughput DNA Sequencing—Concepts and Limitations. BioEssays 2010, 32, 524–536. [Google Scholar] [CrossRef] [PubMed]
- Ambardar, S.; Gupta, R.; Trakroo, D.; Lal, R.; Vakhlu, J. High Throughput Sequencing: An Overview of Sequencing Chemistry. Indian J. Microbiol. 2016, 56, 394–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, T.; Chitnis, N.; Monos, D.; Dinh, A. Next-Generation Sequencing Technologies: An Overview. Hum. Immunol. 2021, 82, 801–811. [Google Scholar] [CrossRef] [PubMed]
- Bronner, I.F.; Quail, M.A. Best Practices for Illumina Library Preparation. Curr. Protoc. Hum. Genet. 2019, 102, e86. [Google Scholar] [CrossRef]
- Rhoads, A.; Au, K.F. PacBio Sequencing and Its Applications. Genom. Proteom. Bioinform. 2015, 13, 278–289. [Google Scholar] [CrossRef] [Green Version]
- Nurk, S.; Koren, S.; Rhie, A.; Rautiainen, M.; Bzikadze, A.V.; Mikheenko, A.; Vollger, M.R.; Altemose, N.; Uralsky, L.; Gershman, A.; et al. The Complete Sequence of a Human Genome. Science 2022, 76, 44–53. [Google Scholar] [CrossRef]
- Jain, M.; Olsen, H.E.; Paten, B.; Akeson, M. The Oxford Nanopore MinION: Delivery of Nanopore Sequencing to the Genomics Community. Genome Biol. 2016, 17, 239. [Google Scholar] [CrossRef] [Green Version]
- Dippenaar, A.; Goossens, S.N.; Grobbelaar, M.; Oostvogels, S.; Cuypers, B.; Laukens, K.; Meehan, C.J.; Warren, R.M.; van Rie, A. Nanopore Sequencing for Mycobacterium Tuberculosis: A Critical Review of the Literature, New Developments, and Future Opportunities. J. Clin. Microbiol. 2022, 60, e00646-21. [Google Scholar] [CrossRef] [PubMed]
- Karst, S.M.; Ziels, R.M.; Kirkegaard, R.H.; Sørensen, E.A.; McDonald, D.; Zhu, Q.; Knight, R.; Albertsen, M. High-Accuracy Long-Read Amplicon Sequences Using Unique Molecular Identifiers with Nanopore or PacBio Sequencing. Nat. Methods 2021, 18, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Amarasinghe, S.L.; Su, S.; Dong, X.; Zappia, L.; Ritchie, M.E.; Gouil, Q. Opportunities and Challenges in Long-Read Sequencing Data Analysis. Genome Biol. 2020, 21, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hon, T.; Mars, K.; Young, G.; Tsai, Y.-C.; Karalius, J.W.; Landolin, J.M.; Maurer, N.; Kudrna, D.; Hardigan, M.A.; Steiner, C.C.; et al. Highly Accurate Long-Read HiFi Sequencing Data for Five Complex Genomes. Sci. Data 2020, 7, 399. [Google Scholar] [CrossRef] [PubMed]
- Wenger, A.M.; Peluso, P.; Rowell, W.J.; Chang, P.-C.; Hall, R.J.; Concepcion, G.T.; Ebler, J.; Fungtammasan, A.; Kolesnikov, A.; Olson, N.D.; et al. Accurate Circular Consensus Long-Read Sequencing Improves Variant Detection and Assembly of a Human Genome. Nat. Biotechnol. 2019, 37, 1155–1162. [Google Scholar] [CrossRef] [PubMed]
- Logsdon, G.A.; Vollger, M.R.; Eichler, E.E. Long-Read Human Genome Sequencing and Its Applications. Nat. Rev. Genet. 2020, 21, 597–614. [Google Scholar] [CrossRef] [PubMed]
- Jain, M.; Koren, S.; Miga, K.H.; Quick, J.; Rand, A.C.; Sasani, T.A.; Tyson, J.R.; Beggs, A.D.; Dilthey, A.T.; Fiddes, I.T.; et al. Nanopore Sequencing and Assembly of a Human Genome with Ultra-Long Reads. Nat. Biotechnol. 2018, 36, 338–345. [Google Scholar] [CrossRef] [Green Version]
- Grünberger, F.; Ferreira-Cerca, S.; Grohmann, D. Nanopore Sequencing of RNA and CDNA Molecules in Escherichia Coli. RNA 2022, 28, 400–417. [Google Scholar] [CrossRef]
- DNA and RNA Sequencing Kits. Available online: https://nanoporetech.com/products/kits (accessed on 14 January 2023).
- Oikonomopoulos, S.; Bayega, A.; Fahiminiya, S.; Djambazian, H.; Berube, P.; Ragoussis, J. Methodologies for Transcript Profiling Using Long-Read Technologies. Front. Genet. 2020, 11, 606. [Google Scholar] [CrossRef]
- Wahl, A.; Huptas, C.; Neuhaus, K. Comparison of RRNA Depletion Methods for Efficient Bacterial MRNA Sequencing. Sci. Rep. 2022, 12, 5765. [Google Scholar] [CrossRef]
- Kraus, A.J.; Brink, B.G.; Siegel, T.N. Efficient and Specific Oligo-Based Depletion of RRNA. Sci. Rep. 2019, 9, 12281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDonald, T.L.; Zhou, W.; Castro, C.P.; Mumm, C.; Switzenberg, J.A.; Mills, R.E.; Boyle, A.P. Cas9 Targeted Enrichment of Mobile Elements Using Nanopore Sequencing. Nat. Commun. 2021, 12, 3586. [Google Scholar] [CrossRef] [PubMed]
- Stangl, C.; de Blank, S.; Renkens, I.; Westera, L.; Verbeek, T.; Valle-Inclan, J.E.; González, R.C.; Henssen, A.G.; van Roosmalen, M.J.; Stam, R.W.; et al. Partner Independent Fusion Gene Detection by Multiplexed CRISPR-Cas9 Enrichment and Long Read Nanopore Sequencing. Nat. Commun. 2020, 11, 2861. [Google Scholar] [CrossRef] [PubMed]
- Giraldez, M.D.; Spengler, R.M.; Etheridge, A.; Godoy, P.M.; Barczak, A.J.; Srinivasan, S.; De Hoff, P.L.; Tanriverdi, K.; Courtright, A.; Lu, S.; et al. Comprehensive Multi-Center Assessment of Small RNA-Seq Methods for Quantitative MiRNA Profiling. Nat. Biotechnol. 2018, 36, 746–757. [Google Scholar] [CrossRef]
- Feng, Y.; Cohen, S.N. Unpaired Terminal Nucleotides and 5′ Monophosphorylation Govern 3′ Polyadenylation by Escherichia Coli Poly(A) Polymerase I. Proc. Natl. Acad. Sci. USA 2000, 97, 6415–6420. [Google Scholar] [CrossRef] [Green Version]
- MacKenzie, M.; Tigert, S.; Lovato, D.; Mir, H.; Zahedi, K.; Barone, S.L.; Brooks, M.; Soleimani, M.; Argyropoulos, C. To Make a Short Story Long: Simultaneous Short and Long RNA Profiling on Nanopore Devices. bioRxiv 2022, 2022.12.16.520507. [Google Scholar] [CrossRef]
- Gleeson, J.; Leger, A.; Prawer, Y.D.J.; Lane, T.A.; Harrison, P.J.; Haerty, W.; Clark, M.B. Accurate Expression Quantification from Nanopore Direct RNA Sequencing with NanoCount. Nucleic Acids Res. 2022, 50, e19. [Google Scholar] [CrossRef]
- Li, Y.; Gutiérrez Moreno, J.J.; Song, Z.; Liu, D.; Wang, M.; Ramiere, A.; Feng, Z.; Niu, Q.J.; Sasaki, T.; Cai, X. Controlled Synthesis of Perforated Oxide Nanosheets with High Density Nanopores Showing Superior Water Purification Performance. ACS Appl. Mater. Interfaces 2022, 14, 18513–18524. [Google Scholar] [CrossRef]
- Zhang, S.; Huang, G.; Versloot, R.C.A.; Bruininks, B.M.H.; de Souza, P.C.T.; Marrink, S.-J.; Maglia, G. Bottom-up Fabrication of a Proteasome–Nanopore That Unravels and Processes Single Proteins. Nat. Chem. 2021, 13, 1192–1199. [Google Scholar] [CrossRef]
- Chen, K.; Kong, J.; Zhu, J.; Ermann, N.; Predki, P.; Keyser, U.F. Digital Data Storage Using DNA Nanostructures and Solid-State Nanopores. Nano Lett. 2019, 19, 1210–1215. [Google Scholar] [CrossRef]
- Ying, Y.-L.; Hu, Z.-L.; Zhang, S.; Qing, Y.; Fragasso, A.; Maglia, G.; Meller, A.; Bayley, H.; Dekker, C.; Long, Y.-T. Nanopore-Based Technologies beyond DNA Sequencing. Nat. Nanotechnol. 2022, 17, 1136–1146. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Ru, K.; Zhang, L.; Huang, Y.; Zhu, X.; Liu, H.; Zetterberg, A.; Cheng, T.; Miao, W. Fluorescence in Situ Hybridization (FISH): An Increasingly Demanded Tool for Biomarker Research and Personalized Medicine. Biomark. Res. 2014, 2, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeck, W.R.; Lee, J.; Robinson, H.; Le, L.P.; Iafrate, A.J.; Nardi, V. A Nanopore Sequencing–Based Assay for Rapid Detection of Gene Fusions. J. Mol. Diagn. 2019, 21, 58–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.Y.; Kong, M.; Oh, J.; Lim, J.; Chung, S.H.; Kim, J.-M.; Kim, J.-S.; Kim, K.-H.; Yoo, J.-C.; Kwak, W. Comparative Evaluation of Nanopore Polishing Tools for Microbial Genome Assembly and Polishing Strategies for Downstream Analysis. Sci. Rep. 2021, 11, 20740. [Google Scholar] [CrossRef]
- Jeck, W.R.; Iafrate, A.J.; Nardi, V. Nanopore Flongle Sequencing as a Rapid, Single-Specimen Clinical Test for Fusion Detection. J. Mol. Diagn. 2021, 23, 630–636. [Google Scholar] [CrossRef]
- Fang, L.; Liu, Q.; Monteys, A.M.; Gonzalez-Alegre, P.; Davidson, B.L.; Wang, K. DeepRepeat: Direct Quantification of Short Tandem Repeats on Signal Data from Nanopore Sequencing. Genome Biol. 2022, 23, 108. [Google Scholar] [CrossRef]
- Ren, Z.-L.; Zhang, J.-R.; Zhang, X.-M.; Liu, X.; Lin, Y.-F.; Bai, H.; Wang, M.-C.; Cheng, F.; Liu, J.-D.; Li, P.; et al. Forensic Nanopore Sequencing of STRs and SNPs Using Verogen’s ForenSeq DNA Signature Prep Kit and MinION. Int. J. Legal Med. 2021, 135, 1685–1693. [Google Scholar] [CrossRef]
- Tytgat, O.; Škevin, S.; Deforce, D.; Van Nieuwerburgh, F. Nanopore Sequencing of a Forensic Combined STR and SNP Multiplex. Forensic Sci. Int. Genet. 2022, 56, 102621. [Google Scholar] [CrossRef]
- Kinkar, L.; Gasser, R.; Webster, B.; Rollinson, D.; Littlewood, D.; Chang, B.; Stroehlein, A.; Korhonen, P.; Young, N. Nanopore Sequencing Resolves Elusive Long Tandem-Repeat Regions in Mitochondrial Genomes. Int. J. Mol. Sci. 2021, 22, 1811. [Google Scholar] [CrossRef]
- Perešíni, P.; Boža, V.; Brejová, B.; Vinař, T. Nanopore Base Calling on the Edge. Bioinformatics 2021, 37, 4661–4667. [Google Scholar] [CrossRef]
- Athreya, N.; Milenkovic, O.; Leburton, J.-P. Interaction Dynamics and Site-Specific Electronic Recognition of DNA-Nicks with 2D Solid-State Nanopores. Npj 2D Mater. Appl. 2020, 4, 32. [Google Scholar] [CrossRef]
- Robertson, K.D. DNA Methylation and Human Disease. Nat. Rev. Genet. 2005, 6, 597–610. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Hu, Y.; Zhou, B.; Bao, Y.; Li, Z.; Gong, C.; Yang, H.; Wang, S.; Xiao, Y. The Role of M6A Modification in Physiology and Disease. Cell Death Dis. 2020, 11, 960. [Google Scholar] [CrossRef] [PubMed]
- Laird, P.W. Principles and Challenges of Genome-Wide DNA Methylation Analysis. Nat. Rev. Genet. 2010, 11, 191–203. [Google Scholar] [CrossRef] [PubMed]
- Schatz, M.C. Nanopore Sequencing Meets Epigenetics. Nat. Methods 2017, 14, 347–348. [Google Scholar] [CrossRef]
- Davenport, C.F.; Scheithauer, T.; Dunst, A.; Bahr, F.S.; Dorda, M.; Wiehlmann, L.; Tran, D.D.H. Genome-Wide Methylation Mapping Using Nanopore Sequencing Technology Identifies Novel Tumor Suppressor Genes in Hepatocellular Carcinoma. Int. J. Mol. Sci. 2021, 22, 3937. [Google Scholar] [CrossRef]
- Liu, Y.; Rosikiewicz, W.; Pan, Z.; Jillette, N.; Wang, P.; Taghbalout, A.; Foox, J.; Mason, C.; Carroll, M.; Cheng, A.; et al. DNA Methylation-Calling Tools for Oxford Nanopore Sequencing: A Survey and Human Epigenome-Wide Evaluation. Genome Biol. 2021, 22, 295. [Google Scholar] [CrossRef]
- Rand, A.C.; Jain, M.; Eizenga, J.M.; Musselman-Brown, A.; Olsen, H.E.; Akeson, M.; Paten, B. Mapping DNA Methylation with High-Throughput Nanopore Sequencing. Nat. Methods 2017, 14, 411–413. [Google Scholar] [CrossRef] [Green Version]
- Petersen, L.M.; Martin, I.W.; Moschetti, W.E.; Kershaw, C.M.; Tsongalis, G.J. Third-Generation Sequencing in the Clinical Laboratory: Exploring the Advantages and Challenges of Nanopore Sequencing. J. Clin. Microbiol. 2019, 58, e01315-19. [Google Scholar] [CrossRef]
- Quick, J.; Loman, N.J.; Duraffour, S.; Simpson, J.T.; Severi, E.; Cowley, L.; Bore, J.A.; Koundouno, R.; Dudas, G.; Mikhail, A.; et al. Real-Time, Portable Genome Sequencing for Ebola Surveillance. Nature 2016, 530, 228–232. [Google Scholar] [CrossRef] [Green Version]
- Wang, J. MinION Nanopore Sequencing of an Influenza Genome. Front. Microbiol. 2015, 6, 766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faria, N.R.; Quick, J.; Claro, I.M.; Thézé, J.; de Jesus, J.G.; Giovanetti, M.; Kraemer, M.U.G.; Hill, S.C.; Black, A.; da Costa, A.C.; et al. Establishment and Cryptic Transmission of Zika Virus in Brazil and the Americas. Nature 2017, 546, 406–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yakovleva, A.; Kovalenko, G.; Redlinger, M.; Liulchuk, M.G.; Bortz, E.; Zadorozhna, V.I.; Scherbinska, A.M.; Wertheim, J.O.; Goodfellow, I.; Meredith, L.; et al. Tracking SARS-CoV-2 Variants Using Nanopore Sequencing in Ukraine in 2021. Sci. Rep. 2022, 12, 15749. [Google Scholar] [CrossRef] [PubMed]
- Vandenbogaert, M.; Kwasiborski, A.; Gonofio, E.; Descorps-Declère, S.; Selekon, B.; Nkili Meyong, A.A.; Ouilibona, R.S.; Gessain, A.; Manuguerra, J.-C.; Caro, V.; et al. Nanopore Sequencing of a Monkeypox Virus Strain Isolated from a Pustular Lesion in the Central African Republic. Sci. Rep. 2022, 12, 10768. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.-T.; Liu, P.-Y.; Shih, P.-W. Homopolish: A Method for the Removal of Systematic Errors in Nanopore Sequencing by Homologous Polishing. Genome Biol. 2021, 22, 95. [Google Scholar] [CrossRef]
- Bayega, A.; Oikonomopoulos, S.; Gregoriou, M.-E.; Tsoumani, K.T.; Giakountis, A.; Wang, Y.C.; Mathiopoulos, K.D.; Ragoussis, J. Nanopore Long-Read RNA-Seq and Absolute Quantification Delineate Transcription Dynamics in Early Embryo Development of an Insect Pest. Sci. Rep. 2021, 11, 7878. [Google Scholar] [CrossRef]
- Zhang, J.; Yan, S.; Chang, L.; Guo, W.; Wang, Y.; Wang, Y.; Zhang, P.; Chen, H.-Y.; Huang, S. Direct MicroRNA Sequencing Using Nanopore-Induced Phase-Shift Sequencing. iScience 2020, 23, 100916. [Google Scholar] [CrossRef] [Green Version]
- Smith, A.M.; Abu-Shumays, R.; Akeson, M.; Bernick, D.L. Capture, Unfolding, and Detection of Individual TRNA Molecules Using a Nanopore Device. Front. Bioeng. Biotechnol. 2015, 3, 91. [Google Scholar] [CrossRef] [Green Version]
- Thomas, N.K.; Poodari, V.C.; Jain, M.; Olsen, H.E.; Akeson, M.; Abu-Shumays, R.L. Direct Nanopore Sequencing of Individual Full Length TRNA Strands. ACS Nano 2021, 15, 16642–16653. [Google Scholar] [CrossRef]
- Liu, Z.; Tao, C.; Li, S.; Du, M.; Bai, Y.; Hu, X.; Li, Y.; Chen, J.; Yang, E. CircFL-Seq Reveals Full-Length Circular RNAs with Rolling Circular Reverse Transcription and Nanopore Sequencing. eLife 2021, 10, e69457. [Google Scholar] [CrossRef]
- Workman, R.E.; Tang, A.D.; Tang, P.S.; Jain, M.; Tyson, J.R.; Razaghi, R.; Zuzarte, P.C.; Gilpatrick, T.; Payne, A.; Quick, J.; et al. Nanopore Native RNA Sequencing of a Human Poly(A) Transcriptome. Nat. Methods 2019, 16, 1297–1305. [Google Scholar] [CrossRef] [PubMed]
- Depledge, D.P.; Srinivas, K.P.; Sadaoka, T.; Bready, D.; Mori, Y.; Placantonakis, D.G.; Mohr, I.; Wilson, A.C. Direct RNA Sequencing on Nanopore Arrays Redefines the Transcriptional Complexity of a Viral Pathogen. Nat. Commun. 2019, 10, 754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soneson, C.; Yao, Y.; Bratus-Neuenschwander, A.; Patrignani, A.; Robinson, M.D.; Hussain, S. A Comprehensive Examination of Nanopore Native RNA Sequencing for Characterization of Complex Transcriptomes. Nat. Commun. 2019, 10, 3359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibrahim, F.; Oppelt, J.; Maragkakis, M.; Mourelatos, Z. TERA-Seq: True End-to-End Sequencing of Native RNA Molecules for Transcriptome Characterization. Nucleic Acids Res. 2021, 49, e115. [Google Scholar] [CrossRef] [PubMed]
- Hackl, T.; Hedrich, R.; Schultz, J.; Förster, F. Proovread: Large-Scale High-Accuracy PacBio Correction through Iterative Short Read Consensus. Bioinformatics 2014, 30, 3004–3011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaffi, S.K.; Galas, D.; Etheridge, A.; Argyropoulos, C. Role of MicroRNAs in Renal Parenchymal Diseases—A New Dimension. Int. J. Mol. Sci. 2018, 19, 1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vieux, K.-F.; Prothro, K.P.; Kelley, L.H.; Palmer, C.; Maine, E.M.; Veksler-Lublinsky, I.; McJunkin, K. Screening by Deep Sequencing Reveals Mediators of MicroRNA Tailing in C. Elegans. Nucleic Acids Res. 2021, 49, 11167–11180. [Google Scholar] [CrossRef]
- Yang, A.; Bofill-De Ros, X.; Shao, T.-J.; Jiang, M.; Li, K.; Villanueva, P.; Dai, L.; Gu, S. 3′ Uridylation Confers MiRNAs with Non-Canonical Target Repertoires. Mol. Cell 2019, 75, 511–522.e4. [Google Scholar] [CrossRef]
- Wang, Y.; Zheng, D.; Tan, Q.; Wang, M.X.; Gu, L.-Q. Nanopore-Based Detection of Circulating MicroRNAs in Lung Cancer Patients. Nat. Nanotechnol. 2011, 6, 668–674. [Google Scholar] [CrossRef]
- Takeuchi, N.; Hiratani, M.; Kawano, R. Pattern Recognition of MicroRNA Expression in Body Fluids Using Nanopore Decoding at Subfemtomolar Concentrations. JACS Au 2022, 2, 1829–1838. [Google Scholar] [CrossRef]
- Greene, J.; Baird, A.-M.; Brady, L.; Lim, M.; Gray, S.G.; McDermott, R.; Finn, S.P. Circular RNAs: Biogenesis, Function and Role in Human Diseases. Front. Mol. Biosci. 2017, 4, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Hou, L.; Zuo, Z.; Ji, P.; Zhang, X.; Xue, Y.; Zhao, F. Comprehensive Profiling of Circular RNAs with Nanopore Sequencing and CIRI-Long. Nat. Biotechnol. 2021, 39, 836–845. [Google Scholar] [CrossRef] [PubMed]
- Lebrigand, K.; Magnone, V.; Barbry, P.; Waldmann, R. High Throughput Error Corrected Nanopore Single Cell Transcriptome Sequencing. Nat. Commun. 2020, 11, 4025. [Google Scholar] [CrossRef] [PubMed]
- Philpott, M.; Watson, J.; Thakurta, A.; Brown, T.; Brown, T.; Oppermann, U.; Cribbs, A.P. Nanopore Sequencing of Single-Cell Transcriptomes with ScCOLOR-Seq. Nat. Biotechnol. 2021, 39, 1517–1520. [Google Scholar] [CrossRef] [PubMed]
- 1D Squared Kit Available in the Store: Boost Accuracy, Simple Prep. Available online: https://nanoporetech.com/about-us/news/1d-squared-kit-available-store-boost-accuracy-simple-prep (accessed on 14 January 2023).
- Teng, H.; Cao, M.D.; Hall, M.B.; Duarte, T.; Wang, S.; Coin, L.J.M. Chiron: Translating Nanopore Raw Signal Directly into Nucleotide Sequence Using Deep Learning. GigaScience 2018, 7, giy037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wick, R.R.; Judd, L.M.; Holt, K.E. Performance of Neural Network Basecalling Tools for Oxford Nanopore Sequencing. Genome Biol. 2019, 20, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, Y.K.; Hendra, C.; Pratanwanich, P.N.; Göke, J. Beyond Sequencing: Machine Learning Algorithms Extract Biology Hidden in Nanopore Signal Data. Trends Genet. 2022, 38, 246–257. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MacKenzie, M.; Argyropoulos, C. An Introduction to Nanopore Sequencing: Past, Present, and Future Considerations. Micromachines 2023, 14, 459. https://doi.org/10.3390/mi14020459
MacKenzie M, Argyropoulos C. An Introduction to Nanopore Sequencing: Past, Present, and Future Considerations. Micromachines. 2023; 14(2):459. https://doi.org/10.3390/mi14020459
Chicago/Turabian StyleMacKenzie, Morgan, and Christos Argyropoulos. 2023. "An Introduction to Nanopore Sequencing: Past, Present, and Future Considerations" Micromachines 14, no. 2: 459. https://doi.org/10.3390/mi14020459