Cyanotoxins and Other Bioactive Compounds from the Pasteur Cultures of Cyanobacteria (PCC)


Abstract
:1. Introduction
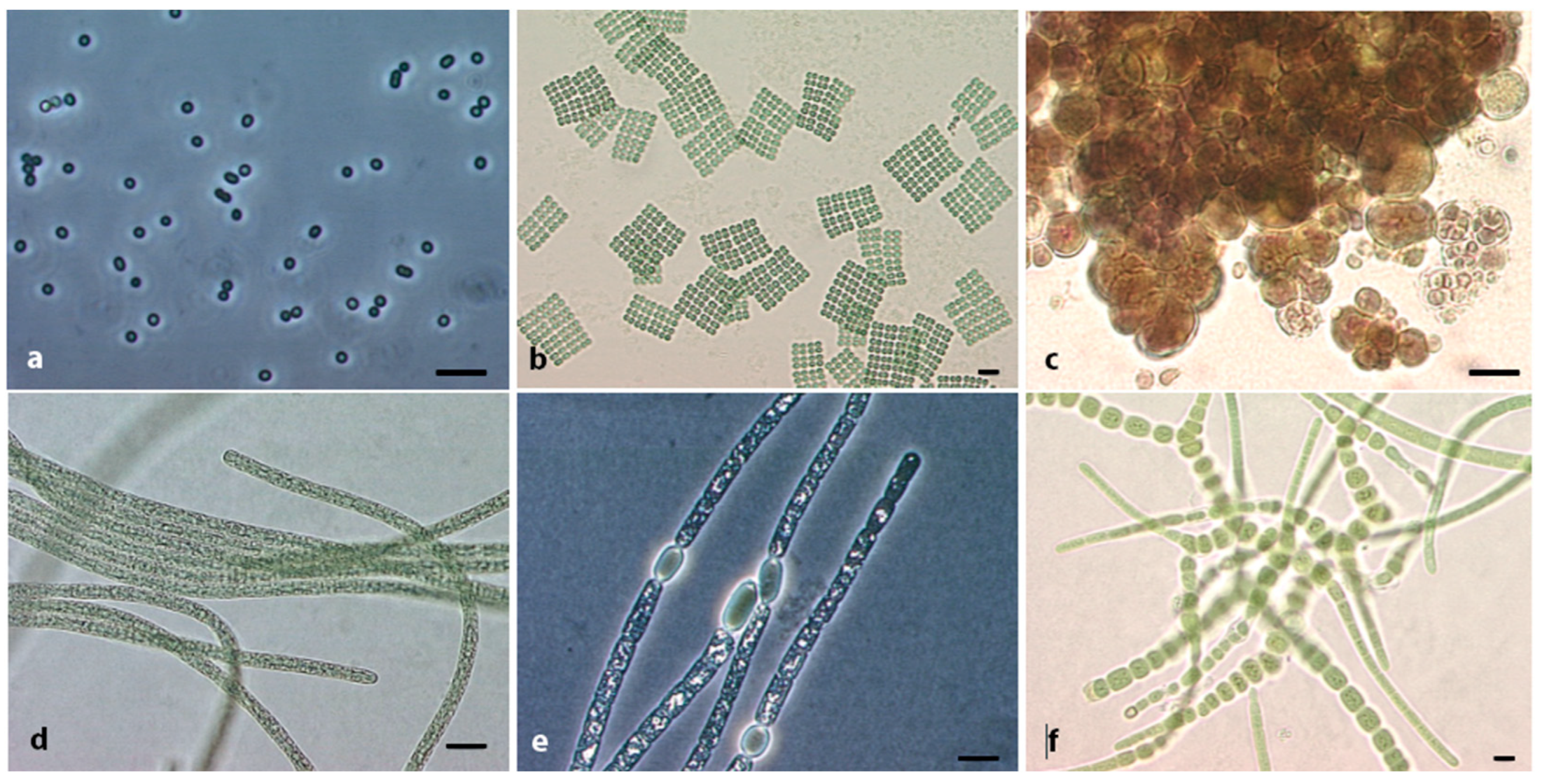
2. The Phylum Cyanobacteria, Diversity in Terms of Morphology and Genome
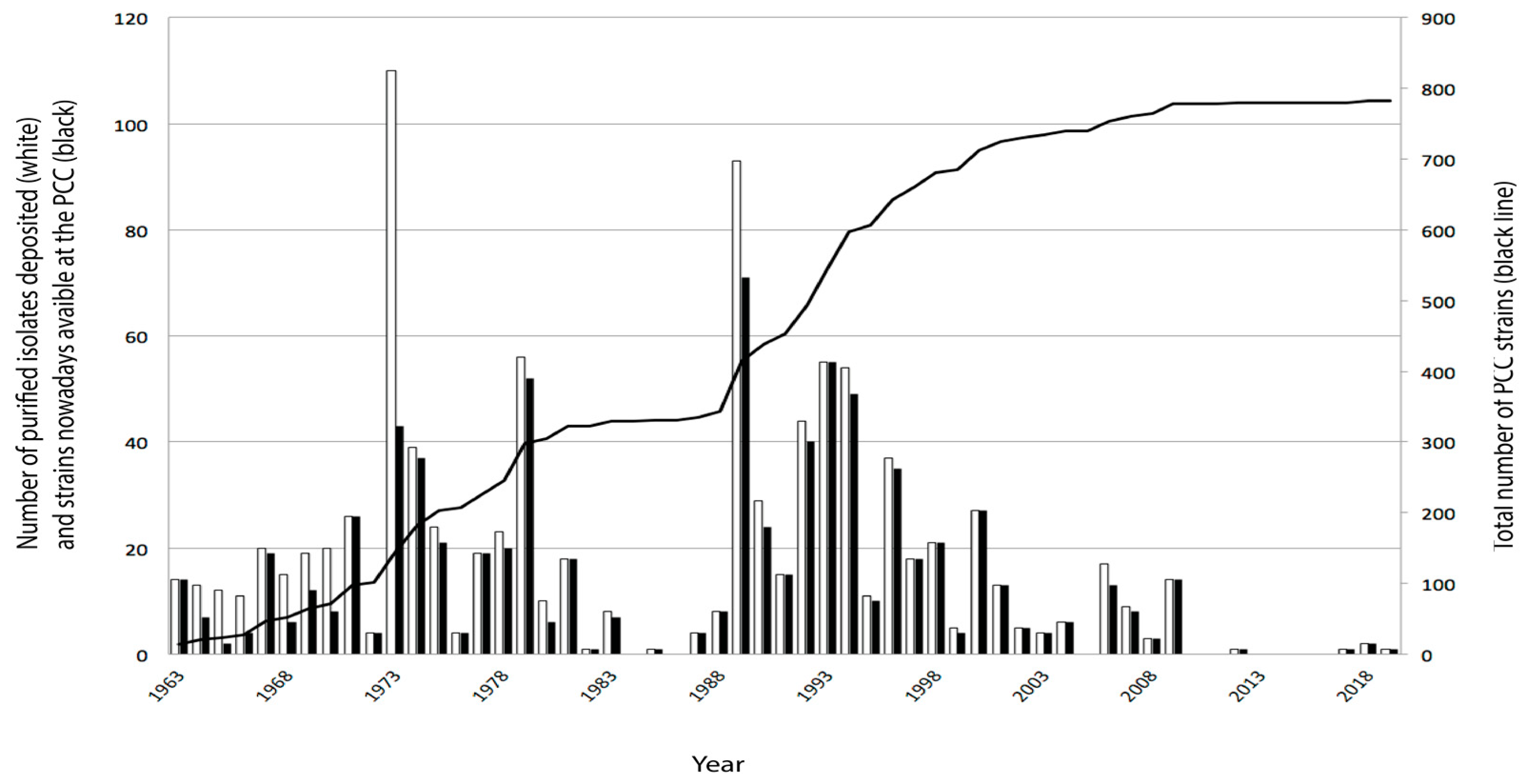
3. Collection of Cyanobacteria and the Pasteur Cultures of Cyanobacteria (PCC)
4. Natural Products of Cyanobacteria—From Toxins to Novel Compounds
5. From Molecules to Bioactivity

{kind=link}
{kind=link}
| NPs or Their BGCs | Type of NPs | Gene Cluster | Strain | Reference |
|---|---|---|---|---|
| Microcyclamide | RiPPs, cyanobactins | mca, 13 kb | Microcystis aeruginosa PCC 7806 | [26] |
| Viridisamide | RiPPs, 1st linear cyanobactins | Variation from pat * | Oscillatoria viridis PCC 7112 | [69] |
| Aeruginosamide B and C3 | Microcystis aeruginosa PCC 9432 | |||
| Muscoride | RiPPs, linear cyanobactin | mus, 12.7 kb | 12 Nostoc strains, in which 6 PCC strains | [61] |
| Tolypamide | RiPPs, cyanobactins | tol, 10.4 kb | Tolypothrix sp. PCC 7601 | [62] |
| Geosmin | RiPPs, sesquiterpene | geosmin synthase | Nostoc spp. PCC 7310 and PCC 7120, Kamptonema sp. PCC 6506 | [70,71] |
| Merosterol A and B + isomer | RiPPs, meroterpene | mst, 29 kb | Scytonema sp. PCC 10023 | [72] |
| Cyclophanes | RiPPs, cyclopetide alkaloids | lsc, 2.6 kb | Lyngbya sp. PCC 8106 and various other strains | [63] |
| Landornamides | RiPPs, proteusins | osp, 12 kb | Kamptonema sp. PCC 6506 and 6 other Kamptonema PCC strains | [73] |
| Kamptornamide | RiPPs, 1st ribosomal fatty-acylated lipo- petides, selidamides | ksp, 6.3 kb | Kamptonema sp. PCC 6506 and Nostoc punctiforme PCC 73102 | [74] |
| Microguanidine amide Aeruginoguanidine BGC | NRPS | agd, 34 kb | 11 Microcystis, in which 7 PCC strains | [75] |
| Hassallidin E | NRPS | has, 48 kb | Planktothrix serta PCC 8927 | [76] |
| Cyanopeptolin | NRPS | oci *, 31.5 kb | Microcystis aeruginosa PCC 7806 Scytonema hofmanni PCC 7110 | [77,78] |
| Scyptolins | NRPS | Scytonema hofmanni PCC 7110 | [79] | |
| Anatoxin-a and dihydroanatoxin | PKS, alkaloid | ana, 20 kb | Kamptonema sp. PCC 6506, Cylindrospermum sp. PCC 7417 and 13 other PCC strains | [36,55] |
| Cylindrospermopsins | PKS, alkaloid | cyr, 42 kb | Kamptonema sp. PCC 6506 | [56] |
| Luminaolide B | Trans AT-PKS | lum, 99 kb | Planktothrix paucivesiculata PCC 9631 | [80,81] |
| Tolytoxin BGC | tto, >100 kb | Scytonema sp. PCC 10023 | ||
| Tolytoxin, Scytophycin | tto, 92.8 kb | Planktothrix sp. PCC 11201 | ||
| Leptolyngbyalide | Trans AT-PKS | lept, 96.7 kb | Leptolyngbya sp. PCC 7375 | [82] |
| Alkene and alkanes | PKS, Hydrocarbon | ols 10 kb | Synechococcus sp. PCC 7002 16 unicellular PCC strains | [83,84,85] |
| Heterocyte glycolipids | PKS, polyunsaturated fatty acid | CF1 * | Nostoc sp. PCC 7120, 18 Nostocales strains and Microchaete sp. PCC 7126 | [86,87,88] |
| Microcystin | NRPS-PKS Hybrid | mcy, 55 kb | Microcystis aeruginosa PCC 7806, Fischerella sp. PCC 9339 | [36,54] |
| Nostopeptolide | NRPS-PKS Hybrid | pks2, 62.7 kb | Nostoc puctiforme ATCC29133/PCC 73102 | [89,90] |
| Aranazoles | NRPS-PKS Hybrid | arz, 43 kb | Fischerella sp. PCC 9339 | [91] |
6. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Garcia-Pichel, F.; Zehr, J.P.; Bhattacharya, D.; Pakrasi, H.B. What’s in a name? The case of cyanobacteria. J. Phycol. 2020, 56, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schirrmeister, B.E.; Gugger, M.; Donoghue, P.C. Cyanobacteria and the Great Oxidation Event: Evidence from genes and fossils. Palaeontology 2015, 58, 769–785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shih, P.M.; Wu, D.; Latifi, A.; Axen, S.D.; Fewer, D.P.; Talla, E.; Calteau, A.; Cai, F.; Tandeau de Marsac, N.; Rippka, R.; et al. Improving the coverage of the cyanobacterial phylum using diversity-driven genome sequencing. Proc. Natl. Acad. Sci. USA 2013, 110, 1053–1058. [Google Scholar] [CrossRef] [Green Version]
- Huisman, J.; Codd, G.A.; Paerl, H.W.; Ibelings, B.W.; Verspagen, J.M.H.; Visser, P.M. Cyanobacterial blooms. Nat. Rev. Microbiol. 2018, 16, 471–483. [Google Scholar] [CrossRef]
- Dittmann, E.; Gugger, M.; Sivonen, K.; Fewer, D.P. Natural product biosynthetic diversity and comparative genomics of the Cyanobacteria. Trends Microbiol. 2015, 23, 642–652. [Google Scholar] [CrossRef]
- Partensky, F.; Hess, W.R.; Vaulot, D. Prochlorococcus, a marine photosynthetic prokaryote of global significance. Microbiol. Mol. Biol. Rev. 1999, 63, 106–127. [Google Scholar] [CrossRef] [Green Version]
- Humbert, J.F.; Barbe, V.; Latifi, A.; Gugger, M.; Calteau, A.; Coursin, T.; Lajus, A.; Castelli, V.; Oztas, S.; Samson, G.; et al. A tribute to disorder in the genome of the bloom-forming freshwater cyanobacterium Microcystis aeruginosa. PLoS ONE 2013, 8, e70747. [Google Scholar] [CrossRef] [PubMed]
- Komarek, J.; Anagnostidis, K. Cyanoprokaryota I (Chroococales); Pascher, A., Ettl, H., Gärtner, G., Heyning, H., Mollenhauer, D., Eds.; Gustav Fischer: Jena, Germany, 1998; Volume 19/1. [Google Scholar]
- Komarek, J.; Anagnostidis, K. Cyanoprokaryota II (Oscillatoriales); Büdel, B., Gärtner, G., Krietniz, L., Schagerl, M., Eds.; Spektrum Akademischer Verlag: Jena, Germany, 2005; Volume 19/2. [Google Scholar]
- Komarek, J. Cyanoprokaryota III (Heterocytous Genera); Springer Spektrum: Berlin/Heidelberg, Germany, 2013; Volume 19/3. [Google Scholar]
- Castenholz, R.W.; Phylum, B.X. Cyanobacteria, Oxygenic Photosynthetic Bacteria. In Bergey’s Manual of Systematic Bacteriology, 2nd ed.; Boone, D.R., Castenholz, R.W., Garrity, G.M., Eds.; Volume 1—The Archaea and the Deeply Branching and Phototrophic Bacteria; Springer: New York, NY, USA, 2001; pp. 473–487. [Google Scholar]
- Gonzalez-Esquer, C.R.; Smarda, J.; Rippka, R.; Axen, S.D.; Guglielmi, G.; Gugger, M.; Kerfeld, C.A. Cyanobacterial ultrastructure in light of genomic sequence data. Photosynth. Res. 2016, 129, 147–157. [Google Scholar] [CrossRef]
- Stanier, R.Y.; Sistrom, W.R.; Hansen, T.A.; Whitton, B.A.; Castenholz, R.W.; Pfennig, N.; Gorlenko, V.N.; Kondratieva, E.N.; Eimhjellen, K.E.; Whittenbury, R.; et al. Proposal to place the nomemclature of the cyanobacteria (blue-green algae) under the rules of the International Code of Nomenclature of Bacteria. Int. J. Syst. Bacteriol. 1978, 28, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Rippka, R.; Waterbury, J.; Cohen-Bazire, G. A cyanobacterium which lacks thylakoids. Arch. Microbiol. 1974, 100, 419–436. [Google Scholar] [CrossRef]
- Guglielmi, G.; Cohen-Bazire, G. Structure et distribution des pores et des perforations de l’enveloppe de peptidoglycane chez quelques cyanobactéries. Protistologica 1982, 18, 151–165. [Google Scholar]
- Guglielmi, G.; Cohen-Bazire, G. Etude taxonomique d’un genre de cyanobactérie oscillatoriacée: Le genre Pseudanabaena Lauterborn. I. Etude ultrastructurale. Protistologica 1984, 20, 377–391. [Google Scholar]
- Campbell, D.; Houmard, J.; Tandeau de Marsac, N. Electron transport regulates cellular differentiation in the filamentous cyanobacterium Calothrix. Plant Cell 1993, 5, 451–463. [Google Scholar] [CrossRef] [PubMed]
- Damerval, T.; Castets, A.M.; Guglielmi, G.; Houmard, J.; Tandeau de Marsac, N. Occurrence and distribution of gas vesicle genes among cyanobacteria. J. Bacteriol. 1989, 171, 1445–1452. [Google Scholar] [CrossRef] [Green Version]
- Forchhammer, K.; Tandeau de Marsac, N. Phosphorylation of the PII protein (glnB gene product) in the cyanobacterium Synechococcus sp. strain PCC 7942: Analysis of in vitro kinase activity. J. Bacteriol. 1995, 177, 5812–5817. [Google Scholar] [CrossRef] [Green Version]
- Herdman, M.; Coursin, T.; Rippka, R.; Houmard, J.; Tandeau de Marsac, N. A new appraisal of the prokaryotic origin of eukaryotic phytochromes. J. Mol. Evol. 2000, 51, 205–213. [Google Scholar] [CrossRef]
- Tandeau de Marsac, N.; Mazel, D.; Damerval, T.; Guglielmi, G.; Capuano, V.; Houmard, J. Photoregulation of gene expression in the filamentous cyanobacterium Calothrix sp. PCC 7601: Light-harvesting complexes and cell differentiation. Photosynth. Res. 1988, 18, 99–132. [Google Scholar] [CrossRef]
- Mazel, D.; Houmard, J.; Tandeau de Marsac, N. A multigene family in Calothrix sp. PCC 7601 encodes phycocyanin, the major component of the cyanobacterial light-harvesting antenna. Mol. Gen. Genet. 1988, 211, 296–304. [Google Scholar] [CrossRef]
- Damerval, T.; Guglielmi, G.; Houmard, J.; Tandeau de Marsac, N. Hormogonium differentiation in the cyanobacterium Calothrix: A photoregulated developmental process. Plant Cell 1991, 3, 191–201. [Google Scholar] [CrossRef]
- Liotenberg, S.; Campbell, D.; Rippka, R.; Houmard, J.; Tandeau de Marsac, N. Effect of the nitrogen source on phycobiliprotein synthesis and cell reserves in a chromatically adapting filamentous cyanobacterium. Microbiology 1996, 142, 611–622. [Google Scholar] [CrossRef] [Green Version]
- Frangeul, L.; Quillardet, P.; Castets, A.M.; Humbert, J.F.; Matthijs, H.C.; Cortez, D.; Tolonen, A.; Zhang, C.C.; Gribaldo, S.; Kehr, J.C.; et al. Highly plastic genome of Microcystis aeruginosa PCC 7806, a ubiquitous toxic freshwater cyanobacterium. BMC Genom. 2008, 9, 274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziemert, N.; Ishida, K.; Quillardet, P.; Bouchier, C.; Hertweck, C.; Tandeau de Marsac, N.; Dittmann, E. Microcyclamide biosynthesis in two strains of Microcystis aeruginosa: From structure to genes and vice versa. Appl. Environ. Microbiol. 2008, 74, 1791–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zilliges, Y.; Kehr, J.C.; Mikkat, S.; Bouchier, C.; Tandeau de Marsac, N.; Börner, T.; Dittmann, E. An extracellular glycoprotein is implicated in cell-cell contacts in the toxic cyanobacterium Microcystis aeruginosa PCC 7806. J. Bacteriol. 2008, 190, 2871–2879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rippka, R.; Deruelles, J.; Waterbury, J.B.; Herdman, M.; Stanier, R.Y. Generic assignments, strain histories and properties of pure cultures of cyanobacteria. J. Gen. Microbiol. 1979, 111, 1–61. [Google Scholar] [CrossRef] [Green Version]
- Rippka, R.; Herdman, M. Pasteur Culture Collection of Cyanobacterial Strains in Axenic Culture. In Catalogue & Taxonomic Handbook; Institut Pasteur: Paris, France, 1992; pp. 1–103. [Google Scholar]
- Palinska, K.A.; Jahns, T.; Rippka, R.; Tandeau de Marsac, N. Prochlorococcus marinus strain PCC 9511, a picoplanktonic cyanobacterium, synthesizes the smallest urease. Microbiology 2000, 146, 3099–3107. [Google Scholar] [CrossRef] [Green Version]
- Adomako, M.; Ernst, D.; Simkovsky, R.; Chao, Y.Y.; Wang, J.; Fang, M.; Bouchier, C.; Lopez-Igual, R.; Mazel, D.; Gugger, M.; et al. Comparative genomics of Synechococcus elongatus explains the phenotypic diversity of the strains. mBio 2022, 13, e0086222. [Google Scholar] [CrossRef]
- Rippka, R.; Coursin, T.; Hess, W.; Lichtlé, C.; Scanlan, D.J.; Palinska, K.A.; Iteman, I.; Partensky, F.; Houmard, J.; Herdman, M. Prochlorococcus marinus Chisholm et al. 1992, subsp. nov. pastoris, strain PCC 9511, the first axenic chlorophyll a2/b2-containing cyanobacterium (Oxyphotobacteria). Int. J. Syst. Evol. Microbiol 2000, 50, 1833–1847. [Google Scholar] [CrossRef] [Green Version]
- Włodarczyk, A.; Selão, T.T.; Norling, B.; Nixon, P.J. Newly discovered Synechococcus sp. PCC 11901 is a robust cyanobacterial strain for high biomass production. Commun. Biol. 2020, 3, 215. [Google Scholar] [CrossRef]
- Jaiswal, D.; Sengupta, A.; Sohoni, S.; Sengupta, S.; Phadnavis, A.G.; Pakrasi, H.B.; Wangikar, P.P. Genome features and biochemical characteristics of a robust, fast growing and naturally transformable cyanobacterium Synechococcus elongatus PCC 11801 isolated from India. Sci. Rep. 2018, 8, 16632. [Google Scholar] [CrossRef]
- Jaiswal, D.; Sengupta, A.; Sengupta, S.; Madhu, S.; Pakrasi, H.B.; Wangikar, P.P. A novel cyanobacterium Synechococcus elongatus PCC 11802 has distinct genomic and metabolomic characteristics compared to its neighbor PCC 11801. Sci. Rep. 2020, 10, 191. [Google Scholar] [CrossRef] [Green Version]
- Calteau, A.; Fewer, D.P.; Latifi, A.; Coursin, T.; Laurent, T.; Jokela, J.; Kerfeld, C.A.; Sivonen, K.; Piel, J.; Gugger, M. Phylum-wide comparative genomics unravel the diversity of secondary metabolism in Cyanobacteria. BMC Genom. 2014, 15, 977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francis, G. Poisonous Australian Lake. Nature 1878, 18, 11–12. [Google Scholar] [CrossRef] [Green Version]
- Edwards, C.; Beattie, K.A.; Scrimgeour, C.M.; Codd, G.A. Identification of anatoxin-A in benthic cyanobacteria (blue-green algae) and in associated dog poisonings at Loch Insh, Scotland. Toxicon 1992, 30, 1165–1175. [Google Scholar] [CrossRef] [PubMed]
- Gugger, M.; Lenoir, S.; Berger, C.; Ledreux, A.; Druart, J.C.; Humbert, J.F.; Guette, C.; Bernard, C. First report in a river in France of the benthic cyanobacterium Phormidium favosum producing anatoxin-a associated with dog neurotoxicosis. Toxicon 2005, 45, 919–928. [Google Scholar] [CrossRef]
- Harding, W.R.; Rowe, N.; Wessels, J.C.; Beattie, K.A.; Codd, G.A. Death of a dog attributed to the cyanobacterial (blue-green algal) hepatotoxin nodularin in South Africa. J. S. Afr. Vet. Assoc. 1995, 66, 256–259. [Google Scholar]
- Kelly, L.T.; Bouma-Gregson, K.; Puddick, J.; Fadness, R.; Ryan, K.G.; Davis, T.W.; Wood, S.A. Multiple cyanotoxin congeners produced by sub-dominant cyanobacterial taxa in riverine cyanobacterial and algal mats. PLoS ONE 2019, 14, e0220422. [Google Scholar] [CrossRef] [Green Version]
- Mahmood, N.A.; Carmichael, W.W.; Pfahler, D. Anticholinesterase poisonings in dogs from a cyanobacterial (blue-green algae) bloom dominated by Anabaena flos-aquae. Am. J. Vet. Res. 1988, 49, 500–503. [Google Scholar]
- Turner, A.D.; Turner, F.R.I.; White, M.; Hartnell, D.; Crompton, C.G.; Bates, N.; Egginton, J.; Branscombe, L.; Lewis, A.M.; Maskrey, B.H. Confirmation Using Triple Quadrupole and High-Resolution Mass Spectrometry of a Fatal Canine Neurotoxicosis following Exposure to Anatoxins at an Inland Reservoir. Toxins 2022, 14, 804. [Google Scholar] [CrossRef]
- Wood, S.A.; Heath, M.W.; Holland, P.T.; Munday, R.; McGregor, G.B.; Ryan, K.G. Identification of a benthic microcystin-producing filamentous cyanobacterium (Oscillatoriales) associated with a dog poisoning in New Zealand. Toxicon 2010, 55, 897–903. [Google Scholar] [CrossRef]
- Azevedo, S.M.; Carmichael, W.W.; Jochimsen, E.M.; Rinehart, K.L.; Lau, S.; Shaw, G.R.; Eaglesham, G.K. Human intoxication by microcystins during renal dialysis treatment in Caruaru-Brazil. Toxicology 2002, 181–182, 441–446. [Google Scholar] [CrossRef]
- Breinlinger, S.; Phillips, T.J.; Haram, B.N.; Mareš, J.; Martínez Yerena, J.A.; Hrouzek, P.; Sobotka, R.; Henderson, W.M.; Schmieder, P.; Williams, S.M.; et al. Hunting the eagle killer: A cyanobacterial neurotoxin causes vacuolar myelinopathy. Science 2021, 371, eaax9050. [Google Scholar] [CrossRef] [PubMed]
- Krienitz, L.; Ballot, A.; Kotut, K.; Wiegand, C.; Pütz, S.; Metcalf, J.S.; Codd, G.A.; Pflugmacher, S. Contribution of hot spring cyanobacteria to the mysterious deaths of Lesser Flamingos at Lake Bogoria, Kenya. FEMS Microbiol. Ecol. 2003, 43, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Skafi, M.; Duy, S.V.; Munoz, G.; Dinh, Q.T.; Simon, D.F.; Juneau, P.; Sauvé, S. Occurrence of microcystins, anabaenopeptins and other cyanotoxins in sh from a freshwater wildlife reserve impacted by harmful cyanobacterial blooms. Toxicon 2021, 194, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Veerman, J.; Kumar, A.; Mishra, D.R. Exceptional landscape-wide cyanobacteria bloom in Okavango Delta, Botswana in 2020 coincided with a mass elephant die-off event. Harmful Algae 2022, 111, 102145. [Google Scholar] [CrossRef]
- Chorus, I.; Welker, M. Toxic Cyanobacteria in Water: A Guide to Their Pubic Health Consequences, Monitoring and Management, 2nd ed.; Welker, I.C.M., Ed.; CRC Press: Boca Raton, FL, USA; on behalf of the World Health Organization: Geneva, Switzerland, 2021. [Google Scholar]
- Trevino-Garrison, I.; DeMent, J.; Ahmed, F.S.; Haines-Lieber, P.; Langer, T.; Ménager, H.; Neff, J.; van der Merwe, D.; Carney, E. Human illnesses and animal deaths associated with freshwater harmful algal blooms-Kansas. Toxins 2015, 7, 353–366. [Google Scholar] [CrossRef] [Green Version]
- Hilborn, E.D.; Beasley, V.R. One health and cyanobacteria in freshwater systems: Animal illnesses and deaths are sentinel events for human health risks. Toxins 2015, 7, 1374–1395. [Google Scholar] [CrossRef] [Green Version]
- Theiss, W.C.; Carmichael, W.W.; Wyman, J.; Bruner, R. Blood pressure and hepatocellular effects of the cyclic heptapeptide toxin produced by the freshwater cyanobacterium (blue-green alga) Microcystis aeruginosa strain PCC-7820. Toxicon 1988, 26, 603–613. [Google Scholar] [CrossRef]
- Tillett, D.; Dittmann, E.; Erhard, M.; von Döhren, H.; Börner, T.; Neilan, B.A. Structural organization of microcystin biosynthesis in Microcystis aeruginosa PCC7806: An integrated peptide-polyketide synthetase system. Chem. Biol. 2000, 7, 753–764. [Google Scholar] [CrossRef] [Green Version]
- Méjean, A.; Mann, S.; Maldiney, T.; Vassiliadis, G.; Lequin, O.; Ploux, O. Evidence that biosynthesis of the neurotoxic alkaloids anatoxin-a and homoanatoxin-a in the cyanobacterium Oscillatoria PCC 6506 occurs on a modular polyketide synthase initiated by L-proline. J. Am. Chem. Soc. 2009, 131, 7512–7513. [Google Scholar] [CrossRef]
- Mazmouz, R.; Chapuis-Hugon, F.; Mann, S.; Pichon, V.; Méjean, A.; Ploux, O. Biosynthesis of cylindrospermopsin and 7-epicylindrospermopsin in Oscillatoria sp. strain PCC 6506: Identification of the cyr gene cluster and toxin analysis. Appl. Environ. Microbiol. 2010, 76, 4943–4949. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Pearson, L.A.; Mazmouz, R.; Liu, T.; Neilan, B.A. Heterologous expression and biochemical analysis reveal a schizokinen-based siderophore pathway in Leptolyngbya (Cyanobacteria). Appl. Environ. Microbiol. 2022, 88, e0237321. [Google Scholar] [CrossRef] [PubMed]
- Morinaka, B.I.; Verest, M.; Freeman, M.F.; Gugger, M.; Piel, J. An orthogonal D2 O-based induction system that provides insights into d-amino acid pattern formation by radical S-adenosylmethionine peptide epimerases. Angew. Chem. Int. Ed. 2017, 56, 762–766. [Google Scholar] [CrossRef]
- Morinaka, B.I.; Vagstad, A.L.; Helf, M.J.; Gugger, M.; Kegler, C.; Freeman, M.F.; Bode, H.B.; Piel, J. Radical S-adenosyl methionine epimerases: Regioselective introduction of diverse D-amino acid patterns into peptide natural products. Angew. Chem. Int. Ed. 2014, 53, 8503–8507. [Google Scholar] [CrossRef] [PubMed]
- Morinaka, B.I.; Lakis, E.; Verest, M.; Helf, M.J.; Scalvenzi, T.; Vagstad, A.L.; Sims, J.; Sunagawa, S.; Gugger, M.; Piel, J. Natural noncanonical protein splicing yields products with diverse β-amino acid residues. Science 2018, 359, 779–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattila, A.; Andsten, R.M.; Jumppanen, M.; Assante, M.; Jokela, J.; Wahlsten, M.; Mikula, K.M.; Sigindere, C.; Kwak, D.H.; Gugger, M.; et al. Biosynthesis of the bis-prenylated alkaloids muscoride A and B. ACS Chem. Biol. 2019, 14, 2683–2690. [Google Scholar] [CrossRef] [PubMed]
- Purushothaman, M.; Sarkar, S.; Morita, M.; Gugger, M.; Schmidt, E.W.; Morinaka, B.I. Genome-mining-based discovery of the cyclic peptide tolypamide and tolF, a Ser/Thr forward O-prenyltransferase. Angew. Chem. Int. Ed. 2021, 60, 8460–8465. [Google Scholar] [CrossRef]
- Nguyen, T.Q.N.; Tooh, Y.W.; Sugiyama, R.; Nguyen, T.P.D.; Purushothaman, M.; Leow, L.C.; Hanif, K.; Yong, R.H.S.; Agatha, I.; Winnerdy, F.R.; et al. Post-translational formation of strained cyclophanes in bacteria. Nat. Chem. 2020, 12, 1042–1053. [Google Scholar] [CrossRef]
- Chlipala, G.E.; Mo, S.; Orjala, J. Chemodiversity in freshwater and terrestrial cyanobacteria—A source for drug discovery. Curr. Drug Targets 2011, 12, 1654–1673. [Google Scholar] [CrossRef]
- Gerwick, W.H.; Fenner, A.M. Drug discovery from marine microbes. Microb. Ecol. 2013, 65, 800–806. [Google Scholar]
- Vijayakumar, S.; Menakha, M. Pharmaceutical applications of cyanobacteria-A review. J. Acute Med. 2015, 5, 15–23. [Google Scholar] [CrossRef] [Green Version]
- Ali Shah, S.A.; Akhter, N.; Auckloo, B.N.; Khan, I.; Lu, Y.; Wang, K.; Wu, B.; Guo, Y.W. Structural diversity, biological properties and applications of natural products from cyanobacteria. A review. Mar. Drugs 2017, 15, 354. [Google Scholar] [CrossRef] [Green Version]
- Demay, J.; Bernard, C.; Reinhardt, A.; Marie, B. Natural products from Cyanobacteria: Focus on beneficial activities. Mar. Drugs 2019, 17, 320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leikoski, N.; Liu, L.; Jokela, J.; Wahlsten, M.; Gugger, M.; Calteau, A.; Permi, P.; Kerfeld, C.A.; Sivonen, K.; Fewer, D.P. Genome mining expands the chemical diversity of the cyanobactin family to include highly modified linear peptides. Chem. Biol. 2013, 20, 1033–1043. [Google Scholar] [CrossRef] [PubMed]
- Melo, N.; Wolff, G.H.; Costa-da-Silva, A.L.; Arribas, R.; Triana, M.F.; Gugger, M.; Riffell, J.A.; DeGennaro, M.; Stensmyr, M.C. Geosmin attracts Aedes aegypti mosquitoes to oviposition sites. Curr. Biol. 2020, 30, 127–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agger, S.A.; Lopez-Gallego, F.; Hoye, T.R.; Schmidt-Dannert, C. Identification of sesquiterpene synthases from Nostoc punctiforme PCC 73102 and Nostoc sp. strain PCC 7120. J. Bacteriol. 2008, 190, 6084–6096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moosmann, P.; Ueoka, R.; Grauso, L.; Mangoni, A.; Morinaka, B.I.; Gugger, M.; Piel, J. Cyanobacterial ent-sterol-like natural products from a deviated ubiquinone pathway. Angew. Chem. Int. Ed. 2017, 56, 4987–4990. [Google Scholar] [CrossRef] [PubMed]
- Bösch, N.M.; Borsa, M.; Greczmiel, U.; Morinaka, B.I.; Gugger, M.; Oxenius, A.; Vagstad, A.L.; Piel, J. Landornamides: Antiviral ornithine-containing ribosomal peptides discovered through genome mining. Angew. Chem. Int. Ed. 2020, 59, 11763–11768. [Google Scholar] [CrossRef]
- Hubrich, F.; Bösch, N.M.; Chepkirui, C.; Morinaka, B.I.; Rust, M.; Gugger, M.; Robinson, S.L.; Vagstad, A.L.; Piel, J. Ribosomally derived lipopeptides containing distinct fatty acyl moieties. Proc. Natl. Acad. Sci. USA 2022, 119, e2113120119. [Google Scholar] [CrossRef]
- Pancrace, C.; Ishida, K.; Briand, E.; Pichi, D.G.; Weiz, A.R.; Guljamow, A.; Scalvenzi, T.; Sassoon, N.; Hertweck, C.; Dittmann, E.; et al. Unique biosynthetic pathway in bloom-forming cyanobacterial genus Microcystis jointly assembles cytotoxic aeruginoguanidines and microguanidines. ACS Chem. Biol. 2019, 14, 67–75. [Google Scholar] [CrossRef] [Green Version]
- Pancrace, C.; Jokela, J.; Sassoon, N.; Ganneau, C.; Desnos-Ollivier, M.; Wahlsten, M.; Humisto, A.; Calteau, A.; Bay, S.; Fewer, D.P.; et al. Rearranged biosynthetic gene cluster and synthesis of hassallidin E in Planktothrix serta PCC 8927. ACS Chem. Biol. 2017, 12, 1796–1804. [Google Scholar] [CrossRef] [Green Version]
- Martin, C.; Oberer, L.; Buschdtt, M.; Weckesser, J. Cyanopeptolins, new depsipeptides from the cyanobacterium Microcystis sp. PCC 7806. J. Antibiot. 1993, 46, 1550–1556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matern, U.; Oberer, L.; Erhard, M.; Herdmand, M.; Weckesser, J. Hofmannolin, a cyanopeptolin from Scytonema hofmanni PCC 7110. Phytochemistry 2003, 64, 1061–1067. [Google Scholar] [CrossRef] [PubMed]
- Matern, U.; Oberer, L.; Falchetto, R.A.; Erhard, M.; König, W.A.; Herdman, M.; Weckesser, J. Scyptolin A and B, cyclic depsipeptides from axenic cultures of Scytonema hofmanni PCC 7110. Phytochemistry 2001, 58, 1087–1095. [Google Scholar] [CrossRef] [PubMed]
- Ueoka, R.; Uria, A.R.; Reiter, S.; Mori, T.; Karbaum, P.; Peters, E.E.; Helfrich, E.J.; Morinaka, B.I.; Gugger, M.; Takeyama, H.; et al. Metabolic and evolutionary origin of actin-binding polyketides from diverse organisms. Nat. Chem. Biol. 2015, 11, 705–712. [Google Scholar] [CrossRef]
- Pancrace, C.; Barny, M.A.; Ueoka, R.; Calteau, A.; Scalvenzi, T.; Pédron, J.; Barbe, V.; Piel, J.; Humbert, J.F.; Gugger, M. Insights into the Planktothrix genus: Genomic and metabolic comparison of benthic and planktic strains. Sci. Rep. 2017, 7, 41181. [Google Scholar] [CrossRef] [Green Version]
- Helfrich, E.J.N.; Ueoka, R.; Dolev, A.; Rust, M.; Meoded, R.A.; Bhushan, A.; Califano, G.; Costa, R.; Gugger, M.; Steinbeck, C.; et al. Automated structure prediction of trans-acyltransferase polyketide synthase products. Nat. Chem. Biol. 2019, 15, 813–821. [Google Scholar] [CrossRef]
- Brito, Ã.; Vieira, J.; Vieira, C.P.; Zhu, T.; Leao, P.N.; Ramos, V.; Lu, X.; Vasconcelos, V.M.; Gugger, M.; Tamagnini, P. Comparative genomics discloses the uniqueness and the biosynthetic potential of the marine cyanobacterium Hyella patelloides. Front. Microbiol. 2020, 11, 1527. [Google Scholar] [CrossRef]
- Coates, R.C.; Podell, S.; Korobeynikov, A.; Lapidus, A.; Pevzner, P.; Sherman, D.H.; Allen, E.E.; Gerwick, L.; Gerwick, W.H. Characterization of cyanobacterial hydrocarbon composition and distribution of biosynthetic pathways. PLoS ONE 2014, 9, e85140. [Google Scholar] [CrossRef] [Green Version]
- Mendez-Perez, D.; Begemann, M.B.; Pfleger, B.F. Modular synthase-encoding gene involved in α-olefin biosynthesis in Synechococcus sp. strain PCC 7002. Appl. Environ. Microbiol. 2011, 77, 4264–4267. [Google Scholar] [CrossRef] [Green Version]
- Bauersachs, T.; Gugger, M.; Schwark, L. Heterocyte glycolipid diketones: A novel type of biomarker in the N2-fixing heterocytous cyanobacterium Microchaete sp. Org. Geochem. 2020, 141, 103976. [Google Scholar] [CrossRef]
- Bauersachs, T.; Miller, S.R.; Gugger, M.; Mudimu, O.; Friedl, T.; Schwark, L. Heterocyte glycolipids indicate polyphyly of stigonematalean cyanobacteria. Phytochemistry 2019, 166, 112059. [Google Scholar] [CrossRef] [PubMed]
- Fan, Q.; Huang, G.; Lechno-Yossef, S.; Wolk, C.P.; Kaneko, T.; Tabata, S. Clustered genes required for synthesis and deposition of envelope glycolipids in Anabaena sp. strain PCC 7120. Mol. Microbiol. 2005, 58, 227–243. [Google Scholar] [CrossRef] [PubMed]
- Liaimer, A.; Jenke-Kodama, H.; Ishida, K.; Hinrichs, K.; Stangeland, J.; Hertweck, C.; Dittmann, E. A polyketide interferes with cellular differentiation in the symbiotic cyanobacterium Nostoc punctiforme. Environ. Microbiol. Rep. 2011, 3, 550–558. [Google Scholar] [CrossRef]
- Liaimer, A.; Helfrich, E.J.N.; Hinrichs, K.; Guljamow, A.; Ishida, K.; Hertweck, C.; Dittmann, E. Nostopeptolide plays a governing role during cellular differentitaion of the symbiotic cyanobacterium Nostoc punctiforme. Proc. Natl. Acad. Sci. USA 2015, 112, 1862–1867. [Google Scholar] [CrossRef] [Green Version]
- Moosmann, P.; Ueoka, R.; Gugger, M.; Piel, J. Aranazoles: Extensively chlorinated nonribosomal peptide-polyketide hybrids from the cyanobacterium Fischerella sp. PCC 9339. Org. Lett. 2018, 20, 5238–5241. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, E.W.; Nelson, J.T.; Rasko, D.A.; Sudek, S.; Eisen, J.A.; Haygood, M.G.; Ravel, J. Patellamide A and C biosynthesis by a microcin-like pathway in Prochloron didemni, the cyanobacterial symbiont of Lissoclinum patella. Proc. Natl. Acad. Sci. USA 2005, 102, 7315–7320. [Google Scholar] [CrossRef] [Green Version]
- Rounge, T.B.; Rohrlack, T.; Tooming-Klunderud, A.; Kristensen, T.; Jakobsen, K.S. Comparison of cyanopeptolin genes in Planktothrix, Microcystis, and Anabaena strains: Evidence for independent evolution within each genus. Appl. Environ. Microbiol. 2007, 73, 7322–7330. [Google Scholar] [CrossRef] [Green Version]
- Patterson, G.M.L.; Smith, C.D.; Kimura, L.H.; Britton, B.A.; Carmeli, S. Action of tolytoxin on cell morphology, cytoskeletal organization, and actin polymerization. Cell Motil. Cytoskelet. 1993, 24, 39–48. [Google Scholar] [CrossRef]
- Zhang, X.; Minale, L.; Zampella, A.; Smith, C.D. Microfilament depletion and circumvention of multiple drug resistance by sphinxolides. Cancer Res. 1997, 57, 3751–3758. [Google Scholar]
- Smith, C.D.; Carmeli, S.; Moore, R.E.; Patterson, G.M. Scytophycins, novel microfilament-depolymerizing agents which circumvent P-glycoprotein-mediated multidrug resistance. Cancer Res. 1993, 53, 1343–1347. [Google Scholar]
- Patterson, G.M.; Carmeli, S. Biological effects of tolytoxin (6-hydroxy-7-O-methyl-scytophycin b), a potent bioactive metabolite from cyanobacteria. Arch. Microbiol. 1992, 157, 406–410. [Google Scholar] [CrossRef] [PubMed]
- Dilsizoglu Senol, A.; Pepe, A.; Grudina, C.; Sassoon, N.; Ueoka, R.; Bousset, L.; Melki, R.; Piel, J.; Gugger, M.; Zurzolo, C. Effect of tolytoxin on tunneling nanotube formation and function. Sci. Rep. 2019, 9, 5741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thiery, I.; Nicolas, L.; Rippka, R.; Tandeau de Marsac, N. Selection of cyanobacteria isolated from mosquito breeding sites as a potential food source for mosquito larvae. Appl. Environ. Microbiol. 1991, 57, 1354–1359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briand, E.; Reubrecht, S.; Mondeguer, F.; Sibat, M.; Hess, P.; Amzil, Z.; Bormans, M. Chemically mediated interactions between Microcystis and Planktothrix: Impact on their growth, morphology and metabolic profiles. Environ. Microbiol. 2019, 21, 1552–1566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briand, E.; Bormans, M.; Gugger, M.; Dorrestein, P.C.; Gerwick, W.H. Changes in secondary metabolic profiles of Microcystis aeruginosa strains in response to intraspecific interactions. Environ. Microbiol. 2016, 18, 384–400. [Google Scholar] [CrossRef] [Green Version]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gugger, M.; Boullié, A.; Laurent, T. Cyanotoxins and Other Bioactive Compounds from the Pasteur Cultures of Cyanobacteria (PCC). Toxins 2023, 15, 388. https://doi.org/10.3390/toxins15060388
Gugger M, Boullié A, Laurent T. Cyanotoxins and Other Bioactive Compounds from the Pasteur Cultures of Cyanobacteria (PCC). Toxins. 2023; 15(6):388. https://doi.org/10.3390/toxins15060388
Chicago/Turabian StyleGugger, Muriel, Anne Boullié, and Thierry Laurent. 2023. "Cyanotoxins and Other Bioactive Compounds from the Pasteur Cultures of Cyanobacteria (PCC)" Toxins 15, no. 6: 388. https://doi.org/10.3390/toxins15060388